
Abstract
Background
Circulating MicroRNAs (miRNAs) carried by microvesicles (MVs) have various physiological and pathological functions by post-transcriptional regulation of gene expression being considered markers for many diseases including diabetes and dyslipidemia. We aimed to identify new common miRNAs both in MVs and plasma that could be predictive biomarkers for diabetic dyslipidemia evolution.
Methods
For this purpose, plasma from 63 participants in the study (17 type 2 diabetic patients, 17 patients with type 2 diabetes and dyslipidemia, 14 patients with dyslipidemia alone and 15 clinically healthy persons without diabetes or dyslipidemia) was used for the analysis of circulating cytokines, MVs, miRNAs and MV-associated miRNAs.
Results
The results uncovered three miRNAs, miR-218, miR-132 and miR-143, whose expression was found to be significantly up-regulated in both circulating MVs and plasma from diabetic patients with dyslipidemia. These miRNAs showed significant correlations with important plasma markers, representative of this pathology. Thus, MV/plasma miR-218 was negatively correlated with the levels of erythrocyte MVs, plasma miR-132 was positively connected with MV miR-132 and negatively with uric acid and erythrocyte plasma levels, and plasma miR-143 was negatively related with creatinine levels and diastolic blood pressure. Also, three miRNAs common to MV and plasma, namely miR-21, miR-122, and miR-155, were identified to be down-regulated and up-regulated, respectively, in diabetic dyslipidemia. In addition, MV miR-21 was positively linked with cholesterol plasma levels and plasma miR-21 with TNFα plasma levels, MV miR-122 was negatively correlated with LDL-c levels and plasma miR-122 with creatinine and diastolic blood pressure and positively with MV miR-126 levels, MV miR-155 was positively associated with cholesterol and total MV levels and negatively with HDL-c levels, whereas plasma miR-155 was positively correlated with Il-1β plasma levels and total MV levels and negatively with MV miR-223 levels.
Conclusions
In conclusion, miR-218, miR-132, miR-143, and miR-21, miR-122, miR-155 show potential as biomarkers for diabetic dyslipidemia, but there is a need for more in-depth studies. These findings bring new information regarding the molecular biomarkers specific to diabetic dyslipidemia and could have important implications for the treatment of patients affected by this pathology.
Similar content being viewed by others


Background
One of many reasons for the severity of vascular disease in diabetic patients is dyslipidemia, also known as diabetic dyslipidemia (DDLP) and defined as lipoprotein metabolism disorder characterized by elevated total cholesterol, the “bad” low-density lipoprotein (LDL) cholesterol and the triglyceride concentrations, and a decrease in the “good” high-density lipoprotein (HDL) cholesterol levels in the blood [1]. In diabetes, both small and large vessels are affected by lipid imbalance. In type 2 diabetes (T2D), characterized by impaired insulin sensitivity, the lipid phenotype includes hypertriglyceridemia, reduced plasma HDL cholesterol and the conversion of LDL to a more atherogenic lipoprotein, termed small dense LDL [2]. However, the dominant lipid defect in insulin resistance remains hypertriglyceridemia, a consequence of increased production and decreased release of triglyceride-rich lipoproteins in fasting as well as in non-fasting state [3]. This feature is commonly associated with a reduction in HDL and an increase in small dense LDL levels [4]. Conventionally low HDL levels were regarded as a consequence of insulin resistance and diabetes. Other studies indicated that low HDL may result in or exacerbate abnormal glucose homeostasis [5]. Even though impaired insulin sensitivity is considered to be the main cause for diabetic dyslipidemia, hyperglycemia alone cannot entirely justify the lipid changes, often detected many years before clinical diagnosis of prediabetes in individuals with normal glucose concentrations [6]. Therefore, unlike type 1 diabetes, lipid metabolism dysregulation characteristic for T2D cannot become completely corrected with glycemic control.
In addition to the known specific biochemical profile of diabetic or dyslipidemic patients, some of the most promising biomarkers could be extracellular vesicles (EVs), particularly microparticles (MPs) or microvesicles (MVs), and microRNAs (miRNAs).
MVs are larger EVs, which together with exosomes (smaller EVs) have important roles within our body. All EVs are quite similar, however, their different way of emerging is very important for their function. MVs, are vesicles of about 100 to 1000 nm in diameter, released by most of the activated or apoptotic cells, found both in tissues and body fluids and involved in cell-to-cell communication. Compared to exosomes, MVs do not require exocytosis for their formation and release. They are generated by the outward budding of a plasma membrane domain, as a result of the increase in intracellular Ca2+ concentration leading to phosphatidylserine (PS) exposure. Therefore, their cargo originates from the cytosolic face of the cell, where many proteins and RNAs involved in cell-to-cell interaction can be found [7,8,9,10]. The specific molecular signature of MVs consists of PS as a key identifier, several surface proteins which are specific to paternal cells and RNA, including miRNAs [11]. MVs express numerous features of cells of origin including specific surface antigens and receptors, so they were named according to the cells they come from, for example endothelial, platelet and leukocyte MVs [12]. In vitro studies confirmed that, high glucose concentration activates endothelial cells (ECs) to discharge significant amount of MVs, called endothelial microvesicles (EMVs), with a distinct molecular composition [13, 14]. These endothelial membrane-derived vesicles are also reported to carry regulating molecules crucial in cell-to-cell communication. All types of MVs are endowed with the ability to transport a specific class of miRNAs in circulation and transfer them to certain cells, such as ECs, endothelial progenitor cells (EPCs), through a tightly regulated process [15]. Following, the miRNAs transferred by MVs could play an important role in the regulation of biological processes involved in the vascular complications in diabetes, turning MVs in key vesicles of the processes underlying intercellular communication in different pathologies including diabetic dyslipidemia [16]. The miRNAs are known as small non-coding RNAs which regulate gene expression post-transcriptionally through the inhibition of translation and/or the degradation of messenger RNA (mRNA) and they have been found in several body fluids, including blood, circulating EVs or associated with lipoproteins or protein complexes [17, 18]. The levels of miRNAs are modified during disease progression and miRNAs’ circulating profiles often reflect their modified expression in the tissue of origin or indicate increased intercellular communication [19]. There are clear data on a possible use of miRNAs as diagnostic biomarkers and therapeutic targets for the endothelial dysfunction in hypertensive patients [20]. In diabetes, alterations of miRNA expressions were assumed that could disturb the production of insulin and occurrence of diabetic complications [21]. In addition, differences in the abundance of pro- and anti-angiogenic miRNAs in the MV-enriched plasma fraction were reported in patients with T2D, suggesting that development of vascular complications is due to impaired angiogenesis in such patients [22]. Reportedly, miRNAs associated with MVs regulate cellular mechanisms involved in vascular homeostasis in the progression of diabetic dyslipidemia [7, 23,24,25]. The most recent methods of diagnosis and therapy target EV and miRNA technologies. We hypothesized that detection of the changes in plasma MV components could be used as early biomarkers to prevent the irreversible effects of diabetic dyslipidemia. The aim of this study was to search whether the newly discovered modifications of the circulating MVs and plasma miRNAs could be predictive for diabetic dyslipidemia evolution. However, to date, MV-associated and circulating miRNAs are not applied as biomarkers in clinical settings for diabetic dyslipidemia.
Methods
Patients and control subjects
A total number of 63 male and female participants, including healthy volunteers, patients with dyslipidemia and patients with type 2 diabetes, selected from National Institute for Diabetes, Nutrition and Metabolic Disease ‘Prof. Dr. Nicolae Constantin Paulescu’, Bucharest, Romania, and who signed the consent form, were enrolled in the study and divided into 4 experimental groups.
Diabetic patients of similar age and body mass index (BMI) and different degrees of cholesterol levels were divided into 2 experimental groups: (1) patients with type 2 diabetes (T2D group, n = 17) and (2) patients with type 2 diabetes and dyslipidemia (T2D-DLP group, n = 17). As control, two experimental groups of participants were also included: (3) patients with dyslipidemia alone (DLP group, n = 14, control for T2D-DLP group) and (4) clinically healthy persons without diabetes or dyslipidemia (C group, n = 15, control for DLP, T2D and T2D-DLP groups).
The division of patients into diabetics and non-diabetics was made according to American Diabetes Association (ADA) criteria by measuring fasting plasma glucose (FPG), a test done first thing in the morning, before breakfast, after a minim of 8 h fasting (means not eating and drinking anything, except water, for a period of 8 h). Diabetes was diagnosed at fasting blood glucose of greater than or equal to 126 mg/dL, and normal was considered at fasting blood glucose under 100 mg/dL.
Clinical and metabolic measurements
The blood was collected by venipuncture from each volunteer or patient included in the study and clinical characteristics (gender, age, BMI, systolic/diastolic blood pressure, heart rate and several biochemical data: transaminases, urea, creatinine, uric acid, hemoglobin and leucocyte) were carried out. Also, to state the metabolic profile, consisting of plasma glucose, glycated hemoglobin (HbA1C), total cholesterol, LDL-cholesterol, HDL-cholesterol, triglyceride, was determined. All tests were performed on sera using Cobas Integra 400 plus analyzer (Roche Diagnostics). Additional blood samples were collected from every participant in the study and stored at − 70 °C for further investigations.
Enzyme-linked immunosorbent assay
Using specific Elisa kits (R&D Systems), the circulating levels of inflammatory factors, interleukin 6 (IL-6), interleukin 1beta (IL-1β), interferon gamma (IFNγ), tumor necrosis factor alfa (TNFα), were measured in sera as indicated by the manufacturer. Elisa plates were analyzed on spectrophotometer (Tecan Infinite M200 PRO).
Microvesicle isolation
Peripheral blood have been collected on anticoagulant (2 vacutainer tubes, blood collection tubes with EDTA K3) and total circulating MVs were separated from plasma by sequential centrifugations, according to the protocol optimized after the one described by [9], as follows: the peripheral blood was centrifuged at 2500g for 10 min at 4 ºC to collect the platelet poor plasma (PPP) from supernatant; subsequently, 200 µL PPP were centrifuged at 16000g for 5 min at 4 ºC to remove residual platelets, apoptotic bodies and collect the platelet free plasma (PFP) in supernatant; finally, MVs obtained in the pellet by centrifugation of PFP at 20000g for 90 min at 4 ºC were washed twice (20000 g, 90 min, 4 ºC) with phosphate buffered saline (PBS), re-suspended in PBS (~ 100 μL) and stored at − 80 °C until further analysis [19].
Microvesicle characterization
Zetasizer Nano ZS (Malvern Instruments, Malvern, United Kingdom) equipped with a solid-state He–Ne laser at 633 nm wavelength, was used to establish the size distribution for MVs. The measurements were carried out in triplicates on MV suspension (10 μL) diluted to 1 mL of PBS. Zetasizer software version 7.03 was used in order to process and analyze the obtained data.
The fluorescence and electron microscopy were applied to investigate the structure and size of MVs. For fluorescence microscopy analysis, MVs were stained with red fluorescent membrane linker dye PKH26 (Invitrogen, Waltham, MA, United States) according to the manufacturer’s instructions and the protocol described by [26]. In brief, MV suspension (20 μL) in PBS, mixed with diluent C (1 mL) and PKH26 (1 mL) stock solution (4 × 10−6 M/L) was incubated at 37 °C for 5 min, the reaction was stopped by adding an equal volume of fetal bovine serum (FBS). MVs were analyzed under a Zeiss Axiovert microscope (Carl Zeiss, Jena, Germany). For electron microscopy analysis, MVs were subjected to the negative staining technique consisting of incubating 5 μL of MVs in suspension for 2 min on carbon-coated copper grids (100 mesh, Agar Scientific) and their subsequent staining with 2% uranyl acetate at room temperature (RT). Image acquisition was carried out at 18˚C using a Talos F200C Transmission Electron Microscope (Fei/Thermo Fisher Scientific Talos F200C TEM) [27].
Microvesicle quantification and analysis
A Gallios Flow Cytometer (Beckman Coulter Life Sciences, CA, USA) was used to quantify the levels of MVs in circulation and analyze them according to the cells of origin from which they come.
