
Abstract
Background
Coronavirus disease 2019 (COVID-19) patients can develop pulmonary fibrosis (PF), which is associated with impaired outcome. We assessed specific leukocytic transcriptome profiles associated with PF and the influence of early dexamethasone (DEXA) treatment on the clinical course of PF in critically ill COVID-19 patients.
Methods
We performed a pre-post design study in 191 COVID-19 patients admitted to the Intensive Care Unit (ICU) spanning two treatment cohorts: the pre-DEXA- (n = 67) and the DEXA-cohort (n = 124). PF was identified based on radiological findings, worsening of ventilatory parameters and elevated circulating PIIINP levels. Longitudinal transcriptome profiles of 52 pre-DEXA patients were determined using RNA sequencing. Effects of prednisone treatment on clinical fibrosis parameters and outcomes were analyzed between PF- and no-PF-patients within both cohorts.
Results
Transcriptome analyses revealed upregulation of inflammatory, coagulation and neutrophil extracellular trap-related pathways in PF-patients compared to no-PF patients. Key genes involved included PADI4, PDE4D, MMP8, CRISP3, and BCL2L15. Enrichment of several identified pathways was associated with impaired survival in a external cohort of patients with idiopathic pulmonary fibrosis. Following prednisone treatment, PF-related profiles reverted towards those observed in the no-PF-group. Likewise, PIIINP levels decreased significantly following prednisone treatment. PF incidence was 28% and 25% in the pre-DEXA- and DEXA-cohort, respectively (p = 0.61). ICU length-of-stay (pre-DEXA: 42 [29–49] vs. 18 [13–27] days, p < 0.001; DEXA: 42 [28–57] vs. 13 [7–24] days, p < 0.001) and mortality (pre-DEXA: 47% vs. 15%, p = 0.009; DEXA: 61% vs. 19%, p < 0.001) were higher in the PF-groups compared to the no-PF-groups within both cohorts. Early dexamethasone therapy did not influence these outcomes.
Conclusions
ICU patients with COVID-19 who develop PF exhibit upregulated coagulation, inflammation, and neutrophil extracellular trap-related pathways as well as prolonged ICU length-of-stay and mortality. This study indicates that early dexamethasone treatment neither influences the incidence or clinical course of PF, nor clinical outcomes.
Similar content being viewed by others

Background
Patients with Coronavirus Disease 2019 (COVID-19)-induced Acute Respiratory Distress Syndrome (ARDS) are at risk of subsequent complications such as a pathological fibroproliferative response [1, 2]. Pulmonary fibrosis (PF) is associated with challenges in mechanical ventilation, prolonged length of stay (LOS) in ICU, higher mortality rates, and chronic symptoms in survivors [3,4,5,6,7].
It is challenging to detect PF at an early stage of ARDS. High N-terminal pro-peptide of type III procollagen (PIIINP) levels in bronchoalveolar lavage (BAL) fluid [8], as well as PIIINP in blood may be used. Also, other circulating fibrosis biomarkers such as hepatocyte growth factor (HGF) [9] and Macrophage Inflammatory Protein-3 alpha (MIP-3α) [10] could be of value. The mechanisms underlying the development of PF are largely unexplored, while knowledge of these pathways may aid early diagnosis and novel treatment targets. Currently, PF in patients with non-COVID-19 ARDS is treated with corticosteroids, which is effective in reducing time on mechanical ventilation and ICU-LOS, especially in those with elevated biomarker concentrations [11, 12].
Initially, care for critically ill COVID-19 patients was limited to supportive treatment. However, since early treatment with the corticosteroid dexamethasone (DEXA) was shown to be beneficial [13], hospitalized COVID-19 patients requiring oxygen suppletion were all treated with DEXA. It is however unknown whether DEXA treatment influences the incidence or severity of PF and whether or not it affects the therapeutic efficacy of later corticosteroid treatment in patients who develop PF. The primary aim of this study in critically ill COVID-19 patients was therefore twofold: (1) to explore transcriptome profiles associated with PF and the response to treatment using longitudinal RNA sequencing of circulating leukocytes. (2) to determine the influence of early dexamethasone treatment on the incidence and time to development of PF, and to assess the therapeutic efficacy of steroids to treat PF both before and after the introduction of early dexamethasone as standard care for critically ill COVID-19 patients.
Methods
Study design and participants
In this prospectively designed pre-post design cohort study, all adult COVID-19 patients admitted to the ICU of Radboud University Medical Center (Radboudumc, Nijmegen, The Netherlands) between March 2020 and April 2021 were screened for inclusion. Patients with comorbidities that might significantly influence the disease course and clinical outcomes (e.g. immunocompromised patients) were excluded. This study was carried out in accordance with the applicable rules concerning the review of research ethics committees and informed consent in the Netherlands. All patients or legal representatives were informed about the details of this cohort study and could decline to participate.
