
Abstract
The aim of this research was to estimate the immunopotentiation effect of brown algae Padina boergesenii water extract on Nile tilapia, Oreochromis niloticus through resistance to Pseudomonas putida infection. Gas Chromatography Mass Spectrometry was utilized to characterize the seaweed phytoconstituents. One hundred and twenty-six fish were divided in triplicates into two equal groups corresponding to two diet variants that used to feed Nile tilapia for 20 successive days: a basal (control), and P. boergesenii water extract supplemented group. Fish samples were collected at 10-days intervals throughout the experiment. Serum biochemical constituents, total antioxidant capacity (TAC), and some immune related genes expression of the spleen and intestinal tissues of experimental fish were studied, as well as histological examination of fish immune tissues. Moreover, following 20 days of feeding, the susceptibility of Nile tilapia to P. putida infection was evaluated to assess the protective effect of the used extract. The findings indicated that the studied parameters were significantly increased, and the best immune response profiles were observed in fish fed P. boergesenii water extract for 20 successive days. A bacterial challenge experiment using P. putida resulted in higher survival within the supplemented fish group than the control. Thus, the lowered post-challenge mortality of the fish may be related to the protection provided by the stimulation of the innate immune system, reduced oxidative stress by higher activity of TAC, and elevated levels of expression of iterleukin-1beta (IL-1β), beta-defensin (β-defensin), and natural killer-lysin (NKl). Moreover, the constituents of the extract used showed potential protective activity for histological features of the supplemented fish group when compared to the control. Collectively, this study presents a great insight on the protective role of P. boergesenii water extract as an additive in Nile tilapia feed which suggests its potential for improving the immune response against P. putida infection.
Similar content being viewed by others

Introduction
Nile tilapia, O. niloticus, aquaculture has experienced a substantial expansion in Egypt and other tropical nations [1]. Nevertheless, the extensive growth of their population is confronted with various infections and specific strategies for handling them that have the potential to harm the well-being of both humans and animals [2]. Disease outbreaks significantly hinder the long-term viability of Nile tilapia aquaculture production. Pseudomonas and other bacterial septicemia provide the greatest risk to Nile tilapia in Egypt [3]. Previous research has indicated that P. putida is a significant bacterial pathogen that impacts a range of fish species [4, 5], resulting in elevated mortality rates and substantial economic loss [6]. P. putida was recorded from Nile tilapia in Egypt and resulted in exophthalmia, ascites, and ulcers on the fish body [5, 7].
Different chemotherapeutic agents can be employed to prevent various illnesses in aquaculture. Nevertheless, their utilization is accompanied by antibiotic resistance against bacteria [8], antibiotic residues in tissues [9], and pollution of the aquatic environment [10]. Therefore, there is a growing demand for new approaches to manage bacterial infections [11] and there is an increasing interest in using creative management approaches to enhance aquatic welfare and ensure the construction of safe food [12]. These alternate measures involve the utilization of natural bioactive substances that activate the immune system of fish.
Seaweeds are inexpensive and readily accessible sources of bioactive chemicals that can be utilized as immunostimulants in aquaculture [13]. The seaweed P. boergesenii is a type of brown algae, that is frequently encountered in coastal areas worldwide. P. boergesenii has lately gained considerable attention in aquaculture due to its diverse range of active principles that possess highly powerful immunostimulant, antioxidant, and antibacterial characteristics [14, 15]. A recent study indicated that consuming P. boergesenii in fish diet may have the ability to combat Pseudomonas aeruginosa infection [16].
Serum biochemical and innate immune markers are employed to assess the impact of feed additives on fish [17, 18]. Recently, there has been a growing focus on studying the alterations in the expression of genes relevant to the immune system. IL-1β plays a crucial role in the inflammatory response in fish [19] and is a key component of the innate immune mechanism [20]. It can activate lymphocytes and other cytokines, which in turn can stimulate phagocytic cells.
Antimicrobial peptides are constituents of the fish’s innate immune system which offer defense against bacteria, and other pathogens [21, 22]. Many AMPs inhibit bacterial colonization by employing either a lytic or ionophoric mechanism [23]. β-defensin and NKL are two prominent and widely recognized groups of antimicrobial peptides (AMPs). β-defensin exhibits potent antibacterial activity against infections that affect fish [24, 25]. Furthermore, the transcriptome expression of this antimicrobial peptide (AMP) was found to be elevated in fish that were given immunostimulants [26]. NKL is synthesized by cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells and exhibits a wide spectrum of activity against many microbial infections [27]. Hence, the target of this work was to investigate the immunomodulatory impacts of P. boergesenii water extract when added to Nile tilapia feed. Serum biochemical indicators, some immune gene expression response, and the histological architecture of the intestine, kidney, liver, and spleen will be evaluated in this study. Furthermore, a challenge experiment will be carried out to evaluate the disease resistance capabilities of Nile tilapia against P. putida infection.
Materials and methods
Collection and identification of brown algae
Samples from seaweed P. boergesenii, a species of brown algae, was collected from the coastal region of the Red Sea near Hurghada city during the summer of 2022. The National Institute of Oceanography and Fisheries (NIOF) gathered and identified the seaweed material. Following the collection process, the algae were promptly rinsed with seawater to eliminate any attached substances and contaminants, such as sand particles and epiphytes, and then sterilized using distilled water. According to Allender and Kraft [28] macroalgae species were taxonomically identified based on their morphology. Samples were left to dry at room temperature, crushed into fine powder, and then sieved following the method of Cho et al. and Gonzalez et al. [29, 30].
Preparation of P. boergesenii water extract
The brown alga, P. boergesenii (Phaeophyceae, Dictyotaceae) watery extract was prepared according to Ghaednia et al. [31]. Briefly, 20 g of powdered sea algae were dissolved in 300 ml of distilled water in a round bottom flask and boiled for about 3 h at 100 °C, then filtrated using filter paper, and the filtered extract was lyophilized using a rotary evaporator till obtain solid watery extract.
Gas chromatography-mass spectrometry (GC/MS) analysis of P. Boergesenii
The P. boergesenii extract underwent analysis using GC-MS at the National Research Center in Dokki, Cairo, Egypt. The specimen was introduced into an HP-5 column (30 m × 0.25µM film thickness) for GC–MS analysis. The PerkinElmer Clarus 580/560 S model system was utilized to convey the gas at a flux rate of 0.8 mL/min. The temperature of the gas chromatography (GC) oven was set to increase at a rate of 2 °C/min., starting from 60 °C and reaching 250 °C. The total ion current is determined by calculating the relative area values of the volatile composition as a proportion of the total.
Diets preparation
The basal diet used in this experiment was purchased from Skretting Company of Animal Nutrition, Egypt, and contained 30% crude protein, 9.5% ash, 6% crude lipid, and 5.22% crude fiber. The control diet, which served as the basal diet, was not supplemented with P. boergesenii water extract. Feeding rate used in this study was chosen according to the earlier finding in Cirrhinus mrigala [16] to detect the most suitable concentration of the P. boergesenii water extract powder. Hence, the supplemented diet was prepared by thoroughly mixing the basal diet with 4.5% concentration of P. boergesenii water extract powder until stiff dough was obtained, and water was used as the binding agent. The dough was prepared into pellets with the help of a hand pelletizer and then cut into small-sized pieces. The pellets were left to dry at room temperature and were preserved in plastic containers at -20oC until use.
Fish samples and experimental design
The methodology and fish experiments undertaken in this study were approved by the Ethical Committee of the Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt. These experiments adhered to the regulations set by the OIE guidelines for the use of animals in research, under the approval No. 06/2024/0179.
