
Abstract
Regenerative medicine relying on cell and gene therapies is one of the most promising approaches to repair tissues. Multipotent mesenchymal stem/stromal cells (MSC), a population of progenitors committing into mesoderm lineages, are progressively demonstrating therapeutic capabilities far beyond their differentiation capacities. The mechanisms by which MSC exert these actions include the release of biomolecules with anti-inflammatory, immunomodulating, anti-fibrogenic, and trophic functions. While we expect the spectra of these molecules with a therapeutic profile to progressively expand, several human pathological conditions have begun to benefit from these biomolecule-delivering properties. In addition, MSC have also been proposed to vehicle genes capable of further empowering these functions. This review deals with the therapeutic properties of MSC, focusing on their ability to secrete naturally produced or gene-induced factors that can be used in the treatment of kidney, lung, heart, liver, pancreas, nervous system, and skeletal diseases. We specifically focus on the different modalities by which MSC can exert these functions. We aim to provide an updated understanding of these paracrine mechanisms as a prerequisite to broadening the therapeutic potential and clinical impact of MSC.
Similar content being viewed by others

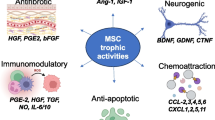

Introduction
The therapeutic promise of multipotent mesenchymal stem/stromal cells (MSC), a population of adult stem cells that can differentiate into cells deriving from mesoderm lineage, is rising [1–3]. MSC, historically isolated from bone marrow (BM), emerged in the biomedical field for their proliferative capacity and the potential to generate skeletal-related tissues [4]. Research therefore originally focused on their ability to differentiate into committed cell types within injured areas. More recently, evidence suggests that other MSC-related mechanisms, such as secretion of cytokines or release of microvesicles (MV), may play a significant role, by promoting the stimulation of endogenous cells, the inhibition of apoptosis, neovascularization, and anti-inflammatory responses [5–7]. Early in vivo evidence suggested that MSC may also induce tolerance [8]. A large body of in vitro data subsequently supported these findings, demonstrating MSC immunosuppressive functions on different immune effectors [9]. These findings revealed that MSC retain unique immunological features, which are paving the way for their clinical application in the treatment of invalidating or deadly immune-related disorders [6, 10, 11].
These MSC secretory functions have been progressively enhanced by cell modification within gene therapy approaches, promoting tissue restoration in a more targeted manner. MSC can be modified to carry therapeutic genes, serving as programmed molecule transmitters to overcome limitations connected with direct injection of beneficial proteins. In particular, these modified MSC can be used when better bioavailability of the desired molecule is required.
The emerging role of these mechanisms in specific contexts can be considered a paradigm changer. A proper understanding of these novel actions by MSC is desirable to allow regenerative therapies to gain robust clinical importance. To date, clinical trials have shown mild or no adverse effects from MSC treatment [10, 12]. Encouraging results have led to a growing number of applications that have received tremendous attention, such as the delivery of therapeutic products to repair neural injury; the amelioration of cardiovascular events; the promotion of bone and cartilage regeneration; and for counteracting liver, pancreas, lung, and kidney disorders. This review dissects these therapeutic applications of MSC, focusing on their ability to spontaneously or artificially secrete paracrine factors to counteract still-challenging human diseases.
The nervous system
Neurodegenerative disorders are attributed to the degeneration of specific neural cells with subsequent functional loss. Cell replacement and gene transfer to diseased or injured brain provide the basis for the development of new treatment strategies for a broad spectrum of human neurological conditions, including multiple sclerosis (MS) [13, 14], amyotrophic lateral sclerosis (ALS) [15, 16], Parkinson’s disease (PD) [17], Huntington’s disease (HD) [18], spinal cord injury (SCI) [19, 20], and stroke [21]. Growing evidence suggests that the effects orchestrated by MSC might only be marginally associated with the generation of newly graft-derived cells [22, 23], and MSC seem more likely to be producing neurotrophic and/or immunomodulatory factors to foster tissue repair in vivo (Table 1; Additional file 1: Link 1.1) [24].
Adipose (AD) or BM-derived MSC reduce the disease severity in a mouse model of MS mainly by influencing the immune response. These effects are especially reported in the early stages, where the autoreactive response against myelin begins to trigger disease development [13, 14]. Similarly, MSC immunosuppression applies to other neurological models, such as Krabbe disease [25].
Early clinical trials demonstrated the safety and feasibility of MSC for MS [26, 27] and, since then, a growing number of clinical studies enforced those observations [28]. Besides the absence of major side effects following intravenous (i.v.) or intrathecal delivery, few studies also showed that MSC therapy can improve or stabilize the course of progressive MS [29]. Structural and functional improvements in visual function have also been reported in some patients [30]. Immunological changes compatible with a reduced pro-inflammatory environment have been described, and are an indirect hint of the possible MSC-mediated actions in the context of MS [31].
Aside from immunomodulation, the beneficial effect of MSC may derive from the induction of local neurogenesis through the secretion of neural growth factors, such as basic fibroblast growth factor (bFGF), platelet-derived growth factor (PDGF), and brain-derived neurotrophic factor (BDNF) [13]. These same factors have been implicated in other experimental settings, such as improving motor performance in transgenic mouse models of ALS [15, 16]. An additional mechanism was reported by Marconi et al., who hypothesized that AD-MSC derived factors influence astrocytic secretome, which in turn can amplify the impact of AD-MSC. Thus, MSC can trigger a virtuous cycle to protect neurons from deterioration in the ALS mouse model [15]. In support of this concept, within a gene therapy context (Table 1; Additional file 1: Link 1.2), Suzuki et al. forced glial cell-derived neurotrophic factor (GDNF) expression in MSC and demonstrated an improvement in motor function in vivo, along with delayed disease progression and increased life span, in a rat model of familial ALS [32].
MSC-derived neurotrophic factors can also attenuate dopaminergic dysfunction and neuronal loss in a model of neurotoxin-induced PD [17], and GDNF-overexpressing MSC placed into the striatum of a PD rat model suggested that genetic modification of MSC holds therapeutic potential for PD [33, 34]. Similarly, Dey et al. demonstrated that human BM-MSC modified to express BDNF were able to reduce neuronal degeneration in a mouse model of HD [35]. MSC were also modified by microRNA encoding sequences targeting adenosine kinase (ADK) in a background of epilepsy, providing evidence that an ADK knockdown in human MSC reduces acute injury and seizures when injected into mouse hippocampus [36, 37]. Neurotrophic factors may also contribute to the reduction of infarct volume in cerebral ischemia; vascular endothelial growth factor (VEGF), epidermal growth factor (EGF), bFGF, BDNF, and GDNF increase after MSC transplantation in rats [21]. More specifically, an in vivo study [38] showed that exogenous EGF improved cerebral ischemic condition by inhibiting free radical generation and/or lipid peroxidation, preventing neuronal damage. For this reason, EGF and other factors were forced into MSC, with favorable outcomes in stroke animal models [39–42].
Based on these pre-clinical findings, several clinical trials explored and confirmed the safety of MSC-based approaches for ischemic strokes, reporting a functional recovery and a reduction in lesion extension a few weeks after i.v. infusion [43–45]. The abundance of pre-clinical data for other neurological conditions, such as PD and SCI, prompted the employment of MSC in pilot clinical studies that generated data on the safety and feasibility of the strategy [46, 47]. Further controlled clinical studies shall now be required to validate these outcomes and generate more evidence for a solid therapeutic benefit.
Beside central nervous tissues, MSC can also promote axonal regrowth in SCI, along with the production of neutrophil-activating protein-2 and neurotrophin (NT)-3 [20]. Thus, BM-MSC have been modified for NT-3 production with partial effects in vivo [48] (Table 1; Additional file 1: Links 1.1 and 1.2).
The heart
There is extreme clinical interest in novel treatments to improve heart function, and cellular therapies show promise [49]. Several types of cell have been so far used with the aim of generating functional cardiomyocytes and/or vascular cells in damaged myocardial tissue. MSC have progressively gained importance within these approaches [50].
Differentiation of transplanted MSC into cardiomyocytes and vessels was originally proposed as the main mechanism underlying their therapeutic action in cardiovascular diseases [51, 52]. More recently, it has been shown that the number of newly generated cells is too low to justify functional improvements, and evidence supports the hypothesis that paracrine mechanisms mediated by MSC may play an essential role in the reparation (Table 1; Additional file 2: Link 2.1) [53]. The mechanisms mediating these effects by paracrine factors are numerous and not completely clear, although it has been demonstrated that they can lead to neovascularization, cytoprotection, and endogenous cardiac regeneration (Fig. 1). More, post-infarction inflammatory and fibrogenic processes, cardiac contractility, and cardiac metabolism may also be influenced in a paracrine fashion [49].

MSC paracrine action/mechanisms in heart regeneration. Soluble factors released by MSC play an essential role in the post-ischemic reparative process improving angiogenesis, cytoprotection, and endogenous cardiac regeneration and reducing fibrosis. Ang-1 angiopoietin 1, HGF hepatocyte growth factor, MSC mesenchymal stem/stromal cells, VEGF vascular endothelial growth factor
The complexity of MSC secretome is hindering a definitive understanding; however, clues on the biological drivers for cardiac regeneration have been emerging and consistent evidence begins to indicate some pivotal players. VEGF is emerging as a critical paracrine factor for MSC-mediated cardioprotection. Several MSC types may also differentially release insulin-like growth factor (IGF)-1, transforming growth factor (TGF)-β2, and EGF [54–56]. AD-MSC are able to secrete numerous angiogenic, arteriogenic, chemotactic, and anti-apoptotic growth factors; for this reason their secretome has been involved in a series of novel strategies to enhance tissue restoration by increased angiogenesis [57–59]. Schenke-Layland et al. showed that AD-MSC accelerated vascularization in infarcted areas, increasing both capillary and arteriole density as a result of paracrine signaling [60]. This mechanism has been supported by other investigators who have considered adult stem cells from other sources administered into animal models post myocardial infarction (MI) [58, 61, 62].
