
Abstract
Background
Tianhuang formula (THF) is a Chinese medicine prescription that is patented and clinically approved, and has been shown to improve energy metabolism, but the underlying mechanism remains poorly understood. The purpose of this study is to clarify the potential mechanisms of THF in the treatment of type 2 diabetes mellitus (T2DM).
Methods
A murine model of T2DM was induced by high-fat diet (HFD) feeding combined with low-dose streptozocin (STZ) injections, and the diabetic mice were treated with THF by gavaging for consecutive 10 weeks. Fasting blood glucose (FBG), serum insulin, blood lipid, mitochondrial Ca2+ (mCa2+) levels and mitochondrial membrane potential (MMP), as well as ATP production were analyzed. The target genes and proteins expression of visceral adipose tissue (Vat) was tested by RT-PCR and western blot, respectively. The underlying mechanism of the regulating energy metabolism effect of THF was further explored in the insulin resistance model of 3T3-L1 adipocytes cultured with dexamethasone (DXM).
Results
THF restored impaired glucose tolerance and insulin resistance in diabetic mice. Serum levels of lipids were significantly decreased, as well as fasting blood glucose and insulin in THF-treated mice. THF regulated mCa2+ uptake, increased MMP and ATP content in VAT. THF increased the mRNA and protein expression of AMPK, phosphorylated AMPK (p-AMPK), MICU1, sirtuin1 (SIRT1) and peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α). THF could increase the mCa2+ level of 3T3-L1 adipocytes and regulate mitochondrial function. The protein expression of AMPK, p-AMPK, mCa2+ uniporter (MCU) and MICU1 decreased upon adding AMPK inhibitor compound C to 3T3-L1 adipocytes and the protein expression of MCU and MICU1 decreased upon adding the MCU inhibitor ruthenium red.
Conclusions
These results demonstrated that THF ameliorated glucose and lipid metabolism disorders in T2DM mice through the improvement of AMPK/MICU1 pathway-dependent mitochondrial function in adipose tissue.
Highlights
• AMPK-MCU/MICU1 mediated mCa2+ uptake in adipose tissue played an important role in insulin resistance
• THF restored impaired glucose tolerance and insulin resistance in diabetic mice induced by HFD/STZ
• THF improved the mitochondrial function of in T2DM mice via AMPK-MICU1 pathway
Similar content being viewed by others

Background
Diabetes mellitus (DM) is a chronic and lifelong illness with growing incidence globally and is characterized by inadequate insulin secretion [1]. Mitochondria are known as cellular ‘power plants’ [2], their dysfunction plays a pivotal role in the processes of diabetes [3], and can lead to a variety of medical problems including neurodegenerative diseases, cardiovascular diseases, cancer, and metabolic disorders. The mitochondrial energy metabolism disorder will be further aggravated during the development of diabetes. They have an influence on each other through common or different mechanisms, and even form a vicious circle. Mitochondrial dysfunction, caused by mCa2+ homeostasis imbalance under a high glucose environment, may be one of the important pathological mechanisms of diabetes and its complications [4]. The mCa2+ transport is a complex and strictly controlled process. Mitochondrial calcium uniporter (MCU) complex has been identified as a major channel located on the inner membrane to regulate Ca2+ transport into mitochondria [5]. Silencing MCU in cultured cells or in vivo in mouse liver severely abrogates mitochondrial Ca2+ uptake [6]. And there is growing evidence that MICU1 is a gatekeeper of MCU-mediated mCa2+ uptake that is essential to prevent mCa2+ overload and associated stress [7]. Therefore, MCU/MICU1 may play a fundamental role in mediating mCa2+ homeostasis in T2DM, and may represent a novel therapeutic target for T2DM.
Insulin resistance (IR) is the principal pathogenesis of T2DM as an initiating risk factor. The main target organs for insulin action are the liver, the muscle and the adipose tissue. Adipose tissue is a dynamic endocrine organ and nutrient sensor that tightly regulates energy supply. Such as insulin can maintain glucose homeostasis by stimulating glucose uptake in adipose tissue and reducing hepatic gluconeogenesis. The recent trend in research on the mechanism of adipose tissue leading to T2DM mainly focuses on specific adipokines, inflammation and metabolism, that is, adipose tissue dysfunction, which promotes inflammation, hyperlipidemia and IR, and finally results in T2DM [8, 9]. However, there are only very limited reports on how mCa2+ homeostasis in adipose tissue affects T2DM. Mitochondria play a fundamental role in maintaining the balance of energy homeostasis in metabolic tissues, including adipose tissues, and participate in the regulation of energy metabolism homeostasis through fatty acid oxidation [10], oxidative phosphorylation (OXPHOS) [11] and mCa2+ uptake [12]. The AMPK is a sensor of cellular energy status that regulates cellular and whole-body energy balance. During cell mitosis, AMPK mediates the phosphorylation on the 57th serine of MCU to promote the large amount of Ca2+ to enter the mitochondria and stimulate ATP produced to change the state of insufficient cell energy [13]. However, the current research evidence is still insufficient to prove that AMPK, MCU and MICU1 are involved in the regulation of mCa2+ uptake in diabetic adipose tissue.