A total of 150 live, healthy, cultured Nile tilapia, with an initial average body weight of 65 ± 6 g, were collected from the Nag Hamady fish hatchery in Qena Governorate. They were transported in oxygenated tanks to the Animal Experiment Lab, Fish Diseases Division, Faculty of Veterinary Medicine, South Valley University.
The fish were acclimated to laboratory conditions for a period of 14 days in fully equipped 500 L fiberglass tanks. During this time, they were fed commercial pellets without any additional substances, twice a day, at a quantity equivalent to 3% of their body weight. The tanks were provided with dechlorinated tape water containing a salinity of 0.2 parts per thousand (ppt), and the duration of light and darkness was regulated to 12 h each. A total of ten Nile tilapia specimens were randomly captured and subsequently tested negative for P. putida using specific primer (Table 1) in polymerase chain reaction which give 380 bp (bp).
Feeding trials for this study were set up and continued for 20 days in three replicates, where acclimated Nile tilapia (n = 126) was randomly divided into 2 equal groups (control and supplemented) that represent the two nutritional experiments (corresponding to the two diet variants). Each group was divided into two subgroups, including 18 (n = 6 per replicate) and 45 (n = 5 per replicate) fish, used for sampling (sampling tanks) to evaluate the serum parameters, histological examination, and gene expression analysis of fish, and for the challenge experiment (challenge tanks), respectively.
During the feeding period, the experimental diets were given to the fish twice a day, at a dosage of 3% of their body weight. With dissolved oxygen at 7 ± 0.5 mg /L, pH at 7 ± 0.4, and water temperature at 25 ± 0.5oC, the tank water was kept in an ideal condition.
Blood and tissue sampling
Blood samples were obtained from the experimental fish (n = 2/group/replicate) at the 10th and 20th days of the feeding period. The fish were randomly caught and anesthetized with 0.05 mL/L clove oil [32]. Blood was collected from the caudal vein by using blood collection tubes. Samples were left for 3–4 h at room temperature to coagulate, then centrifuged at 4000 rpm for 25 min. The serum was collected and stored at − 20oC until used for further analysis.
Fish were euthanized with 0.5 mL/L of clove oil after blood sampling, and then the spleen and intestine were immediately collected (n = 1/group/replicate) and placed in RNAlater (Qiagen) at 4 °C overnight and then stored at − 20 °C until the further analysis.
Analysis of serum biochemical indices
The serum analysis was conducted utilizing commercially available test kits as per the manufacturer’s recommended procedures. The parameters examined were total protein and albumin (Biomed Diagnostics, Badr city, Egypt). The results were determined through colorimetric analysis in a T80 Spectrophotometer (PG Instrument, Leicestershire, UK). To get the globulin level (g/dL), the albumin value is subtracted from the total protein value using a mathematical calculation [33]. The TAC level was assessed after the method of Koracevic et al. [34], utilizing commercially accessible test kits from Bio diagnostic, located in Dokki, Egypt.
Challenge experiment with P. Putida
The pathogenic P. putida strain used for the challenge study was isolated from Nile tilapia, O niloticus, showing typical signs of septicemia during an outbreak in a local commercial farm. The strain was identified by various biochemical characters as pseudomonas species and confirmed molecularly as P. putida using 16s rDNA gene sequencing (GenBank accession no. OM048106, 838 bp). The strain isolated was identified and preserved at -80 °C.
Pseudomonas putida colonies were recovered by cultivating them on pseudomonas agar base and incubating them at a temperature of 28oC for 24 h. Subsequently, one colony was selected and cultured into 100 mL of tryptic soy broth and then incubated at a temperature of 28oC for one night. The bacterial suspension utilized for infecting Nile tilapia was quantified by assessing the optical density by spectrophotometric absorbance at a wavelength of 600 nm, as well as by determining the colony-forming unit (cfu) counts using a standard plate-count method [35].
To restore virulence, the P. putida was administered 3 passages to Nile tilapia via intra-peritoneal injection (500 µL broth culture) before to the actual challenge experiment. Colony morphology and biochemical profile allowed for the probable identification of P. putida after its isolation from the kidneys of injected Nile tilapia. Molecular analysis with primers specific to P. putida validated the identification (Table 1).
In the challenge subgroups (control and supplemented), 3 tanks (n = 15) were set for clinical signs and mortality records (mortality tanks), another 3 tanks (n = 15) were used for post-challenge gene expression analysis and histological examination (challenge sampling tanks) and additional 3 tanks (n = 15) were assigned as bacterial challenge negative control group.
After 20 days of feeding, fish of the challenge tanks (mortality and challenge sampling tanks) from both feeding groups (n = 30/group) were removed from their tanks using a net, anesthetized, and subjected to a 500 µL intraperitoneal injection of P. putida at a concentration of 1 × 106 cfu/mL. The negative challenge control group (n = 15) received an intraperitoneal injection of a 500 µL sterile physiological saline solution. The mortality rate was observed and documented daily for a period of 14 days post-challenge. In addition, samples of moribund and recently dead fish were taken from the kidney to verify P. putida presence as the reason of the documented mortality and observed clinical symptoms. The results were stated as percentage of mortality. After 2 days of post-challenge, 3 fish from each feeding group (1 fish / sampling tank) were randomly chosen and euthanized. The spleen and gut were sampled to obtain total RNA.
Gene expression analysis
Total RNA extraction and cDNA synthesis
The tissue was disrupted and homogenized, and total RNA was extracted from the spleen and intestinal tissues using the RNeasy® Mini kit (Qiagen, Germany) according to the manufacturer’s instructions. The RNA was eluted from the column and its purity and concentration were assessed using a nano-photometer spectrophotometer (NanoDrop™ LITE Spectrophotometer, Thermo Scientific, USA). For cDNA synthesis, 1 µg of total RNA from each sample (with a purity of 2.1 ± 0.2) was reverse transcribed using the RevertAid first strand cDNA synthesis kit (Thermo Scientific, USA) according to the manufacturer’s instructions. Subsequently, the first cDNA products were preserved at a temperature of -20 °C until used for the amplification in the quantitative real-time PCR (qRT-PCR).
Target genes and primers
To analyze the expression of our target immune genes, we evaluated the expressions of IL-1β, β-defensin, and NKL genes using qRT-PCR. The primer sequences utilized in the gene expression study are provided in Table 1. The relative expression was normalized using the Nile tilapia beta-actin (β-actin) and elongation factor 1-alpha (EF1α) genes as reference genes.
Expression of the target genes and data visualization
The HERAPLUS SYBR® Green Master Mix (Willowfort, England) was utilized to perform qRT-PCR, following the instructions provided by the manufacturer. The reaction consisted of 10 µL of 2× SYBR® Green, 1 µL of each primer for the target gene, and 2 µL of cDNA template. The reaction was conducted in a total volume of 20 µL, which was adjusted to 20 µL by adding PCR grade sterile nuclease free water. The thermal cycling protocol for IL-1β, β-defensin, β-actin, and EF1α genes consisted of an initial denaturation step at 95 °C for 3 min. to activate the DNA polymerase, followed by 40 cycles of denaturation at 95 °C for 30 s, combined annealing, and extension at 60 °C for 1 min., and a final extension at 95 °C for 10 s. The thermal cycling protocol for NKL involved an initial denaturation phase at 95 °C for 3 min., followed by 40 cycles of denaturation at 95 °C for 30 s., annealing at 58 °C for 30 s., and extension at 72 °C for 30 s., and a final extension step at 95 °C for 10 s. Fluorescent data was gathered during the extension process. No template control was included in all reactions. The thermal protocol for the melting curve analysis was conducted at the end of the amplification process, with a temperature range of 65℃ to 95℃ with increase by 0.5℃ every 0.05 s., and the cycle threshold (Ct) value was determined. The qRT-PCR tests were conducted using the CFX96™ Real-Time PCR detection system manufactured by BIO-RAD in the United States. The calculation of gene expression for the target genes was performed using the delta-delta Ct (2−ΔΔCt) method as described by Livak and Schmittgen [36].