Other cytoprotective factors such as hepatocyte growth factor (HGF) and angiopoietin (Ang)-1 are released by MSC when delivered into an acute MI rat model, and are associated with a significant improvement in cardiac function through increased angiogenesis and decreased infarct size [60, 63, 64]. Similarly, Li et al. showed an increase in capillary density along with significantly higher VEGF mRNA and protein levels after AD-MSC treatment [55].
Starting from these early understandings of MSC paracrine effects within infarcted regions, several authors selected putative beneficial factors to be introduced in a gene therapy approach (Table 1; Additional file 2: Link 2.2). A promising strategy to treat MI comes from Gao et al., who overexpressed VEGF in rat BM-MSC and generated effective myogenesis, preventing progressive heart dysfunction [65]. Similarly, murine BM-MSC modified by VEGF and/or HGF improved ventricular ejection function and reduced scar size [66]. Others showed that Ang-1 genetically modified rat BM-MSC were able to improve heart function by decreasing infarct area and promoting heart remodeling [67], indicating MSC-based gene therapies as feasible tools for heart regeneration.
Besides pro-angiogenic effects, data have demonstrated that cellular benefits might also be mediated by the activation of survival kinase pathways in response to MSC-secreted cytokines, additionally suggesting prevention of programmed cell death. Such pathways include activation of Akt, extracellular signal-regulated kinase 1/2 (ERK1/2), and signal transducer and activator of transcription 3, and inhibition of p38 mitogen-activated protein kinase, all instrumental in the promotion of cell proliferation [54]. To support this concept after MI, Gnecchi et al. genetically modified rat BM-MSC with Akt, showing that metabolism, glucose uptake, and cytosolic pH were maintained, and cardiac metabolism remodeling was prevented [68].
Emerging aspects from these pre-clinical findings are also related to cell homing and tissue persistence. Both aspects are crucial for clinical outcome in both intra-vessel and intra-MI injections. Considering rat BM-MSC, researchers have demonstrated that by overexpression of C-X-C chemokine receptor type 4 (CXCR4) i.e the stromal cell-derived factor (SDF)-1 receptor (largely involved in progenitor homing and survival) was possible to enhance engraftment within the infarct, thereby improving function and promoting neo-myoangiogenesis [69]. On tissue retainment and survival, recent data revealed that overexpression of cytoprotective proteins capable of enhancing expression of pro-survival genes, such as heme oxygenase-1, is associated with an increase in MSC survival [70].
Inflammation is a detrimental factor for tissue regeneration after MI. Attention has therefore been paid to anti-inflammatory approaches based on MSC. A paradigmatic example comes from the studies of Lee et al., who investigated gene expression of MSC trapped in lungs after i.v. injection in a mouse model of MI [71]. From all identified factors, tumor necrosis factor (TNF)-stimulated gene-6, a known anti-inflammatory molecule, contributed significantly to the amelioration of heart function, reducing infarct size and improving heart remodeling.
A large number of clinical trials have been completed for cardiovascular regeneration and their outcomes published (for extensive revisions see [10, 72]), with results suggesting at least the safety of these approaches. However, emerging data from patients with chronic/acute MI and refractory angina are still contradictory, showing either no significant effects or improvements in cardiac function associated with a reduction of scar tissue [73–76]. Therefore, basic investigations are currently following these studies to provide a better understanding of the optimal MSC source, delivery manner, cell doses, cell persistence, and precise mechanisms of action to ultimately create a more favorable prospect for the clinical uses of MSC for cardiovascular disorders.
The lung
While less prevalent than cardiovascular diseases, several lung pathologies represent unmet clinical needs possibly requiring novel cell based-therapeutic interventions. Investigations into different pulmonary diseases (Table 1; Additional file 3: Link 3.1) have revealed a basic understanding of the possible approaches for treatment that is yet to be consolidated by further pre-clinical research.
Studies on pulmonary fibrosis demonstrate that i.v. and endotracheal administration of MSC attenuates lung injury and fibrosis, suggesting a potential clinical application of MSC for the treatment of lethal idiopathic pulmonary fibrosis [77–79]. The mechanisms of the MSC-mediated amelioration in pulmonary fibrosis are not completely clear and an active participation of MSC through differentiation into alveolar epithelial cells in lung regeneration is still under debate [77, 78]. An endotoxin-induced lung fibrosis model in mice showed an MSC-mediated reduction in pulmonary fibrosis via paracrine downregulation of pro-inflammatory responses by reducing TNF-α and macrophage inflammatory protein (MIP)-2 while increasing the anti-inflammatory interleukin (IL)-10 [80]. Additionally, MSC can also upregulate matrix metalloproteinases (MMP), favoring the establishment of a microenvironment prone to extracellular matrix degradation and fibrosis reduction [77].
In a different context, a syngeneic model of pulmonary emphysema demonstrated that rat MSC reduced apoptosis of alveolar epithelial cells through upregulation of anti-apoptotic B-cell lymphoma (Bcl)-2 gene [81]. Moreover, Akram and colleagues showed that human MSC displayed site-specific migration into alveolar wounds where they secreted paracrine components for alveolar and small airway epithelial wound repair [82]. These paracrine effectors include fibronectin and lumican, known to be involved in corneal, skin, and mucosal healing. In relation to these findings, researchers have additionally gene-modified MSC for lung disorders (Table 1; Additional file 3: Link 3.2), selectively overexpressing either Ang-1 or IL-10. This resulted in a reduction of pro-inflammatory cytokines, increased lung permeability, and improved lung injury in vivo [83–85]. Experimental models of bronchopulmonary dysplasia (BPD) have also been considered; MSC mitigated lung inflammation, preventing lung vascular damage and alveolar growth impairment, ultimately inhibiting lung fibrosis [86].
Very curiously, the use of conditioned media from MSC has been shown to protect alveolar epithelial and lung microvasculature endothelial cells from oxidative stress, prevent oxygen-induced alveolar growth impairment, and stimulate endogenous lung progenitors such as bronchoalveolar stem cells [87]. For this reason, several researchers investigated the role of an emerging class of cell-derived particles, such as MV. Recent evidence suggested that MV can reduce lung inflammation and protein permeability, which in turn prevented the formation of pulmonary edema in Escherichia coli endotoxin-induced acute lung injury [88]. In a mouse model of hypoxic pulmonary hypertension, MV derived from mouse MSC-conditioned medium prevented vascular remodeling and an elevation in right ventricular systolic pressure by suppressing the hypoxic pulmonary influx of macrophages and by inducing pro-inflammatory and pro-proliferative mediators [89]. These data are generating a novel paradigm of tissue restoration by cell-derived bio-products that shall require far deeper investigation to determine the active principle(s) associated with the biological observations in pre-clinical models and in humans. To achieve this goal, several clinical pilot studies are already ongoing for the treatment of BPD, pulmonary emphysema, and pulmonary fibrosis (data extrapolated from www.ClinicalTrials.gov).
The liver
The liver has remarkable regenerative capacity in response to acute injuries. Either hepatic progenitors, the oval cells, or mature hepatocytes are able to re-enter the cell cycle to restore the hepatic mass. However, under chronic damage, these cells lose their ability to regenerate, causing “liver failure” [90]. For this reason, BM-MSC and human umbilical cord (hUC)-derived MSC-based approaches were introduced in early clinical studies [91, 92] for cirrhosis and end-stage liver failure, with improvements in liver function, reduced ascites, and no safety concerns [93–95].
Several studies on animal models reported the beneficial effect of MSC in promoting hepatic regeneration or preventing pathological changes (Table 1; Additional file 4: Link 4.1). The following mechanisms have been proposed to explain this therapeutic effect: homing and differentiation into hepatocytes, secretion of trophic molecules, and suppression of inflammation [96]. The liver homing properties of MSC were confirmed by the demonstration that CXCR4 overexpression enhanced engraftment and improved early liver regeneration [97]. However, reports supporting differentiation of MSC into hepatocytes are controversial [98, 99], so authors began to explore the therapeutic potential of MSC, hypothesizing their ability to produce bioactive factors [100]. These factors include HGF, VEGF, and nerve growth factor, which have the intrinsic ability to support hepatocyte proliferation and thereby facilitate the breakdown of fibrosis [96, 100–104]. To test this hypothesis, Ishikawa et al. genetically increased the expression of HGF in MSC. They observed that modified MSC accumulated in the liver, resulting in a decrease in fibrosis in vivo, thus confirming the homing potential and therapeutic benefits of MSC towards liver fibrosis [105]. Similarly, others introduced gene therapy approaches as outlined in Additional file 4: Link 4.2. MMP and fibrinogen-like protein-1 are also reported to be upregulated, further indicating the potential of MSC to counteract cirrhosis [106, 107]. Interestingly, MSC are also likely to exert an antioxidative action on resident cells by increasing superoxide dismutase activity and inhibiting reactive oxygen species production [108].
The anti-fibrogenic action of MSC has also been enhanced by gene modification. Li et al. overexpressed human alpha-1 antitrypsin, demonstrating that gene-modified MSC engraft into recipient livers and contribute to liver regeneration without eliciting an immune response in vivo [109]. This lack of significant immune response recalls known immunomodulatory properties of MSC that may represent a significant step in restoring liver injury. The local downregulation of pro-inflammatory cytokines and upregulation of anti-inflammatory cytokines, such as IL-10, after MSC transplantation has been reported to significantly improve function and reduce fibrosis [96, 100, 103, 107, 110].
As for lungs, MSC-derived MV are gaining interest in pre-clinical models of liver injury. MSC-MV reversed CCl4-induced injury in mice, through the activation of proliferative and regenerative responses. These in vivo beneficial effects confirmed the in vitro findings where MV sustained higher hepatocyte viability after injuries caused by acetaminophen and H2O2. The higher survival rate in vivo in the MV-treated group was also associated with upregulation of the priming-phase genes (coding for IL-6, TNF-α, and MIP-2) during liver regeneration, which subsequently lead to higher expression of proliferation proteins, such as proliferating cell nuclear antigen and cyclin D1 [111]. In another recent study, MV derived from hUC-MSC were used to treat CCl4-induced mouse liver fibrosis, ameliorating liver injury by inactivating the TGF-β1/Smad signaling pathway and inhibiting the epithelial–mesenchymal transition of hepatocytes [112].
