
Abstract
Background
Previous studies, including Mendelian randomization (MR), have demonstrated type 2 diabetes (T2D) and glycemic traits are associated with increased risk of metabolic dysfunction-associated steatotic liver disease (MASLD). However, few studies have explored the underlying pathway, such as the role of iron homeostasis.
Methods
We used a two-step MR approach to investigate the associations of genetic liability to T2D, glycemic traits, iron biomarkers, and liver diseases. We analyzed summary statistics from various genome-wide association studies of T2D (n = 933,970), glycemic traits (n ≤ 209,605), iron biomarkers (n ≤ 246,139), MASLD (n ≤ 972,707), and related biomarkers (alanine aminotransferase (ALT) and proton density fat fraction (PDFF)). Our primary analysis was based on inverse-variance weighting, followed by several sensitivity analyses. We also conducted mediation analyses and explored the role of liver iron in post hoc analysis.
Results
Genetic liability to T2D and elevated fasting insulin (FI) likely increased risk of liver steatosis (ORliability to T2D: 1.14 per doubling in the prevalence, 95% CI: 1.10, 1.19; ORFI: 3.31 per log pmol/l, 95% CI: 1.92, 5.72) and related biomarkers. Liability to T2D also likely increased the risk of developing liver cirrhosis. Genetically elevated ferritin, serum iron, and liver iron were associated with higher risk of liver steatosis (ORferritin: 1.25 per SD, 95% CI 1.07, 1.46; ORliver iron: 1.15 per SD, 95% CI: 1.05, 1.26) and liver cirrhosis (ORserum iron: 1.31, 95% CI: 1.06, 1.63; ORliver iron: 1.34, 95% CI: 1.07, 1.68). Ferritin partially mediated the association between FI and liver steatosis (proportion mediated: 7%, 95% CI: 2–12%).
Conclusions
Our study provides credible evidence on the causal role of T2D and elevated insulin in liver steatosis and cirrhosis risk and indicates ferritin may play a mediating role in this association.
Highlights
• Ferritin possibly mediates the association of insulin resistance in liver steatosis. Targeting the reduction of ferritin may mitigate the risk of liver steatosis arising from elevated insulin.
• Liver iron is likely to be positively associated with liver steatosis and the irreversible stage, liver cirrhosis, and mediates the association of ferritin and liver steatosis.
• Genetically elevated HbA1c is shown to decrease the risk of liver steatosis due to its erythrocytic property. Caution should be applied when using HbA1c values for MASLD management.
Similar content being viewed by others

Background
Metabolic dysfunction-associated steatotic liver disease (MASLD), formerly known as non-alcoholic fatty liver disease [1], is a significant worldwide health issue that affects approximately 32% of the population [2]. Despite its widespread prevalence, effective treatments for MASLD remain elusive [2]. The spectrum of MASLD ranges from steatosis with > 5% hepatic fat accumulation, and progression to steatohepatitis with ballooning, inflammation, fibrosis, and even liver cirrhosis [3]. The presence of MASLD is related to other major chronic diseases, notably type 2 diabetes (T2D) and insulin resistance, although evidence from earlier studies suggests that this association may be bi-directional [4, 5]. Among possible mechanisms linking these two diseases, ferritin (a protein in cells for iron storage which can be found in hepatocytes) has been proposed to play a role in hepatic steatosis, inflammation, and fibrosis [6]. Meta-analyses of observational studies have shown people with T2D have higher ferritin, which is a risk factor for the severity of advanced fibrosis [7, 8]. Despite these postulated pathways, few studies have investigated the mediating role of ferritin and other iron homeostasis biomarkers in the associations of T2D or glycemic traits in MASLD. However, these studies were mainly observational, and hence are vulnerable to confounding and reverse causation [9, 10].
Mendelian randomization (MR), a study design that relies on genetic variants randomly allocated at conception, is increasingly used to investigate etiologic questions given that it is less prone to confounding than conventional observational studies [11], and tends to give more consistent results with randomized controlled trials, such as the lack of association of HDL-cholesterol in coronary artery disease [12]. Previous MR studies have revealed that genetically elevated fasting insulin (FI) and liability to T2D increase alanine aminotransferase (ALT) and MASLD risk [13,14,15], whereas some MR studies have shown higher systemic iron status and iron homeostasis biomarkers (e.g., ferritin, serum iron, or transferrin saturation (TSAT)) may increase risk of MASLD [16, 17], but with some exceptions [18, 19]. These discrepancies could be driven by a lack of statistical power, or possible biases including highly pleiotropic variants (e.g., rs1800562 in HFE) which relate to the predominant type of hemochromatosis [20]. A recent MR study found genetic liability to T2D and glycemic traits may impact iron homeostasis biomarkers, particularly ferritin, whilst the association of most iron homeostasis biomarkers with T2D was unlikely causal [21]. This suggests a possible mechanistic pathway in which hyperglycemia and elevated insulin relate to MASLD. Although, to the best of our knowledge, this has not been explored using an MR design. Therefore, we conducted a two-step MR study to comprehensively assess the mediating role of different iron homeostasis biomarkers in the associations of genetic liability to T2D and glycemic traits in liver steatosis, liver cirrhosis and its biomarkers (ALT and magnetic resonance imaging (MRI)-derived proton density fat fraction (PDFF)) [22, 23]. We additionally included MRI-derived liver iron as a possible mediator to explore the potential differential effects compared to blood-based measures of iron homeostasis.
Methods
Study design
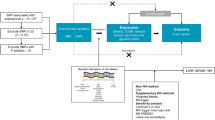
We adopted a two-step MR design using summary statistics from relevant genome-wide association studies (GWAS) (Fig. 1A). To estimate the overall effect, we first used a standard MR approach to evaluate the associations of liability to T2D and glycemic traits (exposures) on liver steatosis, liver cirrhosis and its biomarkers (outcomes) (Fig. 1B). We then extracted the associations of liability to T2D and glycemic traits (exposures) on iron homeostasis biomarkers (mediators) based on our previous study (Step 1, see Additional file 1: Table S1) [21]. Afterwards, we assessed the associations between these selected mediators and the liver-related outcomes (Step 2) (Fig. 1B). We also investigated the role of liver iron content in the associations of iron homeostasis biomarkers with the liver-related outcomes as a post hoc analysis (Fig. 1B). As with all MR analyses, there are three core assumptions, including relevance, independence, and exclusion restriction [23].

Study design of this study and diagram of this two-step Mendelian randomization analysis. A Study design of this study. B Diagram of this two-step Mendelian randomization. Liability to T2D, liability to type 2 diabetes; FG, fasting glucose; FI, fasting insulin; 2hGlu, 2-hour glucose; HbA1c, hemoglobin A1c; TIBC, total iron-binding capacity; TSAT, transferrin saturation; PDFF, proton density fat fraction; ALT, alanine aminotransferase. IVW, inverse-variance weighting; WM, weighted median; MR-PRESSO, Mendelian Randomization Pleiotropy RESidual Sum and Outlier
This study was conducted in accordance with the Strengthening the Reporting of Observational Studies in Epidemiology using MR (STROBE-MR) (Additional file 2: Supplementary Note) [24].