Included patients were divided into two cohorts: patients who were not treated with DEXA (pre-DEXA-cohort, March 2020–August 2020) and patients who received DEXA (6 mg/day, intravenously for 10 days) as part of standard COVID-19 care in accordance to the RECOVERY criteria [13] (DEXA-cohort, August 2020–April, 2021). A subgroup of the DEXA-cohort was also treated with the interleukin (IL)-6 receptor antagonist tocilizumab as part of standard COVID-19 care (single dose of 8 mg/kg, intravenously) [14]. Details on the sensitivity analyses performed in this subgroup are provided in the Additional file 1. Both cohorts were subdivided into groups of patients who were assessed to have developed PF while still in ICU and were treated with prednisone (start dose of 1 mg/kg twice daily, intravenously, PF-groups) and groups of patients who were not (no-PF-groups). In the absence of validated diagnostic criteria of PF, the diagnosis, and therefore the indication for prednisone treatment was at the discretion of the treatment team. All patients were discussed daily in a multidisciplinary meeting including over 15 medical experts, suspicion of PF and initiation of prednisone treatment was based on a combination of radiological findings, worsening ventilatory parameters (e.g. lower PaO2/FiO2 ratio, lower lung compliance and increased ventilatory ratio as a measure of impaired ventilation and increase in dead space ventilation), and an increase in PIIINP plasma levels that were measured three times per week. To analyze the kinetics of fibrosis biomarkers in the days prior to and following the day on which prednisone treatment for PF was initiated, serial data were aligned on the first day of prednisone treatment for PF (PF-day 0). For patients of the no-PF-groups, data were aligned on the median start day of late prednisone treatment in both cohorts separately to correct for time-dependent effects in this group [15, 16].
RNA sequencing
To explore underlying molecular mechanisms of PF development and treatments responses, we performed RNA sequencing on leukocytes isolated from a total of 52 PF- and no-PF-patients of the pre-DEXA-cohort. We used co-expression network analysis on these longitudinal RNA sequencing data using our established hCocena pipeline [17] to identify similarly regulated genes across samples and group these genes into modules. We applied this approach to samples obtained up to day 0 (when prednisone treatment was initiated in PF patients), to identify genes associated with the development of PF (pre-alignment day analysis). To assess the transcriptome response to treatment of PF with prednisone, we applied the same analysis pipeline to samples obtained from day 0 onwards (post-alignment day analysis). See Additional file 1 for a detailed description of RNA sequencing and analysis procedures.
Clinical data and biomarker measurements
See Additional file 1.
Statistical analysis
Differences in baseline characteristics and clinical outcomes between the PF- and no-PF-groups were analyzed using Mann–Whitney U and Fisher’s exact tests for continuous and categorical data, respectively. Differences in kinetics of serially measured data were analyzed using linear mixed effect model analysis on log-transformed data followed by post-hoc Sidak’s multiple comparisons tests. ICU-LOS and mortality were analyzed using log-rank tests during 60 days following ICU admission. A more detailed description of the statistical analysis is presented in Additional file 1.
Results
Patient characteristics
The pre-DEXA-cohort and the DEXA-cohort consisted of 67 and 124 patients, respectively (Fig. 1). Baseline characteristics of both cohorts are listed in Table 1. Prednisone treatment for PF was initiated on day 16 [12,13,14,15,16,17,18,19,20,21] and day 19 [14,15,16,17,18,19,20,21,22,23] following ICU admission in the pre-DEXA- and DEXA-cohorts, respectively (p = 0.11, Table 1). No relevant baseline demographic differences were present between the PF- and no-PF-groups within both cohorts. Furthermore, within the DEXA-cohort, no difference in the proportion of patients who were also treated with tocilizumab as standard COVID-19 care was present between the PF- and no-PF-groups (65% vs. 60%, p = 0.83, Table 1). Also, when comparing the PF-groups between both cohorts, no significant differences were present in patient characteristics and PF-free days from hospital admission onwards (Table 1).

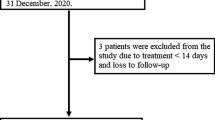
Patient flowchart. COVID-19 Coronavirus Disease 2019, DEXA dexamethasone, ICU intensive care unit, PF pulmonary fibroproliferation
Transcriptome analysis
In blood samples obtained up to day 0 (when prednisone treatment was initiated in PF patients), we identified nine co-expressed modules associated with the development of PF across a total of 3775 genes included in the analysis (pre-alignment day analysis, Fig. 2a, b). These modules are designated by colors gold to wheat (Fig. 2b). Based on linear regression analysis and a predefined set of rules (see Fig. 2a and Additional file 1), we focused on five modules associated with PF: seagreen, lightgreen, maroon, and wheat (upregulated in PF-patients) and turquoise (downregulated in PF-patients). Differential expressed genes over time were visualized by wave plots (Fig. 2c, wave plots of all modules provided in Additional file 1: Fig. S1) and heatmaps of the top 10 significant genes ranked by effect size (Fig. 2d, genes of all modules provided in Additional file 1: Fig. S1). Functional Enrichment Analysis (FEA) on these modules identified associated gene signatures with distinct functional characteristics related to fibrosis (Fig. 2e, all