The Traditional Chinese Medicine Fufang Zhenshu Tiaozhi Formula (FTZ), was developed based on Prof. Jiao Guo's 30 years of clinical experience, and has now been developed into a hospital preparation. Studies showed that FTZ has been well-documented with significant clinical curative effects for hyperglycemia and hyperlipidemia [14, 15]. FTZ has the effect of controlling blood sugar and keeping patients' blood sugar stable for a long time [16], reducing the serum levels of TC, TG, LDL-C and Non-HDL-C in patients [17]. Because of its excellent cost-effective properties, FTZ capsules have been covered by health insurance in the Guangdong Province of China. In clinical and experimental studies over the past few years, a simpler but equivalent formula originated from the FTZ has formed, named Tianhuang formula (THF), which is a patented and clinically approved Chinese medicinal prescription composed of Radix Notoginseng(Panax notoginseng (Burkill) F.H.Chen ex C.Y.Wu & K.M.Feng) and Rhizoma Coptidis (Coptis chinensis Franch). Previous studies have shown THF could regulate lipid metabolism disorders through the gut microbiota-TβMCA-FXR metabolism axis [18], and improve the insulin resistance and glucose intolerance of obese rats induced by high fat and sugar diets [19]. In addition, Radix Notoginseng and Rhizoma Coptidis are both traditional herbal drugs with hundreds of years of usage. A large number of studies have shown that Radix Notoginseng and Rhizoma Coptidis have anti-hyperglycemic activity [20,21,22]. In conclusion, THF has the potential to improve glucose metabolism, but its mechanism has not been fully elucidated.
This study demonstrated that the AMPK-MICU1 pathway mediated mCa2+ uptake, affected mitochondrial function, and caused an imbalance in energy metabolism, further leading to glucose and lipid metabolism disorders in Vat of STZ-induced T2DM mice. THF attenuated T2DM by regulating mCa2+ uptake in Vat which mechanism may be mediated through the AMPK-MICU1 pathway. These data addressed key gaps in our understanding of the AMPK-MICU1 and shed light on the role MICU1 plays in T2DM.
Methods
Herbal materials
Herbs in THF ( Radix Notoginseng and Rhizoma Coptidis) were provided by Zhixin Chinese Herbal Medicine Co., Ltd. (Guangzhou, China. S). Prof. Jiao Guo, Guangdong Pharmaceutical University authenticated the plant material using the Pharmacopeia of the People's Republic of China identification key (ISBN 2020, volume I). The production batch numbers were 210,401 and 210,501. Plant names have been checked with http://www.theplantlist.org.
Preparation and chemical constituents of THF
THF was prepared as follows [19], powdered Radix Notoginseng (400 g) and Rhizoma Coptidis (400 g) were separately extracted triply with 70% ethanol at 80 °C under reflux, each time for 2 h. The extract solution was concentrated in a rotary evaporator to remove ethanol, and then dissolved in water and purified using D101 macro-porous resin (Lanxiao, Xi’an). The resulting purified extract was dried in a vacuum at 60 °C. The quantitative profiling of THF was performed on a U3000 HPLC with a DAD detector (Dionex, USA). The chromatography separation was carried out using a Kromasil C18 column (4.5 × 250 mm, 5 μm in particle size) according to the Pharmacopoeia of the People’s Republic of China (2020), and data were recorded and analyzed on the Chromeleon Console workstation. Finally, the content of eight active components in THF, namely, Ginsenoside Rg1, Ginsenoside Rb1, Ginsenoside Rd, Ginsenoside Re, Notoginsenoside R1, Berberine, Coptisine, and Palmatine, were quantified.
Animals and management ethics protocol
All the animal experiments were approved by the Animal Ethical Committee of Guangdong Pharmaceutical University (SPF2017310). Specific pathogen free (SPF) male C57BL/6 J Narl mice 3–4 weeks of age were purchased from Guangdong Medical Laboratory Animal Center. All mice were housed in a temperature-controlled room at 24 °C ± 2 °C, with a humidity of 60%-70%, and 12 h of light and darkness alternated, standard solid food and water were provided during the experiment.
Induction of hyperglycemia in experimental animals
Mice were divided into two groups, normal and diabetic groups. To induce diabetes, mice were fed HFD (60% fat, 20% protein, 20% carbohydrate, Research Diets, D12492) for 4 weeks [23]. Then, the mice were fasted for 6 h followed by the administration of intraperitoneal (ip) injection of 40 mg/kg STZ for 4 consecutive days based on the previous study by Gilbert ER et al. [24]. The STZ (Sigma, St. Louis, MO, USA) was dissolved in citrate buffer (0.05 M, pH4.5), which was freshly prepared before use. Blood glucose level was checked using an Accu-Chek blood glucometer (Roche Diagnostics, Basel, Switzerland) every 72 h. Stable hyperglycemia was established if the fasting blood glucose (FBG) was ≥ 11.1 mmol/L in the tested animals after 1 week from the STZ injection. Normal mice received i.p. injection of citrate buffer.