Data sources
Genetic instruments for T2D, glycemic traits, iron homeostasis biomarkers, and liver iron as exposures
We selected genetic instruments for our study based on specific criteria. These instruments included single nucleotide polymorphisms (SNPs) that demonstrated strong associations with our exposure of interest, with a significance threshold of P < 5 × 10−8. We ensured that these SNPs were independent of each other, with a linkage disequilibrium (LD) threshold of r2 < 0.001. To reduce potential bias from population stratification, we exclusively used data from individuals of European ancestries. We gathered the genetic instruments for our study from several datasets. For T2D, we obtained data from the Diabetes Meta-Analysis of Trans-Ethics Associations Studies (DIAMANTE) Consortium, which encompassed 80,154 cases and 853,816 controls [25]. Additionally, we obtained information on glycemic traits, including fasting glucose (FG) measured in mmol/l (n = 209,605), 2-hour glucose (2hGlu) measured in mmol/l (n = 64,469), FI in log pmol/l (n = 158,550), and hemoglobin A1c (HbA1c) in percentage (n = 149,006) from the Meta-Analyses of Glucose and Insulin-related traits Consortium (MAGIC) [26]. Furthermore, we incorporated data on four iron homeostasis biomarkers, including ferritin (standard deviation (SD), n = 246,139), serum iron (SD, n = 163,511), total iron-binding capacity (TIBC) (SD, n = 135,430), and TSAT (SD, n = 131,471) from a recent GWAS meta-analysis [27], since these biomarkers represent different aspects of iron homeostasis including iron stores (serum ferritin) and iron transport (serum iron, TIBC, TSAT) (Additional file 1: Tables S2). As part of post hoc analysis, we also included MRI-derived liver iron content data from the UK Biobank (UKB; SD, n = 32,858) [28]. To mitigate potential bias stemming from horizontal pleiotropy and distinguish the effects of iron in general from hereditary hemochromatosis, we implemented a strategy to exclude genetic instruments in ABO and associated with HFE-hemochromatosis (specifically rs1800562 and rs1799945, if any) [20, 29]. Details of data sources and genetic instruments were listed in Additional file 1: Tables S3–S8.
Genetic associations for liver steatosis, liver cirrhosis, and related biomarkers as outcomes
Genetic associations of liver diseases were obtained from relevant GWAS involving individuals of European descent [30]. These studies included liver steatosis, which involved 9491 cases and 876,210 controls, and liver cirrhosis, which encompassed 4809 cases and 967,898 controls [30]. Biomarkers of liver diseases included ALT (SD, n = 344,136) [31], and PDFF (SD, n = 36,116) from UKB (Additional file 1: Table S9) [30]. For outcomes where the standard errors were not provided by authors (i.e., liver steatosis, liver cirrhosis, and PDFF), we calculated these using the corresponding betas and P values based on normal distribution.
Details of these GWAS, including the mean age, sex distribution, and covariates adjustment, are described in Additional file 1: Table S3 and Additional file 2: Supplementary Note. Information pertaining to potential participant overlap across the GWAS is listed in Additional file 1: Table S4. These data were harmonized based on the reported effect allele and effect allele frequencies (EAF). Palindromic variants with intermediate EAF (42% < EAF < 58%) were discarded to avoid ambiguity in the strand direction.
Statistical analyses
Main analyses
We calculated the variance (R2) of the exposures explained by genetic instruments and approximated the overall F-statistic. An F-statistic > 10 suggests that weak instrument bias is unlikely [32]. We used inverse-variance weighting (IVW) with multiplicative random effects as the main analysis, which assumes balanced pleiotropy. We evaluated heterogeneity of the variant-specific Wald ratio through I2 where a high value may imply the presence of invalid instruments [33]. We performed random-effects meta-analysis for liver steatosis and liver cirrhosis as the corresponding GWAS did not provide pooled estimates across the studies for these outcomes [30]. Considering the interpretation of associations with liability to T2D (per log odds), we used a unit of per doubling in the prevalence instead by multiplying the estimates by 0.693 to improve the interpretation [34].
Sensitivity analyses
Various sensitivity analyses based on different assumptions were used to assess the robustness of the main findings. These included MR-Egger (assuming instrument strength independent of direct effect, InSIDE), weighted median (WM, assuming that the majority of selected instruments are valid), and Mendelian Randomization Pleiotropy RESidual Sum and Outlier (MR-PRESSO, outlier-robust) [35]. We also performed random-effects meta-analysis of MR-PRESSO (if available) for liver steatosis and liver cirrhosis as sensitivity analyses. The P value of MR-Egger intercept was used to assess overall horizontal pleiotropy. I2GX statistic was used to detect the possibility of dilution bias of MR-Egger estimates [36]. Lastly, given that HbA1c is influenced by both glycemic and erythrocytic properties, we repeated the analyses regarding HbA1c by classifying the instruments according to the reported clusters (glycemia: 16 SNPs; erythrocytic: 44 SNPs (iron: 3 SNPs; mature red blood cell: 23 SNPs; reticulocyte: 18 SNPs); and unknown: 13 SNPs) based on the original GWAS (see footnote of Additional file 1: Table S10 for details on the classification) [26].
Mediation analyses
Multivariable MR (MVMR) analyses were also used to assess the observed association of mediators on outcomes, adjusting for exposures to control for horizontal pleiotropy [37]. The same method was also used for mediation analysis to assess the associations of exposures on outcomes adjusting for mediators to decompose the total effect into direct and indirect effects. The mediation effects were calculated by the product of coefficients methods, with the standard error derived using the delta method [38, 39].
Power calculation
Based on the significance level of 0.05 and R2 of T2D, glycemic traits, iron homeostasis biomarkers, and liver iron (exposures) explained by the genetic instruments in the analyses, we estimated the effect sizes and odds ratios which we could detect at least 80% study power for the associations with iron homeostasis biomarkers, liver diseases, and related biomarkers (https://sb452.shinyapps.io/power/) [40], and are listed in Additional file 1: Table S11.
Details of these methods can be found in Additional file 2: Supplementary Note.
All analyses were performed using R version 4.2.2 with R packages (“TwoSampleMR” version 0.5.6 [41], “MRPRESSO” version 1.0, “forestplot” version 3.1.1, and “msm” version 1.7).