Histological examination
Liver, anterior kidney, spleen, and intestinal samples were obtained from fish (2/replicate/group) at the 10th and 20th days of the feeding period; moreover, the same tissue samples were collected post-challenge from the challenge sampling tanks. The tissues were transferred to be fixed in 10% buffered formalin for histological examination. The fixed tissues undergo classical histological procedures through consecutive dehydration in the varying series of ethanol solution, clearing in xylene and consequently embedding in paraffin. Stained hematoxylin and eosin (H&E) sections were prepared for histomorphology identification [37].
Statistical analysis
The data analysis and figure construction were done using GraphPad Prism software (version 9.3.0, San Diego, CA, USA). The data were reported as the mean ± standard error of the mean (SEM). Statistical analysis was performed using a One-way analysis of variance test (ANOVA) to compare the two experimental groups. Differences were judged significant when the (P > 0.05). Subsequently, Tukey’s Post-Hoc test was employed to do multiple comparisons.
Results
Identification of the brown algae
The collected seaweed samples were identified as brown algae: P. boergesenii (Phaeophyceae, Dictyotaceae). The taxonomic identification of the seaweed is based on their morphological and anatomical characters.
Gas Chromatography-Mass Spectrometry (GC/MS) analysis
The GC–MS chromatogram (Fig. 1) showed various compounds present in P. boergesenii water extract. The biological activity and the major compounds with their retention time (RT) and peak area (%) are listed in Table 2.

Chromatogram obtained from the GC/MS of the watery extract of P. boergesenii
Serum biochemical parameters
The levels of total protein, albumin, globulin, and A: G ratio were cited in Table 3. It was found that the serum total protein and globulin levels were not significantly different between the supplemented and control groups on the 10th day of feeding, but both were increased in the supplemented group on the 20th day. The serum albumin level was not statistically different between the supplemented and control groups during the feeding experiment. The A: G ratio in the serum of the supplemented group was reduced significantly on the 20th day of feeding. The activity of TAC was significantly increased in the supplemented group (Table 3) during the time course of dietary supplementation with P. boergesenii water extract.
Challenge experiment
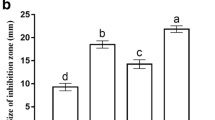
The results were presented in terms of percentage mortality (Fig. 2). The fish group injected with P. putida and fed on basal diet with no additives (control) revealed a total mortality rate of 73.33% within 14 days. They exhibited skin hemorrhages and skin ulceration, fins rot, dark pigmentation, detached scales, abdominal distention and tail and fins rot. Results of re-isolation of bacteria and their identification revealed the same characteristics of the injected bacteria.

Cumulative mortality percent of fish experimental groups challenged with P. putida
In the fish group fed on a diet supplemented with P. boergesenii water extract, a mortality of 26.7% was recorded and less sever clinical signs and lesions were observed during the challenge experiment. Whereas neither clinical signs nor mortality were observed during the experiment in the negative challenge control group.
Tissue distribution, and real-time PCR analysis of the expression levels of IL-1β, β-defensin and NKL
In this study, expression of the genes under study were statically evaluated in the supplemented group in comparison with their expression levels in the control group. There was a variation in the tissue level expression of IL-1β, β-defensin, and NKL in Nile tilapia fed with P. boergesenii water extract. IL-1β expression in the spleen of the supplemented group showed a significant upregulation on the 20th day of feeding of the experimental diet (Fig. 3A). While dietary supplementation with P. boergesenii had no effect on the β-defensin mRNA expression in the spleen of Nile tilapia (Fig. 3B). On the other hand, the expression of NKL gene in the spleen was upregulated during the time course of dietary supplementation with P. boergesenii water extract (Fig. 3C).

Gene expression profiles of IL-1β (A), β-defensin (B) and NKL (C) in the spleen of Nile tilapia following 10 and 20 days of feeding diet supplemented with 4.5% P. boergesenii water extract and 2 days post-challenge with P. putida. Data are presented as mean ± SEM (n = 3). The mean values denoted with different letters within the same experimental days are statistically significant (P < 0.05)
In addition, our study exhibited an upregulation in the IL-1β gene expression level in the intestine of the supplemented group during the feeding period (Fig. 4A). Whereas both AMPs (β-defensin and NKL) expression significantly upregulated in the intestine on the 20th day of feeding of the experimental diet (Fig. 4B & C).

Gene expression profiles of IL-1β (A), β-defensin (B) and NKL (C) in the intestine of Nile tilapia following 10 and 20 days of feeding diet supplemented with 4.5% P. boergesenii water extract and 2 days post-challenge with P. putida. Data are presented as mean ± SEM (n = 3). The mean values denoted with different letters within the same experimental days are statistically significant (P < 0.05)
Changes in the level of expression of the analyzed immune-related genes were also observed in the spleen and intestine after the P. putida challenge (Figs. 3 and 4), where all genes were significantly upregulated in both the spleen and intestine except the β-defensin expression in the spleen.
Histological findings
Pre-challenge fish
Macroscopic results
Liver, anterior kidney, spleen, and intestine of the control and supplemented fish groups seemed to appear healthy without injury.
Microscopic results
Microscopic examination of the liver of the control fish revealed apparently healthy histomorphological structures where it exhibits uniform hepatic parenchyma (Fig. 5A, B). The liver of fish fed P. boergesenii water extract for 10 days manifested symmetrical hepatocytes and activation of kupffer cells (Fig. 5C). Likewise, after 20 days of supplementation, the liver showed regular hepatocytes (Fig. 5D).

(A→D): Light microscopic sections with H &E stain of liver of control Nile tilapia (A, B), Nile tilapia fed diet supplemented with P. boergesenii water extract for 10 days (C) and Nile tilapia fed diet supplemented with P. boergesenii water extract for 20 days (D). (A) regular hepatocytes (arrow) and intact central vein (star). (B) intact hepatic cords (arrow). (C) well organized hepatic cords (arrow) and well identified kupffer cells (star). (D) significant detection of kupffer cells (arrow) beside this uniformly arranged hepatocytes (star)
Microscopic examination of the anterior kidney of the control fish illustrated normal nephritic tubules with minimally aggregated lymphocytes (Fig. 6A, B). Fish fed a diet supplemented with P. boergesenii water extract for 10 days provoked consistent architectures of the anterior kidney. Accordingly, kidney tissues were normally arranged in tubules with deeply infiltrated melanomacrophages (Fig. 6C). Simultaneously, regarding feeding with P. boergesenii for 20 days, the kidney showed regularly distributed tubules; besides this, a discrete distribution of melanomacrophages was observed (Fig. 6D).