To quantify MVs, 10 µL of MVs re-suspended in PBS were mixed with 10 µL of counting beads (1000 beads/µL, 10 µm diameter), incubated for 40 min at RT in the dark, and 100 µL of PBS were added at the time of reading to the flow cytometer for 60 s. The number of MVs (0.1–1 µm diameter) was calculated using the formula: MVs as events/µL = [(MV count/bead count) × bead concentration/µL] × MV purity/100, where: MV count and bead count were collected from the dot-plot representations (X = forward-scatter intensity, Y = side-scatter intensity), and MV purity was evaluated as follows: 10 µL of re-suspended MVs were incubated with 2.5 µL of Annexin V-FITC antibody (binds PS on MVs) in the presence of 2 mM CaCl2 for 40 min at RT in the dark, diluted with 100µL of PBS and read for 60 s at flow cytometer [15, 19].
To analyze MVs according to the types of cells from which they originate in the circulation, fluorescent antibodies (all from SantaCruz Biotechnology, Inc.) against specific cell surface markers were used: CD41 (Integrin alpha-IIb)—specific marker for platelets; CD144 (VE-cadherin)—specific marker for endothelial cells; CD14—specific marker for monocytes; CD18 (Integrin beta chain-2)-specific marker for leucocytes; CD235a (glycophorin A)-specific marker for erythrocytes. In brief, 10 µL of re-suspended MVs were incubated with 2.5 µL of Annexin V-FITC antibody and specific surface marker fluorescent antibody for 40 min at RT in the dark, diluted with 100 µL of PBS and analyzed with a 5000-event cutoff. Data were analyzed with Kaluza Flow Cytometry Analysis Software v2.1 (Beckman Coulter Life Sciences, Indianapolis, IN, USA) and expressed as a percentage of MVs double labeled with Annexin V and antibody specific to the cell of origin. The isotype control experiments were performed as well.
RNA isolation and quantification
Total RNAs, including miRNA, were isolated from 200 µL plasma or purified MVs (obtained from 400 µL plasma and re-suspended in 100 µL PBS) using miRNeasy Serum/Plasma Kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. After, RNAs were eluted in 14 μL RNase-free water. The RNA purity and concentration were evaluated by spectrophotometry using NanoDrop 2000c (Thermo Fisher) and isolated RNAs were kept at − 80 °C until further analysis.
Analysis of miRNA expression in plasma and circulating MVs
TaqMan technology was used to measure the levels of hsa-miR-21-5p (ID: 000397), hsa-miR-34a-5p (ID: 000426), hsa-miR-122-5p (ID: 002245), hsa-miR-126-3p (ID: 002228), hsa-miR-132-3p (ID: 000457), hsa-miR-143-3p (ID: 002249), hsa-miR-155-5p (ID: 002623), hsa-miR-200b-3p (ID: 002251), hsa-miR-212-5p (ID: 000515), hsa-miR-218-5p (ID: 000521) and hsa-miR-223-3p (ID: 002295) (all from Qiagen, Hilden, Germany) in plasma and circulating MVs.
For this purpose, 50 ng RNA were consumed to generate single stranded cDNA using specific stem-loop primers of TaqMan miRNA Assays (RT primer-for transcription) and TaqMan MicroRNA Reverse Transcription Kit. cDNA was amplified with TaqMan Gene expression Master Mix and specific stem-loop primers of TaqMan miRNA Assays (TM primer-for amplification) to profile mature miRNAs by quantitative reverse transcription polymerase chain reaction (qRT-PCR). The investigated miRNA-specific RT/TM primers were used in separate reactions performed in duplicate. For the quantification of miRNAs in plasma it was used synthetic cel-miR-39-5p (ID 000200) (Qiagen, Hilden, Germany) as an exogenous control. This was added as spike-in during RNA isolation from plasma as previously described [28, 29]. Regarding miRNA quantification in circulating MVs, U6 small nuclear RNA (U6 snRNA) (ID001973) (Qiagen, Hilden, Germany) was used as an endogenous control and it was added in separate reactions in the same way as all the other miRNA-specific RT/TM primers were added.
All reactions were performed using Veriti real-time PCR system (for Reverse Transcription Assay) and VIIA7 real-time PCR system (for qRT-PCR Assay), both from Applied Biosystems (Life Technologies Foster City, CA, USA). The data were analyzed using VIIA7 Software v1.2 (Applied Biosystems) with the automatic quantification cycle (Cq) settings. Raw data passed all quality control checks and were filtered by applying cut-offs of Cq < 35 (measure of a good amplification).
MiRNA levels were calculated using the 2−ΔCq method and multiplied by 103 (for plasma). The expression level of each miRNA of interest was normalized to exogenous cel-miR-39-5p for plasma or endogenous snRNU6 for MVs, as previously re-ported [9, 19].
Statistical analysis
Statistical analysis was performed using GraphPad Prism 7.02 software (GraphPad Software Inc., San Diego, CA, USA). The results were expressed as the means ± standard deviation (SD). Differences between experimental groups were determined by one-way ANOVA followed by Bonferroni multiple comparison tests. Statistical significance was set at p < 0.05 (&, *, #), p < 0.01 (&&, **, ##) and p < 0.001(&&&, ***, ###). Spearman rank correlation test was used to assess relationships among continuous variables. A p value less than 0.05 (typically ≤ 0.05) was considered statistically significant.
Results and discussion
The metabolic and inflammatory profile of diabetic patients with dyslipidemia
Demographic and clinical characteristics of the patients
The patients included in the study were divided according to their clinical characteristics into the following groups: (1) patients with type 2 diabetes (T2D group); (2) patients with type 2 diabetes and dyslipidemia (T2D-DLP group); (3) patients with dyslipidemia alone (DLP group), and (4) clinically healthy persons without diabetes or dyslipidemia (C group).
As shown in Table 1, both T2D patients and T2D-DLP patients had poor glycemic control as revealed by significant higher fasting plasma glucose and HbA1c values compared with healthy individuals (C group). Also, patients with dyslipidemia (DLP group) and patients with diabetic dyslipidemia (T2D-DLP group) exhibited significantly higher levels of triglycerides compared with control group. The rest of the metabolic parameters did not significantly differ among the 4 experimental groups. Both systolic and diastolic blood pressure were significantly higher in DLP patients, while T2D patients had only systolic blood pressure significantly higher, compared to healthy subjects. Diabetic patients with dyslipidemia (T2D-DLP group) did not show significant changes in systolic and diastolic blood pressure values (Table 1).
Plasma inflammatory markers of the patients
Elevated values of pro-inflammatory molecules correlated or not with reduction of anti-inflammatory ones, were usually associated with dyslipidemia as well as with diabetes. Also, combined elevation of inflammatory cytokines, led not only to a stronger inflammatory environment, but it could even be essential for the appearance of the diabetic condition. Interestingly, it was shown that patients with elevated levels of IL-6, IL-1β and TNFα had considerably increased risk of developing T2D compared with individuals with higher levels of IL-6 alone. Pro-inflammatory cytokine, IL-1β, has been described as a severe inducer of beta-cell damage. It has been shown that, long lasting elevated IL-1β levels in both obese and T2D patients cause beta-cell dysfunction [30, 31], while low levels of IL-1β lead to β-cell proliferation, beta-cell apoptosis inhibition and enhance glucose-stimulated insulin secretion [32]. However, in vitro studies showed that IL-1β is essential not only for beta-cell compensation, but also for β-cell failure as well [32, 33], meaning that the IL-1β levels may vary in diabetes, depending on the disease stage. Also, age, BMI and various genetic and hormonal factors, can modify the levels of IL-1β in T2D [30, 34], and the treatment with metformin or acarbose decreases IL-1β protein levels in T2D patients [30, 35]. The levels of inflammatory cytokines, IL-6, IL-1β, IFNγ, TNFα, were measured in plasma of the individuals evaluated in our study (Fig. 1).

Pro-inflammatory levels of IL-6 (A), IL-1β (B), IFNγ (C) and TNF-α (D) in plasma from patient groups DLP(n = 14), T2D(n = 17), T2D-DLP(n = 17) and group C (n = 15) of healthy subjects, measured by Elisa method. Data are expressed by means ± SD. &p < 0.05 (vs C), &&p < 0.01 (vs C), *p < 0.05 (vs DLP), #p < 0.05 (vs T2D) (One-Way ANOVA with Bonferroni's Multiple Comparison Test)
As shown in Figure, T2D patients had significantly elevated plasma levels of IL-6, while patients with dyslipidemia alone (DLP group) had significantly increased levels of both IL-6 and IL-1β, when these were compared with the levels measured in healthy individuals (C group). These results are in agreement with those reported by [36] that showed that an imbalance between IL-6 and IL-10 expression led to increased risk of developing T2D, but at the same time they are in disagreement with the results that showed that increased levels of IL-6 associated with those of IL-1β represent an increased risk for developing T2D [37]. In our study, T2D was associated only with an increase in plasma concentrations of IL-6 (Fig. 1A), not with increases in IL-1β, IFNγ and TNFα plasma levels (Fig. 1B–D). In addition, diabetic patients with dyslipidemia (T2D-DLP group) showed significant increases in plasma concentrations of IL-6 both compared to group C and compared to groups T2D and DLP, meaning that both diabetes and dyslipidemia had a cumulative effect on IL-6 plasma levels. (Fig. 1A). It should be noted that, the plasma concentrations of the other investigated inflammatory markers (IL-1β, IFNγ, TNFα) remained unchanged in diabetic dyslipidemia (T2D-DLP group), although IL-1β increased significantly in the patients who had only dyslipidemia (DLP group) (Fig. 1B–D). In addition, a positive correlation was observed between IL-1β levels and systolic blood pressure (r = 0.90; p = 0.03) in patients with diabetic dyslipidemia (Table 2).
Elevated levels of circulating microvesicles in diabetic patients with dyslipidemia
Characterization, quantification and analysis of plasma microvesicles
MVs were isolated through successive centrifugations from plasma of patients and healthy individuals and analyzed by Zetasizer Nano ZS to establish their size. The distribution curve of MVs was unimodal and the value of particle dimension (Z) was between 100 and 1000 nm (Fig. 2A), specific to MVs. In addition, the structure and size of MVs isolated from plasma of patients and healthy subjects, were investigated using both fluorescence (Fig. 2B) and transmission electron microscopy (Fig. 2C), to confirm their size and also to characterize their structure. Image analysis showed that, the majority population of vesicles was found in a range of values specific to MVs (100–1000 nm), and these MV fractions contain heterogeneous particles of larger dimensions.

Analysis of plasma MVs by Zetasizer Nano ZS showing a large population represented by one peak at size range of ~ 100–1000 nm, specific to MVs (Y-axis represents the number of MVs and X-axis the size of MVs) (A). Examination of plasma MVs by fluorescence microscopy using PKH26 staining technique (B), and electron microscopy using negative staining technique (C)
Flow cytometry was used to characterize and quantify the total MVs isolated from plasma of individuals taken into the study, and also to analyze the levels of specific cell derived MVs (Figs. 3, 4). MVs were characterized and quantified by calibration with beads of known size and concentration (1000 beads/µL, 10 µm diameter) (Fig. 3A). In addition, staining of specific marker for MVs, phosphatidylserine (PS), with AnnexinV-FITC, was used to characterize and highlight the purity of MVs isolated from both patient and healthy subject plasma (Fig. 3B).

Characterization and quantification of circulating MVs by flow cytometery in plasma from patient groups DLP (n = 14), T2D (n = 17), T2D-DLP (n = 17) and group C (n = 15) of healthy subjects. Representative dot-plots assigned for MV quantification, based on calibration beads of 10 μm in diameter and 1000/µL concentration (A), and MV purity based on the percentages of MVs positive for Annexin V (B). Annexin-V-positive plasma MV concentrations for all groups of subjects investigated (C). Data are represented as means ± SD. &p < 0.05, &&p < 0.01 (vs C) (One-Way ANOVA with Bonferroni’s Multiple Comparison Test)

The plasma concentrations of MVs from patient groups DLP (n = 14), T2D (n = 17), T2D-DLP (n = 17) and group C (n = 15) of healthy subjects, originating from different cell types: MVs derived from endothelial cells, EMVs: AnnexinV+/CD144+ (A), platelets, PMVs: AnnexinV+/CD41+ (B), leucocytes, LeMVs: AnnexinV+/CD18+ (C), monocytes, MMVs: AnnexinV+/CD14+ (D), and erythrocytes, ErMVs: AnnexinV+/CD235a+ (E). Data are expressed as means ± SD. &p < 0.05, &&p < 0.01, &&&p < 0.001, (vs C), *p < 0.05, **p < 0.01, ***p < 0.001 (vs DLP), #p < 0.05, ##p < 0.01, ###p < 0.001 (vs T2D) (One-Way ANOVA with Bonferroni’s Multiple Comparison Test)
In patients with diabetic dyslipidemia, as well as in patients with diabetes or dyslipidemia alone, the levels of circulating MVs were significantly higher compared to healthy subjects (Fig. 3C). All these data are sustained by previous reports regarding altered levels of MVs in many diseases including hypercholesterolemia [12], diabetes [38, 39], atherosclerosis [10], cancer [11], which suggest that circulating MVs could be used as biomarkers for the onset and progression of certain disease. It should be noted that our results did not highlight differences in the level of circulating MVs between the groups of patients investigated (Fig. 3C).
