
Abstract
Background
Due to the asymptomatic nature of the early stages, chronic kidney disease (CKD) is usually diagnosed at late stages and lacks targeted therapy, highlighting the need for new biomarkers to better understand its pathophysiology and to be used for early diagnosis and therapeutic targets. Given the close relationship between CKD and cardiovascular disease (CVD), we investigated the associations of 233 CVD- and inflammation-related plasma proteins with kidney function decline and aimed to assess whether the observed associations are causal.
Methods
We included 1140 participants, aged 55–74 years at baseline, from the Cooperative Health Research in the Region of Augsburg (KORA) cohort study, with a median follow-up time of 13.4 years and 2 follow-up visits. We measured 233 plasma proteins using a proximity extension assay at baseline. In the discovery analysis, linear regression models were used to estimate the associations of 233 proteins with the annual rate of change in creatinine-based estimated glomerular filtration rate (eGFRcr). We further investigated the association of eGFRcr-associated proteins with the annual rate of change in cystatin C-based eGFR (eGFRcys) and eGFRcr-based incident CKD. Two-sample Mendelian randomization was used to infer causality.
Results
In the fully adjusted model, 66 out of 233 proteins were inversely associated with the annual rate of change in eGFRcr, indicating that higher baseline protein levels were associated with faster eGFRcr decline. Among these 66 proteins, 21 proteins were associated with both the annual rate of change in eGFRcys and incident CKD. Mendelian randomization analyses on these 21 proteins suggest a potential causal association of higher tumor necrosis factor receptor superfamily member 11A (TNFRSF11A) level with eGFR decline.
Conclusions
We reported 21 proteins associated with kidney function decline and incident CKD and provided preliminary evidence suggesting a potential causal association between TNFRSF11A and kidney function decline. Further Mendelian randomization studies are needed to establish a conclusive causal association.
Similar content being viewed by others

Background
Chronic kidney disease (CKD), which is characterized by a progressive loss in kidney function over months or years [1], affected approximately 9.1% of the general population globally in 2017 [2]. Estimated glomerular filtration rate (eGFR), together with albuminuria and blood urea nitrogen, are the most commonly used indicators to evaluate kidney function and diagnose CKD in clinical practice. Kidney function declines with aging, while early stages of CKD remain asymptomatic, resulting in CKD usually being diagnosed in late stages. Besides, there is no targeted therapy for CKD beyond the management of its risk factors, such as diabetes and hypertension. The pathophysiology leading to CKD is not completely understood, and thus, there is a pressing need to identify new biomarkers that may provide new insight into the underlying pathophysiology of CKD, help in early diagnosis, and potentially be used as therapeutic targets.
It has been well documented that CKD is strongly related to cardiovascular diseases (CVD), and they share common mechanisms such as oxidative stress and inflammation [3, 4]. Thus, targeting CVD- and inflammation-related biomarkers may provide a valuable opportunity to identify biomarkers likely to be involved in the pathophysiology of CKD. Advances in proteomics technology, such as proximity extension assay technology using the Olink platform [5], make it possible to measure a large number of targeted biomarkers simultaneously.
An increasingly large number of studies investigating the associations of urinary and circulating proteomic biomarkers with kidney function and the progression of kidney disease have been published in the past decade [6,7,8,9,10,11,12,13,14,15,16]. We have previously identified and replicated 42 proteins associated with kidney function from a panel of inflammatory proteins, and revealed several pathophysiological pathways related to kidney disease using pathway analysis, highlighting the importance of investigating proteomics profiling in the general population [15]. A longitudinal study applying a large proteomics approach also identified novel biomarkers of progression to kidney failure in diabetic patients [16].
The present study aimed to investigate the associations of 233 CVD- and inflammation-related plasma proteins with longitudinal changes in kidney function in a community-based prospective cohort of middle-aged and older adults to uncover biomarkers and pathways involved in longitudinal kidney function decline and CKD development. Furthermore, we aimed to investigate whether the observed associations were potentially causal by using a two-sample Mendelian randomization (MR) design.
Methods
Study population
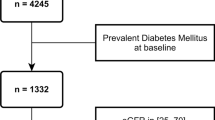
This study was based on the Cooperative Health Research in the Region of Augsburg (KORA) S4/F4/FF4 cohort study [17, 18]. A total of 4261 adults, aged 25–74 years, were included at baseline between 1999 and 2001 (S4) in Germany. Follow-up examinations were conducted after 7 years (F4) and after 14 years (FF4). The present analysis was restricted to participants aged 55–74 years at S4 (N = 1653). The flow chart of study participants is presented in Fig. 1. Participants without data on creatinine-based estimated glomerular filtration rate (eGFRcr, n = 19) or complete proteomics measurements (n = 68) at KORA S4 were excluded, leaving a total of 1566 participants at baseline. For the longitudinal analysis, participants without follow-up information on eGFRcr at both F4 and FF4 (n = 426) were further excluded. Finally, a total of 1140 participants were included, with a median follow-up time of 13.4 (25th percentile: 7.1, 75th percentile: 13.5) years (Fig. 1A). Among these 1140 participants, 638 participants were both followed up at F4 and FF4, 482 participants were only followed up at F4, and 20 participants were only followed up at FF4 (Fig. 1B). Participants with eGFRcr-based CKD at baseline (n = 54) were further excluded when investigating the associations of proteomic biomarkers with incident CKD. The KORA S4, F4, and FF4 studies were approved by the local ethical committee (Number: 99186) and all participants gave written informed consent.

Flowchart of study participants. Abbreviations: eGFRcr, creatinine-based estimated glomerular filtration rate; KORA S4/F4/FF4, the Cooperative Health Research in the Region of Augsburg (KORA) cohort study at baseline ( S4), first follow-up (F4) and second follow-up (FF4)
Measurement of proteomic biomarkers
Plasma samples of baseline participants aged 55–74 years were used to measure proteomic biomarkers. Three Olink panels, including the Proseek Multiplex CVD II, CVD III, and Inflammation panels (Olink, Upsala, Sweden), each covering 92 proteins, were measured using proximity extension assay technology [5]. The assay allows the relative quantification of analyte concentrations and was given as normalized protein expression values on a log2 scale, with higher expression values corresponding to higher protein levels. Details of the measurement process and exclusion of the proteins (e.g., proteins with more than 25% of all data below the limit of detection were excluded) have been reported elsewhere [19, 20]. A total of 233 proteins were included in this study (Additional file 1: Table S1). The Z-score transformation was conducted for all values of proteins in 1566 participants with complete proteomics measurements at baseline, which allows comparing the magnitude of the effect sizes across proteins and Z-score transformation was appropriate for most of our proteins after evaluating the distribution of each protein.