Mice were allocated randomly after 1 week from diabetes induction into three groups (n = 8) whereas the normal mice were allocated randomly after 1 week from ip injection of citrate buffer (n = 8) and treated daily as follows for 6 weeks. Control group: normal mice received oral normal saline only. Model group: animals were fed HFD and received STZ. High-dose THF (THF-H) group: HFD/STZ diabetic mice were treated with THF (120 mg/kg/day). Low-dose THF (THF-L) group: HFD/STZ diabetic mice were treated with THF (60 mg/kg/day) [19]. Metformin (MET) group: HFD/STZ diabetic mice were administered MET (250 mg/kg).
The body weight and food intake of the mice were assessed once per week. The fat mass was detected in the last week of the experiment using the Minispec LF90 Body Composition Analyzer (Bruker). For the oral glucose tolerance test (OGTT), mice were given oral glucose (2 g/kg) after fasting for 6 h to measure their glucose tolerance, and the blood glucose levels at different time points were detected using the Accu-Chek blood glucometer immediately after the initial injection of glucose. The areas under the curves (AUC) of the glucose level over time were calculated to evaluate the glucose tolerance ability of the mice [25].
After all the experiments, all mice were euthanized with 200 mg/kg pentobarbital sodium through intraperitoneal injection, blood samples were collected from the mice’s orbital venous plexus and transferred into a collection tube, which were then centrifuged at 3500 rpm for 30 min at 4 °C, serum samples were prepared and kept at -80 °C [19]. The TC, TG, APN and insulin levels in serum were measured using commercial kits according to the manufacturers' introductions. Adipose tissue was collected and stored at -80 °C until subsequent biochemical analyses and was fixed in 10% buffered formalin for histopathological examination [25]. Animal bodies were taken care of by the Animal Ethical Committee of Guangdong Pharmaceutical University.
Histopathological screening
Adipose tissue specimens were fixed in 10% formalin for 24 h, dehydrated using gradual ethanol concentrations, and embedded in paraffin. Then, the paraffin- embedded specimens were sectioned into 5 μm thick sections and stained with hematoxylin and eosin (H&E)[25]. The adipose tissue (1 mm × 1 mm × 1 mm) was placed in a 2.5% glutaraldehyde fixative solution, fixed overnight at 4 °C, dehydrated, and observed under an electron microscope. Slides were examined under a light microscope (magnification: × 200, Eclipse E200-LED, Nikon, Tokyo, Japan).
Metabolic rate measurements
The mice’s metabolism was evaluated by the Comprehensive Lab Animal Monitoring System (CLAMS, Columbus Instruments) according to the manufacturer's instructions. The mice were placed in individual cages and acclimated to the monitoring system for 24 h. The metabolic rate was evaluated by their carbon dioxide production (VCO2), oxygen consumption (VO2) and heat production over the next 24 h, which were analyzed with the CLAX Research software package (CLAX Research; CLAMS, Columbus Instruments)[25].
Determination of mitochondrial Ca2+, MMP and ATP content in E-Wat and 3T3-L1 adipocyte
Fresh visceral adipose tissue (100 mg) was washed with PBS, then cutted into pieces and added 10 times the amount of pre-cooled mitochondrial separation reagent A, homogenized at low temperature, centrifuged to obtain the precipitate. Then the separated mitochondria in epididymal white adipose tissue (E-Wat) and 3T3-L1 adipocytes were added to the mitochondrial storage solution to detect the contents of mCa2+, MMP and ATP (Beyotime Biotechnology Co., Ltd., Shanghai, China) according to the mitochondrial membrane potential detection kit instructions.
Cell culture and induction of adipocyte differentiation
3T3-L1 preadipocytes were cultured and differentiated into adipocytes by using a previously reported method [26]. Briefly, 3T3-L1 preadipocytes were cultured in DMEM containing 10% bovine calf serum at 37 °C in a 5% CO2 incubator. To induce differentiation, 2-day post-confluent preadipocytes were incubated for 2 days in a differentiation medium containing 10% FBS, 0.5 mM IBMX, 1 μM dexamethasone, and l μg/mL insulin. The medium was then changed to DMEM containing 10% FBS and 1 μg/mL insulin, and cells were cultured for another 2 days. Then cells were incubated in DMEM supplemented with 10% FBS for 2 more days.
RNA extraction and quantitative real-time PCR
Total RNA was extracted using the Trizol reagent (Takara, Dalian, China). The RNA was transcribed into cDNA using a reverse transcription kit (Takara, Dalian, China) according to the manufacturer's instructions, and quantitative real-time PCR was performed using a Light Cycler 480 real-time PCR system (Roche, Switzerland). β- actin was used as an internal control to normalize expression values. The sequences of the PCR primers were listed in Supporting Table 1.
Protein extraction and Western blotting analysis
Western blotting analysis was performed according to the protocol as previously reported [16]. Protein was extracted by RIPA lysis buffer (P1003B, Beyotime, China) and protein concentration was detected by the BCA kit (P0011, Beyotime, China). The primary antibodies were incubated at 4°Covernight, and the information of the brand and item number of the primary antibody were listed in Supporting Table 2. After incubating proper secondary antibodies, the protein bands were visualized by enhanced chemiluminescence (ECL) kit (Bio-Rad, CA, USA) and quantified by Image Pro Plus software.