Results
Genetic instruments of liability to T2D, glycemic traits, iron homeostasis biomarkers, and liver iron
In this study, we included up to 180 SNPs for liability to T2D (F-statistic: 246, R2: 4.5%), 66 SNPs for FG (F-statistic: 154, R2: 4.6%), 37 SNPs for FI (F-statistic: 58, R2: 1.3%), 13 SNPs for 2hGlu (F-statistic: 63, R2: 1.3%), 72 SNPs for HbA1c (F-statistic: 118, R2: 5.4%), 59 SNPs for ferritin (F-statistic: 77, R2: 1.8%), 27 SNPs for serum iron (F-statistic: 161, R2: 2.6%), 31 SNPs for TIBC (F-statistic: 162, R2:3.6%), 26 SNPs for TSAT (F-statistic: 188, R2: 3.6%), and six SNPs for liver iron (F-statistic: 299, R2: 5.2%). The F-statistics indicated low evidence of weak instrument bias (Additional file 1: Table S5).
Associations of liability to T2D and glycemic traits in liver steatosis, liver cirrhosis, ALT, and PDFF
Liability to T2D and impaired FI likely increased risk of liver steatosis (odds ratio (OR)liability to T2D: 1.14 per doubling in the prevalence of T2D, 95% confidence interval (95% CI): 1.10 to 1.19); ORFI: 3.31 per log pmol/l, 95% CI: 1.92 to 5.72) as well as ALT and PDFF (Fig. 2). Liability to T2D also increased liver cirrhosis (ORliability to T2D: 1.07 per doubling in the prevalence, 95% CI: 1.03 to 1.12) (Fig. 2). These results were consistent with meta-analyses using MR-PRESSO estimates which were corrected for potential outliers (Additional file 3: Fig. S1). Although there were signs of heterogeneity based on I2 (e.g., ALT and PDFF), horizontal pleiotropy was not evident in most analyses and findings were generally consistent across sensitivity analyses (Fig. 2; Additional file 1: Table S12). However, the inverse association of HbA1c in liver steatosis (OR: 0.70 per %, 95% CI: 0.51 to 0.96) is likely driven by erythrocytic variants, which was also suggested by the HbA1c signal classification analyses (Additional file 3: Fig. S2; Additional file 1: Table S13). Based on MR-PRESSO analyses, scatter plots, and the forest plot, the inverse association of FG with liver steatosis was likely driven by rs1260326 (GCKR) in UKB (Additional file 1: Table S6; Additional file 3: Fig. S1, S3).

The associations of liability to type 2 diabetes and glycemic traits in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction (MRI) using meta-analysis of inverse variance weighting. A The associations of liability of type 2 diabetes and glycemic traits in liver steatosis and liver cirrhosis. B The associations of type 2 diabetes and glycemic traits in alanine aminotransferase, and proton density fat fraction. Liver steatosis, log odds (n = 885,701 including 9491 cases and 876,210 controls) included UKB (5921 cases), deCODE (785 cases), FinnGen (651 cases), and INTERMOUNTAIN (2134 cases). Liver cirrhosis, log odds (n = 972,707 including 4809 cases and 967,898 controls) included UKB (2301 cases), deCODE (691 cases), FinnGen (1425 cases), and INTERMOUNTAIN (392 cases). Liability to T2D, liability to type 2 diabetes; FG, fasting glucose; FI, fasting insulin; 2hGlu, 2-hour glucose; HbA1c, hemoglobin A1c; ALT, alanine aminotransferase; PDFF, proton density fat fraction; No. of SNPs, number of single nucleotide polymorphisms; IVW, inverse-variance weighting; I2, degree of heterogeneity; 95% CI, 95% confidence interval. *P value < 0.05, **P value < 0.01, ***P value < 0.001
Association of iron homeostasis biomarkers in liver steatosis, liver cirrhosis, ALT, and PDFF
Ferritin was positively associated with liver steatosis (OR: 1.25 per SD, 95% CI: 1.07 to 1.46), with consistent findings from sensitivity analyses (Fig. 3; Additional file 3: Fig. S4; Additional file 1: Table S14). Serum iron was positively associated with liver cirrhosis (OR: 1.31 per SD, 95% CI: 1.06 to 1.63), ALT, and PDFF (Fig. 3). TSAT was positively associated with PDFF with directionally consistent estimates with other liver markers (Fig. 3; Additional file 1: Table S14). Heterogeneity was high across the analyses with variations in estimates across sensitivity analyses. Notably, the association concerning serum iron was attenuated in the MR-PRESSO analyses, likely driven by rs144861591 (ZFP57) (Additional file 3: Fig. S4; Additional file 1: Tables S7 and S14).

The associations of iron homeostasis biomarkers (n ≤ 246,139) in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction using meta-analysis of inverse-variance weighting. A The associations of iron homeostasis biomarkers in liver steatosis and liver cirrhosis. B The associations of iron homeostasis biomarkers in alanine aminotransferase, and proton density fat fraction. Liver steatosis, log odds (n = 885,701 including 9491 cases and 876,210 controls) included UKB (5921 cases), deCODE (785 cases), FinnGen (651 cases), and INTERMOUNTAIN (2134 cases). Liver cirrhosis, log odds (n = 972,707 including 4809 cases and 967,898 controls) included UKB (2301 cases), deCODE (691 cases), FinnGen (1425 cases), and INTERMOUNTAIN (392 cases). TIBC, total iron-binding capacity; TSAT, transferrin saturation; ALT, alanine aminotransferase; PDFF, proton density fat fraction; No. of SNPs, number of single nucleotide polymorphisms; IVW, inverse-variance weighting; I2, degree of heterogeneity; 95% CI, 95% confidence interval. *P value < 0.05, **P value < 0.01, ***P value < 0.001
Post hoc analyses concerning liver iron
Ferritin, serum iron, and TSAT were positively associated with liver iron, and TIBC negatively with liver iron (Fig. 4A). Increased liver iron was associated with higher risk of liver steatosis (OR: 1.15 per SD, 95% CI: 1.05 to 1.26), liver cirrhosis (OR: 1.34 per SD, 95% CI: 1.07 to 1.68), ALT, and PDFF (Fig. 4B–C; Additional file 1: Tables S15–S16). However, liability to T2D or glycemic traits was not associated with liver iron, except for the inverse association of HbA1c in liver iron (β: -0.34 per percentage, 95% CI: − 0.56 to − 0.12) (Additional file 3: Fig. S5). However, the inverse association of HbA1c and liver steatosis via ferritin and liver iron was likely driven by erythrocytic property (Fig. 5; Additional file 3: Fig. S2; Additional file 1: Tables S13, S17).