(A→D): Light paraffin microscopic sections with H &E stain of anterior kidney of control Nile tilapia (A, B), Nile tilapia fed diet supplemented with P. boergesenii water extract for 10 days (C) and Nile tilapia fed diet supplemented with P. boergesenii water extract for 20 days (D). (A) well-identified tubules (arrow) and normal appearance of hematopoietic tissues (star). (B) apparently healthy nephritic tubules (arrow), besides minimal lymphocytes aggregations (star). (C) normal nephritic tubules (arrow), and melanomacrophages infiltration (star). (D) well organized nephritic tubules (arrow)
Microscopic examination of the spleen of the control fish exhibited splenic parenchyma in normal histological status (Fig. 7A, B). While the spleen of the supplemented group exhibited evidence of sharp activation of melanomacrophage cells either for a period of 10- or 20-days supplementation (Fig. 7C, D, respectively).

(A→D): Light paraffin microscopic sections with H &E stain of spleen of control Nile tilapia (A, B), Nile tilapia fed diet supplemented with P. boergesenii water extract for 10 days (C) and Nile tilapia fed diet supplemented with P. boergesenii water extract for 20 days (D). (A) normally arranged splenic parenchyma (arrow). (B) intact splenic capsules (arrow). (C) discrete distribution of melanomacrophages cells (arrow). (D) huge aggregation of melanomacrophages centers (arrow)
Microscopic examination of the intestine of the control fish revealed normally organized mucosa and submucosa layers, as well lack of goblet cells identification (Fig. 8A, B). In parallel to the control intestine histology, fish supplemented with P. boergesenii for 10 and 20 days demonstrated a good appearance of intestinal villi with increased goblet cells activation (Fig. 8C, D, respectively).

(A→D): Light paraffin microscopic sections with H &E stain of intestine of control Nile tilapia (A, B), Nile tilapia fed diet supplemented with P. boergesenii water extract for 10 days (C) and Nile tilapia fed diet supplemented with P. boergesenii water extract for 20 days (D). (A) normal epithelium lined intestinal villi (arrow). (B) less recognized goblet cells (arrow). (C) intact intestinal villi with increased goblet cells (arrow). (D) hyperactivation of goblet cells (arrow)
Post-challenge fish
Macroscopic results
Grossly, the organs under study, comprising the liver, anterior kidney, spleen, and intestine of the challenged supplemented group showed a good criterion. Meanwhile, the organs of the challenged control fish appeared distorted.
Microscopic results
Control fish injected with P. putida exhibited significant histological aberrations in the target organs. Nevertheless, the challenged supplemented fish showed an improvement in histomorphological architectures, as detailed in Table 4; Figs. 9, 10, 11 and 12.
Microscopic examination of the control liver injected with P. putida displayed histological alterations represented by a remarkable degree of degenerative changes in the hepatocytes with cytoplasmic vacuolation and intensive congestion in the blood vessels (Fig. 9A, B). In contrast, the supplemented diet ameliorates the histological lesions that were detected in the control group, which manifested vacuolar degeneration of the hepatocytes and mild congestion in the blood sinusoids. A significant expression in the kupffer cells was observed in the challenge supplemented group when compared with challenge control group (Fig. 9C, D).

(A→D): Light paraffin microscopic sections with H &E stain from liver of control Nile tilapia challenged with P. putida (A, B), liver of Nile tilapia challenged supplemented group (C, D). (A) Light section showing congestion in the central vein (arrow). (B) Light section showing necrosis with vacuolization of the hepatocytes (arrow). (C) Light section showing mild hepatic vacuolation (arrow). (D) Light section showing activation of the kpuffer cells (arrow), and slightly congested blood sinusoids (star)
Microscopic examination of the control anterior kidney injected with P. putida illustrated necrosis of the epithelial lining nephritic tubules, along with cytoplasmic vacuolation, and congestion in the blood vessels. Moderate depletion in the hematopoietic tissues was also seen (Fig. 10A, B). Compared to the challenged control group, healthy histological structures in the kidney of the challenged supplemented fish were observed, where they manifested intact nephritic tubules, and normal hematopoietic tissues (Fig. 10C, D).

(A→D): Light paraffin microscopic sections with H &E stain from anterior kidney of control Nile tilapia challenged with P. putida (A, B), anterior kidney of Nile tilapia challenged supplemented group (C, D). A) Light section showing necrosis of the nephritic tubules (arrow). (B) Light section showing necrosis with cytoplasmic vacuolation of the renal tubular epithelium (arrow), besides congestion in the blood vessels (star). (C) Light section showing regular nephritic tubules (arrow) with normal hematopoietic tissues (star). (D) Light section showing normal kidney histoarchitecture (arrow)
Microscopic examination of the control spleen injected with P. putida showed histopathological lesions in white and red pulps, where red pulps suffered congestion of the hematopoietic tissues, and depletion of white pulp with minimally infiltrated melanomacrophages in the splenic tissues (Fig. 11A, B). In comparison with infected fish spleen, supplemented fish group spleen showed normal splenic pulps, healthy hematopoietic tissues, and massive aggregation of melanomacrophage centers (Fig. 11C, D).

(A→D): Light paraffin microscopic sections with H &E stain from spleen of control Nile tilapia challenged with P. putida (A, B), spleen of Nile tilapia challenged supplemented group (C, D). (A) Light section showing congestion of the red pulp (arrow), moreover vacuolation of white pulp (star). (B) Light section showing minimal infiltration of melanomacrophages (arrow). (C) Light section showing intact white (arrow) and red (star) pulps. (D) Light section showing huge accumulation of melanomacrophages centers (arrow)
Microscopic examination of the control intestine injected with P. putida manifested necrosis of epithelial lining intestinal villi with subsequent disintegration and desquamation of the intestinal tips (Fig. 12A, B). Histological analysis of the intestine of the supplemented fish group showed apparent normal criteria of the intestinal layers with pronounced infiltration of mononuclear cells mainly lymphocytes (Fig. 12C, D).

(A→D): Light paraffin microscopic sections with H & E stain from intestine of control Nile tilapia challenged with P. putida (A, B), intestine of Nile tilapia challenged supplemented group (C, D). (A) Light section showing necrosis with sloughing of the intestinal tips (arrow). (B) High power of Fig. A showing necrosis with sloughing of the tips of intestinal villi (arrow). (C) Light section showing intestinal epithelium lined with normal columnar cells (arrow), besides prominent accumulation of lymphocytes (star). (D) Light section showing apparent intact intestinal villi (arrow)
Discussion
There is convincing evidence that herbal extracts stimulate the immune response and increase the disease resistance. Nevertheless, information concerning the stimulation and disease resistance of fish by the algae is lacking. Thus, our study aimed to investigate the effect of dietary supplementation with P. boergesenii water extract on the immunity and disease resistance of Nile tilapia.
The days of dietary supplementation for optimum induction of the immune response vary corresponding to the immunostimulant used. Herein, feeding Nile tilapia on a diet supplemented with P. boergesenii water extract for 20 successive days reflected a very non-specific immune response. The concurrent increase of both serum protein and globulin might be owed to the constant level of albumin. Elevations in serum protein and globulin concentrations are usually assumed to be correlated with a stronger innate immune response in fish [18] where globulins are created by the mononuclear phagocytes and they are critical components of the immune system.
These results coincide with those of Maghawri et al. [38], who found increases in total protein and globulin levels for the redbelly tilapia after using brown algae (Padina pavonica) as a feed additive. In a similar trend, previous studies in Labeo rohita after being fed diets supplemented with mixed algal extracts of Euglena viridis, Chlorella vulgaris, and Spirulina platensis [39] and diets supplemented with the seaweed, Sargassum wightii, and its fucoidan-rich extract [40].
The total antioxidant capacity is an important marker showing the state of the antioxidant function, including enzymatic and non-enzymatic antioxidants [41]. Our study revealed that dietary supplementation with P. boergesenii water extract for either 10 or 20 successive days significantly increased the TAC. These influences may be due to the structural properties of many secondary metabolites and bioactive compounds in P. boergesenii water extract, which have antioxidant activities that defend fish against oxidative injury, as confirmed by Oliveira et al. [15].