The pancreas
The replacement of functional pancreatic β cells together with immunomodulation is seen as an attractive potential therapy for type 1 diabetes (T1D) [113]. However, extensive application of islet transplantation is hampered by the scarcity of donor tissue, the need for toxic lifelong immunosuppressive drugs, and graft failure usually within a few years [114, 115]. A possible solution to the cited challenges of islet transplantation has been found in MSC, and early trials based on their administration in patients with T1D have been reported. Carlsson et al. showed that autologous MSC treatment of new onset T1D may be a safe and feasible strategy to intervene in the disease process to preserve β-cell function. A randomized and double-blind phase II study is ongoing to validate these encouraging results [116].
From early studies, MSC immunomodulatory and paracrine properties, as opposed to the regenerative properties, are considered to have the greater effect in preventing, arresting, or reversing autoimmunity and ameliorating innate/alloimmune graft rejection (Table 1; Additional file 5: Link 5.1) [113]. At pre-clinical level, independent studies have proven that systemic MSC administration results in a functional recovery and normoglycemia [117–120]. However, the relative mechanisms contributed by MSC to this therapeutic effect are poorly understood. Reported data showed that MSC could differentiate into insulin-producing cells in vitro, indicating that transplantation of these islet-like cells is able to ameliorate hyperglycemia in diabetic rats [113, 121]. However, MSC differentiation has yielded contradictory results, mostly due to the discrepancy between the low level of functional integration of donor MSC and the observed recovery of pancreatic islets [118]. Therefore, the outcome of MSC treatment in diabetes is likely to be achieved by paracrine mechanisms along with immunomodulatory properties that can stimulate β cell repair/regeneration and abrogate immune injury, rather than by direct differentiation into β cells [113].
Although the potential trophic effect of MSC on pancreatic islets is not entirely clear, several MSC-secreted factors, such as IL-6, VEGF-A, HGF, and TGF-β, seem to improve islet cell viability and function by inhibiting apoptosis, inducing β cell proliferation, enhancing β cell insulin response to high glucose, and promoting islet revascularization [122]. Gao et al. demonstrated that conditioned medium from MSC exerts a striking protective effect on isolated islets exposed to streptozotocin (STZ). Moreover, injection of MSC-conditioned media into diabetic mice is able to partly restore the numbers of islets and β cells, and this stem cell paracrine action has been linked to activation of Akt signaling [123].
Besides these trophic functions, MSC immunomodulatory potential has recently gained interest for the treatment of T1D, mainly as an alternative to immunosuppressive drugs (Fig. 2). Ezquer et al. observed that, after i.v. administration in STZ-induced diabetic mice, MSC engraft into secondary lymphoid organs, inhibiting self-reacting T-cell expansion either by inducing regulatory T cells (Tregs) or by shifting the cytokine profile from a pro-inflammatory to an anti-inflammatory one [118]. Similarly, following MSC i.v. injection into NOD mice, others observed an increase in the percentage of Tregs and a shift towards a Th2 cytokine profile, both of which have been shown to help the recovery of islet cells [124].

MSC immunomodulatory properties in pancreatic regeneration. MSC are able to modulate the autoimmune response in T1D either by inducing regulatory T cells or by shifting the cytokine profile from a pro-inflammatory to an anti-inflammatory one. APC Antigen Presenting Cell, CTL Cytotoxic T Lymphocyte, Th1/2 T helper cell type 1/2, Treg regulatory T cell
The recent work by Favaro et al. provides in vitro evidence that some of the immunomodulatory actions of BM-MSC can be vicariated by MSC-derived MV. The observed MV inhibitory effect on glutamic acid decarboxylase (GAD)-stimulated peripheral blood mononuclear cells (PBMC) seems to involve prostaglandin E2 and TGF-β signaling pathways and IL-10. Blockade of MV internalization into PBMC, as well as pre-treatment of MV with RNAse, reduced IL-10 and TGF-β1 transcripts in MV-treated PBMC stimulated with GAD65. Furthermore, levels of mir-21, known to enhance TGF-β signaling, were increased in GAD-stimulated PBMC in the presence of MV [125]. To evaluate this hypothesis, a clinical trial with i.v. infusion of hUC-MSC-MV is ongoing to reduce inflammation and improve the β cell mass in patients with T1D (www.ClinicalTrials.gov: NCT02138331).
MSC have also been genetically modified to reprogram them into a pancreatic β cell lineage as well as to serve as gene delivery vehicles (Table 1; Additional file 5: Link 5.2) [126, 127]. For instance, gene-modified MSC carrying the human insulin gene have been assessed for T1D therapy [128]. Moreover, gene-modified BM-MSC carrying IL-1 receptor antagonist together with HGF or VEGF demonstrated clear advantages in improving the outcome of islet transplantation compared to non-transduced BM-MSC [129, 130]. Besides the potential for MSC to ameliorate T1D outcome, MSC have been evaluated for metabolic control in experimental models of type 2 diabetes (T2D). Si et al. proved that MSC infusion could partially reconstruct islet function and effectively ameliorate hyperglycemia in T2D rats, additionally acting to improve insulin sensitivity by upregulating glucose transporter type 4 expression and elevating phosphorylated insulin receptor substrate 1 and Akt levels in target tissues [131]. A preliminary clinical study involving 22 patients with T2D showed that Wharton’s jelly-derived MSC are able to significantly improve β-cell function without adverse effects [132].
MSC have also been evaluated as a cell-based therapeutic strategy for the treatment of other pancreatic diseases, such as acute pancreatitis (AP). Jung et al. significantly reduced the production of pro-inflammatory cytokines, increasing the production of anti-inflammatory factors by MSC. Curiously, Tregs were specifically recruited into the pancreas and along with MSC induced immunomodulation during AP [133].
The kidney
Several studies have reported multiple beneficial effects of MSC infusion in acute kidney injury (AKI) [134]. Systemically delivered MSC home to kidney after renal injury under the control of several factors, such as SDF-1 and PDGF [135, 136]. In addition, hyaluronic acid was shown to recruit exogenous MSC to injured renal tissue, and enhanced renal regeneration via CD44 [137]. Others report that MSC infusion is followed by an accelerated recovery of renal function compared to non-treated mice. Infused cells were able to partially differentiate into endothelial or smooth muscle cells and contributed to angiogenesis, vasculogenesis, and endothelial repair (Table 1; Additional file 6: Link 6.1) [138]. However, recent data showed that this apparent reparative function of MSC could also be achieved via intraperitoneal injections of MSC-conditioned medium alone, suggesting that MSC may additionally provide paracrine factors with positive impacts on kidney injury [139]. In contrast to these findings, others observed that i.v. infusions of MSC, but not of their conditioned medium, were associated with both the rapid recovery of kidney function and the enhanced survival of the mice [140].
Although extensive clinical studies are still limited in this context, the very interesting results obtained in pre-clinical steps prompted the translation of MSC-based treatments into humans. Preliminary results in a phase I trial using supra-renal aortic injection of allogeneic BM-MSC showed the safety of allogeneic MSC delivery and an amelioration of AKI [141]. Another phase II trial (NCT 01602328) is ongoing to assess the safety and efficacy of MSC in patients developing AKI. An overall view of currently available early data confirms the safety of these treatments. Additional investigations are now required to identify the precise mechanisms of action and confirm a benefit in human kidney disorders [142].
While the exact nature of the putative beneficial factors for the kidney is still under investigation, early evidence points toward IGF-1, HGF, EGF, VEGF [139, 143, 144] and bone morphogenetic protein (BMP)-7 as players capable of restoring kidney function and protecting against fibrosis [145]. To examine these findings, IGF-1 and erythropoietin (EPO) production were enhanced in MSC by gene modification. MSC co-expressing EPO and IGF-1 improved hematocrit levels and heart function in a renal failure mouse model [146]. Similarly, Zhen-Qiang et al. demonstrated that MSC overexpressing BMP-7 were able to improve renal function and regenerate tubular cells [147].
In a cisplatin-induced mouse model of AKI, Morigi et al. showed that UC-MSC could stimulate endogenous target cells to produce regenerative factors, including a robust HGF expression enhanced by hypoxic conditions and inflammatory cytokines [148]. Moreover, Tögel et al. suggested that MSC exert their renal protection through inhibition of pro-inflammatory cytokines [139]. These reparative roles of MSC are likely to be multifactorial and include the provision of cytokines to limit apoptosis, enhance proliferation, and dampen the inflammatory response [145]. This hypothesis has been supported by a gene therapy approach, in which modified MSC expressing tissue kallikrein generated a benefit to tubular injury thanks to regeneration and anti-inflammatory action [149]. Additional gene therapy strategies for kidney repair are reported in Additional file 6: Link 6.2. Besides the mentioned humoral factors, the role of MV secretion in MSC therapy has recently been outlined (Fig. 3). It has been demonstrated that MV released from MSC mimic their beneficial effects for the treatment of a glycerol-induced model of AKI and ischemia–reperfusion injury [150, 151]. The same group described that MV delivery may retain similar efficacy as human BM-MSC injections. Recent reports began deciphering the molecular pathways modulated by MSC-MV in the context of renal regeneration. Specifically, MSC-MV induced the expression of several anti-apoptotic genes, including Bcl-XL, Bcl2, and baculoviral IAP repeat containing 8, in renal tubular epithelial cells while simultaneously downregulating pro-apoptotic genes such as caspase 1, caspase 8, and lymphotoxin-alpha [152].

The therapeutic potential of MSC microvesicles in kidney regeneration. MSC MV mediate anti-apoptotic and pro-proliferative effects, simultaneously reducing oxidative stress to stimulate renal regeneration after acute kidney injury
Thus, MSC-MV may confer an anti-apoptotic phenotype necessary for tissue repair. In addition, MSC-MV stimulate renal cell proliferation by inducing the phosphorylation and subsequent activation of ERK 1/2, and blockade of ERK activation with a chemical inhibitor significantly reduces cell proliferation after MSC-MV treatment [153]. Although the exact molecules in the MV that mediate the anti-apoptotic and pro-proliferative effects have not been identified, these data demonstrate the ability of MSC-MV to simultaneously modulate several different pathways to stimulate renal regeneration. To date and to the best of our knowledge, no clinical study on microvesicles and AKI has been reported.