Further, the levels of plasma MVs derived from endothelial cells (EMVs: CD144+, AnnexinV+), platelets (PMVs: CD41+, AnnexinV+), leukocytes (LeMVs: CD18+, AnnexinV+), monocytes (MMVs: CD14+, AnnexinV+), and erythrocytes (ErMVs: CD235a+, AnnexinV+) were analyzed by flow cytometry for DLP, T2D, T2D-DLP patient groups and control group. The concentrations of plasma EMVs, PMVs, LeMVs, MMVs and ErMVs were calculated, reporting their percentages for double labeling, to the percentage of 100% of the total plasma MVs (AnnexinV+), for which the concentration was previously calculated (Fig. 3) according to the formula presented in materials and methods (Fig. 4).
As shown in Fig. 4, in the case of diabetic patients with dyslipidemia (T2D-DLP group) were found significantly increased concentrations of plasma EMVs, PMVs and LeMVs compared to healthy subjects (C group) or diabetic patients (T2D group) (Fig. 4A–C), and of plasma ErMVs compared to healthy subjects and dyslipidemic patients (DLP group) (Fig. 4E). Diabetic dyslipidemia, however, did not induce changes in plasma concentrations of MMVs compared to any of the investigated groups (Fig. 4D). It should be noted that, except for MMVs, all other types of plasma MVs investigated, respectively EMVs, PMVs, LeMVs and ErMVs, presented significantly increased levels in patients with diabetic dyslipidemia compared to healthy individuals (Fig. 4). There are no studies that highlight the effect of diabetic dyslipidemia on circulating MV levels or on certain cell types of MVs. Our result could be valuable information in finding new biomarkers for monitoring patients with diabetic dyslipidemia.
Significant changes were also observed when plasma MV levels from diabetic patients (T2D group) or patients with dyslipidemia alone (DLP group) were compared to those from healthy subjects (C group). Dyslipidemia was associated with significant increases in plasma concentrations of EMVs, PMVs, LeMVs and MMVs (Fig. 4A–D), while diabetes only with significant increases in plasma concentrations of ErMVs (Fig. 4E). Our results suggest that dyslipidemia and less diabetes generate significant changes in the plasma concentrations of MVs.
Consistent with growing data suggesting that platelet-derived MVs (PMVs) represent over 50% of all circulating MVs, we found that in both diabetic dyslipidemia and dyslipidemia, PMVs are the most abundant in the circulation. Increased PMV levels were found in numerous diseases such as diabetes mellitus, cardiovascular diseases, coagulation disorders, rheumatoid arthritis, cancers, infections and atherosclerosis [29, 40,41,42], as main regulators of inflammation, hemostasis and angiogenesis [29, 41, 42]. Patients with hypercholesterolemia displayed elevated levels of PMVs and MMVs compared to healthy subjects Moreover, in familial hypercholesterolemia, increased concentrations of EMVs, PMVs, LeMVs, MMVs and ErMVs have been found [43].
Although in our study we found that type 2 diabetes induces changes only in the level of plasma concentrations of total MVs and ErMVs, the other types of vesicles investigated having normal values, there are still studies that showed that AnnexinV-positive blood cell microvesicles (total MVs) and particularly EMVs, PMVs and MMVs had higher levels in patients with either type 1 or type 2 diabetes [44] compared with healthy individuals. According to our results, [45] Freeman et al. found that ErMV levels were significantly higher in individuals with diabetes compared with controls [45], while Giannella et al. [13] showed that LeMV levels were not significantly different between T2D patients and controls [46] or pre-diabetic (PreDM) patients. Also, PreDM and T2D patients showed significantly higher levels of circulating activated endothelial cell-derived MVs (AnnexinV+/CD62E+) compared to normal glucose tolerance (NGT) subjects, with the specification that the highest levels were recorded in the case of T2D patients [13, 44]. Moreover, in vitro it was demonstrated that endothelial cells exposed to high glucose can alter EMV content and also induce modifications of cellular function [47]. Also, more data confirmed that the levels of circulating PMVs were increased in diabetic patients compared with controls [48,49,50,51], but after treatment with anti-platelet drugs cilostazol or sarpogrelate hydrochloride their number was significantly decreased [49,50,51]. It has been showed that drugs used in prevention of cardiovascular disease also affect the number and composition of PMVs [51,52,53]. Moreover, patients treated with statins registered lower numbers of extracellular vesicles, especially of those derived from platelets, leukocyte and endothelial cells, compared to untreated patients [53, 54]. However, all these data are inconsistent, since extracellular vesicle concentration often depends on the circumstance of their release, trigger, molecular cargo, or cells of origin [53].
Finally, Hototen and Evans concluded that the levels of circulating MVs, in particular those derived from monocytes, platelets, and endothelial cells, could be used as a biomarker for T2D patients, but considering the treatment plan followed and the onset of disease for each patient. Regarding diabetic dyslipidemia, we can assume that MV concentration might be dependent by the presence of either dyslipidemia (EMVs, PMVs and LeMVs) or diabetes (ErMVs) for each type of MV we investigated.
Furthermore, significant Pearson correlations between circulating MVs (EMVs, PMVs, LeMVs, MMVs, ErMVs) and metabolic parameters or plasma inflammatory cytokines in plasma from diabetic patients with dyslipidemia investigated in our study, are shown in Tables 3 and 4. Positive correlations between PMVs and HbA1c (r = 0.90, p = 0.01), MMVs and ALT (r = 0.96, p = 0.01) or leucocytes (r = 0.91, p = 0.03), and between total MVs and TG (r = 0.92, p = 0.02) or urea (r = 0.86, p = 0.05) can be noticed (Table 3). In addition, negative correlations between MMVs and monocytes (r = 0.94, p = 0.01) and between total MVs and HDL-c (r = 0.91, p = 0.03) can be observed (Table 3). A positive correlation was also observed between MMVs and inflammatory factor IFNγ (r = 0.86, p = 0.05) in diabetic patients with dyslipidemia (Table 4). Until now, there are no studies that highlight the possible correlations that appear in diabetic dyslipidemia between plasma MVs and metabolic parameters or plasma inflammatory markers. There are studies that emphasize the presence of certain correlations only in diabetes or dyslipidemia. Thus, in diabetic patients, PMVs and EMVs were inversely related to HbA1c [53, 55]. Also, Gianella et al. [13] found a negative correlation between CD62E+MVs and plasma glucose of PreDM and NGT individuals. In familial hypercholesterolemia, the levels of circulating MMVs were directly correlated with oxLDL plasma concentrations [54].
Microvesicle-associated and circulating microRNAs in diabetic dyslipidemia
MicroRNA expression profile in plasma microvesicles
Original studies, indicated that extracellular vesicles are important mediators of intercellular communication due to their RNA content, particularly miRNAs, which regulates transcription and translation of several genes [22, 56, 57]. Recently, in a study focused on the relationships between miRNA polymorphisms, insulin signaling pathways and T2D, it was reported that miR-133a-2 rs13040413, let-7a-1 rs13293512 and miR-27a rs895819 may be associated with T2D susceptibility [58]. Common genetic and epigenetic aspects in T2D and cardiovascular disease, including microRNAs and long non-coding RNAs, whose aberrant regulation has been implicated in both disease states, either etiologically or as a cause of their progression, have been discussed by [59]. Furthermore, a growing number of studies has shown that a specific set of miRNAs has an altered expression profile in both the progression of diabetes and dyslipidemia, making these biomolecules potential biomarkers for disease prognosis, diagnosis, and management [21, 22]. They have been reported as biomarkers and due to the beneficial effects that certain drugs such as empagliflozin [60] or physical exercises [61] have on them in patients with heart failure with preserved ejection fraction and diabetes or patients with diabetic heart. Knowing also that, modulation of miRNA expression affects insulin production and the incidence of vascular complications in diabetes [21, 22], in our study we analyzed the expression of 11 miRNAs, from those previously reported to undergo changes during diabetes and dyslipidemia evolution. Selected miRNAs, miR-21-5p, -34a-5p, -122-5p, -126-3p, -132-3p, -143-3p, -155-5p, -200b-3p, -212-5p, -218-5p and -223-3p, were first investigated in MVs isolated from plasma of patients with diabetic dyslipidemia, with diabetes or dyslipidemia alone and of healthy subjects (Figs. 5 and 6). However, a systematic investigation of microvesicle-associated microRNAs (MV miRNAs) in diabetic dyslipidemic patients has not yet been reported to our knowledge.

Relative expression (2−ΔCq, ΔCq = Cq has-miRNA—Cq U6) of miRNAs in MVs from the plasma of patients with dyslipidemia (n = 14), diabetes (n = 17), diabetes associated with dyslipidemia (n = 17) and of healthy subjects taken as control (n = 15): A miR-218-5p; B miR-212-5p; C miR-200b-3p; D miR-143-3p; E miR-132-3p; F miR-34a-5p. Data are expressed by means ± SD. &p < 0.05 (vs C), *p < 0.05 (vs DLP), #p < 0.05 (vs T2D), &&/**/##p < 0.01 (One-Way ANOVA with Bonferroni's Multiple Comparison Test)

Relative expression (2−ΔCq, ΔCq = Cq has-miRNA—Cq U6) of miRNAs in MVs from the plasma of patients with dyslipidemia (n = 14), diabetes (n = 17), diabetes associated with dyslipidemia (n = 17) and of healthy subjects taken as control (n = 15): A miR-21-5p; B miR-122-5p; C miR 126-3p; D miR-155-5p; E miR-223-3p. Data are expressed as means ± SD. &p < 0.05, &&p < 0.01, &&&p < 0.001 (vs C), #p < 0.05 (vs T2D) (One-Way ANOVA with Bonferroni's Multiple Comparison Test)
In diabetic patients with dyslipidemia (T2D-DLP group), we found significantly elevated levels of miR-218, miR-212, miR-200b and miR-143 contained in circulating MVs, compared to all the other experimental groups (Fig. 5:A–D). Also, in diabetic dyslipidemia, an up-regulation of miR-132 and miR-122, contained in circulating MVs, was observed when compared to T2D group (Figs. 5E, 6B). It should be noted that, the miR-132 also registered strong negative correlations with plasma levels of cholesterol (r = 0.86, p = 0.05), uric acid (r = 0.89, p = 0.01) and erythrocytes (r = 0.95, p = 0.01) (Table 5). In the case of miR-34a, a slightly increasing trend was observed for investigated patients compared to healthy subjects, but not statistically significant (Fig. 5F).
The other investigated MV miRNAs, respectively miR-21, miR-122, miR-126, miR-155, had the expression levels significantly reduced for all studied patient groups compared to the control group (Fig. 6A, C, D), while down-regulation of miR-223 was not significant for dyslipidemic and diabetic dyslipidemic groups (Fig. 6E). Importantly, these latter MV miRNAs, except for miR-122, did not have altered gene expression levels between the groups of patients (Fig. 6A, C, D, E).
The possible correlations between miRNAs carried by plasma MVs and clinical (Table 5) or inflammatory (Table 6) parameters in diabetic dyslipidemia were established. Anywise, MV miR-21 and MV miR-155 displayed positive correlations with plasma levels of cholesterol (r = 0.90, p = 0.03, respectively r = 0.88, p = 0.04), MV miR-126 with HbA1c levels (r = 0.91, p = 0,02) and SBP (r = 0.87, p = 0.05) (Table 5), MV miR-155 presented negative associations with plasma levels of HDL-c (r = 0.90, p = 0.03) and MV miR-122 only with LDL-c levels (r = 0.99, p = 0.001) (Table 5). In addition, negative correlations were recorded for MV miR-223 gene expression with IL-1β plasma levels (r = 0.92, p = 0.02) and MV miR-212 with IFNγ levels (r = 0.92, p = 0.02) (Table 6).