Interleukin-1β is an early-expressed proinflammatory cytokine that promotes the immune response by stimulating lymphocytes or generating the release of other cytokines that activate lymphocytes and macrophages [42]. The up-regulated IL-1β expression in our study confirms the immunostimulatory nature of the P. boergesenii water extract. Thus, dietary supplementation will clearly influence the gut mucosa after ingestion of the pellets. This site is a significant component of the mucosal immune system, that is one of the first barriers against fish infection [43], and hence this sensitization could lead to disease resistance. Also, the spleen, as the crucial immune organ, has been used as a marker of immune response in fish [44].
Our findings overlap with Watanuki et al. [45] who found increased IL-1β gene expression by administration of Spirulina sp. in common carp diets. Also, IL-1 gene expression was associated with the feeding of common carp with polysaccharides from the Padina gymnospora [46]. Research reported that algae extract of Capparis spinosa improved the expression of IL-1β in the rainbow [47].
Moreover, this study showed up-regulation of IL-1β 2 days post-challenge with P. putida in the spleen and intestinal tissues, as they are the predilection sites of P. putida, indicating the stronger innate immune response of Nile tilapia. A recent study documented up-regulation of the IL-1β after bacterial challenge in L. rohita fed a diet comprising mixed algal extracts [39].
Antimicrobial peptides are important components of the innate immune system [26]. Therefore, expression studies of the AMPs may offer important data regarding the stimulation of their expression by immunostimulants and their contribution in protection against pathogens.
Only a limited number of studies related to AMPs expression following administration of immunostimulants, and few have demonstrated whether there is time effect on various AMPs in different tissues. This was the focus of the present study, which analyzed the expression of β-defensin and NKL in the intestine as a mucosal tissue and the spleen as a central immune tissue. This study showed that dietary supplementation with P. boergesenii water extract had a various effect on the different AMPs families in tissues. Such changes increase the antibacterial defenses at those tissues, and disease the bacterial resistance affected by the various AMPs families can be predicted. There are numerous causes that explain these changes in gene expression. For example, the expression levels of AMPs could relate to alterations in the leucocyte types in the tissues post-P. boergesenii administration or to alteration in their activation state. The expression of AMPs has been induced by pathogenic challenges [48, 49] as well as by immunostimulants in fish [26, 40].
Antimicrobial peptides have demonstrated their efficacy against gram-negative bacteria [50], including P. putida. Various AMP mechanisms have been examined, leading to the development of a model known as the Shai-Matsuzaki-Huang model. This model postulates that cationic AMPs can utilise their positive charge to attach to the anionic elements of the bacterial cell surface [21]. This attachment is suggested to cause killing of the bacterial cells through the leakage of bacterial cellular components.
Herein, this study is the first to record promoted immune-related gene expression levels in Nile tilapia fed a diet supplemented with P. boergesenii water extract. Therefore, this algae species can be listed as one of the immunostimulants that can modulate the expression of critical immune genes.
In recent years, there has been an increase in interest in algae extraction’s prophylactic abilities against bacterial challenges to aquatic animals. It has been stated that various algae extractions offer resistance to virulent bacteria Aeromonas hydrophila and Edwardsiella tarda in common carp [46], A. hydrophila in tilapia [51], A. hydrophila in rainbow trout [39] and to P. aeruginosa in Cirrhinus mrigala [16]. Nevertheless, in the literature, there is no study on the resistance of the P. boergesenii -fed Nile tilapia to P. putida.
In this study, a fish group fed on a diet supplemented with water extract of P. boergesenii showed a decrease in mortality rate and clinical signs, which could be attributed to the presence of different secondary metabolites such as flavonoids and fatty acids contents [15]. Where the GC-MS analysis of the volatile compounds of the marine algal extract showed that they have antibacterial [52, 53], and antioxidant activities [54, 55] that have a protective role in reducing the ulcerative injury induced by the P. putida toxins. Moreover, as confirmed herein, increased serum globulin may further promote resistance to infection. The findings suggested that the water extract of P. boergesenii has a promising potential immunostimulant, opening the door for the isolation of its active components.
The main lymphoid tissues are the liver, head kidney, spleen, and gastric mucosa in fish [56]. Accordingly, these tissues constitute the primitive analogues of fish lymph nodes for immune function [57]. According to our study, dietary inclusion of algae in aquafeed poses a synergistic effect on cell morphology with improved histoarchitectures, contrary to control fish. Hence, the histological observations on liver, kidney, spleen, and intestinal tissues confirm that the addition of P. boergesenii water extract to the fish feed exerts a histoprotective effect in Nile tilapia fish. These observations confirmed the protective impact of P. boergesenii water extract on the injured livers and other tissues induced by P. putida in the challenge experiment, permitting the alleviation of the histological deformations. This effect was probably mediated by the antioxidant and cytotoxic potential of the P. boergesenii water extract, owing to its flavonoids and fatty acid contents [15]. Algae-derived compounds could enhance the fish’s physiological function as well as change the histological damage [58].
Currently, at the histological level, P. boergesenii stimulates the cells that contribute to the immune response. Hence, the Kupffer cells were significantly activated in the liver sinusoids. Algae-derived bioactive ingredients, that can promote individual’s immune system, should be thought to raise the health condition of fish [59]. These findings are in parallel with Neyrinck et al. [60], who ensured that brown algae, Laminaran, when administered orally, could exert hepatoprotection. Herein, tilapia fed P. boergesenii water extract, followed by prominent activation of Melanomacrophage centers. Melanomacrophages serve as an applicable histological monitor of the immune response in fish [61]. Usage of spirulina (Arthrospira platensis) in feeding Nile tilapia displayed normal histomorphological structures in the liver, kidney, and spleen, with remarkable detection of melanomacrophage centers [62]. Also, the intestinal mucosa recorded proliferation of goblet cells and lymphocyte aggregation. A micro- and macroalgal blend could modulate the immune function of European seabass by supporting the migration of lymphocytes and monocytes to mucosal tissue and activating the proliferation of goblet cells in the intestine [59]. Similarly, Atlantic salmon, when fed an algae diet of Schizochytrium sp., resulted in a positive effect on salmon intestine health by enhancing intestinal mucus and goblet cell production [63]. Moreover, European seabass supplemented Gracilaria gracilis seaweed and Nannochloropsis oceanica microalga, which led to proliferation of goblet cells in the intestine [64].
Conclusion
Feeding Nile tilapia with a diet supplemented with P. boergesenii water extract for 20 days significantly increased the serum globulin level and TAC activity and upregulated the expression of IL-1β, β-defensin, and NKL genes, along with their improvement to the cell morphology of the immune tissues, which all may promote resistance against P. putida infection. The experimental design and conclusion of the results are illustrated in Fig. 13.

The experimental design and conclusion of the results
Data availability
Requests for materials should be addressed to Karima A. Bakry and Walaa F.A. Emeish, Accession numbers: P. putida using 16s rDNA gene sequencing (GenBank accession no. OM048106, 838 bp).