Measurement of kidney function and CKD
The primary outcomes in the present study were the annual rate of change in eGFRcr and incident eGFRcr-based CKD, given their availability at KORA S4, F4, and FF4. Creatinine was assessed in fresh serum using an enzymatic method at S4 (CREA plus, Boehringer, Mannheim, Germany), a modified kinetic rate Jaffe method at F4 and the first part of FF4 (CREA Flex, Dade Behring / Siemens Healthcare Diagnostics Products GmbH), and a Jaffe method for the second part of FF4 (Cobas 8000 instrument, Roche Diagnostics, Mannheim, Germany). Serum creatinine at KORA F4 and FF4 (part 1) were Isotope-Dilution Mass Spectrometry standardized. Kidney function was primarily assessed by eGFRcr calculated using the Chronic Kidney Disease Epidemiology Collaboration (CKD-EPI) Eq. 2009 [21]. A CKD case was defined as eGFRcr < 60 ml/min per 1.73m2 [1]. Participants free of CKD at S4 who had CKD at F4 or FF4 were defined as incident cases. In addition, the new race-free CKD-EPI Eq. 2021 [22] was used to calculate eGFRcr for supplementary analyses. Cystatin C-based eGFR (eGFRcys) was calculated using CKD-EPI cystatin C Eq. 2012 [23] and used for confirmation of significant associations. In comparison to eGFRcr, there were 278, 5, and 17 missing values on eGFRcys at S4, F4, and FF4, respectively, and imputation of missing values of eGFRcys was conducted by using a linear mixed-effects model. Additionally, urine albumin to creatinine ratio (UACR) was also available at F4. Details on measurements of UACR and cystatin C and imputation of missing values of eGFRcys are presented in Additional file 2: Text S1.
Covariates
In all surveys, standardized face-to-face interviews were conducted by trained staff [17], gathering the following information: age, sex, physical activity (active/inactive), smoking status (never/former/current smoker), alcohol consumption (0/0–20.0/ > = 20 g/d), use of antihypertensive medication (yes/no), use of lipid-lowering medication (yes/no), prevalent diabetes (yes/no), prevalent cardiovascular diseases (yes/no), and fasting status (fasting for 8 h or more before blood was taken, yes/no) [24, 25]. Participants who were non-fasting before blood was taken were predominantly participants with diabetes. Anthropometric indices and blood pressure were measured based on standard protocols. High-density lipoprotein-cholesterol and triglycerides were measured in serum on Hitachi 717/917 (Boehringer, Mannheim, Germany), respectively.
Statistical analysis
Continuous variables were reported as mean (standard deviation, SD) for normally distributed data and median (interquartile range) for skewed data. Categorical variables were presented as total numbers with the corresponding percentage.
Identification and confirmation of change in eGFR-associated proteins
For participants with eGFR measured at more than one visit (n = 1140), the annual rate of change in eGFR was calculated as β coefficients from linear regression of eGFR regressed against age at the time of eGFR measurement for each participant, and thus, each participant had 1 annual rate of change in eGFR (Additional file 2: Figure S1). Additional file 2: Figure S2 shows the flowchart of statistical analyses. In the discovery analysis, linear regression models were used to estimate the associations of the 233 proteins with the annual rate of change in eGFRcr. Two models were constructed: model 1 adjusted for age, sex, and eGFRcr at baseline; model 2 further adjusted for body mass index, physical activity, smoking status, alcohol consumption, systolic blood pressure, use of antihypertensive medication, triglycerides (naturally log-transformed), high-density lipoprotein cholesterol, use of lipid-lowering medication, prevalent diabetes, prevalent cardiovascular diseases, and fasting status at baseline. The 66 biomarkers significantly associated with the annual rate of change in eGFRcr were taken to investigate their associations with the annual rate of change in eGFRcys. Given the high proportion of missing values at FF4, we did not use linear mixed-effect models. Benjamini–Hochberg false-discovery rate (FDR) was performed for multiple testing correction, and FDR < 0.05 was considered statistically significant [26]. FDR seemed to be more appropriate to correct for multiple testing in our large-scale proteomics exploratory study, because it controls the rate of false positives while still allowing for the identification of a number of potential biomarkers.
Several sensitivity analyses of associations between the 66 significant biomarkers and the annual rate of change in eGFRcr were conducted based on model 2 described above. The analyses were repeated after exclusion of participants who were non-fasting at the time of blood sampling (n = 113), exclusion of participants who had CKD at baseline (n = 54), exclusion of participants who had an increase in eGFRcr during follow-up (n = 151), or further adjusted for UACR at F4 to control for the confounding effect of albuminuria (UACR values were unavailable at S4). To partially address bias caused by loss to follow-up (due to death or other reasons, Fig. 1B), the inverse probability weighting method [27] was used to calculate a weight for each participant. Each participant’s probability of loss to follow-up (P1) was estimated by logistic regression with loss to follow-up (yes/no) as outcomes, including baseline covariates in the above model 2 as predictors. Inverse probability weighting-weight was calculated as 1/(1-P1) and these weights were applied in model 2 (details are presented in Additional file 2: Text S2).
To further assess the robustness of our results on the annual rate of change in eGFRcr, rapid decline in eGFRcr (yes/no), which was defined as the annual rate of change in eGFRcr < -3 ml/min/1.73 m2/year [28], was used to investigate associations with the 66 eGFRcr-associated biomarkers in logistic regressions, adjusting for the same covariates as in the above model 2.
Identification of incident CKD-associated proteins
The 66 proteins significantly associated with the annual rate of change in eGFRcr were taken to investigate their associations with eGFRcr-based incident CKD, using interval-censored Cox regression models (500 bootstrap samples were used to construct 95% confidence intervals), adjusted for the same covariates in the above model 2, using R package “icenReg v.2.0.15” [29]. Cox proportional hazards models were not appropriate, because we did not know the exact time point of CKD occurrence. In sensitivity analyses, incident eGFRcr-based CKD cases were redefined as follows: Participants free of CKD at S4 or F4, respectively, had to have more than 25% decline in eGFRcr together with eGFRcr < 60 ml/min per 1.73m2 at the following follow-up (i.e., F4 or FF4), or participants free of CKD at S4 had to have more than 50% decline in eGFRcr together with eGFRcr < 60 ml/min per 1.73m2 at FF4 [28]. The controls were defined as eGFRcr ≥ 60 ml/min per 1.73m2 at S4, F4, and FF4.
The proteins consistently associated with the annual rate of change in eGFRcr, the annual rate of change in eGFRcys, and incident CKD were further investigated regarding their associations with UACR at F4 using linear regression. To annotate druggable targets of the identified proteins, related information (e.g., known drugs, corresponding diseases or indications, and clinical trials status) was gathered from Open Targets Platform (https://platform.opentargets.org/) [30] based on their UniProt_IDs. In order to examine the novelty of the identified proteins, relevant publications were searched to check whether the identified proteins had been previously reported to be associated with kidney function and/or CKD.