Importantly, we tried to find possible correlations between MV miRNA expressions, levels of different types of MVs in plasma and total circulating MVs levels. The analysis revealed the presence of some positive correlations between MV miR-126 and the levels of PMVs (r = 0.86, p = 0.05), MV miR-155 and total levels of MVs (r = 0.97, p = 0.01), and also of some negative correlations between MV miR-218 and the levels of ErMVs (r = 0.94, p = 0.01) (Table 7).
Many studies have been performed on exosomes carrying several types of miRNAs and only a few attested the role of MVs in caring regulator miRNAs in diabetes or dyslipidemia.
So far, little is known about miR-218, miR-143 and miR-34a contained in EVs in diabetes. Higher amounts of miR-218 were found in urinary exosomes from T1D children, compared to healthy controls [62]. Exosomal miR-34a has been found increased in T1D and dyslipidemic patients being associated with alterations of lipid metabolism (increased levels of cholesterol and triglycerides) and endothelial function [63]. We have previously showed that MVs released from platelets isolated from hypertensive-hyperlipidemic hamsters, contain significantly increased levels of miR-34a and reduced of miR-143 [29], platelets being considered the main source of extracellular vesicles in plasma [22, 64]. In contradiction, we found here that MVs obtained from patients with diabetes or dyslipidemia alone, carry, just like MVs from healthy subjects, decreased levels of miR-218, miR-143 and miR-34a, but MV miR-218 and MV miR-143 levels were significantly augmented in diabetic patients with dyslipidemia (Fig. 5A, D, F).
EVs derived from beta cells also hold the miRNA profile of the host cells, including miR-212 and miR-132 [65, 66]. In mature pancreatic beta cells miR-212 and miR-132 are key players in insulin secretion [65, 66]. It is believed that miR-132 might have a protecting role against diabetes development by promoting cell growth and reducing cell death in beta cells [67], down-regulated miR-132 being associated with a minor inhibition of cell proliferation [68]. However, contradictory data attested up-regulation of miR-132 expression in several T2D models [68]. Furthermore, mir-132 has been found increased in both EVs in early gestation and placenta exosomes in gestational diabetes patients [69], while in plasma from second-trimester gestational diabetes patients was insignificantly [70]. Two other studies performed on a large group of patients found miR-132 decreased in gestational diabetes pregnancies [71, 72]. In line with these results, we also found a slightly but not significant decrease of miR-132 levels in MVs isolated from T2D patients compared to healthy individuals (Figs. 4, 5E). Our results also highlighted significantly increased levels of miR-132 in MVs from patients with dyslipidemia, compared to control subjects (Fig. 5E), being in agreement with a study that showed that in metabolic syndrome EVs derived from porcine adipose derived stem cells have been found to contain high levels of miR-132 [73]. In our study the levels of miR-212 have been found significantly increased only in MVs from patients with T2D alone, compared to control subjects, while in the group of the patients with dyslipidemia MV miR-212 was not significantly modified (Fig. 5B).
In conclusion, from our results we can infer that only the association of dyslipidemia with diabetes could significantly increase the gene expression levels of miR-218, miR-212, miR-200b, miR-143, and miR-132 contained in plasma MVs (Fig. 5).
Regarding the MV miRNAs that we found to be down-regulated in diabetic dyslipidemia, there are some studies that confirm our results, while others deny them. Thus, circulating EV miR-21 was considered to be a possible biomarker for T1D progression, both in vitro and in vivo studies reporting increased levels of miR-21 transported by EVs. Therefore, up-regulated miR-21 was found in MIN6 beta cell-line, after treatment with inflammatory cytokines and even before hyperglycemia establishment, in NOD mice. In preclinical studies using hamsters with hyperlipidemia and hypertension, expression of miR-21 was found up-regulated in MVs derived from platelets [29]. Moreover, the increased presence of miR-21 has been reported in children with new-onset T1D [74]. Contrary to these results, in our study we found significantly decreased levels of circulating miR-21 carried by MVs in diabetic, dyslipidemic and diabetic dyslipidemic patients (Fig. 6A).
In T1D patients [75] and even in the new-onset T1D cohort [76], another highly overexpressed EV miRNA was miR-122. In the case of exosomes from obese mice, miR-122 was found increased, while these exosomes led to insulin resistance and glucose intolerance when injected in lean mice [77]. Significantly higher levels of miR-122 have been found in placenta of gestational diabetes patients [78]. In contrary, we found down-regulated expression of miR-122 transported by circulating MVs, in T2D compared to control group (Fig. 6B). Even thou medications alter some miRNA cargo in EVs, it has been demonstrated in a very recent study that levels of EV-encapsulated miR-122 were not affected by metformin treatment. Consistent with our results, in metformin-insensitive T2D patients, Peng and collaborators found low levels of exosome miRNA-122, indicating miR-122 as biomarker of metformin insensitivity [79]. As important regulator of glucose and lipid metabolism as well [80,81,82], increased miR-122 showed a specific correlation with monounsaturated and saturated fatty acids within the lipid classes triacylglycerols and cholesterol esters. Also, one year atorvastatin treatment led to significant reduction in both cholesterol and serum miR-122 levels [83]. Contrary to these reports, we found MV miR-122 levels decreased in dyslipidemia alone or diabetic dyslipidemia when compared to healthy individuals (Fig. 6B).
Two others important MV miRNAs investigated by us, which were found in abundance both in platelets and in MVs derived from them [22, 40, 84,85,86,87], were miR-126 and miR-223. In addition to platelets, endothelial cells have been found to express miR-126 and, moreover, it has been shown that the highest amount of circulating miR-126 is found in MVs derived from endothelial cells [22, 84].
Previous studies revealed decreased expression of MV miR-126 in patients with diabetes and prevalent cardiovascular disease [22, 88] and miR-223 within MVs released by platelets from hypertensive-hyperlipidemic hamsters [29]. Our data confirm findings attesting miR-126 as one of the most down-regulated MV miRNA, especially in endothelial cells—derived MVs and in the whole plasma [89,90,91]. Specifically, we found that, the gene expression for MV miR-126 was significantly reduced for all groups of patients included in the study (Fig. 6C), while for MV miR-223 it was significantly reduced only for the group of diabetic patients (Fig. 6E).
Another miRNA found significantly decreased in MVs isolated from patients with diabetic dyslipidemia, with diabetes or dyslipidemia alone, was miR-155 (Fig. 6D). In addition to our results, other clinical studies measuring miR-155 in EVs derived from different sources, pointed out that patients with T1D exhibited lower amounts of miR-155, also correlated with the severity of the diseases. Decreased levels of miR-155 in EVs isolated from blood were also found at patients with diabetic retinopathy compared to diabetic patients with no eye defects or to healthy individuals [92, 93]. Also, urinary exosomes from diabetic patients carry lower amount of miR-155, decreased miR-155 being associated with micro albuminuria in T1D patients [92, 94].
To conclude, we believe that cumulative effect of both diabetes and dyslipidemia, induced MV modifications leading to up-regulation of miR-218, miR-212, miR-200b and miR-143 and down-regulation of miR-21, miR-122, miR-126 and miR-155 carried by MVs in diabetic dyslipidemia. It is important to point out that the same MV miRNAs showed significant correlations with important plasma markers, representative of this pathology. Thus, miR-218 was negatively correlated with the levels of ErMVs, whereas miR-21 was positively linked with cholesterol plasma levels, miR-122 was negatively associated with LDL-c levels, miR-126 was positively connected with HbA1c and PMV levels, and miR-155 was positively related with cholesterol and total MV levels and in the same time negatively connected with HDL-c levels.
MicroRNA expression profile in plasma
Since serum or plasma miRNAs are considered potential diagnostic biomarkers in diabetes [95,96,97] the same miRNAs detected above in plasma MVs have been investigated in circulation. However, there are only few studies to investigate the circulating miRNAs in diabetic dyslipidemia.
Therefore, we measured the levels of miR-21, miR-34a, miR-132, miR-143, miR-223, and we found them significantly up-regulated in plasma from T2D patients with dyslipidemia compared to all the other experimental groups (Fig. 7A–E). Although our results did not show any significant changes in the expressions of these miRNAs neither in diabetes nor in dyslipidemia, we believe that the significantly increased miRNA plasma levels found in diabetes associated with dyslipidemia could be the result of cumulative effect of these two diseases.

Relative expression (2−ΔCq × 103, ΔCq = Cq has-miRNA—Cq cel-miR-39) of miRNAs in plasma from patients with dyslipidemia (n = 14), diabetes (n = 17), diabetes associated with dyslipidemia (n = 17) and of healthy subjects taken as control (n = 15): A miR-21-5p; B miR-34a-5p; C miR 132-3p; D miR-143-3p; E miR-223-3p. Data are expressed as means ± SD. &p < 0.05 (vs C), *p < 0.05 (vs DLP), #p < 0.05 (vs T2D), &&/**/##p < 0.01 (One-Way ANOVA with Bonferroni's Multiple Comparison Test)
MiR-21, known as a key player of cell proliferation and apoptosis, was frequently up-regulated in many types of diseases, being considered diagnostic marker and therapeutic modulator for T2D [98]. Circulating miR-21 may act as powerful indicator even for an early detection of glucose adjustments in pre-diabetes stages [99]. Previous data highlighted the idea that plasma miR-21 is up-regulated in pre-diabetes [99], in T2D [99, 100] and in proliferative diabetic retinopathy [100]. Levels of miR-21 have been linked with clinical metabolic parameters characteristic for obesity, BMI and waist circumference [101]. We have also found a slightly but not significant increase of miR-21 levels in circulation of diabetic patients and in patients exhibiting dyslipidemia alone, compared to healthy individuals (Fig. 7A).
Similarly, people with diabetes or dyslipidemia revealed no significant modifications in circulating levels of miR-34a (Fig. 7B), one of the major miRNA involved in insulin production. MiR-34a was associated with insulin resistance being involved in pancreatic development, as well as in the onset of T2D [98, 102, 103]. There are contradictory data regarding circulating levels of miR-34a in diabetes. Plasma miR-34a was found reduced and negatively correlated with insulin and HOMA-IR [104]. Other data described increased expression of miR-34a in serum samples from T2D patients compared with pre-diabetic or T2D susceptible individuals [105].
Even if, we found no statistically modified expression of circulating miR-132 in diabetes or dyslipidemia (Fig. 7C), very recently, it was demonstrated that circulating levels of miR-132 were decreased in diabetic retinopathy patients in comparison to the ones without retinopathy, and they were inversely correlated with VEGF levels and oxidative stress markers [106].
Contradictory data also exist regarding mir-143 and miR-223 from circulation. Peripheral blood levels of miR-143 have been found down-regulated in the of patients with gestational diabetes [78], whereas elevated levels of miR-143 gene expression were correlated with the degree of obesity and proposed as predictor of insulin resistance in people without diabetes [107]. In line with previous studies, we also described a slightly increased level of miR-143 (Fig. 7D) in sera from patients with dyslipidemia, but this was not statistically significant. Likewise, in our study the differences in miR-223 expression were not significant between groups of patients with T2D, dyslipidemia and control group (Fig. 7E). Low levels of circulating miR-223 were considered marker for T2D onset [108]. Also, hyperglycemia induced increased levels of miR-223, in patients with acute coronary syndrome or stabile angina [109].
In our study we also investigated the levels of miR-122, miR-155, miR-200b, miR-218, miR-212 and miR-126 in plasma from DLP, T2D, T2D-DLP patient groups and control group (Fig. 8A–F). For miR-122 and miR-155 the expression levels were significantly higher in diabetic dyslipidemia (T2D-DLP group) compared to all other studied groups. Additionally, we found markedly increased levels of miR-122 (Fig. 8A) and miR-155 (Fig. 8B) in diabetic patients (T2D group) compared with the healthy subjects (C group). Circulating miR-122 has been found significantly up-regulated in individuals with T2D. A positive correlation between miR-122 and insulin levels, HOMA2-IR index [110], markers of obesity associated inflammation (TNFα, IL-1Ra, procalcitonin) [111,112,113,114,115] were also detected. Moreover, miR-122 has been associated with increased liver enzymes, adiposity, inflammation, and insulin resistance and with a contrasting lipid profile (higher triglycerides and lower HDL) [83]. Regarding mir 155, its expression in diabetes is a little controversial.