Statistical analysis
Data were presented as means ± standard error of mean (SEM). Data sets that involved more than two groups were assessed by one-way ANOVA followed by Newman-Keuls post hoc tests. p < 0.05 was considered statistically significant. GraphPad Prism 6.0 software (GraphPad, CA, USA) was used for statistical analysis and graphics.
Result
Preparation and quantitative profiling of THF
The results of the HPLC fingerprint chromatogram showed that THF was made of Ginsenoside Rg1 (23.82%), Berberine (17.01%), Palmatine (5.11%), Ginsenoside Rb1 (4.58%), Coptisine (4.45%), Panax notoginseng saponin R1 (2.05%), Ginsenoside Re (1.03%), Ginsenoside Rd (0.97%) and some other unidentified components (41.98%) (Fig. 1).

Preparation and quantitative profiling of Tianhuang formula. HPLC fingerprint chromatograms of the extracts of the reference standards (1) Ginsenoside Rg1 (CAS:), (2) Berberine (CAS:2086–83-1), (3) Palmatine (CAS:3486–67-7), (4) Ginsenoside Rb1 (CAS:41,753–43-9), (5) Coptisine (CAS: 3486–66-6), (6) Panax notoginseng saponin R1 (CAS:80,418–24-2), (7) Ginsenoside Re (CAS:52,286–59-6), (8) Ginsenoside Rd (CAS:52,705–93-8)
THF improved the glucose and lipid metabolism in T2DM mice
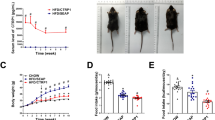
Notably, there was no difference in food intake (Fig. 2A) between the model group and THF group. Body weights as shown in Fig. 2B-C, which were lower in THF group mice relative to model group mice. Compared with the control group, blood glucose levels were significantly increased in the model group and decreased in the THF treatment group (Fig. 2D). The AUC of OGTT results showed significant deterioration in glucose tolerance in the model group. Except for the control group, the glucose levels began to rise after oral glucose, peaking within 15 min, before gradually returning to the initial level. At 60 min, there was a significant difference between the model group and the Met group. After 90 min, glucose levels were significantly reduced in THF-H group (Fig. 2E and F). Compared with the control group, the fasting serum insulin level and IR index of the model group increased significantly, which was alleviated by metformin or THF treatment (Fig. 2G and H).

Comparison of indices during the construction of the HFD/STZ-induced T2DM mice and the treatment effects of THF. A Food intake. B-C Body weight. D Blood glucose. E–F OGTT and AUC analysis of OGTT. G Serum insulin. H HOMA-IR index. I Bat, Sat and Vat weigh. J MRI analysis of lean and fat mass. K Nonestesterified fatty acid (NEFA). L Triglyceride. M Total cholesterol. N Low-density lipoprotein. O High-density lipoprotein. P Adiponectin (APN). Notes:*P < 0.05, **P < 0.01, ***P < 0.001 vs Control, #P < 0.05, ##P < 0.01, ###P < 0.001 vs Model. All the data were presented as the means ± SEMs (n = 6–8)
There were no differences in the weight of brown adipose and subcutaneous adipose among the groups. However, compared with the control group, the weight of E-Wat increased significantly in the model group and decreased significantly in the THF-H group (Fig. 2I). The body fat and the Fat/Weight ratio increased significantly and the Lean/Weight ratio showed a downward trend in the model group, which was significantly adjusted by metformin or THF treatment. It was speculated that the effect of THF on improving glucose and lipid metabolism disorders may be related to E-Wat dysfunction (Fig. 2J).
Furthermore, the nonestesterified fatty acid (NEFA), TC, LDL-C and TG levels in the serum of the model group mice significantly increased compared to the control group, while metformin or THF decreased the serum levels of NEFA, TC, LDL-C and TG (Fig. 2K-N). Serum levels of HDL-C and APN were decreased in HFD/STZ induced mice, while resumed after metformin or THF treatment (Fig. 2O), and the improvement of APN in the THF-H group was significantly better than in the other groups (Fig. 2P).
THF improved the mitochondrial function of VAT in T2DM mice
Histologically, H&E revealed that the brown adipocytes (Bat) were hypertrophy and larger lipid droplets were formed in the adipocytes, and the number of lipid droplets was significantly increased in the model group (Fig. 3A). The visceral adipose cells were full of lipid droplets challenged with the HFD/STZ-induced (Fig. 3B). Observation of E-Wat by electron microscope showed that adipocyte was unclear, the mitochondria were swollen, and the cristae were difficult to distinguish in the model group, whereas THF and metformin treatment were capable to repress those histopathological signatures (Fig. 3C).

THF-treated mice exhibited improved mitochondrial structure. A Representative images of H&E-stained Bat of T2DM mice. B Representative images of H&E-stained epididymal E-Wat of T2DM mice. C Representative images of electron microscopy of E-Wat. D Oxygen consumption (VO2). E Carbon dioxide (VCO2). F-G VCO2/VO2 and AUC analysis of VCO2/VO2. H Mitochondrial Ca2+ levels of E-Wat. I Mitochondrial membrane potential MMP levels of E-Wat. J Relative ATP levels of E-Wat. Notes:*P < 0.05, **P < 0.01, ***P < 0.001 vs Control, #P < 0.05, ##P < 0.01, ###P < 0.001 vs Model. All the data were presented as the means ± SEMs (n = 6–8). Scale: 50 μm
We used CLAMS to determine energy expenditure, and found that THF increased oxygen consumption (VO2) and carbon dioxide production (VCO2) during a 12-h cycle of light and dark. Systemic energy expenditure was significantly increased in THF and Met groups (Fig. 3D-G).