The associations of iron homeostasis biomarkers (blood-based) (n ≤ 246,139) in liver iron content (MRI) (n = 32,858, UKB) and the association of liver iron in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction using Mendelian randomization. A The associations of iron homeostasis biomarkers in liver iron (MRI). B The associations of liver iron in liver steatosis and liver cirrhosis. C The associations of liver iron in alanine aminotransferase and proton density fat fraction alanine aminotransferase, and proton density fat fraction. Liver steatosis, log odds (n = 885,701 including 9491 cases and 876,210 controls) included UKB (5921 cases), deCODE (785 cases), FinnGen (651 cases), and INTERMOUNTAIN (2134 cases). Liver cirrhosis, log odds (n = 972,707 including 4809 cases and 967,898 controls) included UKB (2301 cases), deCODE (691 cases), FinnGen (1425 cases), and INTERMOUNTAIN (392 cases). TIBC, total iron-binding capacity; TSAT, transferrin saturation; ALT, alanine aminotransferase; PDFF, proton density fat fraction; No. of SNPs, number of single nucleotide polymorphisms; IVW, inverse-variance weighting; I2, degree of heterogeneity; 95% CI, 95% confidence interval. *P value < 0.05, **P value < 0.01, ***P value < 0.001

Summary of associations of HbA1c and its signal classification in ferritin, liver iron content (MRI), and liver steatosis (meta-analysis). HbA1c, hemoglobin A1c; Glycemic, probability of the variant in glycemic class, included fasting insulin, 2-hour glucose, and fasting glucose; Reticulocyte, probability of the variant in reticulocyte class, included reticulocyte count, reticulocyte fraction of red cells, immature fraction of reticulocytes, high light scatter reticulocyte count, and high light scatter reticulocyte percentage of red cells; Mature RBC, probability of the variant in mature red blood cell class, included red blood cell count, mean corpuscular volume, hematocrit, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, hemoglobin concentration, and red cell distribution width; Iron, probability of the variant in iron class, included ferritin, transferrin, serum iron, and transferrin saturation; OR, odds ratio; 95% CI, 95% confidence interval
Findings from the mediation analyses
Given that our previous study indicated FI was associated with ferritin (Step 1, Additional file 1: Table S1), and FI and ferritin were associated with liver steatosis (Figs. 2A and 3B), we subsequently assessed mediation using a two-step MR design. The corresponding mediation analysis showed ferritin partially mediated the association of FI in liver steatosis (Fig. 6A; Additional file 3: Fig. S6), where the proportion mediated via ferritin was 7% (95% CI: 2% to 12%) (Fig. 6B). As post hoc analysis, we also assessed the potential mediating role of liver iron between ferritin and liver steatosis and found evidence of partial mediation (proportion mediated: 43%, 95% CI: 38% to 48%) (Fig. 6B).

The associations of fasting insulin in liver steatosis with and without adjustment for ferritin, and summary of the mediating roles of ferritin and liver iron content in the associations of fasting insulin and liver steatosis (meta-analysis). A The associations of fasting insulin in liver steatosis with and without adjustment for ferritin using univariable and multivariable Mendelian randomization analysis. B Summary of the direct effects, indirect effects, and mediation effects of ferritin in the association of fasting insulin and steatosis, and of liver iron in the association of ferritin and steatosis (meta-analysis). FI, fasting insulin; HbA1c, hemoglobin A1c; MVMR, multivariable Mendelian randomization; IVW, inverse-variance weighting; OR, odds ratio; 95% CI, 95% confidence interval. *P value < 0.05, **P value < 0.01, ***P value < 0.001
Discussion
This is the largest MR study to investigate the associations of liability to T2D and glycemic traits in liver steatosis and liver cirrhosis. Our study is supported by previous observational and MR studies [4, 13,14,15, 42, 43] that genetic liability to T2D and FI likely increases the risk of liver steatosis and related biomarkers (e.g., ALT and PDFF), while liability to T2D appears to also increase the risk of liver cirrhosis. Our study also adds by showing, for the first time, iron homeostasis biomarkers and liver-iron content could potentially mediate some of these associations, hence providing genetic evidence that targeting the reduction of ferritin may mitigate the impact of MASLD risk arising from elevated insulin. Based on this study, we also clarify the paradoxical inverse association of HbA1c in liver steatosis is likely driven by the erythrocytic property of HbA1c and informs caution should be applied when using HbA1c values for disease management.
Previous investigations, including MR studies, have demonstrated that T2D and FI are associated with a higher MASLD risk, as well as increased levels of hepatic fat and ALT [4, 13,14,15, 42, 43], suggesting that the associations are likely causal. Similar findings were observed when extended to liver cirrhosis, an advanced stage of steatosis. The underlying mechanisms may include increased insulin resistance, proxied by elevated FI, leading to elevated free fatty acids uptake by liver, and hence increase de novo lipogenesis, lipolysis of dysfunctional adipose tissue, and subsequent risk of hepatic steatosis [3]. These findings imply that modifying risk factors of T2D and insulin resistance might protect against liver steatosis and its progression to liver cirrhosis.
Among the four iron homeostasis biomarkers considered, ferritin was most likely relevant to the development of liver steatosis, whilst serum iron likely increased ALT, a surrogate marker of MASLD. These findings are supported by previous observational and MR studies [9, 16, 17, 44, 45]. Increased ferritin may induce the deposition of hepatic iron in liver macrophages and stellate cells which results in hepatocellular damage and steatosis [6]. Although previous studies speculated that ferritin-MASLD association could be confounded by inflammation, this is not supported by clinical trials and mouse models [46, 47], and hence unlikely explains our findings, especially since MR design is more resistant to confounding compared to conventional observational studies [48]. Elevated serum iron levels may also promote oxidative stress through the production of free radicals in the liver through presence of excess iron [44]. Also, iron overload and high-iron diet are related to ferroptosis, an iron-dependent form of cell death caused by lipid peroxidation, which is a phenomenon related to MASLD progression [49, 50]. Hence, focusing on anti-ferroptosis therapy may help reverse the detrimental effects of elevated insulin in MASLD mediated via ferroptosis, such as Malic Enzyme 1 (ME1) [51], although this requires further investigations using population-based studies. Furthermore, our mediation analysis adds to this hypothesis by suggesting that ferritin partially mediates the association of FI in liver steatosis and hence could be a possible target of intervention amongst people with T2D. However, the positive associations of serum iron in liver cirrhosis and PDFF were attenuated after removing rs144861591 which was strongly related to hereditary hemochromatosis (rs1800562 in HFE, r2 = 0.98) [41]. Although the MR-Egger intercept analyses suggested weak evidence of bias from horizontal pleiotropy, we are uncertain whether these associations are entirely independent of hemochromatosis, which warrants further research. Furthermore, the estimates were directionally consistent across different cohorts, which required verification when larger MASLD GWAS become available.
From our post hoc analyses, iron homeostasis biomarkers associated with liver iron were associated with a higher risk of MASLD and its biomarkers, and liver iron partially mediated the association of ferritin and liver steatosis, which is consistent with previous observational and MR studies [19, 52, 53]. The strong correlation between iron homeostasis biomarkers and liver iron is somewhat expected given that the liver is the main organ for iron storage [54]. Although studies on liver iron and MASLD are generally fewer compared to those of iron homeostasis biomarkers, evidence exists to suggest liver iron content is a strong predictor of liver disease severity [55], with possible pathways including increased inflammation, oxidative stress, and lipid oxidation [56]. Although we did not observe any association of genetic liability to T2D or glycemic traits in liver iron (apart from HbA1c which is likely driven by iron and mature red blood cells, i.e., iron recycling from senescent red blood cells) [21], which is consistent with a recent longitudinal study [42]. However, additional studies should explore whether sex-specific effects exist, as suggested by a previous longitudinal investigation [42].