The skeletal tissues
Failure of bone repair is often associated with a relevant morbidity. Therapies using recombinant BMP with or without biomaterials show promise of becoming a clinically relevant procedure. However, the lack of optimal matrices for controlled, sustained BMP delivery, a short biological half-life of BMP, and the absence of appropriate BMP responsive cells in the fracture environment limit their usefulness [154]. To overcome these limitations, bone engineering methods using MSC and scaffolds provide promising new approaches for bone repair (Table 1; Additional file 7: Link 7.1) [155]. MSC, as key progenitor cells for bone regeneration, have been historically investigated to repair skeletal tissues [156–158] alone or in combination with osteoinductive factors, such as BMP-2 [159, 160]. Pre-clinical and clinical investigations have successfully combined BMP-2 with MSC as therapy for bone defects [161] and several vectors have been tested to provide these trophic factors to skeleton (Table 1; Additional file 7: Link 7.2). In particular, BMP were delivered as liposome-mediated plasmid DNA, adenoviral vectors, and lentiviral vectors [155]. These BMP-2-modified MSC increase alkaline phosphatase activity, mineralization, and cell proliferation, and induce ectopic bone formation, heal critical size bone defects, and repair fracture triggering spinal fusion in vivo [162]. In a recent publication, we revealed that the osteogenic performance of BM-MSC can be empowered by gene modification that introduces Homeobox protein Hox-B7, which in turn promotes an autocrine loop of bFGF—a key player in proliferation and osteogenic differentiation [163]. Beside a direct differentiation into bone cells, Otsuru et al. showed that MSC stimulate bone growth in a model of osteogenesis imperfecta by secreting soluble mediators, ultimately resulting in growth-plate chondrocyte proliferation leading to bone elongation [164].
The historical heritage that MSC retain on bone regeneration represents an advantage in their clinical implementation versus other target tissues. Several clinical studies are ongoing for non-union bone defects, mandible regeneration, osteonecrosis, osteogenesis imperfecta, and vertebral regeneration [165] (www.ClinicalTrials.gov). While these challenging trials are still revealing uncertainties for MSC as a cure for bone defects in humans, they are clearly indicating a path for the development of MSC-based therapeutics on a large scale, with a solid benefit in defined clinical indications [166].
With regards to cartilage, chondrocytes have a limited regenerative potential [167], most likely because of their avascularity and a low cellularity. Although current surgical therapeutic procedures for cartilage repair are clinically useful, they cannot restore a normal articular surface, in particular where inflammatory conditions exist. To overcome these drawbacks, MSC are being considered for their ability to differentiate into cartilage and act as immunosuppressive and anti-inflammatory agents in a variety of cartilage diseases. Among them, Augello et al. reported that a single intraperitoneal injection of allogeneic MSC was sufficient to prevent the occurrence of cartilage erosion in immunized mice, suggesting that MSC might act by inhibiting the activation and proliferation of tissue-specific autoreactive T cell clones, probably by educating antigen-specific Tregs [168].
Similarly, joint destruction caused by persistent inflammation, such as in rheumatoid arthritis (RA), is a possible clinical target for cartilage repair using BM-MSC. A number of studies, based mainly on experimental animal models, have recently provided interesting data on the potential of BM-MSC to suppress local inflammation and tissue damage in RA [169]. Other studies ascribe the significant reduction in the severity of arthritis to the ability of MSC to promote the downregulation of pro-inflammatory cytokines such as TNF-α, IL-1, and interferon-γ and the concomitant upregulation of IL-10 [170, 171].
Based on these in vitro and in vivo evidences, MSC have been introduced in humans for experimental purposes within trials investigating safety and efficacy [10, 12, 72, 172]. Outlining the safety of the approach, the intra-articular delivery of MSC appears promising although still requires additional investigation to definitively ameliorate the chondrogenic actions of MSC [173–175].
Current research offers a growing number of bioactive reagents, including proteins and nucleic acids, that may be used to augment different aspects of the repair process. It is difficult to effectively deliver these agents and gene transfer approaches are being developed to provide their sustained synthesis at sites of damage by MSC delivery. The list of potentially useful cDNAs for cartilage repair comprises members of the TGF-β superfamily, several BMPs, IGF-1, FGF, and EGF. Experimental data generated so far have shown that genetically modified MSC allow sustained transgene expression when transplanted into articular cartilage defects in vivo, and enhance the structural features of cartilaginous tissue repair [176, 177].
Conclusions
This review highlights that injured organs may benefit from MSC as regenerative tools able to differentiate accordingly, secrete useful factors, or both, with the final effect of counteracting damages. The reported data generally indicate amelioration, while less frequently clarifications on mechanism(s) driving the observed therapeutic potential have been addressed. Technicalities limit current understanding; nevertheless, efforts are being made to transfer knowledge from the laboratory to the clinic and vice versa to identify the drivers of the observed MSC impacts. Studies will be implemented on deciphering the ideal MSC tissue source for precise clinical application, as well as on the best delivery manner to exploit MSC potential through better cellular retention and optimized recruitment. More information will be gathered on the still poorly explored potential of MSC, such as whether MV, previously considered to be cell debris, may become an important mediator of intercellular communication. Collectively, this research will contribute to better characterized MSC that can be selected by physicians based on their patient needs, and on more precise information on the pivotal properties of MSC that lead to their enduring benefits.
Abbreviations
- AD:
-
adipose
- ADK:
-
adenosine kinase
- AKI:
-
acute kidney injury
- ALS:
-
amyotrophic lateral sclerosis
- Ang:
-
angiopoietin
- AP:
-
acute pancreatitis
- Bcl:
-
B-cell lymphoma
- BDNF:
-
brain-derived neurotrophic factor
- bFGF:
-
basic fibroblast growth factor
- BM:
-
bone marrow
- BMP:
-
bone morphogenetic protein
- BPD:
-
bronchopulmonary dysplasia
- CXCR4:
-
C-X-C chemokine receptor type 4
- EGF:
-
epidermal growth factor
- EPO:
-
erythropoietin
- ERK 1/2:
-
extracellular signal-regulated kinase 1/2
- GAD:
-
glutamic acid decarboxylase
- GDNF:
-
glial cell-derived neurotrophic factor
- HD:
-
Huntington’s disease
- HGF:
-
hepatocyte growth factor
- IGF:
-
insulin-like growth factor
- IL:
-
interleukin
- i.v.:
-
intravenous
- MI:
-
myocardial infarction
- MIP:
-
macrophage inflammatory protein
- MMP:
-
matrix metalloproteinases
- MS:
-
multiple sclerosis
- MSC:
-
multipotent mesenchymal stem/stromal cells
- MV:
-
microvesicles
- NT:
-
neurotrophin
- PBMC:
-
peripheral blood mononuclear cells
- PD:
-
Parkinson’s disease
- PDGF:
-
platelet-derived growth factor
- RA:
-
rheumatoid arthritis
- SCI:
-
spinal cord injury
- SDF:
-
stromal cell-derived factor
- STZ:
-
streptozotocin
- T1D:
-
type 1 diabetes
- T2D:
-
type 2 diabetes
- TGF:
-
transforming growth factor
- TNF:
-
tumor necrosis factor
- hUC:
-
human umbilical cord
- VEGF:
-
vascular endothelial growth factor
References
Caplan AI. Mesenchymal stem cells. J Orthop Res. 1991;9:641–50.
Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–7.
Friedenstein AJ. Precursor cells of mechanocytes. Int Rev Cytol. 1976;47:327–59.
Burdon TJ, Paul A, Noiseux N, Prakash S, Shum-Tim D. Bone marrow stem cell derived paracrine factors for regenerative medicine: current perspectives and therapeutic potential. Bone Marrow Res. 2011;2011:1–14.
Horwitz EM, Dominici M. How do mesenchymal stromal cells exert their therapeutic benefit? Cytotherapy. 2008;10:771–4.
Murphy MB, Moncivais K, Caplan AI. Mesenchymal stem cells: environmentally responsive therapeutics for regenerative medicine. Exp Mol Med. 2013;45, e54.
Bartholomew A, Sturgeon C, Siatskas M, Ferrer K, McIntosh K, Patil S, et al. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp Hematol. 2002;30:42–8.
English K, French A, Wood KJ. Mesenchymal stromal cells: facilitators of successful transplantation? Cell Stem Cell. 2010;7:431–42.
Sharma RR, Pollock K, Hubel A, McKenna D. Mesenchymal stem or stromal cells: a review of clinical applications and manufacturing practices. Transfusion. 2014;54:1418–37.
Shi Y, Su J, Roberts AI, Shou P, Rabson AB, Ren G. How mesenchymal stem cells interact with tissue immune responses. Trends Immunol. 2012;33:136–43.
Munir H, McGettrick HM. Mesenchymal stem cells therapy for autoimmune disease: risks and rewards. Stem Cells Dev. 2015. [Epub ahead of print].
Constantin G, Marconi S, Rossi B, Angiari S, Calderan L, Anghileri E, et al. Adipose-derived mesenchymal stem cells ameliorate chronic experimental autoimmune encephalomyelitis. Stem Cells. 2009;27:2624–35.
Gerdoni E, Gallo B, Casazza S, Musio S, Bonanni I, Pedemonte E, et al. Mesenchymal stem cells effectively modulate pathogenic immune response in experimental autoimmune encephalomyelitis. Ann Neurol. 2007;61:219–27.
Marconi S, Bonaconsa M, Scambi I, Squintani GM, Rui W, Turano E, et al. Systemic treatment with adipose-derived mesenchymal stem cells ameliorates clinical and pathological features in the amyotrophic lateral sclerosis murine model. Neuroscience. 2013;248C:333–43.
Vercelli A, Mereuta OM, Garbossa D, Muraca G, Mareschi K, Rustichelli D, et al. Human mesenchymal stem cell transplantation extends survival, improves motor performance and decreases neuroinflammation in mouse model of amyotrophic lateral sclerosis. Neurobiol Dis. 2008;31:395–405.
McCoy MK, Martinez TN, Ruhn KA, Wrage PC, Keefer EW, Botterman BR, et al. Autologous transplants of adipose-derived adult stromal (ADAS) cells afford dopaminergic neuroprotection in a model of Parkinson’s disease. Exp Neurol. 2008;210:14–29.