The mitochondrial function of Vat was further tested to investigate whether improvement of glucose and lipid metabolism following THF-treated was related to mCa2+ homeostasis in adipose tissue. The mCa2+ level of E-Wat was significantly reduced in the model group, which was increased significantly by THF treatment. And the levels of MMP and ATP decreased in T2DM mice. However, both metformin and THF-H treatments showed significant improvements (Fig. 3H-J). The mitochondrial function of Wat was impaired in a high-glucose environment and THF attenuated it.
THF improved the mitochondrial function of E-Wat in T2DM mice through the AMPK-MICU1 pathway
The results of the above experiments showed that THF-H improved T2D better than THF-L, indicating that THF improved T2D in a dose-dependent manner, to further investigate whether the protective mechanism of THF on mitochondrial function was linked to the AMPK-MICU1 pathway, we analyzed the mRNA expression levels of energy metabolism-related genes (AMPK, AMPKα, SIRT1, PGC-1α), MCU and MICU1, which decreased in the E-Wat of each group. Notably, MCU mRNA expression levels were indistinguishable between the control group and the model group. THF-H restored the mRNA expression of AMPK, AMPKα, p-AMPK, SIRT1, PGC-1α, and MICU1 (Fig. 4A-F). In addition, HFD/STZ-induced also significantly decreased mitochondrial function-specific genes expression of COX4, TFAM, UQCRb, NDUFS8, SDHb and COX5b, which further were upregulated by THF-H (Fig. 4G-L).

THF-H promoted mitochondrial energy metabolism-related mRNA expression in epididymal white adipose tissue of HFD/STZ-induced T2DM mice. A-B mRNA expression of AMPKα and AMPK. C-D mRNA expression of MCU and MICU1. E–F mRNA expression of SIRT1 and PGC-1α. G-L mRNA expression of COX4, TFAM, UQCRb, NDUFS8, SDHb and COX5b. Notes:*P < 0.05, **P < 0.01, ***P < 0.001 vs Control, #P < 0.05, ##P < 0.01, ###P < 0.001 vs Model. All the data were presented as the means ± SEMs (n = 4–6)
Further, the protein expression levels of AMPK, p-AMPK, SIRT1, PGC-1α, MCU and MICU1 in the Vat of each group were analyzed and found that SIRT1, PGC-1α, AMPK, p-AMPK, and MICU1 decreased in the model group, while increased after metformin or THF treatment, which were consistent with mRNA expression (Fig. 5A-G).

THF promoted mitochondrial energy metabolism-related protein levels in epididymal white adipose tissue of HFD/STZ-induced T2DM mice. A-G Western blot analyses of SIRT1, PGC-1α, MCU, AMPK, p-AMPK and MICU1 protein levels in Vat. β-actin was used as a loading control. Notes:*P < 0.05, **P < 0.01, ***P < 0.001 vs Control, #P < 0.05, ##P < 0.01, ###P < 0.001 vs Model. All the data were presented as the means ± SEMs (n = 3–5). Full-length blots are presented in Supplementary Figure 5
The above results indicated that the mRNA and protein expression levels of the mitochondrial respiratory chain, energy metabolism-related genes and the Ca2+ channel component protein MICU1 and MCU were reduced in E-Wat by HFD/STZ-induced, indicating that mitochondrial dysfunction and energy metabolism disorder were impaired in the E-Wat of T2DM mice, and THF improved the mitochondrial function of E-Wat in T2DM mice which might be related to AMPK-MICU1 pathway.
THF attenuated IR via AMPK-MICU1 pathway in 3T3-L1 adipocytes
In order to further reveal that THF's improvement of mitochondrial function was related to the AMPK-MICU1 mediated mCa2+ uptaker, we tested mCa2+, MMP and ATP levels, and the protein expression of AMPK, p-AMPK, MCU, MICU1 in 3T3-L1 adipocyte.
Compared to the control group, the model group had significantly higher glucose levels. Simultaneously, the levels of mCa2+, MMP, ATP and the protein expression levels of AMPK, p-AMPK, MCU and MICU1 decreased. The level of mCa2+, MMP and ATP increased significantly after THF treatment. The results in vitro were also consistent with those in vivo.
To further investigate the relationship between AMPK and MCU, 3T3-L1 adipocytes were treated with compound C and ruthenium red, which specifically inhibited the protein expression of AMPK and MCU. The result showed the levels of mCa2+, MMP and ATP decreased, and the protein expression of AMPK, p-AMPK, MICU1 and MCU decreased too, resulting in elevated glucose concentration after compound C treatment. Similar results were found after intervention with ruthenium red, the difference was that there was no significant change in the protein expression of AMPK and p-AMPK, thus demonstrating that IR in 3T3-L1 adipocytes was related to the AMPK-MICU1 pathway, and AMPK was upstream of MICU1.