Relative expression (2−ΔCq × 103, ΔCq = Cq has-miRNA—Cq cel-miR-39) of miRNAs in plasma from patients with dyslipidemia (n = 14), diabetes (n = 17), diabetes associated with dyslipidemia (n = 17) and control subjects (n = 15): A miR-122-5p; B miR-155-5p; C miR 200-3p; D miR-218-5p; E miR-212-5p; F miR-126-3p. Data are expressed as means ± SD. &p < 0.05 (vs C), *p < 0.05 (vs DLP), # p < 0.05 (vs T2D) (One-Way ANOVA with Bonferroni's Multiple Comparison Test)
Dysregulation of miR-155 expression was proven to predict the severity of diabetes as well as the development of diabetes nephropathy, neuropathy, and retinopathy. A number of recent studies reported low serum levels of miR-155 in patients with T2D compared to healthy individuals [116,117,118]; while other data certify increased miR-155 in plasma or serum from T1D [119] and diabetes complications like nephropathy [120, 121] and retinopathy [92, 122]. MiR-155 was considered to be crucial for β-cell fitness [92]. Preclinical studies suggested that decreased miR-155 expression could be the key player in the mechanisms that underline the altered pancreatic function in T2D patients [117]. An insufficient level of miR-155 reduced β-cell growth in response to hypercholesterolemia [123], and only an up-regulation could help β-cell adaptation to overnutrition [92]. Any interference in this process could markedly contribute in the transition from pre-diabetes to T2D [92]. Furthermore high-fat diet altered the phenotype of cells in the islets of Langerhans, thereby insulin-secreting β-cells being re-differentiated into glucagon-producing α-cells [124].
For miR-200b, miR-218, miR-212 and miR-126, the expression levels were not significantly higher in diabetic dyslipidemia (T2D-DLP group) compared to all other studied groups. Nevertheless, the results revealed the higher levels of miR-200b in patients exhibiting T2D and dyslipidemia when compared to patients with dyslipidemia alone (Fig. 8C). Also, in our study, markedly elevated levels of plasma miR-218 were identified in all groups of patients investigated (DLP, T2D, T2D-DLP) compared to control group (Fig. 8D). In diabetes the expressions of circulating miR-218 and miR-200b have not been well described so far. It is known that, in diabetic retinopathy, the levels of miR-218 were up-regulated [125], while the levels of miR-200b were down-regulated [126].
Another miRNA associated with glucose imbalance is miR-212. It has been identified at children susceptible to metabolic disorders since the age of 16-and 17 years, later, at the age of 18, being associated with the onset of pre-diabetes [127]. We found circulating miR-212 significantly up-regulated in T2D group compared to control one (Fig. 8E). Even if, circulating miR-212 has been identified as marker of atherosclerosis and has been associated with three of the standard cardiovascular risk markers, HbA1c, HDL cholesterol, and lipoprotein a [128], in our study the levels of circulating miR-212 were found little but not markedly raised in patients with dyslipidemia or diabetes associated with dyslipidemia compared to healthy individuals.
Decreased miR-126 has been previously detected in T2D patients [129,130,131], and in diabetic patients with retinopathy [130]. Liu and collaborators, described miR-126 as potential marker for treatment response, showing that serum miR-126 increased after 6 months of diet and physical exercises in subjects with impaired glucose tolerance, as well as in T2D patients, to the latter insulin treatment being added [131]. The results were in line with data obtained by Ramezani-Aliakbari and collaborators on animal model. They showed protective effect of trimetazidine (a medicine used to prevent angina attacks), on the altered plasmatic levels of miR-126 in diabetic rats [132]. Together with its target gene IRS-1, miR-126 may be potential molecular target for the prevention and treatment of diabetic retinopathy. Also, circulating miR-126 has been associated with obesity and T2D and could be used to diagnose T2D at early stages [130, 131]. In line with these findings, we have also found, decreased levels of sera miR-126 in all groups of patients, the only statistically significant modification registered in the group of patients with dyslipidemia alone (Fig. 8F).
We then investigated possible correlations between significantly modified plasma miRNAs and clinical (Table 8) or inflammatory parameters (Table 9) in diabetes dyslipidemia. Besides the positive correlation between miR-200b with hemoglobin levels in plasma (r = 0.96, p = 0.007), negative associations were also observed between both miR-122 and miR-143 with creatinine (r = 0.90, p = 0.03 for miR-122; r = 0.89, p = 0.04 for miR-143) and diastolic blood pressure (r = 0.93, p = 0.02 for miR-122; r = 0.94, p = 0.01 for miR-143), and between miR-132 with uric acid (r = 0.88, p = 0.05) and erythrocytes (r = 0.95, p = 0.01) plasma levels (Table 8). Moreover, Il-1β plasma levels were positively correlated with miR-223 (r = 0.91, p = 0.03) and miR-155 (r = 0.89, p = 0.04) levels and negatively with miR-21 (r = 0.88, p = 0.05) plasma levels (Table 9). Also, TNFα levels were positively correlated with miR-21 (r = 0.94, p = 0.01) and miR-34a (r = 0.96, p = 0.01) and negatively correlated with miR-223 (r = 0.89, p = 0.04) (Table 9).
Association of plasma miRNAs with circulating MVs and some of their subtypes in plasma of diabetes patients with dyslipidemia (Table 10), revealed positive powerful correlations between miR-126 and PMVs (r = 0.86, p = 0.05) and miR-155 and total MVs (r = 0.97, p = 0.005), and negative correlations between miR-218 and ErMVs (r = 0.94, p = 0.01) (Table 10).
Finally, we followed the relationships between plasma miRNAs and microvesicle-associated miRNAs (MV miRNAs) (Table 11) and found powerful positive correlations between the levels of plasma miR-21 and MV miR-223 (r = 0.95, p = 0.01), plasma miR-132 and MV miR-132 (r = 0.88, p = 0.04) and plasma miR-122 and MV miR-126 (r = 0.90, p = 0.03) (Table 11). In addition, some significant negative correlations between plasma miR-223 and MV miR-223 (r = 0.98, p = 0.002), plasma miR-126 and MV miR-122, (r = 0.90, p = 0.03) and plasma miR-155 and MV miR-223 (r = 0.97, p = 0.003) were also identified (Table 11).
Regarding the analysis of miRNAs in plasma, we can conclude that the levels of miR-21, miR-34a, miR-132, miR-143, miR-223, miR-122, miR-155, miR-200b and miR-218 were up-regulated in diabetic dyslipidemia (T2D-DLP group) as a cumulative result of the alterations induced by both T2D and dyslipidemia. These plasma miRNAs found modified in diabetic dyslipidemia presented significant correlations with some important markers associated with this pathology. Thus: plasma miR-21 was positively correlated with TNFα plasma levels and MV miR-223 and negatively correlated with Il-1β plasma levels; plasma miR-34a was positively associated with TNFα plasma levels; plasma miR-132 was positively connected with MV miR-132 and negatively with uric acid and erythrocytes plasma levels; plasma miR-143 was negatively related with creatinine and diastolic blood pressure; plasma miR-223 was positively linked with Il-1β plasma levels and negatively with TNFα plasma levels and MV miR-223 levels; plasma miR-122 was positively associated with MV miR-126 levels and negatively with creatinine and diastolic blood pressure; plasma miR-155 was positively correlated with Il-1β plasma levels and total MV levels and negatively with MV miR-223 levels; plasma miR-200b was positively linked with hemoglobin levels and plasma miR-218 was negatively related with the levels of ErMVs.
Of all, in diabetes dyslipidemia, alterations of two of the above miRNAs, were related with one of the diseases more than with the other one. Thus, the modified miR-122 was induced by dyslipidemia more than by diabetes in both plasma MVs and the whole circulation, whereas for miR-132 the association was obvious especially within plasma MVs. Because neither in diabetes, nor in dyslipidemia, the levels of miR-21, miR-143, miR-155 and miR-218 were not significantly modified, we can assume that the changes in their expression were the result of imbalances occurred in diabetes dyslipidemia itself more than in diabetes or dyslipidemia alone.
Taking into account that miRNAs contained in MVs significantly up-regulated were miR-218, miR-212, miR-200b and miR-143, and those significantly down-regulated were miR-21, miR-122, miR-126 and miR-155, and miRNAs in plasma significantly up-regulated were miR-21, miR-34a, miR-132, miR-143, miR-223, miR-122, miR-155 and miR-218, we can conclude that miRNAs common to MVs and plasma significantly increased were miR-218, miR-132 and miR-143. In addition, three miRNAs common to MV and plasma, namely miR-21, miR-122 and miR-155, which evolved in different directions of decrease and increase, respectively, were identified.
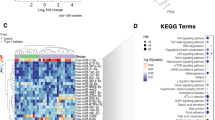
It is interesting to report that in diabetic dyslipidemia, of the three miRNAs whose levels were found higher in circulation as well as within plasma MVs, miR-143 was associated with Parkinson’s disease, miR-132 with hippo signaling pathway and proteoglycans in cancer, and miR-218 with genes involved in fatty acids biosynthesis, proteoglycans in cancer, viral carcinogenesis, pathways in cancer, biotin metabolism, ECM-extracellular matrix receptor interaction, hepatitis B, p53 signaling pathway (Fig. 9).

Heat map showing three similarly expressed miRNAs from circulation and MVs in T2D-DLP patients (n = 17)
Also, two of the three miRNAs found to be expressed differently in plasma and MVs of diabetic dyslipidemic patients, respectively miR-21 and miR-155, were associated with genes involved in various pathologies, namely: miR-21 in lysine degradation, fatty acid degradation, proteoglycans in cancer, colorectal cancer, and miR-155 in hepatitis B (Fig. 10).

Heat map showing three differentially expressed miRNAs from circulation and MVs in T2D-DLP patients (n = 17)
Strengths and limitations of the study
The study has several strong points that deserve to be considered: (1) The patients enrolled in the study did not have obesity or other disorders or even cardiovascular diseases. Also, they have not been exposed to infection or vaccination with COVID-19. Any of these conditions could have been potential sources of variation and could have affected the relevance of the study in the context of the investigated pathology, diabetic dyslipidemia; (2) The results regarding the expression of miRNAs in microvesicles and plasma were correlated with several metabolic and inflammatory parameters, constituting the basis for the development of future therapeutic strategies; (3) Identification of a specific set of miRNAs (miR-218, miR-132, miR-143, and miR-21, miR-122, miR-155), common to microvesicles and plasma and with a modified expression profile in diabetic dyslipidemia, makes these miRNAs to be considered potential biomarkers for the diagnosis and prognosis of this pathology, but also specific targets for the application of an effective therapy.
There are limitations of the study that need to be mentioned: (1) MVs collected from plasma isolated from all the participants, were stored at − 80 °C until use but not more than 2–3 weeks. Still, all the tests performed for their characterization proved that their number and structure were not affected by the freezing/thawing procedure; (2) Experiments characterizing MVs (large EVs) by Zetasizer Nano ZS, fluorescence and electron microscopy showed the presence of an extremely small, even negligible, percentage of smaller vesicles (exosomes/small EVs). It is important to note that, the majority population of vesicles was found in a range of values specific to MVs (100–1000 nm); (3) Circulating miRNAs was not quantified in the plasma depleted of MVs to be able to state with certainty that the miRNAs detected in the plasma belong to a rather large percentage to the MVs that the plasma contains; (4) To understand the origin of miRNAs expressed differently in MVs, the sorting of different types of MVs may help in future experiments; (5) Most of the patients with diabetes dyslipidemia were under treatment for diabetes and dyslipidemia and their BMI was higher compared to that of diabetic patients and control subjects; (6) Because these data are cross-sectional, a concluding remark on miRNA and MV evolution characteristic to a particular patient cannot be drown.
Despite all these limitations, the strengths of this study make our data pathophysiological relevant, demonstrating that MV levels, as well as their miRNA content, can represent a consistent tool for the early detection of diabetes complications.