associated gene signatures provided in Additional file 1: Fig. S1). For instance, ‘inflammatory response’, ‘interferon (IFN)-γ response’, ‘IFN-α responses’, ‘response to virus’, ‘COVID-19’ and ‘influenza’ are enriched in the seagreen module, in keeping with the fact that inflammation is an important driver of fibrotic processes [18]. Hence, these data suggest a more pronounced response to (viral) infections, leading to more severe inflammation in COVID-19 patients who developed PF compared to COVID-19 patients who did not. In accordance, ‘regulation of interleukin-6 production’ and ‘myeloid cell differentiation’ were enriched in the lightgreen module and play key roles in both inflammation and development of PF [19,20,21]. The wheat module showed enrichment for ‘coagulation’ and ‘platelet activation’, and previous work has shown that the coagulation pathway is involved in fibroproliferative responses [22]. Accordingly, prevalence rates of pulmonary embolisms (PE) during stay in ICU and use of therapeutic low molecular weight heparins (LMWH) were compared between PF and no-PF groups. No differences in prevalence rates of PE were present between PF and no-PF groups (71% vs. 65%, respectively, p = 1.00). Therapeutic dosage of LMWH were administered in 75% of PF patients compared to 42% of no-PF patients (p = 0.04). ‘Neutrophil extracellular trap (NET) formation’ and ‘chromatin assembly’, were enriched in the maroon module. Interestingly, the release of NETs has been shown to play a role in the development of organ fibrosis [23] and their release is dependent on histone modification by peptidylarginine deiminase 4 (PADI4) [23, 24], which was one of the top 10 genes in the lightgreen module (Fig. 2d). The turquoise module which was downregulated in PF-patients, showed enrichment of ‘proteasomal protein catabolic process’ and ‘ubiquitin-mediated proteolysis’. Dysregulation of the ubiquitin–proteasome pathway is linked to multiple conditions, including fibrotic diseases [25], implicating that the ubiquitin–proteasome pathway is less functional in COVID-19 patients with PF. Finally, several specific genes which were distinctly upregulated in PF patients have previously been linked to fibrotic processes, including PDE4D [26], MMP8 [27], CRISP3 [28], and BCL2L15 [29] (all in maroon module, Fig. 2d). Additionally, to explore relationships between the gene modules and clinical outcomes of fibrosis, we performed gene set variation analysis on leukocyte gene expression data of a published cohort of patients with idiopathic pulmonary fibrosis (IPF, Fig. 2a) [30]. Four-year survival of IPF patients who showed enrichment of the genes in each module was compared to outcome of patients who exhibited no enrichment. Strikingly, survival of IPF patients who showed enrichment for genes in the maroon module was significantly worse (p = 0.019, Fig. 2f, survival plots for all modules provided in Additional file 1: Fig. S2).

Summary of bulk RNA-seq data pre-alignment day (day 0, the day prednisone treatment was initiated in patients with PF). a Depicts an overview of the complete cohort and general workflow per data set (dashed: pre-alignment day data set; solid: post-alignment day data set). b Shows the expression profile across the time points prior to the alignment day per module in a heatmap split by condition. The amount of samples per timepoint is displayed in brackets. Tiles are colored based on the group fold change (GFC) and modules are represented by their respective colors. Percentage of LMEM genes per module are depicted in a barplot (colored based on significance in the LMEM) and total numbers are shown as a ratio of LMEM genes to module genes. c Displays the mean expression per fibrosis-related module filtered by the LMEM genes over time prior to the alignment day. Lines and confidence intervals are colored according to the condition. d Depicts the mean expression of the top 10 LMEM genes per fibrosis-related module ordered by effect size for all conditions and time points prior to the alignment day. Modules are colored accordingly and effect size is indicated by the dot size. e Shows significant representative functional enrichment terms from GO and KEGG database as well as the hallmark gene set of the Molecular Signature Database per fibrosis-related module. Modules names are displayed on the x-axis and the respectively colored squares indicate the enrichment of a functional term in the module. f Displays the Kaplan–Meier plot of patients with idiopathic pulmonary fibrosis with and without enrichment of genes in the maroon module. Lines are colored based on the enrichment of the LMEM genes in the maroon module in the reference dataset using GSVA
The transcriptome response to treatment of PF with prednisone was assessed in blood samples obtained from day 0 onwards (post-alignment day analysis, see Fig. 2a). This analysis revealed nine co-expression modules (Fig. 3a). Applying the linear regression analysis and the predefined set of rules, led us to focus on two modules: slategray and wheat (Fig. 3b, c, wave plots and top 10 genes of all modules are provided in Additional file 1: Fig. S3). Genes in both modules were upregulated in the PF-group on day 0 and converged towards the no-PF-group afterwards, suggesting a treatment effect. Similar to the pre-alignment day analysis, both modules are enriched for multiple inflammatory and coagulation pathways (Fig. 3d, all associated gene signatures provided in Additional file 1: Fig. S3). Furthermore, the slategray module showed enrichment for ‘epithelial mesenchymal transition’, which was previously implicated in the development of organ fibrosis [31,32,33].