Abbreviations
- P. boergesenii :
-
Padina boergesenii
- O. niloticus :
-
Oreochromis niloticus
- P. putida :
-
Pseudomonas putida
- GC-MS:
-
Gas Chromatography Mass Spectrometry
- ppt:
-
Parts per thousand
- TAC:
-
Total antioxidant capacity
- IL-1β :
-
Interleukin-1beta
- AMPs:
-
Antimicrobial peptides
- β-defensin :
-
Beta-defensin
- NKl :
-
Natural killer-lysin
- NK:
-
Natural killer
- CTLs:
-
Cytotoxic T lymphocytes
- cfu:
-
Colony-forming unit
- rDNA:
-
Recombinant Deoxyribonucleic acid
- RNA:
-
Ribonucleic acid
- Ct:
-
Cycle threshold
- H&E:
-
Hematoxylin and eosin
- SEM:
-
Standard Error of Mean
- ANOVA:
-
Analysis of variance
- RT:
-
Retention time
- A:G ratio:
-
Albumin/globulin ratio
- cDNA:
-
Complementary Deoxyribonucleic acid
- β -actin :
-
Beta actin
- EF1α :
-
Elongation factor 1 alpha
- qRT-PCR:
-
Quantitative real-time Polymerase chain reaction
References
Selim KM, Reda RM. Improvement of immunity and disease resistance in the Nile tilapia, Oreochromis niloticus, by dietary supplementation with Bacillus amyloliquefaciens. Fish Shellfish Immunol. 2015;44(2):496–503.
Syahidah A, Saad CR, Mohd Daud H, Abdelhadi YM. Status and potential of herbal applications in aquaculture: a review. Iran J Fisheries Sci. 2015;14:27–44.
Eissa A. Clinical and Laboratory Manual of Fish diseases. Germany: LAP Lambert Academic Publishing; 2016. p. 140.
Altinok I, Kayis S, Capkin E. Pseudomonas putida infection in rainbow trout. Aquaculture. 2006;261:850–5.
Enany M, Eidaroos N, Eltamimy N. Microbial causes of Summer Mortality in Farmed Fish in Egypt. Suez Canal Veterinary Med J SCVMJ. 2019;24:45–56.
Mao Z, Qiu Y, Zheng L, Chen J, Yang J. Development of a visual loop-mediated isothermal amplification method for rapid detection of the bacterial pathogen Pseudomonas putida of the large yellow croaker (Pseudosciaena crocea). J Microbiol Methods. 2012;89(3):179–84.
El-Barbary M, Hal A. Phenotypic and genotypic characterization of some Pseudomonas sp. Associated with Burkholderia cepacia isolated from various infected fishes. J Aquaculture Res Dev. 2017;8(7):1–7.
Huys G, Rhodes G, McGann P, Denys R, Pickup R, Hiney M, Smith P, Swings J. Characterization of oxytetracycline-resistant heterotrophic bacteria originating from hospital and freshwater fishfarm environments in England and Ireland. Syst Appl Microbiol. 2000;23(4):599–606.
Angulo FJ, Nargund VN, Chiller TC. Evidence of an association between use of anti-microbial agents in food animals and anti-microbial resistance among bacteria isolated from humans and the human health consequences of such resistance. J Veterinary Med B Infect Dis Veterinary Public Health. 2004;51(8–9):374–9.
Hektoen H, Berge JA, Hormazabal V, Yndestad M. Persistence of antibacterial agents in marine sediments. Aquaculture. 1995;133:175–84.
Defoirdt T, Sorgeloos P, Bossier P. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr Opin Microbiol. 2011;14:251–8.
Akhter N, Wu B, Memon AM, Mohsin M. Probiotics and prebiotics associated with aquaculture: a review. Fish Shellfish Immunol. 2015;45(2):733–41.
Balaraman D, Subramanian V. Applications of immunostimulants in aquaculture: a review. Int J Curr Microbiol Appl Sci. 2016;5:447–53.
Kayalvizhi K, Subramanian V, Anantharaman P, Kandasamy K. Antimicrobial activity of seaweeds from the gulf of mannar. Int J Pharm Appl. 2012;3:306–14.
Oliveira NM, Meira CLC, Aguiar R, Oliveira DM, Moura CWN, Filho S. Biological activities of extracts from Padina boergesenii and Sargassum stenophyllum, Seaweeds naturally found in baia de todos os santos, Brazil. Int J Pharm Pharm Sci. 2015;7:350–3.
Ragunath C, Ramasubramanian V. Dietary Effect of Padina boergesenii on growth, Immune Response, and Disease Resistance against Pseudomonas aeruginosa in Cirrhinus mrigala. Appl Biochem Biotechnol. 2022;194(5):1881–97.
Yılmaz S, Acar Ü, Kesbiç OS, Gültepe N, Ergün S. Effects of dietary allspice, Pimenta dioica powder on physiological responses of Oreochromis mossambicus under low pH stress. SpringerPlus. 2015;4:719.
Yilmaz S, Ergün S, Çelik E. Effect of Dietary spice supplementations on Welfare Status of Sea Bass, Dicentrarchus labrax L. Proc Natl Acad Sci India Sect B: Biol Sci. 2016;86:229–37.
Bo YX, Song XH, Wu K, Hu B, Sun BY, Liu ZJ, Fu JG. Characterization of interleukin-1β as a proinflammatory cytokine in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015;46(2):584–95.
Pooley NJ, Tacchi L, Secombes CJ, Martin SA. Inflammatory responses in primary muscle cell cultures in Atlantic salmon (Salmo salar). BMC Genomics. 2013;14:747.
Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389–95.
Cuesta A, Meseguer J, Esteban MA. Molecular and functional characterization of the gilthead seabream beta-defensin demonstrate its chemotactic and antimicrobial activity. Mol Immunol. 2011;48:1432–8.
Smith VJ, Desbois AP, Dyrynda EA. Conventional and unconventional antimicrobials from fish, marine invertebrates and micro-algae. Mar Drugs. 2010;8(4):1213–62.
Falco A, Chico V, Marroquí L, Perez L, Coll JM, Estepa A. Expression and antiviral activity of a beta-defensin-like peptide identified in the rainbow trout (Oncorhynchus mykiss) EST sequences. Mol Immunol. 2008;45(3):757–65.
Xu W, Faisal M. Defensin of the zebra mussel (Dreissena polymorpha): molecular structure, in vitro expression, antimicrobial activity, and potential functions. Mol Immunol. 2010;47(11–12):2138–47.
Marel M, Adamek M, Gonzalez SF, Frost P, Rombout JH, Wiegertjes GF, Savelkoul HF, Steinhagen D. Molecular cloning and expression of two β-defensin and two mucin genes in common carp (Cyprinus carpio L.) and their up-regulation after β-glucan feeding. Fish Shellfish Immunol. 2012;32(3):494–501.
Andrä J, Monreal D, Martinez de Tejada G, Olak C, Brezesinski G, Gomez SS, Goldmann T, Bartels R, Brandenburg K, Moriyon I. Rationale for the design of shortened derivatives of the NK-lysin-derived antimicrobial peptide NK-2 with improved activity against Gram-negative pathogens. J Biol Chem. 2007;282(20):14719–28.
Allender BM, Kraft GT. The marine algae of Lord Howe Island (New South Wales): the Dictyotales and Cutleriales (Phaeophyta). Brunonia. 1983;6:73–130.
Cho JY, Jin HJ, Lim HJ, Whyte JNC, Hong YK. Growth activation of the microalga isochrysisgalbana by the aqueous extract of the seaweed Monostromanitidum. J Appl Phycol. 1999;10:561–7.
Gonzalez DVA, Platas G, Basilio A. Screening of antimicrobial activities in red, green and brown macroalgae from Gran Canaria (Canary Islands, Spain). Int Microbiol. 2001;4:35–40.