The inverse association of FG in liver steatosis was inconsistent with the findings of liability to T2D, insulin, and glycemic signal class of HbA1c. Although these glycemic traits are well-characterized risk factors for T2D, recent studies indicated that they may have distinct relations with metabolomic signatures and CVD risk [57, 58]. However, our sensitivity analysis suggested this paradoxical finding could be driven by rs1260326 (GCKR), which encodes glucokinase regulatory protein (GKRP) (a primarily liver-specific protein) and is linked to decreased blood glucose by altering GKRP’s function of suppressing glucokinase activity and enhancing hepatic glycolysis, and concurrently increase total hepatic triglycerides [59], resulting in the risk allele of rs1260326 (T) decreasing the risk of T2D but increasing liver steatosis [30], and impacting the overall analyses. Although any effects observed for this GCKR risk variant could be arguably the downstream effects of glucose level variation (i.e., vertical pleiotropy and hence valid), given its high relevance in glucose metabolism, the increased circulating lipids could also arguably as an effect independent of glucose (i.e., horizontal pleiotropy and hence invalid) [60, 61]. As the overall inverse association is also most apparent in UKB, this may also be a reflection of selection bias distorting genetic associations in this cohort [62]. Given these issues, the paradoxical findings of glucose with liver steatosis should be interpreted with caution. Similarly, the unexpected inverse association of HbA1c and liver steatosis, although observed in a previous study [63], may be a reflection of the complex properties of HbA1c. The level of HbA1c is impacted by hyperglycemia and iron deficiency, where an earlier study showed iron deficiency can shift the distribution of HbA1c [64]. This is supported by the lack of association of HbA1c with liver steatosis when we restricted our analyses to only glycemia-related instruments (Fig. 5) and is more consistent with the glucose analyses without GCKR variant. These findings illustrated the challenges of using HbA1c solely as a reflection of hyperglycemia and are largely relevant to diabetes management.
Despite using an MR study design which is less prone to confounding than conventional observational studies, limitations remain. First, MR has stringent, unverifiable assumptions. Whilst confounding is less likely given genetics are randomly allocated at conception and with the use of GWAS which controls for population stratification, the possibility of exclusion restriction violation cannot be ruled out completely. Nevertheless, we applied different statistical methods with various assumptions that yielded comparable results. We also did not include variants which could be highly pleiotropic, such as the variants harboring HFE-hemochromatosis (rs1800562/rs1799945) and ABO gene region as per our previous study [21]. Although there was participant overlap in some of the GWAS used, which may bias our estimates towards the observational association in instances where weak instruments are used, this is unlikely given the large F-statistics used. Second, although we mainly focused on the overall analyses which combined different cohorts to maximize statistical power, these individual cohorts did not always give consistent associations (e.g., the associations of FI in liver cirrhosis, Fig. 2A). These discrepancies may have been driven by multiple factors such as participant selection criteria, or statistical power, and variation in genetic architecture in the underlying populations [65]. Third, we did not explore potential sex differences given the lack of sex-specific GWAS summary statistics whereas early studies suggested possible sex-specific associations of glycemic traits in liver iron and liver fat [42]. Additional studies with individual-level data (e.g., UK Biobank) might be helpful to address this limitation. Fourth, the effect sizes of liver steatosis and cirrhosis may be underestimated as the case definition in the MASLD GWAS is only based on ICD-10 diagnostic codes and hence may have missed patients who were symptomless and hence not being diagnosed in clinical settings [30], and UKB participants are more healthy and hence are vulnerable to selection bias [66]. However, our findings were directionally consistent with previous MR studies of liability to T2D, glycemic traits, and iron biomarkers in MASLD using GWAS with liver biopsy specimens definition [14, 16, 17, 19, 67]. Lastly, we only included studies of European ancestry. Whether the observed associations extend to other ethnic groups warrants further investigation.
Conclusions
Our study highlights the role of T2D and FI in liver steatosis and cirrhosis etiology, and provides mechanistic insights into the mediating role of ferritin in the positive association of FI with liver steatosis. Whether targeting the reduction of ferritin in those with increased insulin would reduce the risk of MASLD requires further bespoke investigation.
Availability of data and materials
All data used in this study can be found in the cited references and the URLs in the Acknowledgements and Supplementary Materials.
Abbreviations
- 2hGlu:
-
2-hour glucose
- ALT:
-
Alanine aminotransferase
- DIAMANTE:
-
Diabetes Meta-Analysis of Trans-Ethics associations studies
- EAF:
-
Effect allele frequencies
- FG:
-
Fasting glucose
- FI:
-
Fasting insulin
- GWAS:
-
Genome-wide association studies
- HbA1c:
-
Hemoglobin A1c
- InSIDE:
-
Instrument Strength Independent of Direct Effect
- IVW:
-
Inverse-variance weighting
- LD:
-
Linkage disequilibrium
- MAGIC:
-
Meta-Analyses of Glucose and Insulin-related traits Consortium
- MASLD:
-
Metabolic dysfunction-associated steatotic liver disease
- MR:
-
Mendelian randomization
- MR-PRESSO:
-
Mendelian Randomization Pleiotropy RESidual Sum and Outlier
- MVMR:
-
Multivariable Mendelian randomization
- PDFF:
-
Proton density fat fraction
- R2 :
-
Variance of the exposures explained by genetic instruments
- SD:
-
Standard deviation
- SNPs:
-
Single nucleotide polymorphisms
- STROBE-MR:
-
Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian randomization
- T2D:
-
Type 2 diabetes
- TIBC:
-
Total iron-binding capacity
- TSAT:
-
Transferrin saturation
- UKB:
-
UK Biobank
- WM:
-
Weighted median
References
Rinella ME, Lazarus JV, Ratziu V, Francque SM, Sanyal AJ, Kanwal F, Romero D, Abdelmalek MF, Anstee QM, Arab JP, et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. Hepatology. 2023;78(6):1966–86.
Riazi K, Azhari H, Charette JH, Underwood FE, King JA, Afshar EE, Swain MG, Congly SE, Kaplan GG, Shaheen AA. The prevalence and incidence of NAFLD worldwide: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2022;7(9):851–61.
Brunt EM, Wong VW, Nobili V, Day CP, Sookoian S, Maher JJ, Bugianesi E, Sirlin CB, Neuschwander-Tetri BA, Rinella ME. Nonalcoholic fatty liver disease. Nat Rev Dis Primers. 2015;1:15080.
Younossi ZM, Golabi P, de Avila L, Paik JM, Srishord M, Fukui N, Qiu Y, Burns L, Afendy A, Nader F. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J Hepatol. 2019;71(4):793–801.
Mantovani A, Petracca G, Beatrice G, Tilg H, Byrne CD, Targher G. Non-alcoholic fatty liver disease and risk of incident diabetes mellitus: an updated meta-analysis of 501 022 adult individuals. Gut. 2021;70(5):962–9.