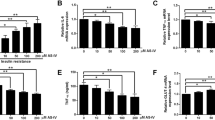
The protein expressions of AMPK, p-AMPK and MICU1 were significantly decreased compared with THF group after compound C and ruthenium red treatment, which further verified THF treatment attenuated IR by regulating the expression of genes and proteins associated via the AMPK-MICU1 pathway in 3T3-L1 adipocytes (Fig. 6A-I).

THF promoted mitochondrial function in 3T3-L1 adipocytes cells induced by dexamethasone. A Glucose. B-D The levels of mCa2+, MMP and ATP in 3T3-L1 adipocytes cells. E-I The protein expression of AMPK, p-AMPK, MICU1 and MCU. β-actin was used as a loading control. Notes:*P < 0.05, **P < 0.01, ***P < 0.001 vs Control, #P < 0.05, ##P < 0.01, ###P < 0.001 vs Model,▲P < 0.05, ▲▲P < 0.01, ▲▲▲P < 0.001 vs THF. All the data were presented as the means ± SEMs (n = 3–5). Full-length blots are presented in Supplementary Figure 6
Discussion
T2DM has a high incidence and serious harm to human health, affects the metabolism of multiple organs and systems throughout the body, and causes a variety of complications [19]. IR is one of the major causes of T2DM [27], mitochondria dysfunction is tightly associated with IR, therefore, protecting mitochondrial function is a feasible approach to attenuate IR. The adipose tissue is one of the target organs for insulin action, however, how mitochondria in adipose tissue affect IR has to be further investigated.
A fundamental function of mitochondria is to produce ATP through oxidative phosphorylation and provides energy to the body. When the structure and function of mitochondria are damaged, it will directly lead to slower oxidative decomposition of glucose and reduce ATP production. Previous studies have found that under IR conditions, the mitochondrial structure is damaged and ATP production is significantly reduced [28]. After mitochondrial oxidative phosphorylation and the level of ATP production is increased, insulin sensitivity is increased and IR is further alleviated [29]. So enhancing mitochondrial function also promote insulin sensitivity, and improve insulin resistance.
Mitochondria play an essential role in the processes of energy production, signal transduction, oxidative stress and so on. Mitochondrial dysfunction is inextricably linked to metabolic diseases such as T2DM [30] through mechanisms including mCa2+ disorder and the decreased activity of the electron transport chain complex [31]. Ca2+ is an important regulatory ion and mitochondrial dysfunction caused by the imbalance of mCa2+ homeostasis is considered to be a significant pathological mechanism of diabetes [32], while MCU/MICU1 is a key molecule that regulates mCa2+ uptake [7]. A long-term high glucose environment will cause the decrease of MCU expression, which lead to the weakening of the uptake of Ca2+ by the mitochondria or the imbalance of mCa2+ homeostasis [33], and futher lead to abnormal mitochondrial electron transport chains and decrease in MMP [4, 34]. Due to the reduction of mCa2+ uptake, the activity of rate-limiting enzymes of the tricarboxylic acid cycle is reduced, and the production of ATP is significantly reduced [35]. Suarez J et al. prove that the genes and proteins expression of MCU is related to mCa2+ concentrations in the heart of diabetic mice [36]. The above-mentioned studies indicate that the MCU and MICU1 act as the main channel for mCa2+ uptake and regulate the uptake of Ca2+ by mitochondria. Experimentally, we found similar phenomena through experiments, in the present study, the genes and proteins expression of MCU and MICU1 decreased, the uptake of mCa2+ was reduced, MMP and ATP were also significantly reduced in the E-Wat of T2DM mice. The genes and proteins expression of MCU and MICU1 in 3T3-L1 adipocytes were also decreased under a high glucose environment. The uptake of mCa2+ decreased, and MMP and ATP were significantly decreased too.
Ca2+, as the second messenger factor, is a pivotal signal in the transmission mechanism of mitochondrial energy activity. In the mitochondrial oxidative respiratory chain complexes, Ca2+ can enhance the activity of oxidative phosphorylation, thereby increasing the production of ATP [37]. At the same time, MCU and MICU1 are major highly selective channels for mCa2+ uptake, and the transportation of Ca2+ depends on the electrochemical gradient of MMP [38]. In a high glucose environment, the MMP of 3T3-L1 adipocytes decreases, forming a vicious cycle [39]. Previous studies have shown that mCa2+ effects membrane potential and ATP production. No research has confirmed whether decreased uptake of mCa2+ in adipocytes in a high-glucose environment leads to abnormal MMP and ATP production. It is unclear whether AMPK participates in the regulation of mCa2+ homeostasis in adipose tissue, thereby affecting the occurrence of diabetes. In the present study, we found systemic energy expenditure was significantly decreased, along with a significant decrease in MMP, ATP and mCa2+ levels in Vat of T2DM mice. The mRNA expression of COX5b, NDUFS8, SDHb, UQCRb and mitochondrial function-specific genes TFAM and COX4 decreased. The mRNA and protein expression levels of AMPK, p-AMPK, MICU1, SIRT1 and PGC-1α decreased. It could be seen that AMPK in Vat was involved in the regulation of mCa2+, increasing mCa2+, MMP and ATP levels.