Summary of bulk RNA-seq data post-alignment day (day 0, the day prednisone treatment was initiated in patients with PF). a Shows the expression profile across the time points post-alignment day per module in a heatmap split by condition. The amount of samples per timepoint is displayed in brackets. Tiles are colored based on the group fold change (GFC) and modules are represented by their respective colors. Percentage of LMEM genes per module are depicted in a barplot (colored based on significance in the LMEM) and total numbers are shown as a ratio of LMEM genes to module genes. b Displays the mean expression per fibrosis-related module filtered by the LMEM genes over time post-alignment day. Lines and confidence intervals are colored according to the condition. c Depicts the mean expression of the top 10 LMEM genes per fibrosis-related module ordered by effect size for all conditions and time points post-alignment day. Modules are colored accordingly and effect size is indicated by the dot size. d Shows significant representative functional enrichment terms from GO and KEGG database as well as the hallmark gene set of the Molecular Signature Database per fibrosis-related module. Modules names are displayed on the x-axis and the respectively colored squares indicate the enrichment of a functional term in the module
There was no evidence that any of the module genes were regulated differentially between PF and no-PF groups by pro-fibrotic cytokines e.g. TGF-β.
Fibrosis biomarkers and ventilatory parameters
Peak PIIINP levels in the PF-groups were observed on day 3 following start of prednisone treatment and on the day prednisone treatment was initiated for PF (day 0) for the pre-DEXA- and DEXA-cohorts, respectively. PIIINP levels decreased significantly following prednisone treatment (Fig. 4ab). These kinetics were not observed in the no-PF-groups (Fig. 4ab). Unlike PIIINP, no significant between-group differences in circulating levels of HGF and MIP-3α within both cohorts were present (Additional file 1: Fig. S4). Following initiation of prednisone treatment, the dynamic lung compliance remained lower in the PF-groups of both cohorts compared with the no-PF-groups during the entire follow-up period (Fig. 4c, d). In the pre-DEXA-cohort, the ventilatory ratio decreased following initiation of prednisone treatment in the PF-group, whereas no relevant changes in ventilatory ratio were observed in the no-PF-group (Fig. 4e). In the DEXA-cohort, the ventilatory ratio of the PF-group remained higher compared to the no-PF-group on all ensuing timepoints following initiation of prednisone treatment (Fig. 4f). In the pre-DEXA-cohort, PaO2/FiO2 ratio of the PF-group remained lower compared to the no-PF-group for several days following start of prednisone treatment, while this was not the case in the DEXA-cohort (Fig. 4g, h). Kinetics and values on individual timepoints of all ventilatory parameters were similar between PF patients of both cohorts (Additional file 1: Fig. S5). So, overall, early DEXA treatment did not influence the subsequent response to steroid therapy in PF patients.

Circulating procollagen type III concentrations and clinical parameters. Differences between the pulmonary fibrosis (PF)- and no-PF-groups in kinetics of circulating procollagen type III (PIIINP) in a pre-DEXA-cohort and b DEXA-cohort, dynamic lung compliance in c pre-DEXA-cohort and d DEXA-cohort, ventilatory ratio in e pre-DEXA-cohort and f DEXA-cohort, and PaO2/FiO2 ratio in e pre-DEXA-cohort and f DEXA-cohort within 9 days (PIIINP) or 8 days (ventilatory parameters) prior to and 12 days following the alignment day (PF-day 0, start of prednisone treatment in the PF-groups). P-values on the left and the right of each panel reflect between-group differences over time for the days prior to and following PF-day 0, respectively, and were calculated using linear mixed models analysis (time * group interaction factor). Data presented as geometric mean with 95% confidence intervals. *p-value < 0.05 on the corresponding timepoint, calculated using Sidak’s post-hoc multiple comparisons tests
Clinical outcomes
PF incidence was 28% and 25% in the pre-DEXA- and DEXA-cohorts, respectively (p = 0.61). Time on ventilator, LOS in ICU and mortality were higher in the PF-groups compared to the no-PF-groups within both cohorts (Table 1, Fig. 5). Furthermore, within both cohorts, PF-patients who survived their ICU stay had a prolonged time on mechanical ventilation and ICU stay compared to no-PF-patients who survived. None of the clinical outcomes differed between the PF-groups of both cohorts (Table 1), again indicating no influence of early DEXA treatment on these clinical response to steroid treatment for PF. When dividing the DEXA-cohort into subgroups of patients who were also treated with tocilizumab and those who were not, similar differences in clinical outcomes between the PF- and no-PF-groups were observed as in the main analysis (Additional file 1: Table S1). Furthermore, no differences in clinical outcomes were present between PF-patients who were co-treated with tocilizumab and PF-patients who were not (Additional file 1: Table S1).

Clinical outcomes. Differences between the pulmonary fibrosis (PF)- and no PF-groups in length of stay (LOS) in the intensive care unit (ICU) in a the pre-DEXA-cohort and b the DEXA-cohort, and 60-day hospital mortality in c the pre-DEXA-cohort and d the DEXA-cohort. Kaplan–Meier curves are depicted and p-values were calculated using log-rank tests. For analysis of 60-day hospital mortality, patients who were discharged alive from the hospital or were still in the ICU or hospital on day 60 were censored at day 60. Numbers at risk are shown below graphs
Discussion
Our study reveals that several genes and signaling pathways that were previously linked to organ fibrosis are upregulated in critically ill COVID-19 patients who develop PF, including inflammatory processes, coagulation, and NET-related pathways. Furthermore, we demonstrate that some of the identified pathways are associated with worse long-term outcomes of fibrotic diseases. Following initiation of steroid treatment for PF, multiple upregulated pathways in the PF-group converged towards expression levels observed in the no-PF-group. Likewise, circulating PIIINP levels reverted to concentrations similar to those observed in the no-PF-group following treatment of PF with steroids. Whereas several clinical ventilatory parameters also stabilized or improved after treatment, these largely remained worse in PF-patients compared to no-PF-patients. Importantly, this treatment response was not influenced by early dexamethasone treatment. Finally, PF was associated with a prolonged length of stay in the ICU and higher mortality rates, which was also not influenced by early dexamethasone treatment or co-treatment with tocilizumab.