Mendelian randomization analysis
We applied a two-sample MR design using the largest genome-wide association studies (GWAS) results to date. Additional file 2: Figure S3 shows the process of MR analysis, and details of MR analysis are described in Additional file 2: Text S3 [31,32,33,34,35,36,37]. Briefly, selection of single nucleotide polymorphisms (SNPs) for proteins [31] and extraction of SNPs-eGFR decline associations [32] were from European ancestry population-based GWAS. To test the assumption of MR that instrumental variables are not associated with confounders, associations between selected SNPs and other traits were searched for in the PhenoScanner V2 [33]. One SNP (rs198389) was excluded given its associations with blood pressure (Additional file 1: Table S2), leaving 17 proteins for MR analysis (Additional file 1: Table S3). Wald ratio was calculated since only one SNP was available for each protein. MR analyses were performed using R package “TwoSampleMR v.0.5.6” [34].
Pathway enrichment analysis
To characterize biological pathways that are enriched for the identified proteins, a Gene Ontology enrichment analysis was performed, using R package “clusterProfiler v.4.0.5” [38]. To investigate the potential biological pathways linking identified biomarkers and kidney function, the analysis was limited to the biological process subontology, using Fisher's exact test.
All analyses were conducted by R version 4.1.0 (R Development Core Team, Vienna, Austria) and RStudio version 1.4.1717 (RStudio, Boston, MA, USA).
Results
Characteristics of the study population
Table 1 shows the baseline characteristics of participants. The 1140 participants had a mean age of 63.3 (SD, 5.36) years. The median annual rate of change in eGFRcr was -1.04 ml/min/1.73 m2/year (Additional file 2: Figure S4A). Characteristics and kidney function of all participants over the study period are shown in Additional file 1: Table S4. The mean eGFRcr was 82.9 (SD, 12.2) ml/min/1.73 m2 at S4 and declined to 67.6 (SD, 15.3) ml/min/1.73 m2 at FF4. Figure 1B shows the number of participants and reasons for loss to follow-up and Additional file 1: Table S5 shows baseline characteristics for participants with and without follow-up information on eGFRcr. Participants lost to follow-up tended to be older, were more frequently smokers, consumed less alcohol, were less physically active, and had higher systolic blood pressure and prevalence of diabetes.
Associations of proteomic biomarkers with kidney function decline
After adjustment for age, sex, and baseline eGFRcr in model 1, 95 out of 233 biomarkers were inversely associated with the annual rate of change in eGFRcr (FDR < 0.05), which means that in comparison to participants with lower levels of biomarkers at baseline, participants with higher levels of biomarkers had a faster decline in eGFRcr (Additional file 1: Table S6). After adjustment for additional covariates in model 2, 66 biomarkers showed inverse associations with change in eGFRcr (62 of these were also significant in model 1) (Additional file 1: Table S6 & Fig. 2). The top 3 biomarkers with the lowest FDR were KIM1 (FDR = 9.51E-09, β = -0.292), NT-proBNP (FDR = 1.62E-06, β = -0.249), and EPHB4 (FDR = 1.62E-06, β = -0.233). The β coefficients for biomarkers significantly associated with eGFRcr decline ranged from -0.292 to -0.098 ml/min/1.73 m2/year. When calculating eGFRcr using the 2021 equation, the correlation coefficient between the annual rate of change in eGFRcr-2009 and eGFRcr-2021 was 0.998 (Additional file 2: Figure S4D). Among the 233 biomarkers, 67 biomarkers were associated with eGFRcr-2021 decline (FDR < 0.05, β ranged from -0.301 to -0.104, Additional file 1: Table S7), and 65 out of these 67 were associated with eGFRcr-2009 decline. In 3 sensitivity analyses removing participants with non-fasting status, CKD, or increase in eGFRcr, 60, 62, and 63 of the 66 biomarkers remained significant, respectively (Additional file 1: Table S8 & Additional file 2: Figure S5). In sensitivity analyses model 2d (further adjusted for UACR) and 2e (inverse probability weighting-weight was applied), all 66 biomarkers remained significant, indicating that albuminuria and bias caused by loss to follow-up may not affect our associations (Additional file 1: Table S8). When investigating associations with rapid eGFRcr decline defined as the annual rate of change in eGFRcr < -3 ml/min/1.73 m2/year, 61 out of 66 biomarkers were positively associated with rapid eGFRcr decline, with odds ratios ranging from 1.29 to 2.09 (Additional file 1: Table S9).

Association of 233 proteomic biomarkers with the annual rate of change in eGFRcr. Detailed results of beta coefficients and FDR for the association of 233 proteins with change in eGFRcr are presented in Additional file 1: Table S6 for model 2. Abbreviations: eGFRcr, creatinine-based estimated glomerular filtration rate; FDR, Benjamini–Hochberg false-discovery rate. Full names of the biomarkers can be found in Additional file 1: Table S1
When investigating their associations with the annual rate of change in eGFRcys, 38 out of 66 biomarkers were inversely associated with change in eGFRcys at levels of FDR < 0.05 (Additional file 1: Table S10 & Additional file 2: Figure S6), and associations of the total 233 biomarkers with the annual rate of change in eGFRcys are presented in Additional file 1: Table S11. Associations between baseline characteristics and the annual rate of change in eGFRcr and eGFRcys are presented in Additional file 1: Table S12.
Associations of proteomic biomarkers with eGFRcr-based incident CKD
Among 1086 participants free of eGFRcr-based CKD at baseline, 262 cases of incident CKD were identified during 11,849 person-years of follow-up. Twenty-eight out of the 66 eGFRcr change-associated biomarkers were associated with incident CKD in model 2 (FDR < 0.05, Additional file 1: Table S10 & Additional file 2: Figure S7). The hazard ratios (HRs) for biomarkers with significant associations with incident CKD ranged from 1.16 to 1.52. The top 3 biomarkers with the highest HRs and lowest FDR for the associations were TRAIL-R2 (HR, 1.52), TNFRSF9 (HR, 1.51), and TNFRSF11A (HR, 1.49). In sensitivity analyses using the alternative definition of incident cases, 166 cases were identified and 30 out of 66 biomarkers were associated with incident CKD (FDR < 0.05, HRs ranged from 1.21 to 1.67), with 27 biomarkers overlapping with the initially identified 28 biomarkers (Additional file 1: Table S9).