We additionally found a decrease in protein expression of MCU and MICU1 in 3T3-L1 adipocytes cultured in a high-glucose environment. After adding AMPK inhibitor Compound C and MCU inhibitor ruthenium red, the protein expression of AMPK, p-AMPK, MCU and MICU1 decreased. It was found that the decreased expression of MCU and MICU1 in a high glucose environment evoked inhibition of mCa2+ uptake, MMP and ATP production. Through the results, we found that the down-regulation of MCU and MICU1 mediated mitochondrial dysfunction and led to IR in 3T3-L1 adipocytes cultured in a high-glucose environment. We confirmed that the energy metabolism disorder of T2DM may be related to the mitochondrial dysfunction caused by the decreased expression of MCU and MICU1, and the abnormal mCa2+ uptake caused by the MCU-mediated mitochondrial dysfunction, which may be the pathogenesis of T2DM.
Natural Chinese herbal medicines have been proven to have a wide range of pharmacological effects. Screening drugs that improve insulin sensitivity from natural Chinese herbal medicines may be a potential strategy for T2DM. T2DM belongs to the “Danzhuo” in TCM, which was first put forward by Prof. Jiao Guo from the Guangdong Pharmaceutical University. THF is composed of Radix Notoginseng and Rhizoma Coptidis, based on the theory of traditional Chinese medicine "TiaoGan QiShu HuaZuo". Studies have shown that THF has the effect of improving hepatosteatosis and glucose intolerance in diet-induced obese rats [19], but the underlying mechanism still needs to be revealed. We investigated the effect of THF on HFD/STZ-induced T2DM mice, and found that THF not only regulated glucose metabolism, but also improved insulin sensitivity. Furthermore, APNs are inflammatory cytokines associated with obesity and insulin resistance [25]. THF administration led to significant improvement in ANP level. Thus, these findings suggest that THF exerts protective effects against T2DM. Surprisingly, there were no differences in the weight of brown adipose and subcutaneous adipose among the groups, while the weight of E-Wat increased significantly in the model group and significantly adjusted by metformin or THF treatment, so we speculated that the effect of THF on improving glucose and lipid metabolism disorders may be related to E-Wat dysfunction. We futher found after THF administering, the mCa2+, MMP and ATP increased, and the mRNA and protein expression levels of AMPK, MICU1, SIRT1, PGC1α, and MICU1 also increased in E-Wat of T2DM mice.
To investigate whether THF reduces mCa2+ uptake via AMPK/MICU1 pathway, we then used compound C and ruthenium red, which specifically inhibited the protein expression of AMPK and MCU. The results showed that THF increased the level of mCa2+ in 3T3-L1 adipocytes, regulated mitochondrial function, thereby improving IR, and its effect might be partly achieved by regulating mCa2+ uptake disorders via AMPK/MICU1 pathway.
There were several limitations in the study. First, liver and skeletal muscle also play an important role in mitochondrial energy metabolism, so study the AMPK-MICU1 pathway in the liver and skeletal muscle to regulate mCa2+ uptake disorders is also very valuable. Second, the study only investigated the relationship between AMPK and MICU1 on 3T3-L1 adipocytes, establish a tissue-specific MICU1 gene knockout model for further research may make the conclusion more convincing.
Conclusion
In this study, we verified the effect of MICU1 as an influential regulator of mCa2+ homeostasis in a high-glucose environment. AMPK-MICU1 might be an important pathway for adipocyte energy metabolism disorders under high a glucose environment and was involved in the pathogenesis of T2DM. Furthermore, our study showed that THF treatment attenuated diabetes by regulating adipocyte mitochondrial function by AMPK/MICU1 pathway in vivo and in vitro.
Availability of data and materials
The data used and/or investigated during the present study are accessible from the corresponding author on reasonable request.
Abbreviations
- T2DM:
-
Type 2 diabetes mellitus
- THF:
-
Tianhuang formula
- AMPK:
-
AMP-activated protein kinase
- MICU1:
-
Mitochondrial Ca2+ uptake 1
- mCa2 + :
-
Mitochondrial calcium
- HFD:
-
High fat diet
- STZ:
-
Streptozocin
- FBG:
-
Fasting blood glucose
- VAT:
-
Visceral adipose tissue
- DXM:
-
Dexamethasone
- MMP:
-
Mitochondrial membrane potential
- p-AMPK:
-
Phosphorylated AMPK
- SIRT1:
-
Sirtuin1
- PGC-1α:
-
Peroxisome proliferator-activated receptor-γ coactivator-1α
- MCU:
-
Mitochondrial Ca2+ uniporte
- DM:
-
Diabetes mellitus
- IR:
-
Insulin resistance
- OXPHOS:
-
Oxidative phosphorylation
- ATP:
-
Adenosine triphosphate
- THF-H:
-
High-dose THF
- THF-L:
-
Low-dose THF
- MET:
-
Metformin
- OGTT:
-
Oral glucose tolerance test
- AUC:
-
Area under the curve
- LDL-C:
-
Low density lipoprotein cholesterol
- TC:
-
Total cholesterol
- TG:
-
Triglycerides
- HDL-C:
-
High density lipoprotein cholesterol
- NEFA:
-
Nonestesterified fatty acid
- APN:
-
Adiponectin
- MMP:
-
Mitochondrial membrane potential
- BAT:
-
Brown adipocytes
- VAT:
-
Visceral adipose tissue
References
Magliano DJ, Sacre JW, Harding JL, Gregg EW, Zimmet PZ, Shaw JE. Young-onset type 2 diabetes mellitus-implications for morbidity and mortality. Nat Rev Endocrinol. 2020;16:321–31.