Up to now, multiple studies have described long-term symptoms of COVID-19 in both ICU and non-ICU COVID-19 patients, including pulmonary sequalae [34,35,36,37]. The exact underlying mechanisms for these long-lasting symptoms are still unclear, but the development of PF likely plays a role. Therefore, and because of its high mortality, it is of paramount importance to detect and treat PF at an early stage. In non-COVID-19 ARDS patients, it was shown that corticosteroid treatment is effective in shortening ICU-LOS and reducing mortality rates [11]. Accordingly, we hypothesized that early dexamethasone treatment would result in a lower incidence or less severe course of excessive PF and subsequent more pronounced improvement of clinical pulmonary outcomes in patients of our DEXA-cohort compared to the pre-DEXA-cohort. In contrast, we did not observe differences in incidence rates or clinical outcomes between both cohorts.
Although it did not reach statistical significance, time from ICU admission until the initiation of prednisone to treat PF was 3 days later in the DEXA-cohort. One may argue that prolongation of the early dexamethasone treatment as standard care in this subgroup of critically ill COVID-19 patients could further delay or even prevent the development of PF. Additionally, the dose of dexamethasone used as standard treatment for COVID-19 is considerably lower than the equivalent corticosteroid dose of prednisone used for the treatment of PF. For example, a patient with PF of 80 kg would be treated with 160 mg prednisone daily, approximately equivalent to 24 mg dexamethasone [38], and thus several times higher than the 6 mg dexamethasone dose used as standard treatment for COVID-19. Therefore, we cannot exclude that prolongation of treatment, or increasing the dosage of early dexamethasone may mitigate the development of PF and improve clinical outcomes in critically ill COVID-19 patients. Of interest, a recently published study compared the effects of daily administration of 6 mg and 12 mg dexamethasone for 10 days in 982 severely ill COVID-19 patients and showed better clinical outcomes in the 12 mg group, while the incidence of serious adverse effects was similar [39]. To investigate the effects of early dexamethasone treatment on incidence rates and mortality of all hospitalized COVID-19 patients and the effects of prolongation/intensification of dexamethasone treatment on the development of PF in critically ill COVID-19 patients, randomized controlled trials should be performed.
Our longitudinal transcriptome analysis provided clues for novel therapeutic targets for prevention or treatment of PF in critically ill COVID-19 patients. One of the most strongly upregulated genes in PF-patients, MMP8, is related to bleomycin-induced fibrosis in mice [27, 40], that treatment with MMP8 inhibitors may be beneficial. Similar, inhibition of PDE4, also markedly upregulated in PF patients, prevented PF in bleomycin-treated mice [26]. Interestingly, the PDE4 inhibitor roflumilast is already licensed for the treatment of severe COPD and asthma [41, 42].
This study has several limitations. First, early dexamethasone treatment was not randomized. As a consequence, bias related to the initial response to dexamethasone is likely present, especially because this treatment is often started on the ward. Therefore, it is possible that several patients of the pre-DEXA-cohort would not have required ICU admission if they would have received dexamethasone on the ward. On the other hand, our data are as observed in current clinical practice. Second, later on, patients were treated with prednisone when PF was identified based on radiological findings, worsening of ventilatory parameters and elevated circulating PIIINP levels which were available to the treating physicians. Ideally, PF should be diagnosed based on high PIIINP levels in BAL fluids and typical high-resolution computed tomography (HRCT) images [4, 8]. However, during the COVID-19 pandemic, it was not feasible to perform repeated BALs and HRCTs, and circulating PIIINP levels have shown promise for evaluation of disease progression and treatment efficacy in non-COVID-19 ARDS patients [12, 43]. Also, the differences in transcriptome profiles between PF- and no-PF-patients support the PF diagnosis, and the changes in PIIINP kinetics, pulmonary parameters, and gene expression patterns following prednisone treatment illustrate therapeutic efficacy. Third, the transcriptome analyses were performed on total leukocytes. With this approach, changes in blood count differential (i.e. the percentages of e.g. monocytes, lymphocytes, and neutrophils) influence gene expression levels. Unfortunately, we do not have data available on blood count differentials to assess the magnitude of this effect in our analyses. Fourth, since the relatively small sample size, our study was possibly underpowered to draw conclusions on the effects of early dexamethasone treatment on incidence and clinical outcomes of COVID-19 patients with PF. However, we did not find any (non-significant) indications to a major influence of early dexamethasone treatment on these outcomes. Last, the observational design of our study could have introduced confounding because of the rapidly increasing knowledge of the disease and the so-called learning curve during the pandemic. Since we compared data of two cohorts of COVID-19 patients who were admitted to the ICU during different periods in time, the possibility that the introduction of dexamethasone and tocilizumab as standard treatment in COVID-19 was not the only difference in treatment between both cohorts has to be acknowledged. For example, differences in virulence of the dominant SARS-CoV-2 strain during each period of time might also be of influence on our study outcomes. Preferably, a RCT in hospitalized COVID-19 patients should be performed to more accurately determine the effects of early dexamethasone treatment on the incidence, clinical course and outcomes of PF, although this would now raise ethical dilemmas.