Lin Y-T, Chern Y, Shen C-KJ, Wen H-L, Chang Y-C, Li H, et al. Human mesenchymal stem cells prolong survival and ameliorate motor deficit through trophic support in Huntington’s disease mouse models. PLoS One. 2011;6:1–17.
Gu W, Zhang F, Xue Q, Ma Z, Lu P, Yu B. Transplantation of bone marrow mesenchymal stem cells reduces lesion volume and induces axonal regrowth of injured spinal cord. Neuropathology. 2010;30:205–17.
Yang C-C, Shih Y-H, Ko M-H, Hsu S-Y, Cheng H, Fu Y-S. Transplantation of human umbilical mesenchymal stem cells from Wharton’s jelly after complete transection of the rat spinal cord. PLoS One. 2008;3, e3336.
Wakabayashi K, Nagai A, Sheikh AM, Shiota Y, Narantuya D, Watanabe T, et al. Transplantation of human mesenchymal stem cells promotes functional improvement and increased expression of neurotrophic factors in a rat focal cerebral ischemia model. J Neurosci Res. 2010;88:1017–25.
Kokaia Z, Martino G, Schwartz M, Lindvall O. Cross-talk between neural stem cells and immune cells: the key to better brain repair? Nat Neurosci. 2012;15:1078–87.
Uccelli A, Laroni A, Freedman MS. Mesenchymal stem cells for the treatment of multiple sclerosis and other neurological diseases. Lancet Neurol. 2011;10:649–56.
Drago D, Cossetti C, Iraci N, Gaude E, Musco G, Bachi A, et al. The stem cell secretome and its role in brain repair. Biochimie. 2013;95:2271–85.
Ripoll CB, Flaat M, Klopf-Eiermann J, Fisher-Perkins JM, Trygg CB, Scruggs BA, et al. Mesenchymal lineage stem cells have pronounced anti-inflammatory effects in the twitcher mouse model of Krabbe’s disease. Stem Cells. 2011;29:67–77.
Karussis D, Karageorgiou C, Vaknin-Dembinsky A, Gowda-Kurkalli B, Gomori JM, Kassis I, et al. Safety and immunological effects of mesenchymal stem cell transplantation in patients with multiple sclerosis and amyotrophic lateral sclerosis. Arch Neurol. 2010;67:1187–94.
Yamout B, Hourani R, Salti H, Barada W, El-Hajj T, Al-Kutoubi A, et al. Bone marrow mesenchymal stem cell transplantation in patients with multiple sclerosis: a pilot study. J Neuroimmunol. 2010;227:185–9.
Dulamea A. Mesenchymal stem cells in multiple sclerosis - translation to clinical trials. J Med Life. 2015;8:24–7.
Bonab MM, Sahraian MA, Aghsaie A, Karvigh SA, Hosseinian SM, Nikbin B, et al. Autologous mesenchymal stem cell therapy in progressive multiple sclerosis: an open label study. Curr Stem Cell Res Ther. 2012;7:407–14.
Connick P, Kolappan M, Patani R, Scott MA, Crawley C, He X-L, et al. The Mesenchymal Stem Cells in Multiple Sclerosis (MSCIMS) trial protocol and baseline cohort characteristics: an open-label pre-test: post-test study with blinded outcome assessments. Trials. 2011;12:62.
Llufriu S, Sepúlveda M, Blanco Y, Marín P, Moreno B, Berenguer J, et al. Randomized placebo-controlled phase II trial of autologous mesenchymal stem cells in multiple sclerosis. PLoS One. 2014;9, e113936.
Suzuki M, McHugh J, Tork C, Shelley B, Hayes A, Bellantuono I, et al. Direct muscle delivery of GDNF with human mesenchymal stem cells improves motor neuron survival and function in a rat model of familial ALS. Mol Ther. 2008;16:2002–10.
Glavaski-Joksimovic A, Virag T, Mangatu TA, McGrogan M, Wang XS, Bohn MC. Glial cell line-derived neurotrophic factor-secreting genetically modified human bone marrow-derived mesenchymal stem cells promote recovery in a rat model of Parkinson’s disease. J Neurosci Res. 2010;88:2669–81.
Moloney TC, Rooney GE, Barry FP, Howard L, Dowd E. Potential of rat bone marrow-derived mesenchymal stem cells as vehicles for delivery of neurotrophins to the parkinsonian rat brain. Brain Res. 2010;1359:33–43.
Dey ND, Bombard MC, Roland BP, Davidson S, Lu M, Rossignol J, et al. Genetically engineered mesenchymal stem cells reduce behavioral deficits in the YAC 128 mouse model of Huntington’s disease. Behav Brain Res. 2010;214:193–200.
Boison D. Engineered adenosine-releasing cells for epilepsy therapy: human mesenchymal stem cells and human embryonic stem cells. Neurother J Am Soc Exp Neurother. 2009;6:278–83.
Ren G, Li T, Lan JQ, Wilz A, Simon RP, Boison D. Lentiviral RNAi-induced downregulation of adenosine kinase in human mesenchymal stem cell grafts: a novel perspective for seizure control. Exp Neurol. 2007;208:26–37.
Peng H, Wen TC, Tanaka J, Maeda N, Matsuda S, Desaki J, et al. Epidermal growth factor protects neuronal cells in vivo and in vitro against transient forebrain ischemia- and free radical-induced injuries. J Cereb Blood Flow Metab. 1998;18:349–60.
Horita Y, Honmou O, Harada K, Houkin K, Hamada H, Kocsis JD. Intravenous administration of glial cell line-derived neurotrophic factor gene-modified human mesenchymal stem cells protects against injury in a cerebral ischemia model in the adult rat. J Neurosci Res. 2006;84:1495–504.
Ikeda N, Nonoguchi N, Zhao MZ, Watanabe T, Kajimoto Y, Furutama D, et al. Bone marrow stromal cells that enhanced fibroblast growth factor-2 secretion by herpes simplex virus vector improve neurological outcome after transient focal cerebral ischemia in rats. Stroke. 2005;36:2725–30.
Kurozumi K, Nakamura K, Tamiya T, Kawano Y, Kobune M, Hirai S, et al. BDNF gene-modified mesenchymal stem cells promote functional recovery and reduce infarct size in the rat middle cerebral artery occlusion model. Mol Ther. 2004;9:189–97.
Miki Y, Nonoguchi N, Ikeda N, Coffin RS, Kuroiwa T, Miyatake S. Vascular endothelial growth factor gene-transferred bone marrow stromal cells engineered with a herpes simplex virus type 1 vector can improve neurological deficits and reduce infarction volume in rat brain ischemia. Neurosurgery. 2007;61:586–95.
Bang OY, Lee JS, Lee PH, Lee G. Autologous mesenchymal stem cell transplantation in stroke patients. Ann Neurol. 2005;57:874–82.
Honmou O, Houkin K, Matsunaga T, Niitsu Y, Ishiai S, Onodera R, et al. Intravenous administration of auto serum-expanded autologous mesenchymal stem cells in stroke. Brain. 2011;134:1790–807.
Lee JS, Hong JM, Moon GJ, Lee PH, Ahn YH, Bang OY, et al. A long-term follow-up study of intravenous autologous mesenchymal stem cell transplantation in patients with ischemic stroke. Stem Cells. 2010;28:1099–106.
Pal R, Venkataramana NK, Bansal A, Balaraju S, Jan M, Chandra R, et al. Ex vivo-expanded autologous bone marrow-derived mesenchymal stromal cells in human spinal cord injury/paraplegia: a pilot clinical study. Cytotherapy. 2009;11:897–911.
Venkataramana NK, Kumar SKV, Balaraju S, Radhakrishnan RC, Bansal A, Dixit A, et al. Open-labeled study of unilateral autologous bone-marrow-derived mesenchymal stem cell transplantation in Parkinson’s disease. Transl Res. 2010;155:62–70.
Zhang W, Yan Q, Zeng Y-S, Zhang X-B, Xiong Y, Wang J-M, et al. Implantation of adult bone marrow-derived mesenchymal stem cells transfected with the neurotrophin-3 gene and pretreated with retinoic acid in completely transected spinal cord. Brain Res. 2010;1359:256–71.
Gnecchi M, Danieli P, Cervio E. Mesenchymal stem cell therapy for heart disease. Vascul Pharmacol. 2012;57:48–55.
Madonna R, Rokosh G, De Caterina R, Bolli R. Hepatocyte growth factor/Met gene transfer in cardiac stem cells--potential for cardiac repair. Basic Res Cardiol. 2010;105:443–52.
Pittenger MF, Martin BJ. Mesenchymal stem cells and their potential as cardiac therapeutics. Circ Res. 2004;95:9–20.
Tomita S, Li R-K, Weisel RD, Mickle DAG, Kim E-J, Sakai T, et al. Autologous transplantation of bone marrow cells improves damaged heart function. Circulation. 1999;100:II–247–56.
Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adult stem cell signaling and therapy. Circ Res. 2008;103:1204–19.
Jiang Z, Hu X, Yu H, Xu Y, Wang L, Chen H, et al. Human endometrial stem cells confer enhanced myocardial salvage and regeneration by paracrine mechanisms. J Cell Mol Med. 2013;17:1247–60.
Li B, Zeng Q, Wang H, Shao S, Mao X, Zhang F, et al. Adipose tissue stromal cells transplantation in rats of acute myocardial infarction. Coron Artery Dis. 2007;18:221–7.
Sadat S, Gehmert S, Song Y-H, Yen Y, Bai X, Gaiser S, et al. The cardioprotective effect of mesenchymal stem cells is mediated by IGF-I and VEGF. Biochem Biophys Res Commun. 2007;363:674–9.
Madonna R, Geng Y-J, Caterina RD. Adipose tissue-derived stem cells characterization and potential for cardiovascular repair. Arterioscler Thromb Vasc Biol. 2009;29:1723–9.
Nagaya N, Fujii T, Iwase T, Ohgushi H, Itoh T, Uematsu M, et al. Intravenous administration of mesenchymal stem cells improves cardiac function in rats with acute myocardial infarction through angiogenesis and myogenesis. Am J Physiol Heart Circ Physiol. 2004;287:H2670–6.