Nevertheless, additional prospective studies are needed to certify the actual relevance of miR-218, miR-132, miR-143, and miR-21, miR-122, miR-155 as diagnostic and prognostic biomarkers of diabetes dyslipidemia in the overall population.
Conclusions
Our findings suggest that the presence of dyslipidemia more than diabetes in the pathology of diabetes dyslipidemia, seems to play an important role in the up-regulation and down-regulation of some miRNAs, such as miR-218, miR-132, miR-143, and miR-21, miR-122, miR-155, detected in both plasma and circulating MVs.
Up-regulated miR-218, miR-132 and miR-143 and down-regulated miR-21, miR-122, and miR-155 were associated with important plasma markers representative of diabetes dyslipidemia, but also with different signaling pathways, including p53 signaling pathway and hippo signaling pathway, and other pathologies such as Parkinson’s disease, colorectal cancer, hepatitis B. Also, these miRNAs were shown to control some process within the body: miR-218, miR-132 and miR-143 to control fatty acids biosynthesis, biotin metabolism, extracellular matrix-receptor interaction, and miR-21, miR-122, and miR-155 lysine and fatty acid degradation.
Understanding the regulatory networks of miR-143-3p, miR-132-3p, miR-218-5p and miR-21, miR-122, and miR-155 is essential for the development of therapies for diabetes dyslipidemia as well as for the identification of new biomarkers which will reflect the severity as well as the progression of the disease.
Availability of data and materials
The original contributions presented in the study are included in the article, additional inquiries can be addressed to the corresponding author.
Abbreviations
- miRNAs:
-
MicroRNAs
- MVs:
-
Microvesicles
- DDLP:
-
Diabetic dyslipidemia
- LDL:
-
Low-density lipoprotein
- HDL:
-
High-density lipoprotein
- T2D:
-
Type 2 diabetes
- PS:
-
Phosphatidylserine
- EMPs:
-
Endothelial microparticles
- PMPs:
-
Platelet microparticles
- LeMPs:
-
Leukocyte microparticles
- ECs:
-
Endothelial cells
- EPCs:
-
Endothelial progenitor cells
- mRNA:
-
Messenger RNA
- BMI:
-
Body mass index
- T2D-DLP:
-
Type 2 diabetes and dyslipidemia
- DLP:
-
Dyslipidemia
- C:
-
Control
- HbA1C:
-
Glycated haemoglobin
- IL-6:
-
Interleukin 6
- IL-1β:
-
Interleukin 1beta
- IFNγ:
-
Interferon gamma
- TNFα:
-
Tumor necrosis factor alfa
- PFP:
-
Phosphate buffered saline
- FBS:
-
Fetal bovine serum
- RT:
-
Room temperature
- qRT-PCR:
-
Quantitative reverse transcription polymerase chain reaction
- snRNA:
-
Small nuclear RNA
- Cq:
-
Quantification cycle
- EMVs:
-
Microvesicles derived from endothelial cells
- PMVs:
-
Microvesicles derived from platelets
- LeMVs:
-
Microvesicles derived from leucocytes
- MMVs:
-
Microvesicles derived monocytes
- ErMVs:
-
Microvesicles derived from erythrocytes
- PreDM:
-
Pre-diabetic
- NGT:
-
Normal glucose tolerance
- MV miRNAs:
-
Microvesicle-associated microRNAs
References
Goldberg IJ. Clinical review 124: Diabetic dyslipidemia: causes and consequences. J Clin Endocrinol Metab. 2001;86(3):965–71.
Krauss RM. Heterogeneity of plasma low-density lipoproteins and atherosclerosis risk. Curr Opin Lipidol. 1994;5(5):339–49.
Adiels M, Olofsson SO, Taskinen MR, Boren J. Overproduction of very low-density lipoproteins is the hallmark of the dyslipidemia in the metabolic syndrome. Arterioscler Thromb Vasc Biol. 2008;28(7):1225–36.
Guerin M, Le Goff W, Lassel TS, Van Tol A, Steiner G, Chapman MJ. Atherogenic role of elevated CE transfer from HDL to VLDL(1) and dense LDL in type 2 diabetes: impact of the degree of triglyceridemia. Arterioscler Thromb Vasc Biol. 2001;21(2):282–8.
Drew BG, Rye KA, Duffy SJ, Barter P, Kingwell BA. The emerging role of HDL in glucose metabolism. Nat Rev Endocrinol. 2012;8(4):237–45.
Lorenzo C, Hartnett S, Hanley AJ, Rewers MJ, Wagenknecht LE, Karter AJ, et al. Impaired fasting glucose and impaired glucose tolerance have distinct lipoprotein and apolipoprotein changes: the insulin resistance atherosclerosis study. J Clin Endocrinol Metab. 2013;98(4):1622–30.
Parrizas M, Novials A. Circulating microRNAs as biomarkers for metabolic disease. Best Pract Res Clin Endocrinol Metab. 2016;30(5):591–601.
Keller MP, Choi Y, Wang P, Davis DB, Rabaglia ME, Oler AT, et al. A gene expression network model of type 2 diabetes links cell cycle regulation in islets with diabetes susceptibility. Genome Res. 2008;18(5):706–16.
Alexandru N, Andrei E, Niculescu L, Dragan E, Ristoiu V, Georgescu A. Microparticles of healthy origins improve endothelial progenitor cell dysfunction via microRNA transfer in an atherosclerotic hamster model. Acta Physiol. 2017;221(4):230–49.
Georgescu A, Simionescu M. Extracellular vesicles: versatile nanomediators, potential biomarkers and therapeutic agents in atherosclerosis and COVID-19-related thrombosis. Int J Mol Sci. 2021;22(11):5967.
Simionescu N, Zonda R, Petrovici AR, Georgescu A. The multifaceted role of extracellular vesicles in glioblastoma: microRNA nanocarriers for disease progression and gene therapy. Pharmaceutics. 2021;13(7):988.
Georgescu A, Alexandru N, Andrei E, Titorencu I, Dragan E, Tarziu C, et al. Circulating microparticles and endothelial progenitor cells in atherosclerosis: pharmacological effects of irbesartan. J Thromb Haemost. 2012;10(4):680–91.
Giannella A, Radu CM, Franco L, Campello E, Simioni P, Avogaro A, et al. Circulating levels and characterization of microparticles in patients with different degrees of glucose tolerance. Cardiovasc Diabetol. 2017;16(1):118.
Burger D, Turner M, Xiao F, Munkonda MN, Akbari S, Burns KD. High glucose increases the formation and pro-oxidative activity of endothelial microparticles. Diabetologia. 2017;60(9):1791–800.
Alexandru N, Andrei E, Safciuc F, Dragan E, Balahura AM, Badila E, et al. Intravenous administration of allogenic cell-derived microvesicles of healthy origins defend against atherosclerotic cardiovascular disease development by a direct action on endothelial progenitor cells. Cells. 2020;9(2):423.
Alexandru N, Badila E, Weiss E, Cochior D, Stepien E, Georgescu A. Vascular complications in diabetes: microparticles and microparticle associated microRNAs as active players. Biochem Biophys Res Commun. 2016;472(1):1–10.
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97.
Vickers KC, Remaley AT. Lipid-based carriers of microRNAs and intercellular communication. Curr Opin Lipidol. 2012;23(2):91–7.
Simionescu N, Nemecz M, Petrovici AR, Nechifor IS, Buga RC, Dabija MG, et al. Microvesicles and microvesicle-associated microRNAs reflect glioblastoma regression: microvesicle-associated miR-625–5p has biomarker potential. Int J Mol Sci. 2022;23(15):8398.
Nemecz M, Alexandru N, Tanko G, Georgescu A. Role of microRNA in endothelial dysfunction and hypertension. Curr Hypertens Rep. 2016;18(12):87.
Shantikumar S, Caporali A, Emanueli C. Role of microRNAs in diabetes and its cardiovascular complications. Cardiovasc Res. 2012;93(4):583–93.
Stepien EL, Durak-Kozica M, Kaminska A, Targosz-Korecka M, Libera M, Tylko G, et al. Circulating ectosomes: Determination of angiogenic microRNAs in type 2 diabetes. Theranostics. 2018;8(14):3874–90.
Fajans SS, Bell GI. MODY: history, genetics, pathophysiology, and clinical decision making. Diabetes Care. 2011;34(8):1878–84.
Vaisse C, Clement K, Guy-Grand B, Froguel P. A frameshift mutation in human MC4R is associated with a dominant form of obesity. Nat Genet. 1998;20(2):113–4.
Gabcova-Balaziova D, Stanikova D, Vohnout B, Huckova M, Stanik J, Klimes I, et al. Molecular-genetic aspects of familial hypercholesterolemia. Endocr Regul. 2015;49(3):164–81.
Georgescu A, Alexandru N, Andrei E, Dragan E, Cochior D, Dias S. Effects of transplanted circulating endothelial progenitor cells and platelet microparticles in atherosclerosis development. Biol Cell. 2016;108(8):219–43.
Comarita IK, Vilcu A, Constantin A, Procopciuc A, Safciuc F, Alexandru N, et al. Therapeutic potential of stem cell-derived extracellular vesicles on atherosclerosis-induced vascular dysfunction and its key molecular players. Front Cell Dev Biol. 2022;10:817180.
Niculescu LS, Simionescu N, Sanda GM, Carnuta MG, Stancu CS, Popescu AC, et al. MiR-486 and miR-92a identified in circulating HDL discriminate between stable and vulnerable coronary artery disease patients. PLoS ONE. 2015;10(10):e0140958.
Alexandru N, Constantin A, Nemecz M, Comarita IK, Vilcu A, Procopciuc A, et al. hypertension associated with hyperlipidemia induced different microRNA expression profiles in plasma, platelets, and platelet-derived microvesicles; effects of endothelial progenitor cell therapy. Front Med. 2019;6:280.
Alfadul H, Sabico S, Al-Daghri NM. The role of interleukin-1beta in type 2 diabetes mellitus: a systematic review and meta-analysis. Front Endocrinol. 2022;13:901616.
Tong HV, Luu NK, Son HA, Hoan NV, Hung TT, Velavan TP, et al. Adiponectin and pro-inflammatory cytokines are modulated in Vietnamese patients with type 2 diabetes mellitus. J Diabetes Investig. 2017;8(3):295–305.
Zhao G, Dharmadhikari G, Maedler K, Meyer-Hermann M. Possible role of interleukin-1beta in type 2 diabetes onset and implications for anti-inflammatory therapy strategies. PLoS Comput Biol. 2014;10(8):e1003798.
Maedler K, Schumann DM, Sauter N, Ellingsgaard H, Bosco D, Baertschiger R, et al. Low concentration of interleukin-1beta induces FLICE-inhibitory protein-mediated beta-cell proliferation in human pancreatic islets. Diabetes. 2006;55(10):2713–22.
Banerjee J, Dhas Y, Mishra N. Middle-aged Indians with type 2 diabetes are at higher risk of biological ageing with special reference to serum CDKN2A. J Diabetes Res. 2020;2020:7569259.
Mo D, Liu S, Ma H, Tian H, Yu H, Zhang X, et al. Effects of acarbose and metformin on the inflammatory state in newly diagnosed type 2 diabetes patients: a one-year randomized clinical study. Drug Des Devel Ther. 2019;13:2769–76.
Ayelign B, Negash M, Andualem H, Wondemagegn T, Kassa E, Shibabaw T, et al. Association of IL-10 (- 1082 A/G) and IL-6 (- 174 G/C) gene polymorphism with type 2 diabetes mellitus in Ethiopia population. BMC Endocr Disord. 2021;21(1):70.
Spranger J, Kroke A, Mohlig M, Hoffmann K, Bergmann MM, Ristow M, et al. Inflammatory cytokines and the risk to develop type 2 diabetes: results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes. 2003;52(3):812–7.
Alexandru N, Procopciuc A, Vilcu A, Comarita IK, Bӑdilӑ E, Georgescu A. Extracellular vesicles-incorporated microRNA signature as biomarker and diagnosis of prediabetes state and its complications. Rev Endocr Metab Disord. 2022;23(3):309–32.
Mihaela G, Nicoleta A, Stefania Lucia M, Alina C, Miruna N, Alexandru F, et al. Part one: extracellular vesicles as valuable players in diabetic cardiovascular diseases. In: De Ana Gil B, Jose Antonio Reales C, editors., et al., Extracellular vesicles and their importance in human health. Rijeka: IntechOpen; 2019. p. 6.