Conclusions
In critically ill COVID-19 patients who develop PF, coagulation, inflammation and NET-related pathways are upregulated, ICU-LOS is prolonged and mortality is higher. This study indicates that early dexamethasone treatment neither influences the incidence or clinical course of PF, nor the outcomes of this subgroup of critically ill COVID-19 patients. Steroid treatment normalized PF-related RNA profiles and PIIINP levels, while clinical parameters stabilized, but remained aberrant compared to critically ill COVID-19 patients without signs of PF.
Availability of data and materials
The RNA sequencing data are made available at the European Genome-Phenome Archive (EGA) under accession numbers EGAS00001005735 and EGAS00001006407, which is hosted by the EBI and the CRG. The R package to bin and align longitudinal clinical data is available here: https://doi.org/10.5281/zenodo.6594554. Other datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Burnham EL, Janssen WJ, Riches DWH, Moss M, Downey GP. The fibroproliferative response in acute respiratory distress syndrome: mechanisms and clinical significance. Eur Respir J. 2014;43:276–85.
Cabrera-Benitez NE, Laffey JG, Parotto M, Spieth PM, Villar J, Zhang H, Slutsky AS. Mechanical ventilation-associated lung fibrosis in acute respiratory distress syndrome: a significant contributor to poor outcome. Anesthesiology. 2014;121:189–98.
Martin C, Papazian L, Payan M-J, Saux P, Gouin F. Pulmonary fibrosis correlates with outcome in adult respiratory distress syndrome: a study in mechanically ventilated patients. Chest. 1995;107:196–200.
Ichikado K, Muranaka H, Gushima Y, Kotani T, Nader HM, Fujimoto K, Johkoh T, Iwamoto N, Kawamura K, Nagano J, et al. Fibroproliferative changes on high-resolution CT in the acute respiratory distress syndrome predict mortality and ventilator dependency: a prospective observational cohort study. BMJ Open. 2012;2: e000545.
Gao J, Chu W, Duan J, Li J, Ma W, Hu C, Yao M, Xing L, Yang Y. Six-month outcomes of post-ARDS pulmonary fibrosis in patients with H1N1 pneumonia. Front Mol Biosci. 2021;8: 640763.
Herridge MS, Cheung AM, Tansey CM, Matte-Martyn A, Diaz-Granados N, Al-Saidi F, Cooper AB, Guest CB, Mazer CD, Mehta S, et al. One-year outcomes in survivors of the acute respiratory distress syndrome. N Engl J Med. 2003;348:683–93.
Masclans JR, Roca O, Muñoz X, Pallisa E, Torres F, Rello J, Morell F. Quality of life, pulmonary function, and tomographic scan abnormalities after ARDS. Chest. 2011;139:1340–6.
Forel JM, Guervilly C, Hraiech S, Voillet F, Thomas G, Somma C, Secq V, Farnarier C, Payan MJ, Donati SY, et al. Type III procollagen is a reliable marker of ARDS-associated lung fibroproliferation. Intensive Care Med. 2015;41:1–11.
Yamanouchi H, Fujita J, Yoshinouchi T, Hojo S, Kamei T, Yamadori I, Ohtsuki Y, Ueda N, Takahara J. Measurement of hepatocyte growth factor in serum and bronchoalveolar lavage fluid in patients with pulmonary fibrosis. Respir Med. 1998;92:273–8.
Chu X, Jin Q, Chen H, Wood GC, Petrick A, Strodel W, Gabrielsen J, Benotti P, Mirshahi T, Carey DJ, et al. CCL20 is up-regulated in non-alcoholic fatty liver disease fibrosis and is produced by hepatic stellate cells in response to fatty acid loading. J Transl Med. 2018;16:108.
Meduri GU, Golden E, Freire AX, Taylor E, Zaman M, Carson SJ, Gibson M, Umberger R. Methylprednisolone infusion in early severe ARDS: results of a randomized controlled trial. Chest. 2007;131:954–63.
Meduri GU, Tolley EA, Chinn A, Stentz F, Postlethwaite A. Procollagen types I and III aminoterminal propeptide levels during acute respiratory distress syndrome and in response to methylprednisolone treatment. Am J Respir Crit Care Med. 1998;158:1432–41.
Horby P, Lim WS, Emberson JR, Mafham M, Bell JL, Linsell L, Staplin N, Brightling C, Ustianowski A, Elmahi E, et al. Dexamethasone in hospitalized patients with covid-19. N Engl J Med. 2021;384:693–704.
Gordon AC, Mouncey PR, Al-Beidh F, Rowan KM, Nichol AD, Arabi YM, Annane D, Beane A, van Bentum-Puijk W, Berry LR, et al. Interleukin-6 receptor antagonists in critically ill patients with covid-19. N Engl J Med. 2021;384:1491–502.