Ghaednia B, Mehrabi M, Mirbakhsh M, Yeganeh V, Dashtiannasab A. Administration of hot-water extract of Padina boergesenii via immersion route to enhance haemolymph-immune responses of Fenneropenaeus indicus (Edwards). Aquac Res. 42 (2010).
Hamackova J. Clove oil as an anaesthetic for different freshwater fish species. Bulgarian J Agricultural Sci. 2006;12:185.
Busher J. Serum albumins and globulin. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.;Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, UK, Chap. 101. (1990).
Koracevic D, Koracevic G, Djordjevic V, Andrejevic S, Cosic V. Method for the measurement of antioxidant activity in human fluids. J Clin Pathol. 2001;54(5):356–61.
Goldman E, Green LH. Practical Handbook of Microbiology. 3rd ed. Boca Raton, FL, USA: CRC; 2015.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2–∆∆CT method. Methods. 2001;25:402–8.
Bancroft JD, Gamble M. Theory and practice of histological techniques. 6th Edition, Churchill Livingstone, Elsevier, China. (2008).
Maghawri A, Marzouk SS, El-Din E, Zaghloul HM, G.Y., Nashaat M. Dietary Effect of Microalga Padina pavonica richness with fatty acids on the growth and Immune Status of Coptodon zilli. Egypt J Aquat Biology Fisheries. 2023;27(6):225–42.
Sattanathan G, Liu W-C, Padmapriya S, Pushparaj K, Sureshkumar S, Lee J-W, Balasubramanian B, Kim IH. Effects of Dietary Blend of Algae Extract supplementation on growth, biochemical, Haemato-Immunological response, and Immune Gene expression in Labeo rohita with Aeromonas hydrophila Post-challenges. Fishes. 2023;8(1):7.
Gora A, Sahu N, Sahoo S, Rehman S, Dar S, Ahmad I, Agarwal D. Effect of dietary Sargassum Wightii and its fucoidan-rich extract on growth, immunity, disease resistance and antimicrobial peptide gene expression in Labeo rohita. Int Aquat Res. 2018;10:115–31.
El-Naggar M, Salaah S, El-Shabaka H, El-Rahman A, Khalil M, Suloma A. Efficacy of dietary chitosan and chitosan nanoparticles supplementation on health status of Nile tilapia, Oreochromis niloticus (L). Aquaculture Rep. 19 (2021).
Low C, Wadsworth S, Burrells C, Secombes CJ. Expression of immune genes in turbot (Scophthalmus maximus) fed a nucleotide-supplemented diet. Aquaculture. 2003;221:23–40.
Alvarez-Pellitero P. Fish immunity and parasite infections: from innate immunity to immunoprophylactic prospects. Vet Immunol Immunopathol. 2008;126(3–4):171–98.
Vazirzadeh A, Dehghan F, Kazemeini R. Changes in growth, blood immune parameters and expression of immune related genes in rainbow trout (Oncorhynchus mykiss) in response to diet supplemented with Ducrosia Anethifolia essential oil. Fish Shellfish Immunol. 2017;69:164–72.
Watanuki H, Ota K, Tassakka AR, Kato A, T., Sakai M. Immunostimulant effects of dietary Spirulina platensis on carp, Cyprinus carpio. Aquaculture. 2006;258:157–63.
Rajendran P, Subramani PA, Michael D. Polysaccharides from marine macroalga, Padina gymnospora improve the nonspecific and specific immune responses of Cyprinus carpio and protect it from different pathogens. Fish Shellfish Immunol. 2016;58:220–8.
Bilen S, Altunoglu YC, Ulu F, Biswas G. Innate immune and growth promoting responses to caper (Capparis spinosa) extract in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2016;57:206–12.
Chen Y, Zhao H, Zhang X, Luo H, Xue X, Li Z, Yao B. Identification, expression and bioactivity of Paramisgurnus dabryanus β-defensin that might be involved in immune defense against bacterial infection. Fish Shellfish Immunol. 2013;35(2):399–406.
Huang Y, Zheng Q, Niu J, Tang J, Wang B, Abarike ED, Lu Y, Cai J, Jian J. NK-lysin from Oreochromis niloticus improves antimicrobial defence against bacterial pathogens. Fish Shellfish Immunol. 2018;72:259–65.
Hsieh JC, Pan CY, Chen JY. Tilapia hepcidin (TH)2–3 as a transgene in transgenic fish enhances resistance to Vibrio vulnificus infection and causes variations in immune-related genes after infection by different bacterial species. Fish Shellfish Immunol. 2010;29(3):430–9.
Abu-Elala N, Galal MK, Abd-Elsalam RM, Mohey-Elsaeed O, Ragaa NM. Effects of Dietary Supplementation of Spirulina platensis and garlic on the growth performance and expression levels of Immune-related genes in Nile tilapia (Oreochromis niloticus). J Aquaculture Res Dev. 2016;7:433.
Deepak P, Sowmiya R, Balasubramani G, Perumal P. Phytochemical profiling of Turbinaria ornata and its antioxidant and anti-proliferative effects. J Taibah Univ Med Sci. 2017;12(4):329–37.
Jinfeng EC, Mohamad Rafi MI, Chai Hoon K, Kok Lian H. Yoke Kqueen, C. Analysis of chemical constituents, antimicrobial and anticancer activities of dichloromethane extracts of Sordariomycetes sp. endophytic fungi isolated from Strobilanthes Crispus. World J Microbiol Biotechnol. 2017;33(1):5.
Chandrasekaran M, Senthilkumar A, Venkatesalu V. Antibacterial and antifungal efficacy of fatty acid methyl esters from the leaves of Sesuvium portulacastrum L. Eur Rev Med Pharmacol Sci. 2011;15(7):775–80.
Shaaban MT, Ghaly MF, Fahmi SM. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. J Basic Microbiol. 2021;61(6):557–68.
Koppang EO, Fischer U, Moore L, Tranulis MA, Dijkstra JM, Köllner B. Hordvik, I. Salmonid T cells assemble in the thymus, spleen and in novel interbranchial lymphoid tissue. J Anat. 2010;217(6):728–39.
Sundaresan M. Ultrastructure of spleen in the freshwater fish, Tilapia mossambica (Peters). Eur Acad Res. 2014;2(2):2894–908.
Abo-Shady AM, Gheda SF, Ismail GA, Cotas J, Pereira L, Abdel-Karim OH. Antioxidant and antidiabetic activity of algae. Life. 2023;13(2):460.
Ferreira M, Machado M, Mota CS, Abreu H, Silva J, Maia MR, Valente LM. Micro-and macroalgae blend modulates the mucosal and systemic immune responses of European seabass (Dicentrarchus labrax) upon infection with Tenacibaculum maritimum. Aquaculture. 2023;566:739222.
Neyrinck AM, Mouson A, Delzenne NM. Dietary supplementation with laminaran, a fermentable marine ß(1–3) glucan, protects against hepatoxicity induced by LPS in rat by modulating immune response in the hepatic tissue. Int Immunopharmacol. 2007;7:1497–506.
Steinel NC, Bolnick DI. Melanomacrophage centers as a histological indicator of immune function in fish and other poikilotherms. Front Immunol. 2017;8:827.
Mabrouk MM, Ashour M, Labena A, Zaki MA, Abdelhamid AF, Gewaily MS, Ayoub HF. Nanoparticles of Arthrospira platensis improves growth, antioxidative and immunological responses of Nile tilapia (Oreochromis niloticus) and its resistance to Aeromonas hydrophila. Aquac Res. 2022;53(1):125–35.