Diehl P, Fricke A, Sander L, Stamm J, Bassler N, Htun N, et al. Microparticles: major transport vehicles for distinct microRNAs in circulation. Cardiovasc Res. 2012;93(4):633–44.
Melki I, Tessandier N, Zufferey A, Boilard E. Platelet microvesicles in health and disease. Platelets. 2017;28(3):214–21.
Zaldivia MTK, McFadyen JD, Lim B, Wang X, Peter K. Platelet-derived microvesicles in cardiovascular diseases. Front Cardiovasc Med. 2017;4:74.
Suades R, Padro T, Crespo J, Sionis A, Alonso R, Mata P, et al. Liquid biopsy of extracellular microvesicles predicts future major ischemic events in genetically characterized familial hypercholesterolemia patients. Arterioscler Thromb Vasc Biol. 2019;39(6):1172–81.
Sabatier F, Darmon P, Hugel B, Combes V, Sanmarco M, Velut JG, et al. Type 1 and type 2 diabetic patients display different patterns of cellular microparticles. Diabetes. 2002;51(9):2840–5.
Freeman DW, Noren Hooten N, Eitan E, Green J, Mode NA, Bodogai M, et al. Altered extracellular vesicle concentration, cargo, and function in diabetes. Diabetes. 2018;67(11):2377–88.
Randriamboavonjy V, Mann WA, Elgheznawy A, Popp R, Rogowski P, Dornauf I, et al. Metformin reduces hyper-reactivity of platelets from patients with polycystic ovary syndrome by improving mitochondrial integrity. Thromb Haemost. 2015;114(3):569–78.
Martinez MC, Andriantsitohaina R. Extracellular vesicles in metabolic syndrome. Circ Res. 2017;120(10):1674–86.
Nomura S, Shouzu A, Omoto S, Hayakawa T, Kagawa H, Nishikawa M, et al. Effect of cilostazol on soluble adhesion molecules and platelet-derived microparticles in patients with diabetes. Thromb Haemost. 1998;80(3):388–92.
Omoto S, Nomura S, Shouzu A, Hayakawa T, Shimizu H, Miyake Y, et al. Significance of platelet-derived microparticles and activated platelets in diabetic nephropathy. Nephron. 1999;81(3):271–7.
Shouzu A, Nomura S, Hayakawa T, Omoto S, Shimizu H, Miyake Y, et al. Effect of sarpogrelate hydrochloride on platelet-derived microparticles and various soluble adhesion molecules in diabetes mellitus. Clin Appl Thromb Hemost. 2000;6(3):139–43.
Noren Hooten N, Evans MK. Extracellular vesicles as signaling mediators in type 2 diabetes mellitus. Am J Physiol Cell Physiol. 2020;318(6):C1189–99.
Balaphas A, Meyer J, Sadoul K, Fontana P, Morel P, Gonelle-Gispert C, et al. Platelets and platelet-derived extracellular vesicles in liver physiology and disease. Hepatol Commun. 2019;3(7):855–66.
Paola GS. Extracellular vesicles number and cell subtype may be influenced by diabetes mellitus and metformin in patients at high cardiovascular risk. Nutr Metabol Cardiovasc Dis. 2022. https://doi.org/10.1016/j.numecd.2022.09.010.
Suades R, Padro T, Alonso R, Mata P, Badimon L. Lipid-lowering therapy with statins reduces microparticle shedding from endothelium, platelets and inflammatory cells. Thromb Haemost. 2013;110(2):366–77.
Giro O, Jimenez A, Pane A, Badimon L, Ortega E, Chiva-Blanch G. Extracellular vesicles in atherothrombosis and cardiovascular disease: Friends and foes. Atherosclerosis. 2021;330:61–75.
Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–9.
Srinivasan S, Selvan ST, Archunan G, Gulyas B, Padmanabhan P. MicroRNAs -the next generation therapeutic targets in human diseases. Theranostics. 2013;3(12):930–42.
Zhu Z, Zhang Y, Bai R, Yang R, Shan Z, Ma C, et al. Association of genetic polymorphisms in microRNAs with type 2 diabetes mellitus in a Chinese population. Front Endocrinol. 2020;11:587561.
De Rosa S, Arcidiacono B, Chiefari E, Brunetti A, Indolfi C, Foti DP. Type 2 diabetes mellitus and cardiovascular disease: genetic and epigenetic links. Front Endocrinol. 2018;9:2.
Mone P, Lombardi A, Kansakar U, Varzideh F, Jankauskas SS, Pansini A, et al. Empagliflozin improves the microRNA signature of endothelial dysfunction in patients with heart failure with preserved ejection fraction and diabetes. J Pharmacol Exp Ther. 2023;384(1):116–22.
Lew JK, Pearson JT, Schwenke DO, Katare R. Exercise mediated protection of diabetic heart through modulation of microRNA mediated molecular pathways. Cardiovasc Diabetol. 2017;16(1):10.
Kong Q, Guo X, Guo Z, Su T. Urinary exosome miR-424 and miR-218 as biomarkers for type 1 diabetes in children. Clin Lab. 2019. https://doi.org/10.7754/Clin.Lab.2018.180921.
Ibrahim AA, Wahby AA, Ashmawy I, Saleh RM, Soliman H. Association of exosomal miR-34a with markers of dyslipidemia and endothelial dysfunction in children and adolescents with T1DM. J Clin Res Pediatr Endocrinol. 2020;12(4):401–9.
Stepien E, Stankiewicz E, Zalewski J, Godlewski J, Zmudka K, Wybranska I. Number of microparticles generated during acute myocardial infarction and stable angina correlates with platelet activation. Arch Med Res. 2012;43(1):31–5.
Malm HA, Mollet IG, Berggreen C, Orho-Melander M, Esguerra JL, Goransson O, et al. Transcriptional regulation of the miR-212/miR-132 cluster in insulin-secreting beta-cells by cAMP-regulated transcriptional co-activator 1 and salt-inducible kinases. Mol Cell Endocrinol. 2016;424:23–33.
Mollet IG, Malm HA, Wendt A, Orho-Melander M, Eliasson L. Integrator of stress responses calmodulin binding transcription activator 1 (Camta1) regulates miR-212/miR-132 expression and insulin secretion. J Biol Chem. 2016;291(35):18440–52.
Eliasson L, Esguerra JLS. MicroRNA networks in pancreatic islet cells: normal function and type 2 diabetes. Diabetes. 2020;69(5):804–12.
Mziaut H, Henniger G, Ganss K, Hempel S, Wolk S, McChord J, et al. MiR-132 controls pancreatic beta cell proliferation and survival through Pten/Akt/Foxo3 signaling. Mol Metab. 2020;31:150–62.
Liu C, Feng H, Zhang L, Guo Y, Ma J, Yang L. MicroRNA-143-3p levels are reduced in the peripheral blood of patients with gestational diabetes mellitus and influences pancreatic beta-cell function and viability. Exp Ther Med. 2023;25(2):81.
Tagoma A, Alnek K, Kirss A, Uibo R, Haller-Kikkatalo K. MicroRNA profiling of second trimester maternal plasma shows upregulation of miR-195-5p in patients with gestational diabetes. Gene. 2018;672:137–42.
Zhao C, Dong J, Jiang T, Shi Z, Yu B, Zhu Y, et al. Early second-trimester serum miRNA profiling predicts gestational diabetes mellitus. PLoS ONE. 2011;6(8):e23925.
Zhou X, Xiang C, Zheng X. miR-132 serves as a diagnostic biomarker in gestational diabetes mellitus and its regulatory effect on trophoblast cell viability. Diagn Pathol. 2019;14(1):119.
Meng Y, Eirin A, Zhu XY, Tang H, Chanana P, Lerman A, et al. The metabolic syndrome alters the miRNA signature of porcine adipose tissue-derived mesenchymal stem cells. Cytometry A. 2018;93(1):93–103.
Lakhter AJ, Pratt RE, Moore RE, Doucette KK, Maier BF, DiMeglio LA, et al. Beta cell extracellular vesicle miR-21-5p cargo is increased in response to inflammatory cytokines and serves as a biomarker of type 1 diabetes. Diabetologia. 2018;61(5):1124–34.
Akerman L, Casas R, Ludvigsson J, Tavira B, Skoglund C. Serum miRNA levels are related to glucose homeostasis and islet autoantibodies in children with high risk for type 1 diabetes. PLoS ONE. 2018;13(1):e0191067.
Tesovnik T, Kovac J, Pohar K, Hudoklin S, Dovc K, Bratina N, et al. Extracellular vesicles derived human-miRNAs modulate the immune system in type 1 diabetes. Front Cell Dev Biol. 2020;8:202.
Castano C, Kalko S, Novials A, Parrizas M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc Natl Acad Sci USA. 2018;115(48):12158–63.
Liu X, Qiu H. Placenta-derived exosomes and gestational diabetes mellitus. Diabetes Metab Syndr Obes. 2022;15:1391–404.
Peng H, Hou M, Wu Z, Wang J, Zhou M, Zhuang X, et al. Plasma exosomal miR-122 regulates the efficacy of metformin via AMPK in type 2 diabetes and hepatocellular carcinoma. Heliyon. 2022;8(11):e11503.
Willeit P, Skroblin P, Kiechl S, Fernandez-Hernando C, Mayr M. Liver microRNAs: potential mediators and biomarkers for metabolic and cardiovascular disease? Eur Heart J. 2016;37(43):3260–6.
Barajas JM, Reyes R, Guerrero MJ, Jacob ST, Motiwala T, Ghoshal K. The role of miR-122 in the dysregulation of glucose-6-phosphate dehydrogenase (G6PD) expression in hepatocellular cancer. Sci Rep. 2018;8(1):9105.
Fernandez-Hernando C, Ramirez CM, Goedeke L, Suarez Y. MicroRNAs in metabolic disease. Arterioscler Thromb Vasc Biol. 2013;33(2):178–85.
Willeit P, Skroblin P, Moschen AR, Yin X, Kaudewitz D, Zampetaki A, et al. Circulating microRNA-122 is associated with the risk of new-onset metabolic syndrome and type 2 diabetes. Diabetes. 2017;66(2):347–57.
Hunter MP, Ismail N, Zhang X, Aguda BD, Lee EJ, Yu L, et al. Detection of microRNA expression in human peripheral blood microvesicles. PLoS ONE. 2008;3(11):e3694.
Laffont B, Corduan A, Ple H, Duchez AC, Cloutier N, Boilard E, et al. Activated platelets can deliver mRNA regulatory Ago2*microRNA complexes to endothelial cells via microparticles. Blood. 2013;122(2):253–61.
Zhang YY, Zhou X, Ji WJ, Shi R, Lu RY, Li JL, et al. Decreased circulating microRNA-223 level predicts high on-treatment platelet reactivity in patients with troponin-negative non-ST elevation acute coronary syndrome. J Thromb Thrombol. 2014;38(1):65–72.
Chyrchel B, Toton-Zuranska J, Kruszelnicka O, Chyrchel M, Mielecki W, Kolton-Wroz M, et al. Association of plasma miR-223 and platelet reactivity in patients with coronary artery disease on dual antiplatelet therapy: a preliminary report. Platelets. 2015;26(6):593–7.
Jansen F, Wang H, Przybilla D, Franklin BS, Dolf A, Pfeifer P, et al. Vascular endothelial microparticles-incorporated microRNAs are altered in patients with diabetes mellitus. Cardiovasc Diabetol. 2016;15:49.
Olivieri F, Spazzafumo L, Bonafe M, Recchioni R, Prattichizzo F, Marcheselli F, et al. MiR-21-5p and miR-126a-3p levels in plasma and circulating angiogenic cells: relationship with type 2 diabetes complications. Oncotarget. 2015;6(34):35372–82.
Jansen F, Yang X, Hoelscher M, Cattelan A, Schmitz T, Proebsting S, et al. Endothelial microparticle-mediated transfer of MicroRNA-126 promotes vascular endothelial cell repair via SPRED1 and is abrogated in glucose-damaged endothelial microparticles. Circulation. 2013;128(18):2026–38.
Zampetaki A, Kiechl S, Drozdov I, Willeit P, Mayr U, Prokopi M, et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ Res. 2010;107(6):810–7.