Kooistra EJ, Waalders NJB, Grondman I, Janssen NAF, de Nooijer AH, Netea MG, van de Veerdonk FL, Ewalds E, van der Hoeven JG, Kox M, Pickkers P. Anakinra treatment in critically ill COVID-19 patients: a prospective cohort study. Crit Care. 2020;24:688.
Kooistra EJ, van Berkel M, van Kempen NF, van Latum CRM, Bruse N, Frenzel T, van den Berg MJW, Schouten JA, Kox M, Pickkers P. Dexamethasone and tocilizumab treatment considerably reduces the value of C-reactive protein and procalcitonin to detect secondary bacterial infections in COVID-19 patients. Crit Care. 2021;25:281.
Aschenbrenner AC, Mouktaroudi M, Krämer B, Oestreich M, Antonakos N, Nuesch-Germano M, Gkizeli K, Bonaguro L, Reusch N, Baßler K, et al. Disease severity-specific neutrophil signatures in blood transcriptomes stratify COVID-19 patients. Genome Med. 2021;13:7.
Ueha S, Shand F, Matsushima K. Cellular and molecular mechanisms of chronic inflammation-associated organ fibrosis. Front Immunol. 2012;3:71.
Saito F, Tasaka S, Inoue K-I, Miyamoto K, Nakano Y, Ogawa Y, Yamada W, Shiraishi Y, Hasegawa N, Fujishima S, et al. Role of interleukin-6 in bleomycin-induced lung inflammatory changes in mice. Am J Respir Cell Mol Biol. 2008;38:566–71.
Le TT, Karmouty-Quintana H, Melicoff E, Le TT, Weng T, Chen NY, Pedroza M, Zhou Y, Davies J, Philip K, et al. Blockade of IL-6 trans signaling attenuates pulmonary fibrosis. J Immunol. 2014;193:3755–68.
Florez-Sampedro L, Song S, Melgert BN. The diversity of myeloid immune cells shaping wound repair and fibrosis in the lung. Regeneration (Oxford, England). 2018;5:3–25.
Chambers RC, Scotton CJ. Coagulation cascade proteinases in lung injury and fibrosis. Proc Am Thorac Soc. 2012;9:96–101.
Chrysanthopoulou A, Mitroulis I, Apostolidou E, Arelaki S, Mikroulis D, Konstantinidis T, Sivridis E, Koffa M, Giatromanolaki A, Boumpas DT, et al. Neutrophil extracellular traps promote differentiation and function of fibroblasts. J Pathol. 2014;233:294–307.
Suzuki M, Ikari J, Anazawa R, Tanaka N, Katsumata Y, Shimada A, Suzuki E, Tatsumi K. PAD4 deficiency improves bleomycin-induced neutrophil extracellular traps and fibrosis in mouse lung. Am J Respir Cell Mol Biol. 2020;63:806–18.
Inui N, Sakai S, Kitagawa M. Molecular pathogenesis of pulmonary fibrosis, with focus on pathways related to TGF-β and the ubiquitin-proteasome pathway. Int J Mol Sci. 2021;22:6107.
Udalov S, Dumitrascu R, Pullamsetti SS, Al-tamari HM, Weissmann N, Ghofrani HA, Guenther A, Voswinckel R, Seeger W, Grimminger F, Schermuly RT. Effects of phosphodiesterase 4 inhibition on bleomycin-induced pulmonary fibrosis in mice. BMC Pulm Med. 2010;10:26.
Craig VJ, Quintero PA, Fyfe SE, Patel AS, Knolle MD, Kobzik L, Owen CA. Profibrotic activities for matrix metalloproteinase-8 during bleomycin-mediated lung injury. J Immunol. 2013;190:4283–96.
Mathai SK, Cardwell J, Metzger F, Powers J, Walts AD, Kropski JA, Eickelberg O, Hauck SM, Yang IV, Schwartz DA. Preclinical pulmonary fibrosis circulating protein biomarkers. Am J Respir Crit Care Med. 2020;202:1720–4.
Safaeian L, Abed A, Vaseghi G. The role of Bcl-2 family proteins in pulmonary fibrosis. Eur J Pharmacol. 2014;741:281–9.
Herazo-Maya Jose D, Noth I, Duncan Steven R, Kim S, Ma S-F, Tseng George C, Feingold E, Juan-Guardela Brenda M, Richards Thomas J, Lussier Y, et al. Peripheral blood mononuclear cell gene expression profiles predict poor outcome in idiopathic pulmonary fibrosis. Sci Transl Med. 2013;5:205ra136.
Kalluri R, Neilson EG. Epithelial–mesenchymal transition and its implications for fibrosis. J Clin Invest. 2003;112:1776–84.
Stone RC, Pastar I, Ojeh N, Chen V, Liu S, Garzon KI, Tomic-Canic M. Epithelial–mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res. 2016;365:495–506.
Guarino M, Tosoni A, Nebuloni M. Direct contribution of epithelium to organ fibrosis: epithelial–mesenchymal transition. Hum Pathol. 2009;40:1365–76.