We found that 21 proteins were consistently associated with faster decline in eGFRcr, faster decline in eGFRcys, and higher risk of incident CKD (Table 2, Additional file 1: Table S10 & Fig. 3). Among these 21 biomarkers, 17 were also positively associated with higher levels of UACR at F4 (Additional file 1: Table S13). In the discovery of their potential to serve as drug targets, we found 10 out of 21 have been used as drug targets for drugs to treat a wide range of diseases or indications, and IL2-RA has been used as drug target for kidney failure and CKD treatment (Additional file 1: Table S14). All 21 identified biomarkers have been reported in previous proteomic studies (Additional file 1: Table S15). The pairwise correlations of these 21 biomarkers are shown in Additional file 2: Figure S8. After correction for multiple testing, 199 out of 210 pairs of correlations were significant, with a mean correlation coefficient of 0.35 (range 0.14 to 0.88) for significant correlations.

Overlap of proteomic biomarkers associated with kidney function decline and incident CKD. Kidney function decline includes both the annual rate of change in eGFRcr and the annual rate of change in eGFRcys. Detailed results are presented in Additional file 1: Table S10. Abbreviations: CKD, chronic kidney disease; eGFRcr, creatinine-based estimated glomerular filtration rate; eGFRcys, cystatin C-based estimated glomerular filtration rate; HR, hazard ratio. Full names of the biomarkers can be found in Additional file 1: Table S1
Mendelian randomization analysis
The MR analysis results show a potential causal association of a higher level of TNFRSF11A (β = 0.024, P-value = 0.030) with faster eGFR decline (β > 0 means a positive association with eGFR decline [32]). However, no significant associations were observed for any of the proteins after multiple corrections (FDR > 0.05, Table 3).
Pathway enrichment analysis
A total of 254 pathways that reached statistical significance (FDR < 0.05) were identified when using 21 biomarkers (Table 2 & Fig. 3). The top 15 enriched pathways were characterized by processes relating to the response of a tumor necrosis factor (TNF) stimulus, T cell proliferation, monocyte chemotaxis, and regulation of lymphocyte and leukocyte chemotaxis (Table 4 & Additional file 2: Figure S9).
Discussion
In this prospective cohort study, we investigated the associations of 233 proteins with longitudinal change in kidney function and incident CKD among 1140 participants. A total of 66 biomarkers were associated with the annual rate of change in eGFRcr in discovery analysis, and 21 biomarkers out of these, were found to be also associated with both the annual rate of change in eGFRcys and incident CKD. Using a two-sample MR approach, we provided preliminary evidence suggesting a potential causal association between TNFRSF11A and kidney function decline (P-value = 0.030, FDR = 0.463).
All 21 biomarkers that we identified were associated with greater kidney disease risk, probably because we targeted 233 CVD- and inflammation-related biomarkers. Our results were consistent with previous studies investigating proteomic biomarkers measured by the same Olink panels and kidney function [8, 12, 15]. Some of the proteins that we identified are well-known biomarkers of kidney function, such as kidney injury molecule (KIM1), TNF-R1, TNF-R2, and TNF-related apoptosis-inducing ligand receptors 2 (TRAIL-R2), supporting the feasibility of proteomic analysis to identify biomarkers of kidney function decline. In the present study, KIM1 was the biomarker with the strongest association with kidney function decline. KIM1 has been extensively studied and represents a potential biomarker of tubular injury in both animals and humans [39, 40]. Longitudinal studies have also reported that urinary and blood KIM1 are positively associated with kidney function decline, incident CKD, and CKD progression in both diabetic patients and the general population [8, 13, 14, 39, 41, 42]. When investigating the associations with incident CKD, TRAIL-R2 was the biomarker with the strongest association in our study. The best-understood function of TRAIL-R2 is the induction of apoptosis [43]. TRAIL-R2 has been found to be associated with kidney function decline in several proteomic studies [8, 10, 11, 13]. For example, TRAIL-R2 was the biomarker with the strongest association with kidney function decline among 80 CVD-related plasma proteins in a longitudinal study [10]. Several other TNF superfamily receptors (TNFRSF) were found to be related to kidney function decline in our study, including TNF-R1, TNF-R2, TNFRSF9, and TNFRSF11A. In our pathway enrichment analysis, TNF response- and inflammatory response-related pathways were in the top 15 pathways related to kidney function and CKD (Table 4 & Additional file 2: Figure S9). Thus, our study provides additional evidence that TNF signaling pathways and inflammation may play a role in the pathophysiology of CKD. Similarly, a previous study identified a panel of 17 proteins from 194 plasma inflammatory proteins to be associated with end-stage kidney disease (ESKD) risk in diabetic patients, and these 17 proteins were enriched for TNF superfamily receptors [9]. The findings supported the involvement of immune response mechanisms in the development of CKD, which is also consistent with our pathway analysis, implicating T cell proliferation-related mechanisms involved in kidney function pathophysiology. Three chemokines, including C–C motif chemokine 3 (CCL3), C–C motif chemokine 7 (CCL7), and lymphotactin (XCL1), were involved in more than half of the top 15 pathways, and have been reported to be inversely associated with kidney function in previous studies [8, 10, 12, 13, 15]. Chemokines may play a key role in guiding inflammatory cells into the sites of inflammation in kidneys and recruiting immune cells such as T cells and monocytes during the development of chronic kidney injury [44]. Increasing evidence suggests that chemokines and their receptors may be potential targets for anti-inflammatory therapy in CKD [45].
Another important biomarker we identified was N-terminal prohormone brain natriuretic peptide (NT-proBNP), which has been shown to be a reliable biomarker for diagnosis of heart failure and prognostic evaluation among patients with heart failure [46]. In the present study, NT-proBNP was the second strongest biomarker associated with kidney function decline, which is in line with findings from other KORA cohort-based studies [47, 48]. Similarly, a recent proteomic study found that higher plasma NT-proBNP was associated with worsening kidney function among 5131 patients with type 2 diabetes [8]. Community-based longitudinal studies have also found that blood NT-proBNP is positively associated with kidney function decline and incident CKD [49, 50]. The exact mechanisms explaining the link between NT-proBNP and kidney function decline remain unclear. Several pathways have been proposed to explain the association. The increase in blood NT-proBNP can result from cardiac stretch, volume overload, and venous congestion, which in turn, are potential risk factors of kidney function decline. For example, volume overload or venous congestion can lead to an increase in central venous pressure, which has been reported to be associated with impaired kidney function [51, 52]. On the other hand, NT-proBNP is partially dependent on kidney clearance for elimination, so the concentration of NT-proBNP accumulates with impaired kidney function [53, 54]. Thus, NT-proBNP could be only a marker for other kidney-damaging risk factors rather than a causal risk factor itself. Further studies are warranted to explore the underlying mechanisms. In addition to NT-proBNP, we also identified another heart failure-related biomarker, adrenomedullin, which has previously been found to play a pathophysiological role in kidney disease [55].