Valenti L, Corradini E, Adams LA, Aigner E, Alqahtani S, Arrese M, Bardou-Jacquet E, Bugianesi E, Fernandez-Real JM, Girelli D, et al. Consensus Statement on the definition and classification of metabolic hyperferritinaemia. Nat Rev Endocrinol. 2023;19(5):299–310.
Liu J, Li Q, Yang Y, Ma L. Iron metabolism and type 2 diabetes mellitus: A meta-analysis and systematic review. J Diabetes Investig. 2020;11(4):946–55.
Kowdley KV, Belt P, Wilson LA, Yeh MM, Neuschwander-Tetri BA, Chalasani N, Sanyal AJ, Nelson JE. Serum ferritin is an independent predictor of histologic severity and advanced fibrosis in patients with nonalcoholic fatty liver disease. Hepatology. 2012;55(1):77–85.
Brudevold R, Hole T, Hammerstrøm J. Hyperferritinemia is associated with insulin resistance and fatty liver in patients without iron overload. PLoS ONE. 2008;3(10):e3547.
Ryan JD, Armitage AE, Cobbold JF, Banerjee R, Borsani O, Dongiovanni P, Neubauer S, Morovat R, Wang LM, Pasricha SR et al: Hepatic iron is the major determinant of serum ferritin in NAFLD patients. Liver Int 2018, 38(1):164-173.
Davies NM, Holmes MV, Smith GD. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;362:k601.
Voight BF, Peloso GM, Orho-Melander M, Frikke-Schmidt R, Barbalic M, Jensen MK, Hindy G, Hólm H, Ding EL, Johnson T, et al. Plasma HDL cholesterol and risk of myocardial infarction: a mendelian randomisation study. Lancet. 2012;380(9841):572–80.
De Silva NMG, Borges MC, Hingorani AD, Engmann J, Shah T, Zhang X, Luan J, Langenberg C, Wong A, Kuh D, et al. Liver function and risk of type 2 diabetes: bidirectional mendelian randomization study. Diabetes. 2019;68(8):1681–91.
Yuan S, Chen J, Li X, Fan R, Arsenault B, Gill D, Giovannucci EL, Zheng JS, Larsson SC. Lifestyle and metabolic factors for nonalcoholic fatty liver disease: Mendelian randomization study. Eur J Epidemiol. 2022;37(7):723–33.
Liu Z, Zhang Y, Graham S, Wang X, Cai D, Huang M, Pique-Regi R, Dong XC, Chen YE, Willer C, et al. Causal relationships between NAFLD, T2D and obesity have implications for disease subphenotyping. J Hepatol. 2020;73(2):263–76.
Liu C, Chen Y, Zhang Z, Xie J, Yu C, Xu L, Li Y. Iron status and NAFLD among European populations: a bidirectional two-sample mendelian randomization study. Nutrients. 2022;14(24):5324.
Wang K, Yang F, Zhang P, Yang Y, Jiang L. Genetic effects of iron levels on liver injury and risk of liver diseases: A two-sample Mendelian randomization analysis. Front Nutr. 2022;9:964163.
Xie J, Huang H, Liu Z, Li Y, Yu C, Xu L, Xu C. The associations between modifiable risk factors and nonalcoholic fatty liver disease: A comprehensive Mendelian randomization study. Hepatology. 2023;77(3):949–64.
He H, Liao S, Zeng Y, Liang L, Chen J, Tao C. Causal relationships between metabolic-associated fatty liver disease and iron status: Two-sample Mendelian randomization. Liver Int. 2022;42(12):2759–68.
Pilling LC, Atkins JL, Melzer D. Genetic modifiers of penetrance to liver endpoints in HFE hemochromatosis: associations in a large community cohort. Hepatology. 2022;76:1735–45.
Liang Y, Luo S, Wong THT, He B, Schooling CM, Au Yeung SL. Association of iron homeostasis biomarkers in type 2 diabetes and glycaemic traits: a bidirectional two-sample Mendelian randomization study. Int J Epidemiol. 2023;52(6):1914–25.
Wong VW, Adams LA, de Lédinghen V, Wong GL, Sookoian S. Noninvasive biomarkers in NAFLD and NASH - current progress and future promise. Nat Rev Gastroenterol Hepatol. 2018;15(8):461–78.
Sanderson E, Glymour MM, Holmes MV, Kang H, Morrison J, Munafò MR, Palmer T, Schooling CM, Wallace C, Zhao Q, et al. Mendelian randomization. Nat Rev Methods Primers. 2022;2(1):6.
Skrivankova VW, Richmond RC, Woolf BAR, Davies NM, Swanson SA, VanderWeele TJ, Timpson NJ, Higgins JPT, Dimou N, Langenberg C, et al. Strengthening the reporting of observational studies in epidemiology using mendelian randomisation (STROBE-MR): explanation and elaboration. BMJ. 2021;375:n2233.
Mahajan A, Spracklen CN, Zhang W, Ng MCY, Petty LE, Kitajima H, Yu GZ, Rüeger S, Speidel L, Kim YJ, et al. Multi-ancestry genetic study of type 2 diabetes highlights the power of diverse populations for discovery and translation. Nat Genet. 2022;54(5):560–72.
Chen J, Spracklen CN, Marenne G, Varshney A, Corbin LJ, Luan J, Willems SM, Wu Y, Zhang X, Horikoshi M, et al. The trans-ancestral genomic architecture of glycemic traits. Nat Genet. 2021;53(6):840–60.
Bell S, Rigas AS, Magnusson MK, Ferkingstad E, Allara E, Bjornsdottir G, Ramond A, Sørensen E, Halldorsson GH, Paul DS, et al. A genome-wide meta-analysis yields 46 new loci associating with biomarkers of iron homeostasis. Commun Biol. 2021;4(1):156.
Liu Y, Basty N, Whitcher B, Bell JD, Sorokin EP, van Bruggen N, Thomas EL, Cule M. Genetic architecture of 11 organ traits derived from abdominal MRI using deep learning. Elife. 2021;10:e65554.
Li S, Schooling C. A phenome-wide association study of ABO blood groups. BMC Med. 2020;18(1):1–11.
Sveinbjornsson G, Ulfarsson MO, Thorolfsdottir RB, Jonsson BA, Einarsson E, Gunnlaugsson G, Rognvaldsson S, Arnar DO, Baldvinsson M, Bjarnason RG, et al. Multiomics study of nonalcoholic fatty liver disease. Nat Genet. 2022;54(11):1652–63.
Elsworth B, Lyon M, Alexander T, Liu Y, Matthews P, Hallett J, Bates P, Palmer T, Haberland V, Smith GD et al: The MRC IEU OpenGWAS data infrastructure. bioRxiv. 2020.08.10.244293
Ben Elsworth, Matthew Lyon, Tessa Alexander, Yi Liu, Peter Matthews, Jon Hallett, Phil Bates, Tom Palmer, Valeriia Haberland, George Davey Smith, Jie Zheng, Philip Haycock, Tom R Gaunt, Gibran Hemani bioRxiv. 2020;08(10):244293. https://doi.org/10.1101/2020.08.10.244293.