Rehman J, Traktuev D, Li J, Merfeld-Clauss S, Temm-Grove CJ, Bovenkerk JE, et al. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation. 2004;109:1292–8.
Schenke-Layland K, Strem BM, Jordan MC, DeEmedio MT, Hedrick MH, Roos KP, et al. Adipose tissue-derived cells improve cardiac function following myocardial infarction. J Surg Res. 2009;153:217–23.
Miyahara Y, Nagaya N, Kataoka M, Yanagawa B, Tanaka K, Hao H, et al. Monolayered mesenchymal stem cells repair scarred myocardium after myocardial infarction. Nat Med. 2006;12:459–65.
Uemura R, Xu M, Ahmad N, Ashraf M. Bone marrow stem cells prevent left ventricular remodeling of ischemic heart through paracrine signaling. Circ Res. 2006;98:1414–21.
Bai X, Alt E. Myocardial regeneration potential of adipose tissue-derived stem cells. Biochem Biophys Res Commun. 2010;401:321–6.
Bai X, Yan Y, Song Y-H, Seidensticker M, Rabinovich B, Metzele R, et al. Both cultured and freshly isolated adipose tissue-derived stem cells enhance cardiac function after acute myocardial infarction. Eur Heart J. 2010;31:489–501.
Gao F, He T, Wang H, Yu S, Yi D, Liu W, et al. A promising strategy for the treatment of ischemic heart disease: mesenchymal stem cell-mediated vascular endothelial growth factor gene transfer in rats. Can J Cardiol. 2007;23:891–8.
Deuse T, Peter C, Fedak PWM, Doyle T, Reichenspurner H, Zimmermann WH, et al. Hepatocyte growth factor or vascular endothelial growth factor gene transfer maximizes mesenchymal stem cell-based myocardial salvage after acute myocardial infarction. Circulation. 2009;120:S247–254.
Sun L, Cui M, Wang Z, Feng X, Mao J, Chen P, et al. Mesenchymal stem cells modified with angiopoietin-1 improve remodeling in a rat model of acute myocardial infarction. Biochem Biophys Res Commun. 2007;357:779–84.
Gnecchi M, He H, Melo LG, Noiseaux N, Morello F, de Boer RA, et al. Early beneficial effects of bone marrow-derived mesenchymal stem cells overexpressing Akt on cardiac metabolism after myocardial infarction. Stem Cells. 2009;27:971–9.
Zhang D, Fan G-C, Zhou X, Zhao T, Pasha Z, Xu M, et al. Over-expression of CXCR4 on mesenchymal stem cells augments myoangiogenesis in the infarcted myocardium. J Mol Cell Cardiol. 2008;44:281–92.
Kearns-Jonker M, Dai W, Gunthart M, Fuentes T, Yeh H-Y, Gerczuk P, Pera M, Mummery C, Kloner RA. Genetically engineered mesenchymal stem cells influence gene expression in donor cardiomyocytes and the recipient heart. J Stem Cell Res Ther. 2012; S1.
Lee RH, Pulin AA, Seo MJ, Kota DJ, Ylostalo J, Larson BL, et al. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell. 2009;5:54–63.
Sousa BR, Parreira RC, Fonseca EA, Amaya MJ, Tonelli FMP, Lacerda SMSN, et al. Human adult stem cells from diverse origins: an overview from multiparametric immunophenotyping to clinical applications. Cytometry A. 2014;85:43–77.
Chen S, Fang W, Qian J, Ye F, Liu Y, Shan S, et al. Improvement of cardiac function after transplantation of autologous bone marrow mesenchymal stem cells in patients with acute myocardial infarction. Chin Med J (Engl). 2004;117:1443–8.
Friis T, Haack-Sørensen M, Mathiasen AB, Ripa RS, Kristoffersen US, Jørgensen E, et al. Mesenchymal stromal cell derived endothelial progenitor treatment in patients with refractory angina. Scand Cardiovasc J. 2011;45:161–8.
Mohyeddin-Bonab M, Mohamad-Hassani M-R, Alimoghaddam K, Sanatkar M, Gasemi M, Mirkhani H, et al. Autologous in vitro expanded mesenchymal stem cell therapy for human old myocardial infarction. Arch Iran Med. 2007;10:467–73.
Williams AR, Trachtenberg B, Velazquez DL, McNiece I, Altman P, Rouy D, et al. Intramyocardial stem cell injection in patients with ischemic cardiomyopathy: functional recovery and reverse remodeling. Circ Res. 2011;108:792–6.
Moodley Y, Atienza D, Manuelpillai U, Samuel CS, Tchongue J, Ilancheran S, et al. Human umbilical cord mesenchymal stem cells reduce fibrosis of bleomycin-induced lung injury. Am J Pathol. 2009;175:303–13.
Ortiz LA, Gambelli F, McBride C, Gaupp D, Baddoo M, Kaminski N, et al. Mesenchymal stem cell engraftment in lung is enhanced in response to bleomycin exposure and ameliorates its fibrotic effects. Proc Natl Acad Sci U S A. 2003;100:8407–11.
Rojas M, Xu J, Woods CR, Mora AL, Spears W, Roman J, et al. Bone marrow-derived mesenchymal stem cells in repair of the injured lung. Am J Respir Cell Mol Biol. 2005;33:145–52.
Gupta N, Su X, Popov B, Lee JW, Serikov V, Matthay MA. Intrapulmonary delivery of bone marrow-derived mesenchymal stem cells improves survival and attenuates endotoxin-induced acute lung injury in mice. J Immunol. 2007;179:1855–63.
Zhen G, Liu H, Gu N, Zhang H, Xu Y, Zhang Z. Mesenchymal stem cells transplantation protects against rat pulmonary emphysema. Front Biosci J Virtual Libr. 2008;13:3415–22.
Akram KM, Samad S, Spiteri MA, Forsyth NR. Mesenchymal stem cells promote alveolar epithelial cell wound repair in vitro through distinct migratory and paracrine mechanisms. Respir Res. 2013;14:9.
Manning E, Pham S, Li S, Vazquez-Padron RI, Mathew J, Ruiz P, et al. Interleukin-10 delivery via mesenchymal stem cells: a novel gene therapy approach to prevent lung ischemia-reperfusion injury. Hum Gene Ther. 2010;21:713–27.
Mei SHJ, McCarter SD, Deng Y, Parker CH, Liles WC, Stewart DJ. Prevention of LPS-induced acute lung injury in mice by mesenchymal stem cells overexpressing angiopoietin 1. PLoS Med. 2007;4, e269.
Xu J, Qu J, Cao L, Sai Y, Chen C, He L, et al. Mesenchymal stem cell-based angiopoietin-1 gene therapy for acute lung injury induced by lipopolysaccharide in mice. J Pathol. 2008;214:472–81.
O’Reilly M, Thébaud B. The promise of stem cells in bronchopulmonary dysplasia. Semin Perinatol. 2013;37:79–84.
Tropea KA, Leder E, Aslam M, Lau AN, Raiser DM, Lee J-H, et al. Bronchioalveolar stem cells increase after mesenchymal stromal cell treatment in a mouse model of bronchopulmonary dysplasia. Am J Physiol Lung Cell Mol Physiol. 2012;302:L829–837.
Zhu Y, Feng X, Abbott J, Fang X, Hao Q, Monsel A, et al. Human mesenchymal stem cell microvesicles for treatment of Escherichia coli endotoxin-induced acute lung injury in mice. Stem Cells. 2014;32:116–25.
Lee C, Mitsialis SA, Aslam M, Vitali SH, Vergadi E, Konstantinou G, et al. Exosomes mediate the cytoprotective action of mesenchymal stromal cells on hypoxia-induced pulmonary hypertension. Circulation. 2012;126:2601–11.
Puglisi MA, Tesori V, Lattanzi W, Piscaglia AC, Gasbarrini GB, D’Ugo DM, et al. Therapeutic implications of mesenchymal stem cells in liver injury. BioMed Res Int. 2011;2011:860578.
Meier RPH, Müller YD, Morel P, Gonelle-Gispert C, Bühler LH. Transplantation of mesenchymal stem cells for the treatment of liver diseases, is there enough evidence? Stem Cell Res. 2013;11:1348–64.
Prasajak P. Mesenchymal stem cells: current clinical applications and therapeutic potential in liver diseases. J Bone Marrow Res. 2014;02:1–9.
Kharaziha P, Hellström PM, Noorinayer B, Farzaneh F, Aghajani K, Jafari F, et al. Improvement of liver function in liver cirrhosis patients after autologous mesenchymal stem cell injection: a phase I-II clinical trial. Eur J Gastroenterol Hepatol. 2009;21:1199–205.
Mohamadnejad M, Alimoghaddam K, Mohyeddin-Bonab M, Bagheri M, Bashtar M, Ghanaati H, et al. Phase 1 trial of autologous bone marrow mesenchymal stem cell transplantation in patients with decompensated liver cirrhosis. Arch Iran Med. 2007;10:459–66.
Zhang Z, Lin H, Shi M, Xu R, Fu J, Lv J, et al. Human umbilical cord mesenchymal stem cells improve liver function and ascites in decompensated liver cirrhosis patients. J Gastroenterol Hepatol. 2012;27:112–20.
Li Q, Zhou X, Shi Y, Li J, Zheng L, Cui L, et al. In vivo tracking and comparison of the therapeutic effects of MSCs and HSCs for liver injury. PLoS One. 2013;8, e62363.
Du Z, Wei C, Yan J, Han B, Zhang M, Peng C, et al. Mesenchymal stem cells overexpressing C-X-C chemokine receptor type 4 improve early liver regeneration of small-for-size liver grafts. Liver Transpl. 2013;19:215–25.
Banas A, Teratani T, Yamamoto Y, Tokuhara M, Takeshita F, Osaki M, et al. Rapid hepatic fate specification of adipose-derived stem cells and their therapeutic potential for liver failure. J Gastroenterol Hepatol. 2009;24:70–7.
di Bonzo LV, Ferrero I, Cravanzola C, Mareschi K, Rustichell D, Novo E, et al. Human mesenchymal stem cells as a two-edged sword in hepatic regenerative medicine: engraftment and hepatocyte differentiation versus profibrogenic potential. Gut. 2008;57:223–31.