Jankauskas SS, Gambardella J, Sardu C, Lombardi A, Santulli G. Functional role of miR-155 in the pathogenesis of diabetes mellitus and its complications. Noncoding RNA. 2021;7(3):39.
Mazzeo A, Beltramo E, Lopatina T, Gai C, Trento M, Porta M. Molecular and functional characterization of circulating extracellular vesicles from diabetic patients with and without retinopathy and healthy subjects. Exp Eye Res. 2018;176:69–77.
Barutta F, Tricarico M, Corbelli A, Annaratone L, Pinach S, Grimaldi S, et al. Urinary exosomal microRNAs in incipient diabetic nephropathy. PLoS ONE. 2013;8(11):e73798.
Kraczkowska W, Stachowiak L, Plawski A, Jagodzinski PP. Circulating miRNA as potential biomarkers for diabetes mellitus type 2: should we focus on searching for sex differences? J Appl Genet. 2022;63(2):293–303.
Guay C, Regazzi R. Circulating microRNAs as novel biomarkers for diabetes mellitus. Nat Rev Endocrinol. 2013;9(9):513–21.
Mirra P, Raciti GA, Nigro C, Fiory F, D’Esposito V, Formisano P, et al. Circulating miRNAs as intercellular messengers, potential biomarkers and therapeutic targets for Type 2 diabetes. Epigenomics. 2015;7(4):653–67.
Chakraborty C, Doss CG, Bandyopadhyay S, Agoramoorthy G. Influence of miRNA in insulin signaling pathway and insulin resistance: micro-molecules with a major role in type-2 diabetes. Wiley Interdiscip Rev RNA. 2014;5(5):697–712.
La Sala L, Mrakic-Sposta S, Tagliabue E, Prattichizzo F, Micheloni S, Sangalli E, et al. Circulating microRNA-21 is an early predictor of ROS-mediated damage in subjects with high risk of developing diabetes and in drug-naive T2D. Cardiovasc Diabetol. 2019;18(1):18.
Jiang Q, Lyu XM, Yuan Y, Wang L. Plasma miR-21 expression: an indicator for the severity of Type 2 diabetes with diabetic retinopathy. Biosci Rep. 2017;37(2):BSR20160589.
Ghorbani S, Mahdavi R, Alipoor B, Panahi G, Nasli Esfahani E, Razi F, et al. Decreased serum microRNA-21 level is associated with obesity in healthy and type 2 diabetic subjects. Arch Physiol Biochem. 2018;124(4):300–5.
Wei R, Yang J, Liu GQ, Gao MJ, Hou WF, Zhang L, et al. Dynamic expression of microRNAs during the differentiation of human embryonic stem cells into insulin-producing cells. Gene. 2013;518(2):246–55.
Mone P, de Donato A, Varzideh F, Kansakar U, Jankauskas SS, Pansini A, et al. Functional role of miR-34a in diabetes and frailty. Front Aging. 2022;3:949924.
Ahmadpour F, Nourbakhsh M, Razzaghy-Azar M, Khaghani S, Alipoor B, Abdolvahabi Z, et al. The association of plasma levels of miR-34a AND miR-149 with obesity and insulin resistance in obese children and adolescents. Acta Endocrinol. 2018;14(2):149–54.
Kong L, Zhu J, Han W, Jiang X, Xu M, Zhao Y, et al. Significance of serum microRNAs in pre-diabetes and newly diagnosed type 2 diabetes: a clinical study. Acta Diabetol. 2011;48(1):61–9.
Pramanik S, Saha C, Chowdhury S, Bose C, Bhattacharyya NP, Mondal LK. Decreased levels of miR-126 and miR-132 in plasma and vitreous humor of non-proliferative diabetic retinopathy among subjects with Type-2 diabetes mellitus. Diabetes Metab Syndr Obes. 2022;15:345–58.
Yuzbashian E, de Campos Zani SC, Zarkash M, Asghari G, Hedayati M, Khalaj A, et al. Elevated miR-143 and miR-34a gene expression in human visceral adipose tissue are associated with insulin resistance in non-diabetic adults: a cross-sectional study. Eat Weight Disord. 2022;27(8):3419–28.
Sanchez-Ceinos J, Rangel-Zuniga OA, Clemente-Postigo M, Podadera-Herreros A, Camargo A, Alcala-Diaz JF, et al. miR-223-3p as a potential biomarker and player for adipose tissue dysfunction preceding type 2 diabetes onset. Mol Ther Nucleic Acids. 2021;23:1035–52.
Simionescu N, Niculescu LS, Carnuta MG, Sanda GM, Stancu CS, Popescu AC, et al. Hyperglycemia determines increased specific microRNAs levels in sera and HDL of acute coronary syndrome patients and stimulates microRNAs production in human macrophages. PLoS ONE. 2016;11(8):e0161201.
Mononen N, Lyytikainen LP, Seppala I, Mishra PP, Juonala M, Waldenberger M, et al. Whole blood microRNA levels associate with glycemic status and correlate with target mRNAs in pathways important to type 2 diabetes. Sci Rep. 2019;9(1):8887.
Lischka J, Schanzer A, Hojreh A, Ba-Ssalamah A, de Gier C, Valent I, et al. Circulating microRNAs 34a, 122, and 192 are linked to obesity-associated inflammation and metabolic disease in pediatric patients. Int J Obes. 2021;45(8):1763–72.
Bussler S, Penke M, Flemming G, Elhassan YS, Kratzsch J, Sergeyev E, et al. Novel insights in the metabolic syndrome in childhood and adolescence. Horm Res Paediatr. 2017;88(3–4):181–93.
El Kassas GM, Shehata MA, El Wakeel MA, Amer AF, Elzaree FA, Darwish MK, et al. Role of procalcitonin as an inflammatory marker in a sample of Egyptian children with simple obesity. Open Access Maced J Med Sci. 2018;6(8):1349–53.
Abbasi A, Corpeleijn E, Postmus D, Gansevoort RT, de Jong PE, Gans RO, et al. Plasma procalcitonin is associated with obesity, insulin resistance, and the metabolic syndrome. J Clin Endocrinol Metab. 2010;95(9):E26-31.
Puder JJ, Varga S, Kraenzlin M, De Geyter C, Keller U, Muller B. Central fat excess in polycystic ovary syndrome: relation to low-grade inflammation and insulin resistance. J Clin Endocrinol Metab. 2005;90(11):6014–21.
Akhbari M, Khalili M, Shahrabi-Farahani M, Biglari A, Bandarian F. Expression level of circulating cell free miR-155 gene in serum of patients with diabetic nephropathy. Clin Lab. 2019;65(8):1493–9.
Lin X, Qin Y, Jia J, Lin T, Lin X, Chen L, et al. MiR-155 enhances insulin sensitivity by coordinated regulation of multiple genes in mice. PLoS Genet. 2016;12(10):e1006308.
Polina ER, Oliveira FM, Sbruzzi RC, Crispim D, Canani LH, Santos KG. Gene polymorphism and plasma levels of miR-155 in diabetic retinopathy. Endocr Connect. 2019;8(12):1591–9.
Assmann TS, Recamonde-Mendoza M, Punales M, Tschiedel B, Canani LH, Crispim D. MicroRNA expression profile in plasma from type 1 diabetic patients: case-control study and bioinformatic analysis. Diabetes Res Clin Pract. 2018;141:35–46.
Bai X, Luo Q, Tan K, Guo L. Diagnostic value of VDBP and miR-155-5p in diabetic nephropathy and the correlation with urinary microalbumin. Exp Ther Med. 2020;20(5):86.
Wang G, Wu B, Zhang B, Wang K, Wang H. LncRNA CTBP1-AS2 alleviates high glucose-induced oxidative stress, ECM accumulation, and inflammation in diabetic nephropathy via miR-155-5p/FOXO1 axis. Biochem Biophys Res Commun. 2020;532(2):308–14.
Prado MSG, de Jesus ML, de Goes TC, Mendonca LSO, Kaneto CM. Downregulation of circulating miR-320a and target gene prediction in patients with diabetic retinopathy. BMC Res Notes. 2020;13(1):155.
Zhu M, Wei Y, Geissler C, Abschlag K, Corbalan Campos J, Hristov M, et al. Hyperlipidemia-Induced microRNA-155-5p Improves beta-cell function by targeting Mafb. Diabetes. 2017;66(12):3072–84.
Lu J, Hamze Z, Bonnavion R, Herath N, Pouponnot C, Assade F, et al. Reexpression of oncoprotein MafB in proliferative beta-cells and Men1 insulinomas in mouse. Oncogene. 2012;31(31):3647–54.
Yao R, Yao X, Liu R, Peng J, Tian T. Glucose-induced microRNA-218 suppresses the proliferation and promotes the apoptosis of human retinal pigment epithelium cells by targeting RUNX2. Biosci Rep. 2019;39(12):BSR20192580.
da Costa ESMED, Polina ER, Crispim D, Sbruzzi RC, Lavinsky D, Mallmann F, et al. Plasma levels of miR-29b and miR-200b in type 2 diabetic retinopathy. J Cell Mol Med. 2019;23(2):1280–7.
Joglekar MV, Kunte PS, Wong WKM, Bhat DS, Satoor SN, Patil RR, et al. Circulating microRNAs from early childhood and adolescence are associated with pre-diabetes at 18 years of age in women from the PMNS cohort. J Dev Orig Health Dis. 2022;13(6):806–11.
Jeong HS, Kim JY, Lee SH, Hwang J, Shin JW, Song KS, et al. Synergy of circulating miR-212 with markers for cardiovascular risks to enhance estimation of atherosclerosis presence. PLoS ONE. 2017;12(5):e0177809.
Yazdanpanah Z, Kazemipour N, Kalantar SM, Vahidi Mehrjardi MY. Plasma miR-21 as a potential predictor in prediabetic individuals with a positive family history of type 2 diabetes mellitus. Physiol Rep. 2022;10(2):e15163.
Zhang T, Li L, Shang Q, Lv C, Wang C, Su B. Circulating miR-126 is a potential biomarker to predict the onset of type 2 diabetes mellitus in susceptible individuals. Biochem Biophys Res Commun. 2015;463(1–2):60–3.
Liu Y, Gao G, Yang C, Zhou K, Shen B, Liang H, et al. The role of circulating microRNA-126 (miR-126): a novel biomarker for screening prediabetes and newly diagnosed type 2 diabetes mellitus. Int J Mol Sci. 2014;15(6):10567–77.
Ramezani-Aliakbari F, Badavi M, Dianat M, Mard SA, Ahangarpour A. Trimetazidine increases plasma microRNA-24 and microRNA-126 levels and improves dyslipidemia, inflammation and hypotension in diabetic rats. Iran J Pharm Res. 2020;19(3):248–57.
Acknowledgements
The authors appreciate the dedicated work of research assistant Marilena Isachi (biochemistry) and clinical assistant Camelia Otilia Tudor.
Funding
This work was supported by a grant from the Romanian National Authority for Scientific Research, CNCS–UEFISCDI, project no. PN-III-P1-1.2-PCCDI-2017-0527 (Contract no. 83PCCDI/2018) and by the Romanian Academy.
Author information
Authors and Affiliations
Contributions
AG and MN participated in study conception and design. MN, DSS, IKC, AC, CG and GT performed the acquisition of data. MN, IKC, AC and AG participated in analysis and interpretation of data. MN, IKC, AG drafted the manuscript and AG revised the manuscript. All authors read and approved the submitted version of manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study involving human participants was approved by the Ethics Committee of the Institute of Cellular Biology and Pathology “Nicolae Simionescu” in accordance with Decision no.11/08.08.2017 (project no. PN-III-P1-1.2-PCCDI2017-0527/Contract no. 83PCCDI/2018). The participants provided written informed consent for their contribution in this study.
Consent for publications
Not applicable.
Competing interests
The authors MN, DSS, IKC, AC, GT, CG, AG, declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Nemecz, M., Stefan, D.S., Comarița, I.K. et al. Microvesicle-associated and circulating microRNAs in diabetic dyslipidemia: miR-218, miR-132, miR-143, and miR-21, miR-122, miR-155 have biomarker potential. Cardiovasc Diabetol 22, 260 (2023). https://doi.org/10.1186/s12933-023-01988-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12933-023-01988-0