Bowden J, Hemani G, Davey Smith G. Invited commentary: detecting individual and global horizontal pleiotropy in Mendelian randomization-a job for the humble heterogeneity statistic? Am J Epidemiol. 2018;187(12):2681–5.
Burgess S, Labrecque JA. Mendelian randomization with a binary exposure variable: interpretation and presentation of causal estimates. Eur J Epidemiol. 2018;33(10):947–52.
Slob EAW, Burgess S. A comparison of robust Mendelian randomization methods using summary data. Genet Epidemiol. 2020;44(4):313–29.
Bowden J, Del Greco MF, Minelli C, Davey Smith G, Sheehan NA, Thompson JR. Assessing the suitability of summary data for two-sample Mendelian randomization analyses using MR-Egger regression: the role of the I2 statistic. Int J Epidemiol. 2016;45(6):1961–74.
Burgess S, Thompson SG. Multivariable Mendelian randomization: the use of pleiotropic genetic variants to estimate causal effects. Am J Epidemiol. 2015;181(4):251–60.
Carter AR, Gill D, Davies NM, Taylor AE, Tillmann T, Vaucher J, Wootton RE, Munafò MR, Hemani G, Malik R, et al. Understanding the consequences of education inequality on cardiovascular disease: mendelian randomisation study. BMJ. 2019;365:l1855.
Tofighi D, MacKinnon DP. RMediation: an R package for mediation analysis confidence intervals. Behav Res Methods. 2011;43(3):692–700.
Burgess S. Sample size and power calculations in Mendelian randomization with a single instrumental variable and a binary outcome. Int J Epidemiol. 2014;43(3):922–9.
Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, Laurin C, Burgess S, Bowden J, Langdon R, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018;7:e34408.
Niedermayer F, Su Y, von Krüchten R, Thorand B, Peters A, Rathmann W, Roden M, Schlett CL, Bamberg F, Nattenmüller J, et al. Trajectories of glycaemic traits exhibit sex-specific associations with hepatic iron and fat content: Results from the KORA-MRI study. Liver Int. 2023;43:2153–66.
Bugianesi E, Gastaldelli A, Vanni E, Gambino R, Cassader M, Baldi S, Ponti V, Pagano G, Ferrannini E, Rizzetto M. Insulin resistance in non-diabetic patients with non-alcoholic fatty liver disease: sites and mechanisms. Diabetologia. 2005;48(4):634–42.
Ruhl CE, Everhart JE. Relation of elevated serum alanine aminotransferase activity with iron and antioxidant levels in the United States. Gastroenterology. 2003;124(7):1821–9.
He A, Zhou Z, Huang L, Yip KC, Chen J, Yan R, Li R. Association between serum iron and liver transaminases based on a large adult women population. J Health Popul Nutr. 2023;42(1):69.
Beaton MD, Chakrabarti S, Adams PC. Inflammation is not the cause of an elevated serum ferritin in non-alcoholic fatty liver disease. Ann Hepatol. 2014;13(3):353–6.
Altamura S, Müdder K, Schlotterer A, Fleming T, Heidenreich E, Qiu R, Hammes HP, Nawroth P, Muckenthaler MU. Iron aggravates hepatic insulin resistance in the absence of inflammation in a novel db/db mouse model with iron overload. Mol Metab. 2021;51:101235.
Smith GD, Lawlor DA, Harbord R, Timpson N, Day I, Ebrahim S. Clustered environments and randomized genes: a fundamental distinction between conventional and genetic epidemiology. PLoS Med. 2007;4(12):e352.
Wang H, An P, Xie E, Wu Q, Fang X, Gao H, Zhang Z, Li Y, Wang X, Zhang J, et al. Characterization of ferroptosis in murine models of hemochromatosis. Hepatology. 2017;66(2):449–65.
Wang S, Liu Z, Geng J, Li L, Feng X. An overview of ferroptosis in non-alcoholic fatty liver disease. Biomed Pharmacother. 2022;153:113374.
Fang X, Zhang J, Li Y, Song Y, Yu Y, Cai Z, Lian F, Yang J, Min J, Wang F. Malic enzyme 1 as a novel anti-ferroptotic regulator in hepatic ischemia/reperfusion injury. Adv Sci (Weinh). 2023;10(13):e2205436.
Buzzetti E, Petta S, Manuguerra R, Luong TV, Cabibi D, Corradini E, Craxì A, Pinzani M, Tsochatzis E, Pietrangelo A. Evaluating the association of serum ferritin and hepatic iron with disease severity in non-alcoholic fatty liver disease. Liver Int. 2019;39(7):1325–34.
Nelson JE, Wilson L, Brunt EM, Yeh MM, Kleiner DE, Unalp-Arida A, Kowdley KV. Relationship between the pattern of hepatic iron deposition and histological severity in nonalcoholic fatty liver disease. Hepatology. 2011;53(2):448–57.
Anderson GJ, Frazer DM. Hepatic iron metabolism. Semin Liver Dis. 2005;25(4):420–32.
Ganne-Carrié N, Christidis C, Chastang C, Ziol M, Chapel F, Imbert-Bismut F, Trinchet JC, Guettier C, Beaugrand M. Liver iron is predictive of death in alcoholic cirrhosis: a multivariate study of 229 consecutive patients with alcoholic and/or hepatitis C virus cirrhosis: a prospective follow up study. Gut. 2000;46(2):277–82.
Fernandez M, Lokan J, Leung C, Grigg A. A critical evaluation of the role of iron overload in fatty liver disease. J Gastroenterol Hepatol. 2022;37(10):1873–83.
Wong THT, Mo JMY, Zhou M, Zhao JV, Schooling CM, He B, Luo S, Au Yeung SL. A two-sample Mendelian randomization study explores metabolic profiling of different glycemic traits. Commun Biol. 2024;7(1):293.
Yuan S, Mason AM, Burgess S, Larsson SC. Differentiating associations of glycemic traits with atherosclerotic and thrombotic outcomes: mendelian randomization investigation. Diabetes. 2022;71(10):2222–32.
Singh C, Jin B, Shrestha N, Markhard AL, Panda A, Calvo SE, Deik A, Pan X, Zuckerman AL, Ben Saad A, et al. ChREBP is activated by reductive stress and mediates GCKR-associated metabolic traits. Cell Metab. 2024;36(1):144-158.e147.
Sliz E, Sebert S, Würtz P, Kangas AJ, Soininen P, Lehtimäki T, Kähönen M, Viikari J, Männikkö M, Ala-Korpela M, et al. NAFLD risk alleles in PNPLA3, TM6SF2, GCKR and LYPLAL1 show divergent metabolic effects. Hum Mol Genet. 2018;27(12):2214–23.