A similar previous study by Grams et al. [14] investigated associations of 4877 proteins measured by the SomaScan platform with a composite outcome of more than 50% eGFR decline or ESKD among 3 American-based cohorts, including 2 kidney disease-related cohorts (1 of them was an African American cohort). However, our study was based on a community-based cohort of a relatively healthy European population. As differences in dietary habits and genetic background between ethnically diverse populations may affect both levels of protein expression and kidney function, it is important to verify observed associations in independent populations from various regions. Furthermore, most of the CKD cases we identified did not yet progress to ESKD (only 2 out of 262 incident cases had ESKD, i.e., eGFRcr < 15 ml/min per 1.73m2). Thus, we mainly focused on the annual rate of kidney function decline as an outcome rather than severe eGFR decline (e.g., ≥ 50% decline) or ESKD. Of note, our sensitivity analyses on rapid kidney function decline and redefinition of incident CKD cases show robust results (Additional file 1: Table S9). In our study, we targeted 233 CVD- and inflammation-related proteins measured by the Olink platform based on prior knowledge of close CKD-CVD relations, providing a more targeted approach to uncover pathways and mechanisms underlying kidney disease compared to the more comprehensive SomaScan platform used in the previous study [14]. However, it is worth noting that the smaller number of proteins measured by the Olink platform may be seen as a disadvantage compared to the SomaScan platform [56]. In a study by Rooney et al. [57] comparing correlations of 417 proteins that overlapped between the Olink and SomaScan platforms in 427 participants, the median Spearman correlation coefficient was 0.53 (range -0.21 to 0.97) and only 19% of the correlation coefficients were higher than 0.8. When Rooney et al. [57] further investigated associations of the overlapping proteins with eGFR, Olink platform-based proteins demonstrated more associations with eGFR, particularly in the group of proteins with Spearman correlation coefficients less than 0.3. Katz et al. [56] reported similar results and showed that the median Spearman correlation coefficient of proteins that overlapped between the two platforms was 0.45. These results show that findings from proteomic studies can be affected by the used platform, but the superiority of one platform over the other has not been clearly established yet. Thus, proteomic studies based on different platforms are important and our study adds to the existing literature in the field.
In MR analysis, our results show preliminary evidence suggesting a potential causal association of TNFRSF11A with eGFR decline (P-value = 0.030). A previous GWAS in 583 coronary patients observed an association of a polymorphism located within the genomic region of TNFRSF11A with kidney function decline [58]. In line with these associations, longitudinal studies that examined proteomics of kidney function also reported that plasma TNFRSF11A was positively associated with kidney function [8, 11]. MR is an effective approach to provide a robust and less biased estimate of causal associations, but our MR analysis was limited by the availability of GWAS, especially GWAS of proteins measured using the Olink platform. As GWAS by Sun et al. [31] reported only cis-SNPs with significance at the level p < 3.4E-11, each protein had only 1 cis-SNP as instrument, which limited the possibility to test the robustness of our results in sensitivity analyses and may have reduced our statistical power [59]. Additionally, the presence of overlapping participants in the 2 GWAS used in the present MR analysis may have caused bias [35], although this potential bias did not significantly change our MR results when using a maximum likelihood method to address it (details in Additional file 2: Text S3 & Additional file 1: Table S16). Further MR analyses based on multiple instruments GWAS summary statistics from larger populations are warranted.
Study strengths include the assessment of a large number of proteomic biomarkers and the use of a large prospective cohort study, with a median follow-up time of 13.4 years and two follow-up visits for most participants. The Strengthening the Reporting of Observational Studies in Epidemiology checklists for cohort studies and MR studies are presented in Additional file 1: Table S17 & 18, respectively. However, several limitations should also be considered. First, we used FDR to correct for multiple testing, which is less conservative and may increase the risk of false positives compared to Bonferroni correction. Furthermore, we did not validate the identified biomarkers in external cohorts since all our 21 identified biomarkers have been reported in previous proteomic studies (Additional file 1: Table S15). Second, there may be misclassification resulting from measurement errors due to different measurement methods of serum creatinine and cystatin C at S4/F4/FF4, and imputation of missing value of eGFRcys. Additionally, we defined CKD cases based on a single creatinine measurement, which does not fulfill the diagnosis of CKD in clinical practice that the presence of eGFRcr < 60 ml/min per 1.73m2 persists for more than 3 months. Thus, because our data may not be ideally suited for predictive analysis due to various limitations, such as a suboptimal CKD diagnosis and lack of external validation cohorts, we did not develop a prediction model for the development and progression of CKD. Our primary aim was to identify potential biomarkers related to kidney function decline, which may contribute to the development of predictive models, diagnostic strategies, and therapeutic targets in the future. Third, there may be selection bias due to loss to follow-up, but the results of sensitivity analysis with inverse probability weights suggest that this may not affect our result remarkably. Finally, although we adjusted for multiple confounders, we were unable to adjust for albuminuria at baseline due to lack of data.
Conclusions
In conclusion, we found 21 known proteins to be associated with kidney function decline and incident CKD in a Caucasian community-based population and provided further evidence regarding new diagnostic or prognostic biomarkers and therapeutic targets for CKD. Although the current underpowered MR analysis failed to find convincing evidence for causal associations of the 21 proteins with kidney function decline, our results provide preliminary evidence suggesting a potential causal association between TNFRSF11A and kidney function decline and a role of TNF signaling pathways in the pathophysiology of CKD. Further MR studies are needed to establish and validate a conclusive causal association.
Availability of data and materials
The informed consent given by KORA study participants does not cover data posting in public databases. Cooperation partners can obtain permission to use KORA data under the terms of a project agreement (https://helmholtz-muenchen.managed-otrs.com/external).
Abbreviations
- CKD:
-
Chronic kidney disease
- CVD:
-
Cardiovascular disease
- eGFR:
-
Estimated glomerular filtration rate
- eGFRcr:
-
Creatinine-based estimated glomerular filtration rate
- eGFRcys:
-
Cystatin c-based estimated glomerular filtration rate
- ESKD:
-
End-stage kidney disease
- FDR:
-
Benjamini–Hochberg false-discovery rate
- GWAS:
-
Genome-wide association study
- HR:
-
Hazard ratio
- KORA:
-
Cooperative Health Research in the Region of Augsburg
- MR:
-
Mendelian randomization
- SD:
-
Standard deviation
- SNP:
-
Single nucleotide polymorphism
- TNF:
-
Tumor necrosis factor
- UACR:
-
Urine albumin-creatinine ratio
References
Webster AC, Nagler EV, Morton RL, Masson P. Chronic Kidney Disease. Lancet. 2017;389(10075):1238–52.