Banas A, Teratani T, Yamamoto Y, Tokuhara M, Takeshita F, Osaki M, et al. IFATS Collection: in vivo therapeutic potential of human adipose tissue mesenchymal stem cells after transplantation into mice with liver injury. Stem Cells. 2008;26:2705–12.
Kanazawa H, Fujimoto Y, Teratani T, Iwasaki J, Kasahara N, Negishi K, et al. Bone marrow-derived mesenchymal stem cells ameliorate hepatic ischemia reperfusion injury in a rat model. PLoS One. 2011;6, e19195.
Lin N, Hu K, Chen S, Xie S, Tang Z, Lin J, et al. Nerve growth factor-mediated paracrine regulation of hepatic stellate cells by multipotent mesenchymal stromal cells. Life Sci. 2009;85:291–5.
Parekkadan B, van Poll D, Suganuma K, Carter EA, Berthiaume F, Tilles AW, et al. Mesenchymal stem cell-derived molecules reverse fulminant hepatic failure. PLoS One. 2007;2, e941.
Yan Y, Xu W, Qian H, Si Y, Zhu W, Cao H, et al. Mesenchymal stem cells from human umbilical cords ameliorate mouse hepatic injury in vivo. Liver Int. 2009;29:356–65.
Ishikawa H, Jo J-I, Tabata Y. Liver anti-fibrosis therapy with mesenchymal stem cells secreting hepatocyte growth factor. J Biomater Sci Polym Ed. 2012;23:2259–72.
Higashiyama R, Inagaki Y, Hong YY, Kushida M, Nakao S, Niioka M, et al. Bone marrow–derived cells express matrix metalloproteinases and contribute to regression of liver fibrosis in mice. Hepatology. 2007;45:213–22.
Xagorari A, Siotou E, Yiangou M, Tsolaki E, Bougiouklis D, Sakkas L, et al. Protective effect of mesenchymal stem cell-conditioned medium on hepatic cell apoptosis after acute liver injury. Int J Clin Exp Pathol. 2013;6:831–40.
Cho K, Woo S, Seoh J, Han H, Ryu K. Mesenchymal stem cells restore CCl4-induced liver injury by an antioxidative process. Cell Biol Int. 2012;36:1267–74.
Li H, Zhang B, Lu Y, Jorgensen M, Petersen B, Song S. Adipose tissue-derived mesenchymal stem cell-based liver gene delivery. J Hepatol. 2011;54:930–8.
Tsai P-C, Fu T-W, Chen Y-MA, Ko T-L, Chen T-H, Shih Y-H, et al. The therapeutic potential of human umbilical mesenchymal stem cells from Wharton’s jelly in the treatment of rat liver fibrosis. Liver Transpl. 2009;15:484–95.
Tan CY, Lai RC, Wong W, Dan YY, Lim S-K, Ho HK. Mesenchymal stem cell-derived exosomes promote hepatic regeneration in drug-induced liver injury models. Stem Cell Res Ther. 2014;5:76.
Li T, Yan Y, Wang B, Qian H, Zhang X, Shen L, et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013;22:845–54.
Chhabra P, Brayman KL. Stem cell therapy to cure type 1 diabetes: from hype to hope. Stem Cells Transl Med. 2013;2:328–36.
Liu M, Han ZC. Mesenchymal stem cells: biology and clinical potential in type 1 diabetes therapy. J Cell Mol Med. 2008;12:1155–68.
Lysy PA, Weir GC, Bonner-Weir S. Concise review: pancreas regeneration: recent advances and perspectives. Stem Cells Transl Med. 2012;1:150–9.
Carlsson P-O, Schwarcz E, Korsgren O, Le Blanc K. Preserved β-cell function in type 1 diabetes by mesenchymal stromal cells. Diabetes. 2015;64:587–92.
Boumaza I, Srinivasan S, Witt WT, Feghali-Bostwick C, Dai Y, Garcia-Ocana A, et al. Autologous bone marrow-derived rat mesenchymal stem cells promote PDX-1 and insulin expression in the islets, alter T cell cytokine pattern and preserve regulatory T cells in the periphery and induce sustained normoglycemia. J Autoimmun. 2009;32:33–42.
Ezquer F, Ezquer M, Contador D, Ricca M, Simon V, Conget P. The antidiabetic effect of mesenchymal stem cells is unrelated to their transdifferentiation potential but to their capability to restore Th1/Th2 balance and to modify the pancreatic microenvironment. Stem Cells. 2012;30:1664–74.
Jurewicz M, Yang S, Augello A, Godwin JG, Moore RF, Azzi J, et al. Congenic mesenchymal stem cell therapy reverses hyperglycemia in experimental type 1 diabetes. Diabetes. 2010;59:3139–47.
Lee RH, Seo MJ, Reger RL, Spees JL, Pulin AA, Olson SD, et al. Multipotent stromal cells from human marrow home to and promote repair of pancreatic islets and renal glomeruli in diabetic NOD/scid mice. Proc Natl Acad Sci U S A. 2006;103:17438–43.
Phadnis SM, Joglekar MV, Dalvi MP, Muthyala S, Nair PD, Ghaskadbi SM, et al. Human bone marrow-derived mesenchymal cells differentiate and mature into endocrine pancreatic lineage in vivo. Cytotherapy. 2011;13:279–93.
Davis NE, Hamilton D, Fontaine MJ. Harnessing the immunomodulatory and tissue repair properties of mesenchymal stem cells to restore β cell function. Curr Diab Rep. 2012;12:612–22.
Gao X, Song L, Shen K, Wang H, Qian M, Niu W, et al. Bone marrow mesenchymal stem cells promote the repair of islets from diabetic mice through paracrine actions. Mol Cell Endocrinol. 2014;388:41–50.
Fiorina P, Jurewicz M, Augello A, Vergani A, Dada S, La Rosa S, et al. Immunomodulatory function of bone marrow-derived mesenchymal stem cells in experimental autoimmune type 1 diabetes. J Immunol. 2009;183:993–1004.
Favaro E, Carpanetto A, Lamorte S, Fusco A, Caorsi C, Deregibus MC, et al. Human mesenchymal stem cell-derived microvesicles modulate T cell response to islet antigen glutamic acid decarboxylase in patients with type 1 diabetes. Diabetologia. 2014;57:1664–73.
Limbert C, Päth G, Ebert R, Rothhammer V, Kassem M, Jakob F, et al. PDX1- and NGN3-mediated in vitro reprogramming of human bone marrow-derived mesenchymal stromal cells into pancreatic endocrine lineages. Cytotherapy. 2011;13:802–13.
Moriscot C, de Fraipont F, Richard M-J, Marchand M, Savatier P, Bosco D, et al. Human bone marrow mesenchymal stem cells can express insulin and key transcription factors of the endocrine pancreas developmental pathway upon genetic and/or microenvironmental manipulation in vitro. Stem Cells. 2005;23:594–603.
Chen NKF, Tan SY, Udolph G, Kon OL. Insulin expressed from endogenously active glucose-responsive EGR1 promoter in bone marrow mesenchymal stromal cells as diabetes therapy. Gene Ther. 2010;17:592–605.
Mundra V, Wu H, Mahato RI. Genetically modified human bone marrow derived mesenchymal stem cells for improving the outcome of human islet transplantation. PLoS One. 2013;8, e77591.
Wu H, Lu W, Mahato RI. Mesenchymal stem cells as a gene delivery vehicle for successful islet transplantation. Pharm Res. 2011;28:2098–109.
Si Y, Zhao Y, Hao H, Liu J, Guo Y, Mu Y, et al. Infusion of mesenchymal stem cells ameliorates hyperglycemia in type 2 diabetic rats: identification of a novel role in improving insulin sensitivity. Diabetes. 2012;61:1616–25.
Liu X, Zheng P, Wang X, Dai G, Cheng H, Zhang Z, et al. A preliminary evaluation of efficacy and safety of Wharton’s jelly mesenchymal stem cell transplantation in patients with type 2 diabetes mellitus. Stem Cell Res Ther. 2014;5:57.
Jung KH, Song SU, Yi T, Jeon M, Hong S, Zheng H, et al. Human bone marrow–derived clonal mesenchymal stem cells inhibit inflammation and reduce acute pancreatitis in rats. Gastroenterology. 2011;140:998–1008.e4.
Reinders ME, Fibbe WE, Rabelink TJ. Multipotent mesenchymal stromal cell therapy in renal disease and kidney transplantation. Nephrol Dial Transplant. 2010;25:17–24.
Burton CJ, Combe C, Walls J, Harris KP. Secretion of chemokines and cytokines by human tubular epithelial cells in response to proteins. Nephrol Dial Transplant. 1999;14:2628–33.
Tögel F, Isaac J, Hu Z, Weiss K, Westenfelder C. Renal SDF-1 signals mobilization and homing of CXCR4-positive cells to the kidney after ischemic injury. Kidney Int. 2005;67:1772–84.
Herrera MB, Bussolati B, Bruno S, Morando L, Mauriello-Romanazzi G, Sanavio F, et al. Exogenous mesenchymal stem cells localize to the kidney by means of CD44 following acute tubular injury. Kidney Int. 2007;72:430–41.
Chen J, Park H-C, Addabbo F, Ni J, Pelger E, Li H, et al. Kidney-derived mesenchymal stem cells contribute to vasculogenesis, angiogenesis and endothelial repair. Kidney Int. 2008;74:879–89.
Tögel F, Hu Z, Weiss K, Isaac J, Lange C, Westenfelder C. Administered mesenchymal stem cells protect against ischemic acute renal failure through differentiation-independent mechanisms. Am J Physiol Ren Physiol. 2005;289:F31–42.
Xing L, Cui R, Peng L, Ma J, Chen X, Xie R-J, et al. Mesenchymal stem cells, not conditioned medium, contribute to kidney repair after ischemia-reperfusion injury. Stem Cell Res Ther. 2014;5:101.
Westenfelder C, Tögel FE. Protective actions of administered mesenchymal stem cells in acute kidney injury: relevance to clinical trials. Kidney Int Suppl. 2011;1:103–6.