Gamberini L, Mazzoli CA, Prediletto I, Sintonen H, Scaramuzzo G, Allegri D, Colombo D, Tonetti T, Zani G, Capozzi C, et al. Health-related quality of life profiles, trajectories, persistent symptoms and pulmonary function one year after ICU discharge in invasively ventilated COVID-19 patients, a prospective follow-up study. Respir Med. 2021;189: 106665.
Morin L, Savale L, Pham T, Colle R, Figueiredo S, Harrois A, Gasnier M, Lecoq AL, Meyrignac O, Noel N, et al. Four-month clinical status of a cohort of patients after hospitalization for COVID-19. JAMA. 2021;325:1525–34.
Latronico N, Peli E, Calza S, Rodella F, Novelli MP, Cella A, Marshall J, Needham DM, Rasulo FA, Piva S. Physical, cognitive and mental health outcomes in 1-year survivors of COVID-19-associated ARDS. Thorax. 2021;77(3):300–3.
Heesakkers H, van der Hoeven JG, Corsten S, Janssen I, Ewalds E, Simons KS, Westerhof B, Rettig TCD, Jacobs C, van Santen S, et al. Clinical outcomes among patients with 1-year survival following intensive care unit treatment for COVID-19. JAMA. 2022;327(6):559–65.
Schimmer BP, Funder JW. ACTH, adrenal steroids, and pharmacology of the adrenal cortex. In: Brunton LL, Chabner BA, Knollmann BC, editors. Goodman & Gilman’s: the pharmacological basis of therapeutics. 12th ed. New York: McGraw-Hill Education; 2015.
Granholm A, Munch MW, Myatra SN, Vijayaraghavan BKT, Cronhjort M, Wahlin RR, Jakob SM, Cioccari L, Kjær MN, Vesterlund GK, et al. Dexamethasone 12 mg versus 6 mg for patients with COVID-19 and severe hypoxaemia: a pre-planned, secondary Bayesian analysis of the COVID STEROID 2 trial. Intensive Care Med. 2021;48:1–11.
García-Prieto E, González-López A, Cabrera S, Astudillo A, Gutiérrez-Fernández A, Fanjul-Fernandez M, Batalla-Solís E, Puente XS, Fueyo A, López-Otín C, Albaiceta GM. Resistance to bleomycin-induced lung fibrosis in MMP-8 deficient mice is mediated by interleukin-10. PLoS ONE. 2010;5: e13242.
Pagès L, Gavaldà A, Lehner MD. PDE4 inhibitors: a review of current developments (2005–2009). Expert Opin Ther Pat. 2009;19:1501–19.
Li H, Zuo J, Tang W. Phosphodiesterase-4 inhibitors for the treatment of inflammatory diseases. Front Pharmacol. 2018;9:1048.
Steinberg KP, Hudson LD, Goodman RB, Hough CL, Lanken PN, Hyzy R, Thompson BT, Ancukiewicz M, National Heart Lung, Blood Institute Acute Respiratory Distress Syndrome Clinical Trials Network. Efficacy and safety of corticosteroids for persistent acute respiratory distress syndrome. N Engl J Med. 2006;354:1671–84.
Acknowledgements
EK takes responsibility for (is the guarantor of) the content of the manuscript, including the data and analysis. Dr. Kooistra was funded by a Radboudumc Coronavirus Disease 2019 grant (Radboudfonds). The authors have no financial or nonfinancial disclosures. The authors thank Denise Waanders, Nicole Waalders, Wout Claassen, Hidde Heesakkers, Tirsa van Schaik, Leonie Buijsse, Maud Lamberts, Jesper Walburgh Schmidt, Marc Pickkers, Celine van Latum, Noortje van Kempen, Aron Jansen, Harmke Duindam and Dirk van Lier (all affiliated to the Department of Intensive Care, Radboudumc, Nijmegen, The Netherlands) for their contribution to data and sample collection. The authors also thank Doorlène van Tienoven for her analytical expertise in PIIINP measurements, and Niklas Bruse for developing a R package to bin and align longitudinal clinical data (https://doi.org/10.5281/zenodo.6594554).
Funding
EK was funded by a Radboudumc Coronavirus Disease 2019 grant (Radboudfonds).
Author information
Authors and Affiliations
Contributions
EK, AvH, MK an PP designed the study. EK and JvdV were responsible for data collection from the electronical patient files and stored data in the database. EK was responsible for sample collection. AvH, RS and JG performed laboratory analyses of the clinical data. KD, MNG, AA, TU and JS were responsible for the RNA sequencing analyses. KM designed the plots of RNA sequencing analyses. MvdB and JvdH were involved in the study as clinical experts. EK, AvH, MK and PP analyzed and interpreted the clinical data. EK, KD, AA, TU, JS, MK and PP interpreted the RNAseq data. EK drafted the manuscript. All authors revised and approved the final manuscript for publication.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was carried out in accordance with the applicable rules concerning the review of research ethics committees and informed consent in the Netherlands. All patients or legal representatives were informed about the details of this cohort study and could decline to participate.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Additional Methods, Additional Table S1 and Additional Figures S1–S5.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kooistra, E.J., Dahm, K., van Herwaarden, A.E. et al. Molecular mechanisms and treatment responses of pulmonary fibrosis in severe COVID-19. Respir Res 24, 196 (2023). https://doi.org/10.1186/s12931-023-02496-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12931-023-02496-1