GBD-CKD-Collaboration. Global, regional, and national burden of chronic kidney disease, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 2020;395(10225):709–33.
Laffin LJ, Bakris GL. Intersection between chronic kidney disease and cardiovascular disease. Curr Cardiol Rep. 2021;23(9):117.
Podkowinska A, Formanowicz D. Chronic Kidney Disease as Oxidative Stress- and Inflammatory-Mediated Cardiovascular Disease. Antioxidants. 2020;9(8):752.
Assarsson E, Lundberg M, Holmquist G, Bjorkesten J, Thorsen SB, Ekman D, Eriksson A, Rennel Dickens E, Ohlsson S, Edfeldt G, et al. Homogenous 96-plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PLoS One. 2014;9(4):e95192.
Good DM, Zurbig P, Argiles A, Bauer HW, Behrens G, Coon JJ, Dakna M, Decramer S, Delles C, Dominiczak AF, et al. Naturally occurring human urinary peptides for use in diagnosis of chronic kidney disease. Mol Cell Proteomics. 2010;9(11):2424–37.
Matias-Garcia P, Wilson R, Guo Q, Zaghlool S, Eales J, Xu X, Charchar F, Dormer J, Maalmi H, Schlosser P et al: Plasma Proteomics of Renal Function: A Trans-ethnic Meta-analysis and Mendelian Randomization Study. J Am Soc Nephrol. 2021.
Ferreira JP, Rossignol P, Bakris G, Mehta C, White WB, Zannad F. Blood and Urine Biomarkers Predicting Worsening Kidney Function in Patients with Type 2 Diabetes Post-Acute Coronary Syndrome: An Analysis from the EXAMINE Trial. Am J Nephrol. 2021;52(12):969–76.
Niewczas MA, Pavkov ME, Skupien J, Smiles A, Md Dom ZI, Wilson JM, Park J, Nair V, Schlafly A, Saulnier PJ, et al. A signature of circulating inflammatory proteins and development of end-stage renal disease in diabetes. Nat Med. 2019;25(5):805–13.
Carlsson AC, Ingelsson E, Sundstrom J, Carrero JJ, Gustafsson S, Feldreich T, Stenemo M, Larsson A, Lind L, Arnlov J. Use of proteomics to investigate kidney function decline over 5 years. Clin J Am Soc Nephrol. 2017;12(8):1226–35.
Ihara K, Skupien J, Krolewski B, Md Dom ZI, O’Neil K, Satake E, Kobayashi H, Rashidi NM, Niewczas MA, Krolewski AS. A profile of multiple circulating tumor necrosis factor receptors associated with early progressive kidney decline in Type 1 Diabetes is similar to profiles in autoimmune disorders. Kidney Int. 2021;99(3):725–36.
Maalmi H, Herder C, Strassburger K, Urner S, Jandeleit-Dahm K, Zaharia OP, Karusheva Y, Bongaerts BWC, Rathmann W, Burkart V, et al. Biomarkers of Inflammation and Glomerular Filtration Rate in Individuals with Recent-Onset Type 1 and Type 2 Diabetes. J Clin Endocrinol Metab. 2020;105(12):e4370–81.
Lind L, Sundstrom J, Larsson A, Lampa E, Arnlov J, Ingelsson E. Longitudinal effects of aging on plasma proteins levels in older adults - associations with kidney function and hemoglobin levels. PLoS One. 2019;14(2): e0212060.
Grams ME, Surapaneni A, Chen J, Zhou L, Yu Z, Dutta D, Welling PA, Chatterjee N, Zhang J, Arking DE, et al. Proteins Associated with Risk of Kidney Function Decline in the General Population. J Am Soc Nephrol. 2021;32(9):2291–302.
Nano J, Schottker B, Lin JS, Huth C, Ghanbari M, Garcia PM, Maalmi H, Karrasch S, Koenig W, Rothenbacher D, et al. Novel biomarkers of inflammation, kidney function and chronic kidney disease in the general population. Nephrol Dial Transplant. 2022;37(10):1916–26.
Kobayashi H, Looker HC, Satake E, Saulnier PJ, Md Dom ZI, O’Neil K, Ihara K, Krolewski B, Galecki AT, Niewczas MA, et al. Results of untargeted analysis using the SOMAscan proteomics platform indicates novel associations of circulating proteins with risk of progression to kidney failure in diabetes. Kidney Int. 2022;102(2):370–81.
Holle R, Happich M, Lowel H, Wichmann HE, Group MKS. KORA–a research platform for population based health research. Gesundheitswesen. 2005;67(Suppl 1):S19-25.
von Elm E, Altman DG, Egger M, Pocock SJ, Gotzsche PC, Vandenbroucke JP, Initiative S. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: guidelines for reporting observational studies. Lancet. 2007;370(9596):1453–7.
Huemer MT, Bauer A, Petrera A, Scholz M, Hauck SM, Drey M, Peters A, Thorand B. Proteomic profiling of low muscle and high fat mass: a machine learning approach in the KORA S4/FF4 study. J Cachexia Sarcopenia Muscle. 2021;12(4):1011–23.
Petrera A, von Toerne C, Behler J, Huth C, Thorand B, Hilgendorff A, Hauck SM. Multiplatform approach for plasma proteomics: complementarity of olink proximity extension assay technology to mass spectrometry-based protein profiling. J Proteome Res. 2021;20(1):751–62.
Levey AS, Stevens LA, Schmid CH, Zhang YP, Castro AF, Feldman HI, Kusek JW, Eggers P, Van Lente F, Greene T, et al. A new equation to estimate glomerular filtration rate. Ann Intern Med. 2009;150(9):604–12.
Inker LA, Eneanya ND, Coresh J, Tighiouart H, Wang D, Sang Y, Crews DC, Doria A, Estrella MM, Froissart M et al: New Creatinine- and Cystatin C-Based Equations to Estimate GFR without Race. N Engl J Med. 2021.
Inker LA, Schmid CH, Tighiouart H, Eckfeldt JH, Feldman HI, Greene T, Kusek JW, Manzi J, Van Lente F, Zhang YL, et al. Estimating glomerular filtration rate from serum creatinine and cystatin C. N Engl J Med. 2012;367(1):20–9.
Meisinger C, Thorand B, Schneider A, Stieber J, Doring A, Lowel H. Sex differences in risk factors for incident type 2 diabetes mellitus: the MONICA Augsburg cohort study. Arch Intern Med. 2002;162(1):82–9.
Laxy M, Knoll G, Schunk M, Meisinger C, Huth C, Holle R. Quality of Diabetes Care in Germany Improved from 2000 to 2007 to 2014, but Improvements Diminished since 2007. Evidence from the Population-Based KORA Studies. Plos One. 2016;11(10):e0164704.
Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J R Stat Soc Ser B-Stat Methodol. 1995;57(1):289–300.
Weuve J, Tchetgen Tchetgen EJ, Glymour MM, Beck TL, Aggarwal NT, Wilson RS, Evans DA. Mendes de Leon CF: Accounting for bias due to selective attrition: the example of smoking and cognitive decline. Epidemiology. 2012;23(1):119–28.
Gorski M, Jung B, Li Y, Matias-Garcia PR, Wuttke M, Coassin S, Thio CHL, Kleber ME, Winkler TW, Wanner V, et al. Meta-analysis uncovers genome-wide significant variants for rapid kidney function decline. Kidney Int. 2021;99(4):926–39.
Anderson-Bergman C. icenReg: regression models for interval censored data in R. J Stat Softw. 2017;81(12):1–23.
Ochoa D, Hercules A, Carmona M, Suveges D, Baker J, Malangone C, Lopez I, Miranda A, Cruz-Castillo C, Fumis L, et al. The next-generation Open Targets Platform: reimagined, redesigned, rebuilt. Nucleic Acids Res. 2023;51(D1):D1353–9.
Sun BB, Chiou J, Traylor M, Benner C, Hsu Y-H, Richardson TG, Surendran P, Mahajan A, Robins C, Vasquez-Grinnell SG et al: Genetic regulation of the human plasma proteome in 54,306 UK Biobank participants. bioRxiv 2022:2022.2006.2017.496443. https://www.biorxiv.org/content/10.1101/2022.06.17.496443v1 .
Gorski M, Rasheed H, Teumer A, Thomas LF, Graham SE, Sveinbjornsson G, Winkler TW, Günther F, Stark KJ, Chai J-F et al: Genetic loci and prioritization of genes for kidney function decline derived from a meta-analysis of 62 longitudinal genome-wide association studies. Kidney Int. 2022.
Kamat MA, Blackshaw JA, Young R, Surendran P, Burgess S, Danesh J, Butterworth AS, Staley JR. PhenoScanner V2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics. 2019;35(22):4851–3.
Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, Laurin C, Burgess S, Bowden J, Langdon R, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018;7:e34408.
Burgess S, Davies NM, Thompson SG. Bias due to participant overlap in two-sample Mendelian randomization. Genet Epidemiol. 2016;40(7):597–608.
Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37(7):658–65.
Skrivankova VW, Richmond RC, Woolf BAR, Yarmolinsky J, Davies NM, Swanson SA, VanderWeele TJ, Higgins JPT, Timpson NJ, Dimou N, et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. JAMA. 2021;326(16):1614–21.
Wu T, Hu E, Xu S, Chen M, Guo P, Dai Z, Feng T, Zhou L, Tang W, Zhan L, et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation. 2021;2(3):100141.
Arici M. Kidney injury molecule-1: a successful quest for a predictive kidney disease marker? Nephrol Dial Transplant. 2020;35(2):194–7.
Ichimura T, Bonventre JV, Bailly V, Wei H, Hession CA, Cate RL, Sanicola M. Kidney injury molecule-1 (KIM-1), a putative epithelial cell adhesion molecule containing a novel immunoglobulin domain, is up-regulated in renal cells after injury. J Biol Chem. 1998;273(7):4135–42.
Schmidt IM, Srivastava A, Sabbisetti V, McMahon GM, He J, Chen J, Kusek JW, Taliercio J, Ricardo AC, Hsu CY, et al. Plasma Kidney Injury Molecule 1 in CKD: Findings From the Boston Kidney Biopsy Cohort and CRIC Studies. Am J Kidney Dis. 2022;79(2):231-243.e231.
Colombo M, Looker HC, Farran B, Hess S, Groop L, Palmer CNA, Brosnan MJ, Dalton RN, Wong M, Turner C, et al. Serum kidney injury molecule 1 and β(2)-microglobulin perform as well as larger biomarker panels for prediction of rapid decline in renal function in type 2 diabetes. Diabetologia. 2019;62(1):156–68.
Bertsch U, Roder C, Kalthoff H, Trauzold A. Compartmentalization of TNF-related apoptosis-inducing ligand (TRAIL) death receptor functions: emerging role of nuclear TRAIL-R2. Cell Death Dis. 2014;5:e1390.
Chung AC, Lan HY. Chemokines in renal injury. J Am Soc Nephrol. 2011;22(5):802–9.
Vielhauer V, Anders HJ. Chemokines and chemokine receptors as therapeutic targets in chronic kidney disease. Front Biosci (Schol Ed). 2009;1(1):1–12.
Lam CSP, Li YH, Bayes-Genis A, Ariyachaipanich A, Huan DQ, Sato N, Kahale P, Cuong TM, Dong Y, Li X, et al. The role of N-terminal pro-B-type natriuretic peptide in prognostic evaluation of heart failure. J Chin Med Assoc. 2019;82(6):447–51.
Luchner A, Behrens G, Stritzke J, Markus M, Stark K, Peters A, Meisinger C, Leitzmann M, Hense HW, Schunkert H, et al. Long-term pattern of brain natriuretic peptide and N-terminal pro brain natriuretic peptide and its determinants in the general population: contribution of age, gender, and cardiac and extra-cardiac factors. Eur J Heart Fail. 2013;15(8):859–67.
Luchner A, Hengstenberg C, Lowel H, Trawinski J, Baumann M, Riegger GA, Schunkert H, Holmer S. N-terminal pro-brain natriuretic peptide after myocardial infarction: a marker of cardio-renal function. Hypertension. 2002;39(1):99–104.
Sasaki T, Oishi E, Nagata T, Sakata S, Chen S, Furuta Y, Honda T, Yoshida D, Hata J, Tsuboi N, et al. N-Terminal Pro-B-Type Natriuretic Peptide and Incident CKD. Kidney Int Rep. 2021;6(4):976–85.
Bansal N, Katz R, Dalrymple L, de Boer I, DeFilippi C, Kestenbaum B, Park M, Sarnak M, Seliger S, Shlipak M. NT-proBNP and troponin T and risk of rapid kidney function decline and incident CKD in elderly adults. Clin J Am Soc Nephrol. 2015;10(2):205–14.
Uthoff H, Breidthardt T, Klima T, Aschwanden M, Arenja N, Socrates T, Heinisch C, Noveanu M, Frischknecht B, Baumann U, et al. Central venous pressure and impaired renal function in patients with acute heart failure. Eur J Heart Fail. 2011;13(4):432–9.