Kousoulaki K, Østbye TKK, Krasnov A, Torgersen JS, Mørkøre T, Sweetman J. Metabolism, health and fillet nutritional quality in Atlantic salmon (Salmo salar) fed diets containing n-3-rich microalgae. Journal of nutritional science. 4,e24 (2015).
Batista S, Pereira R, Oliveira B, Baião LF, Je ssen F, Tulli F, Messina M, Silva JL, Abreu H, Valente LMP. Exploring the potential of seaweed Gracilaria gracilis and microalga Nannochloropsis Oceanica, single or blended, as natural dietary ingredients for European seabass Dicentrarchus labrax. J Appl Phycol. 2020;32:2041–59.
Altinok I. Multiplex PCR assay for detection of four major bacterial pathogens causing rainbow trout disease. Dis Aquat Organ. 2011;93(3):199–206.
Chang CI, Chen LH, Hu YF, Wu CC, Tsai JM. Determining the cleavage site for the mature antimicrobial peptide of Nile tilapia β-defensin using 2D electrophoresis, Western blot, and mass spectrometry analysis. Fish Shellfish Immunol. 2017;62:41–6.
Choi K, Lehmann DW, Harms CA, Law JM. Acute hypoxia-reperfusion triggers immunocompromise in Nile tilapia. J Aquat Anim Health. 2007;19(2):128–40.
He AY, Liu CZ, Chen LQ, Ning LJ, Zhang ML, Li EC, Du ZY. Identification, characterization and nutritional regulation of two isoforms of acyl-coenzyme A oxidase 1 gene in Nile tilapia (Oreochromis niloticus). Gene. 2014;545(1):30–5.
Pang JC, Gao FY, Lu MX, Ye X, Zhu HP, Ke XL. Major histocompatibility complex class IIA and IIB genes of Nile tilapia Oreochromis Niloticus: genomic structure, molecular polymorphism and expression patterns. Fish Shellfish Immunol. 2013;34(2):486–96.
Ryssel H, Kloeters O, Germann G, Schäfer T, Wiedemann G, Oehlbauer M. The antimicrobial effect of acetic acid–an alternative to common local antiseptics? Burns: J Int Soc Burn Injuries. 2009;35(5):695–700.
Essam N, Rajendrasozhan S, Hassan S, Sharawy S, Humaidi J. Phytochemical screening of different organic crude extracts from the stem bark of Ziziphus spina-christi (L.). Biomedical Research (India). 29,1645–1652 (2018).
Mohamad OAA, Li L, Ma JB, Hatab S, Xu L, Guo JW, Rasulov BA, Liu YH, Hedlund BP, Li WJ. Evaluation of the antimicrobial activity of Endophytic Bacterial Populations from Chinese Traditional Medicinal Plant Licorice and characterization of the Bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium Dahliae. Front Microbiol. 2018;9:924.
Meechaona R, Sengpracha W, Banditpuritat J, Kawaree R, Phutdhawong W. Fatty acid content and antioxidant activity of Thai bananas. Maejo Int J Sci Technol. 2007;1:222–8.
Othman AR, Abdullah N, Ahmad S, Ismail IS, Zakaria MP. Elucidation of in-vitro anti-inflammatory bioactive compounds isolated from Jatropha curcas L. plant root. BMC Complement Altern Med. 2015;15(1):11.
Sahu MK, Suthakaran S, Ghosh SC, Singh D, Das A, Jha H. Anticancer activity of secondary metabolite isolated from the rhizospheric fungus Fusarium oxysporum isolate-ABRF1, 2-propenoic acid, pentadecyl ester. Asian J Nat Prod Biochem. 21(2) (2023).
Kuppuswamy KM, Jonnalagadda B, Arockiasamy S. GC-MS analysis of chloroform extract of Croton Bonplandianum. Int J Pharma Bio Sci. 2013;4:613–7.
Yang L, Ge S, Huang J, Bao X. Synthesis of novel (E)-2-(4-(1H-1,2,4-triazol-1-yl)styryl)-4- (alkyl/arylmethyleneoxy) quinazoline derivatives as antimicrobial agents. Mol Diversity. 2018;22(1):71–82.
Malathi K, Anand A, Sudha R. Ethyl Iso-allocholate from a Medicinal Rice Karungkavuni inhibits Dihydropteroate synthase in Escherichia coli: a Molecular Docking and Dynamics Study. Indian J Pharm Sci. 2016;78(6):780–8.
Elagbar ZA, Naik RR, Shakya AK, Bardaweel SK. Fatty Acids Analysis, Antioxidant and Biological Activity of Fixed Oil of Annona muricata L. Seeds. Journal of Chemistry. 2016,6948098 (2016).
Sheela D, Uthayakumari F. GC-MS analysis of bioactive constituents from coastal sand dune taxon-Sesuvium portulacastrum. Bioscience Discovery. 2013;4:47–53.
Agarwal A, Prajapati R, Raza SK, Thakur LK. GC-MS analysis and antibacterial activity of aerial parts of Quisqualis indica plant extracts. Indian J Pharm Educ Res. 2017;51(2):329–36.
Asnaashari S, Delazar A, Safarzadeh E, Tabibi H, Mollaei S, Rajabi A, Asgharian P. Phytochemical analysis and various Biological activities of the Aerial Parts of Scrophularia Atropatana growing in Iran. Iran J Pharm Research: IJPR. 2019;18(3):1543–55.
Sowmya S, Palanisamy PC, Gopalakrishnan VK. Chromatographic and spectrophotometric analysis of bioactive compounds from Cayratia trifolia (L.) stem. Int J Pharm Pharm Sci. 2016;8:56–64.
Chelliah R, Ramakrishnan SR, Antony U. Nutritional quality of Moringa oleifera for its bioactivity and antibacterial properties. Int Food Res J. 2017;24:825–33.
Dahham SS, Tabana YM, Iqbal MA, Ahamed MB, Ezzat MO, Majid AS, Majid AM. The Anticancer, antioxidant and Antimicrobial properties of the Sesquiterpene β-Caryophyllene from the essential oil of Aquilaria crassna. Molecules. 2015;20(7):11808–29.
Nile SH, Keum YS, Nile AS, Jalde SS, Patel RV. Antioxidant, anti-inflammatory, and enzyme inhibitory activity of natural plant flavonoids and their synthesized derivatives. J Biochem Mol Toxicol. 2018;32(1):e22002.
Acknowledgements
The Science, Technology, and Innovation Funding Authority (STDF) and the Egyptian Knowledge Bank (EKB) have partnered to provide open access funding. The authors would like to thank the members of the Cell and Molecular Physiology Research Center, Faculty of Veterinary Medicine, South Valley University, Qena, Egypt, for their help and support the facilities needed for the molecular investigation.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
K. A. B., M. N., Z.A., E. k., M. K. N. K.s, P. E. M., M.A. K., A.A.A.A, H. H. A., W. F.A. E. contributed to study concept, design, data acquisition, data analysis, interpretation, manuscript editing, and manuscript review.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The methodology and fish experiments undertaken in this study were approved by the Ethical Committee of the Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt. These experiments adhered to the regulations set by the OIE guidelines for the use of animals in research, under the approval No. 06/2024/0179. All procedures were carried out in conformity with the applicable rules and regulations. The study was conducted in accordance with the ARRIVE (Animals in Research: Reporting In Vivo Experiments) criteria.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Bakry, K.A., Nasr, M., Al-Amgad, Z. et al. Resistance of Nile tilapia fed with Padina boergesenii extract to Pseudomonas putida infection. BMC Vet Res 20, 281 (2024). https://doi.org/10.1186/s12917-024-04115-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-024-04115-7