Bianchi F, Sala E, Donadei C, Capelli I, La Manna G. Potential advantages of acute kidney injury management by mesenchymal stem cells. World J Stem Cells. 2014;6:644–50.
Imberti B, Morigi M, Tomasoni S, Rota C, Corna D, Longaretti L, et al. Insulin-like growth factor-1 sustains stem cell–mediated renal repair. J Am Soc Nephrol. 2007;18:2921–8.
Bi B, Schmitt R, Israilova M, Nishio H, Cantley LG. Stromal cells protect against acute tubular injury via an endocrine effect. J Am Soc Nephrol. 2007;18:2486–96.
Hopkins C, Li J, Rae F, Little M. Stem cell options for kidney disease. J Pathol. 2009;217:265–81.
Kucic T, Copland IB, Cuerquis J, Coutu DL, Chalifour LE, Gagnon RF, et al. Mesenchymal stromal cells genetically engineered to overexpress IGF-I enhance cell-based gene therapy of renal failure-induced anemia. Am J Physiol Renal Physiol. 2008;295:F488–496.
Zhen-Qiang F, Bing-Wei Y, Yong-Liang L, Xiang-Wei W, Shan-Hong Y, Yuan-Ning Z, et al. Localized expression of human BMP-7 by BM-MSCs enhances renal repair in an in vivo model of ischemia-reperfusion injury. Genes Cells. 2012;17:53–64.
Morigi M, Rota C, Montemurro T, Montelatici E, Lo Cicero V, Imberti B, et al. Life-sparing effect of human cord blood-mesenchymal stem cells in experimental acute kidney injury. Stem Cells. 2010;28:513–22.
Hagiwara M, Shen B, Chao L, Chao J. Kallikrein-modified mesenchymal stem cell implantation provides enhanced protection against acute ischemic kidney injury by inhibiting apoptosis and inflammation. Hum Gene Ther. 2008;19:807–19.
Bruno S, Grange C, Deregibus MC, Calogero RA, Saviozzi S, Collino F, et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J Am Soc Nephrol. 2009;20:1053–67.
Gatti S, Bruno S, Deregibus MC, Sordi A, Cantaluppi V, Tetta C, et al. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia–reperfusion-induced acute and chronic kidney injury. Nephrol Dial Transplant. 2011;26:1474–83.
Bruno S, Grange C, Collino F, Deregibus MC, Cantaluppi V, Biancone L, et al. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS One. 2012;7, e33115.
Zhou Y, Xu H, Xu W, Wang B, Wu H, Tao Y, et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against cisplatin-induced renal oxidative stress and apoptosis in vivo and in vitro. Stem Cell Res Ther. 2013;4:34.
Franceschi RT, Yang S, Rutherford RB, Krebsbach PH, Zhao M, Wang D. Gene therapy approaches for bone regeneration. Cells Tissues Organs. 2004;176:95–108.
Liu J, Chen W, Zhao Z, Xu HHK. Reprogramming of mesenchymal stem cells derived from iPSCs seeded on biofunctionalized calcium phosphate scaffold for bone engineering. Biomaterials. 2013;34:7862–72.
Jethva R, Otsuru S, Dominici M, Horwitz EM. Cell therapy for disorders of bone. Cytotherapy. 2009;11:3–17.
Marolt D, Knezevic M, Novakovic GV. Bone tissue engineering with human stem cells. Stem Cell Res Ther. 2010;1:10.
Xiao Y, Mareddy S, Crawford R. Clonal characterization of bone marrow derived stem cells and their application for bone regeneration. Int J Oral Sci. 2010;2:127–35.
Wang EA, Rosen V, D’Alessandro JS, Bauduy M, Cordes P, Harada T, et al. Recombinant human bone morphogenetic protein induces bone formation. Proc Natl Acad Sci U S A. 1990;87:2220–4.
Wozney JM, Rosen V, Celeste AJ, Mitsock LM, Whitters MJ, Kriz RW, et al. Novel regulators of bone formation: molecular clones and activities. Science. 1988;242:1528–34.
Chen D, Zhao M, Mundy GR. Bone morphogenetic proteins. Growth Factors. 2004;22:233–41.
Hong D, Chen H-X, Ge R, Li J-C. Genetically engineered mesenchymal stem cells: the ongoing research for bone tissue engineering. Anat Rec Hoboken 2007. 2010;293:531–7.
Candini O, Spano C, Murgia A, Grisendi G, Veronesi E, Piccinno MS, et al. Mesenchymal progenitors aging highlights a miR-196 switch targeting HOXB7 as master regulator of proliferation and osteogenesis. Stem Cells. 2015;33:939–50.
Otsuru S, Gordon PL, Shimono K, Jethva R, Marino R, Phillips CL, et al. Transplanted bone marrow mononuclear cells and MSCs impart clinical benefit to children with osteogenesis imperfecta through different mechanisms. Blood. 2012;120:1933–41.
Gómez-Barrena E, Rosset P, Müller I, Giordano R, Bunu C, Layrolle P, et al. Bone regeneration: stem cell therapies and clinical studies in orthopaedics and traumatology. J Cell Mol Med. 2011;15:1266–86.
Qin Y, Guan J, Zhang C. Mesenchymal stem cells: mechanisms and role in bone regeneration. Postgrad Med J. 2014;90:643–7.
O’Driscoll SW. The healing and regeneration of articular cartilage. J Bone Joint Surg Am. 1998;80:1795–812.
Augello A, Tasso R, Negrini SM, Cancedda R, Pennesi G. Cell therapy using allogeneic bone marrow mesenchymal stem cells prevents tissue damage in collagen-induced arthritis. Arthritis Rheum. 2007;56:1175–86.
Kastrinaki M-C, Papadaki H. Mesenchymal stromal cells in rheumatoid arthritis: biological properties and clinical applications. Curr Stem Cell Res Ther. 2009;4:61–9.
Greish S. Human umbilical cord mesenchymal stem cells as treatment of adjuvant rheumatoid arthritis in a rat model. World J Stem Cells. 2012;4:101.
Mao F, Xu W-R, Qian H, Zhu W, Yan Y-M, Shao Q-X, et al. Immunosuppressive effects of mesenchymal stem cells in collagen-induced mouse arthritis. Inflamm Res. 2010;59:219–25.
Norambuena GA, Khoury M, Jorgensen C. Mesenchymal stem cells in osteoarticular pediatric diseases: an update. Pediatr Res. 2012;71:452–8.
Wakitani S, Okabe T, Horibe S, Mitsuoka T, Saito M, Koyama T, et al. Safety of autologous bone marrow-derived mesenchymal stem cell transplantation for cartilage repair in 41 patients with 45 joints followed for up to 11 years and 5 months. J Tissue Eng Regen Med. 2011;5:146–50.
Davatchi F, Abdollahi BS, Mohyeddin M, Shahram F, Nikbin B. Mesenchymal stem cell therapy for knee osteoarthritis. Preliminary report of four patients. Int J Rheum Dis. 2011;14:211–5.
Davatchi F, Sadeghi Abdollahi B, Mohyeddin M, Nikbin B: Mesenchymal stem cell therapy for knee osteoarthritis: 5 years follow-up of three patients. Int J Rheum Dis. 2015. doi: 10.1111/1756-185X.12670. [Epub ahead of print].
Madry H, Cucchiarini M. Clinical potential and challenges of using genetically modified cells for articular cartilage repair. Croat Med J. 2011;52:245–61.
Steinert AF, Nöth U, Tuan RS. Concepts in gene therapy for cartilage repair. Injury. 2008;39:S97–113.
Acknowledgements
This work was supported in part by the Italian Ministry of Health “Bando Cellule Staminali 2008” (MD, PP), Regione Emilia Romagna (MD, PP), AIRC IG 2012 n. 12755 (MD), and the Associazione ASEOP (MD, PP).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
ND, FR, GG, GG, CS, OC, SO, FC, PP participated in literature search, wrote the manuscript, and prepared the tables; EHM and MD conceived the manuscript concept, wrote the manuscript, and prepared the tables. All authors read and approved the final manuscript.
Additional files
Additional file 1:
Link 1.1 Overview of the main experimental findings on the impact of wild-type MSC in diseases associated with the central and peripheral nervous system. Link 1.2 Overview of the main experimental findings on the impact of gene-modified MSC in diseases associated with the central and peripheral nervous system. (DOCX 34 kb)
Additional file 2:
Link 2.1 Overview of the main pre-clinical findings on the impact of wild-type MSC in heart diseases. Link 2.2 Overview of the main pre-clinical findings on the impact of gene-modified MSC in heart diseases. (DOCX 39 kb)
Additional file 3:
Link 3.1 Overview of the main pre-clinical findings on the impact of wild-type MSC in lung diseases. Link 3.2 Overview of the main pre-clinical findings on the impact of gene-modified MSC in lung diseases. (DOCX 24 kb)
Additional file 4:
Link 4.1 Overview of the main pre-clinical findings on the impact of wild-type MSC in liver diseases. Link 4.2 Overview of the main pre-clinical findings on the impact of gene-modified MSC in liver diseases. (DOCX 22 kb)
Additional file 5:
Link 5.1 Overview of the main pre-clinical findings on the impact of wild-type MSC in pancreatic diseases. Link 5.2 Overview of the main pre-clinical findings on the impact of gene-modified MSC in pancreatic diseases. (DOCX 20 kb)
Additional file 6:
Link 6.1 Overview of the main pre-clinical findings on the impact of wild-type MSC in renal diseases. Link 6.2 Overview of the main pre-clinical findings on the impact of gene-modified MSC in renal diseases. (DOCX 29 kb)
Additional file 7:
Link 7.1 Overview of the main pre-clinical findings on the impact of wild-type MSC in skeletal diseases. Link 7.2 Overview of the main pre-clinical findings on the impact of gene-modified MSC in skeletal diseases. (DOCX 24 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
D’souza, N., Rossignoli, F., Golinelli, G. et al. Mesenchymal stem/stromal cells as a delivery platform in cell and gene therapies. BMC Med 13, 186 (2015). https://doi.org/10.1186/s12916-015-0426-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12916-015-0426-0