Damman K, van Deursen VM, Navis G, Voors AA, van Veldhuisen DJ, Hillege HL. Increased central venous pressure is associated with impaired renal function and mortality in a broad spectrum of patients with cardiovascular disease. J Am Coll Cardiol. 2009;53(7):582–8.
van Kimmenade RR, Januzzi JL Jr, Bakker JA, Houben AJ, Rennenberg R, Kroon AA, Crijns HJ, van Dieijen-Visser MP, de Leeuw PW, Pinto YM. Renal clearance of B-type natriuretic peptide and amino terminal pro-B-type natriuretic peptide a mechanistic study in hypertensive subjects. J Am Coll Cardiol. 2009;53(10):884–90.
Luchner A, Hengstenberg C, Lowel H, Riegger GA, Schunkert H, Holmer S. Effect of compensated renal dysfunction on approved heart failure markers: direct comparison of brain natriuretic peptide (BNP) and N-terminal pro-BNP. Hypertension. 2005;46(1):118–23.
Nishikimi T. Adrenomedullin in the kidney-renal physiological and pathophysiological roles. Curr Med Chem. 2007;14(15):1689–99.
Katz DH, Robbins JM, Deng S, Tahir UA, Bick AG, Pampana A, Yu Z, Ngo D, Benson MD, Chen ZZ, et al. Proteomic profiling platforms head to head: Leveraging genetics and clinical traits to compare aptamer- and antibody-based methods. Sci Adv. 2022;8(33):eabm5164.
Rooney MR, Chen J, Ballantyne CM, Hoogeveen RC, Tang O, Grams ME, Tin A, Ndumele CE, Zannad F, Couper DJ, et al. Comparison of Proteomic Measurements Across Platforms in the Atherosclerosis Risk in Communities (ARIC) Study. Clin Chem. 2023;69(1):68–79.
Leiherer A, Muendlein A, Rein P, Saely CH, Kinz E, Vonbank A, Fraunberger P, Drexel H. Genome-wide association study reveals a polymorphism in the podocyte receptor RANK for the decline of renal function in coronary patients. PLoS One. 2014;9(12):e114240.
Burgess S, Davey Smith G, Davies NM, Dudbridge F, Gill D, Glymour MM, Hartwig FP, Holmes MV, Minelli C, Relton CL, et al. Guidelines for performing Mendelian randomization investigations. Wellcome Open Res. 2019;4:186.
Acknowledgements
We thank all participants for their long-term commitment to the KORA study, the staff for data collection and research data management and the members of the KORA Study Group (https://www.helmholtz-munich.de/en/epi/cohort/kora) who are responsible for the design and conduct of the study. JSL would like to thank the China Scholarship Council (CSC) for the financial support (No. 202008440343).
Funding
Open Access funding enabled and organized by Projekt DEAL. The KORA study was initiated and financed by the Helmholtz Zentrum München – German Research Center for Environmental Health, which is funded by the German Federal Ministry of Education and Research (BMBF) and by the State of Bavaria. Data collection in the KORA study is done in cooperation with the University Hospital of Augsburg. Proteomics measurements were supported by Helmholtz Institute for Metabolic, Obesity and Vascular Research – Project Initiative 2018 (HI-MAG). The funder had no role in study design, data collection, analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
JSL drafted the analysis plan, performed the statistical analysis, interpreted the data, and wrote the first draft of the manuscript. JN, CM, and AnP contributed to the analysis plan and data interpretation. BT designed the study, contributed to the analysis plan and data interpretation. AgP, SH, TZ, WK, AnP, and BT contributed data. All authors read and approved the final manuscript. JSL and BT had primary responsibility for the final content.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The KORA S4, F4, and FF4 studies were approved by the local ethical committee (Number: 99186) and all participants gave written informed consent.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. Normalized expressionvalues for proteomic biomarkers at baseline; Table S2. SNPs-traitsassociations retrieved from PhenoScanner V2 for proteins-related SNPs; Table S3. Harmonized summary statisticsused in Mendelian randomization analysis; TableS4. Characteristics of participants over the study period; Table S5. Baseline characteristics forparticipants with and without follow-up information on eGFRcr; Table S6. Longitudinal associationsbetween 233 proteomic biomarkers and the annual rate of change in eGFRcr; Table S7. Longitudinal associations between 233 proteomicbiomarkers and the annual rate of change in CKD-EPI Equation 2009-based andCKD-EPI Equation 2021-based eGFRcr;Table S8. Sensitivity analyses oflongitudinal associations between 66 proteomic biomarkers and the annual rateof change in eGFRcr; Table S9. Sensitivityanalyses of association of 66 proteomic biomarkers with rapid decline in eGFRcrand CKD incidence; Table S10. Associationof 66 proteomic biomarkers with the annual rate of change in eGFRcys and CKDincidence; Table S11. Longitudinalassociations between 233 proteomic biomarkers and the annual rate of change ineGFRcys; Table S12. Longitudinal associationsbetween baseline characteristics and the annual rate of change in eGFRcr andeGFRcys; Table S13. Association of 21 proteomic biomarkers with UACR; Table S14. Summary of druggable targetsand their corresponding disease of the 21 identified proteins; Table S15. Proteomic biomarkersassociated with kidney function and/or chronic kidney disease in previouscross-sectional or longitudinal proteomic studies; Table S16. Evaluation of bias due to participant overlap intwo-sample Mendelian randomization analysis; Table S17. STROBEStatement—checklist of items in reports of observational studies; Table S18. STROBE-MR checklist of recommended items in reports of Mendelianrandomization studies.
Additional file 2:
Text S1. Assessment of kidneyoutcomes; TextS2. Inverse probabilityweighting; Text S3. Mendelianrandomization analysis; Figure S1. Exampleof the annual rate of change in eGFR for each participant; Figure S2. Flowchart of statistical analyses; Figure S3. Genetic instrument selection and data harmonization forMendelian randomization analysis; FigureS4. Distribution and correlation between the annual rate of change ineGFRcr and eGFRcys; Figure S5. Overlapof proteomic biomarkers between biomarkers associated with the annual rate ofchange in eGFRcr in several sensitivity analyses; Figure S6. Longitudinal associations between 66 proteomicbiomarkers and the annual rate of change in eGFRcys; Figure S7. Association of 66 proteomic biomarkers with eGFRcr-basedCKD incidence; Figure S8. Pairwisecorrelation matrix between the 21 identified proteomic biomarkers; Figure S9. Pathway enrichment analysisof the 21 identified biomarkers showing top biological processes related tokidney function.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lin, Js., Nano, J., Petrera, A. et al. Proteomic profiling of longitudinal changes in kidney function among middle-aged and older men and women: the KORA S4/F4/FF4 study. BMC Med 21, 245 (2023). https://doi.org/10.1186/s12916-023-02962-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12916-023-02962-z