Hemani G, Bowden J, Davey Smith G. Evaluating the potential role of pleiotropy in Mendelian randomization studies. Hum Mol Genet. 2018;27(R2):R195-r208.
Schoeler T, Speed D, Porcu E, Pirastu N, Pingault JB, Kutalik Z. Participation bias in the UK Biobank distorts genetic associations and downstream analyses. Nat Hum Behav. 2023;7(7):1216–27.
Christman AL, Lazo M, Clark JM, Selvin E. Low glycated hemoglobin and liver disease in the U.S. population. Diabetes Care. 2011;34(12):2548–50.
Kim C, Bullard KM, Herman WH, Beckles GL. Association between iron deficiency and A1C Levels among adults without diabetes in the National Health and Nutrition Examination Survey, 1999–2006. Diabetes Care. 2010;33(4):780–5.
Lim ET, Würtz P, Havulinna AS, Palta P, Tukiainen T, Rehnström K, Esko T, Mägi R, Inouye M, Lappalainen T, et al. Distribution and medical impact of loss-of-function variants in the finnish founder population. PLoS Genet. 2014;10(7):e1004494.
Fry A, Littlejohns TJ, Sudlow C, Doherty N, Adamska L, Sprosen T, Collins R, Allen NE. Comparison of sociodemographic and health-related characteristics of UK Biobank participants with those of the general population. Am J Epidemiol. 2017;186(9):1026–34.
Anstee QM, Darlay R, Cockell S, Meroni M, Govaere O, Tiniakos D, Burt AD, Bedossa P, Palmer J, Liu YL, et al. Genome-wide association study of non-alcoholic fatty liver and steatohepatitis in a histologically characterised cohort☆. J Hepatol. 2020;73(3):505–15.
Acknowledgements
Data related to iron homeostasis were provided by S.Bell et al. in 2021. Information concerning liver steatosis, liver cirrhosis, and proton density fat fraction was contributed by G. Sveinbjornsson et al. in 2022. These datasets are available at https://www.decode.com/summarydata/. Additionally, data on type 2 diabetes were contributed by DIAMANTE investigators and can be accessed at http://diagram-consortium.org/. Summary data on fasting glucose, 2-hour glucose, fasting insulin, and hemoglobin A1c were provided by MAGIC investigators and are available at http://www.magicinvestigators.org/. For information on alanine aminotransferase, you can refer to UK Biobank (version 3, March 2018) and R. Mitchell et al.’s work in 2019, extracted from the MR-Base platform (https://gwas.mrcieu.ac.uk/). Lastly, data on liver iron were contributed by Y. Liu et al. in 2021 and can be found in the GWAS Catalog (Study accession: GCST90016674).
Funding
This study was partly funded by the Health and Medical Research Fund, Food and Health Bureau, HKSAR Government, Hong Kong, China (CFS-HKU1). The funder had no role in the design, analyses, interpretation of results, or writing of the paper.
Author information
Authors and Affiliations
Contributions
Y.L. and S.L.A.Y. designed the study. Y.L. wrote the analysis plan, performed the data analyses, and interpreted the results, with feedback from S.L.A.Y. S.L. and J.M.YM. cross-checked the results. Y.L. wrote the first draft of the manuscript with critical feedback and revisions from S.L.A.Y., S.L., S.B., J.M.Y.M., B.H., Y.Z., and X.B. All authors read and approved the final manuscript. Y.L. is the guarantor of work.
Authors’ Twitter handles
Twitter handles: @WingwingL88 (Ying Liang), @ShanLuo7 (Shan Luo), @AnnabelHE3 (Baoting He), @Ryan_Au_Yeung (Shiu Lun Au Yeung).
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Since this study relied on publicly available summary statistics from GWAS, ethics approval was not required. The respective ethics approvals for the GWAS were obtained by the relevant investigators.
Consent for publication
Not applicable.
Competing interests
SLAY received honoraria from SomaLogic for scientific presentations on proteomic studies that was unrelated to this study. The authors declare that they have no other competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
12916_2024_3486_MOESM1_ESM.xlsx
Additional file 1: Table S1. The associations of liability to type 2 diabetes and glycemic traits in iron homeostasis biomarkers in our previous study (Liang et al., 2023). Table S2. Description of iron homeostasis biomarkers. Table S3. Information of the data sources used in this study. Table S4. Participant overlap information in the genome-wide associations studies used in this study. Table S5. The genetic instruments information of type 2 diabetes, glycemic traits, iron homeostasis biomarkers, and liver iron after harmonization. Table S6. Genetic instruments information of liability to type 2 diabetes and glycemic traits as exposures. Table S7. Genetic instruments information of iron homeostasis biomarkers as exposures. Table S8. Genetic instruments information of liver iron as exposures. Table S9. Genetic instruments information of liver steatosis, liver cirrhosis, ALT, PDFF, and liver iron as outcomes. Table S10. Genetic information of HbA1c signal classification. Table S11. Study power of this two-step Mendelian randomization study. Table S12. The associations of liability to type 2 diabetes and glycemic traits in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction using Mendelian randomization analysis. Table S13. The associations of HbA1c signal classification in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction using Inverse-variance weighting. Table S14. The associations of iron homeostasis biomarkers in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction using Mendelian randomization analysis. Table S15. The associations of iron homeostasis biomarkers in liver iron using Mendelian randomization analysis. Table S16. The associations of liver iron in liver steatosis, liver cirrhosis, alanine aminotransferase, and proton density fat fraction using Mendelian randomization analysis. Table S17. The associations of HbA1c signal classification in iron homeostasis biomarkers and liver iron using Inverse-variance weighting.
12916_2024_3486_MOESM3_ESM.pdf
Additional file 3: Fig. S1 The associations of liability to type 2 diabetes and glycemic traits in liver steatosis and liver cirrhosis using meta-analysis of Inverse-variance weighting and MR-PRESSO (outlier-corrected). Fig. S2 The associations of HbA1c signal classifications in liver steatosis, ferritin, and liver iron using Mendelian randomization. Fig. S3 (a) Scatter plots and (b) forest plot of fasting glucose and liver steatosis in UK Biobank, deCODE, FinnGen, and INTERMOUNTAIN. Fig. S4 The associations of iron homeostasis biomarkers in liver steatosis and liver cirrhosis using meta-analysis of Inverse-variance weighting and MR-PRESSO (outlier-corrected). Fig. S5 The association of liability to type 2 diabetes and glycemic traits in liver iron (MRI) using Inverse-variance weighting. Fig. S6 The associations of fasting insulin, ferritin, and liver iron in liver steatosis with mediators and exposures adjustment using multivariable Mendelian randomization analysis.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liang, Y., Luo, S., Bell, S. et al. Do iron homeostasis biomarkers mediate the associations of liability to type 2 diabetes and glycemic traits in liver steatosis and cirrhosis: a two-step Mendelian randomization study. BMC Med 22, 270 (2024). https://doi.org/10.1186/s12916-024-03486-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12916-024-03486-w