Raimundo N. Mitochondrial pathology: stress signals from the energy factory. Trends Mol Med. 2014;20:282–92.
Rocha M, Apostolova N, Diaz-Rua R, Muntane J, Victor VM. Mitochondria and T2D: role of autophagy, ER stress, and inflammasome. Trends Endocrinol Metab. 2020;31:725–41.
Sorrentino A, Borghetti G, Zhou Y, Cannata A, Meo M, Signore S, Anversa P, Leri A, Goichberg P, Qanud K, Jacobson JT, Hintze TH, Rota M. Hyperglycemia induces defective Ca2+ homeostasis in cardiomyocytes. Am J Physiol Heart Circ Physiol. 2017;312:H150–61.
Wang CH, Wei YH. Role of mitochondrial dysfunction and dysregulation of Ca2+ homeostasis in the pathophysiology of insulin resistance and type 2 diabetes. J Biomed Sci. 2017;24:70.
Baughman JM, Perocchi F, Girgis HS, Plovanich M, Belcher-Timme CA, Sancak Y, Bao XR, Strittmatter L, Goldberger O, Bogorad RL, Koteliansky V, Mootha VK. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature. 2011;476:341–5.
Mallilankaraman K, Doonan P, Cardenas C, Chandramoorthy HC, Muller M, Miller R, Hoffman NE, Gandhirajan RK, Molgo J, Birnbaum MJ, Rothberg BS, Mak DO, Foskett JK, Madesh M. MICU1 is an essential gatekeeper for MCU-mediated mitochondrial Ca2+ uptake that regulates cell survival. Cell. 2012;151:630–44.
Chait A, Den-Hartigh LJ. Adipose tissue distribution, inflammation and its metabolic consequences, including diabetes and cardiovascular disease. Front Cardiovasc Med. 2020;7:22.
Longo M, Zatterale F, Naderi J, Parrillo L, Formisano P, Raciti GA, Beguinot F, Miele C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int J Mol Sci. 2019;20:2358.
Lee J, Ellis JM, Wolfgang MJ. Adipose fatty acid oxidation is required for thermogenesis and potentiates oxidative stress-induced inflammation. Cell Rep. 2015;10:266–79.
Lee JH, Park A, Oh KJ, Lee SC, Kim WK, Bae KH. The role of adipose tissue mitochondria: regulation of mitochondrial function for the treatment of metabolic diseases. Int J Mol Sci. 2019;20:4924.
Arruda AP, Hotamisligil GS. Calcium homeostasis and organelle function in the pathogenesis of obesity and diabetes. Cell Metab. 2015;22:381–97.
Zhao H, Li T, Wang K, Zhao F, Chen J, Xu G, Zhao J, Li T, Chen L, Li L, Xia Q, Zhou T, Li HY, Li AL, Finkel T, Zhang XM, Pan X. AMPK-mediated activation of MCU stimulates mitochondrial Ca2+ entry to promote mitotic progression. Nat Cell Biol. 2019;21:476–86.
PengYJ, Liu XM, Meng ZZ, Zhan HX, Piao SH, Rong XL, Guo J. Zhenzhu Tiaozhi Capsule combined with conventional western medicine in the treatment of 35 elderly patients with type 2 diabetes: a real world pragmatic study. J Tradit Chin Med. 2022;63:43–49.
Huang X, Zhan H, Yang J, Peng L, Piao S, Wang L, Lan T, Rong X, Guo J. Long-term effect of Zhenzhu Tiaozhi Capsule (FTZ) on hyperlipidemia: 2-year results from a retrospective study using electronic medical records. Evid Based Complement Alternat Med. 2021;2021:6264414.
Wu MW, Huang XQ, Zhan HX, Yang J, Peng LF, Liu Q, Jin YH, Chen YY, Xu JJ, Li SY, Mei YZ, Piao SH, Rong XL, Guo J. Effect of Zhenzhu Tiaozhi Capsule on Hb -A1c in patient with type 2 diabetes mellitus: a real world study. Chin J ETMF. 2021;27:110–7.
Chen YY, Jin YH, Zhan HX, Liu Q, Yang J, Peng LF, Li X, Piao SH, Rong XL, Guo J. A real-world study on efficacy and safety of Zhenzhu Tiaozhi Capsule in treating hyperlipidemia. Pharmacol Clin Chin Mat Med. 2022;38:163–7.
Yang L, Chen KC, Luo DS, Guo J. Efficacy and mechanism of Tianhuang Formula on regulating lipid metabolism disorders in senile mice based on gut microbiota-TβMCA-FXR axis. Pharmacol Clin Chin Mat Med. 2023;39:18–24.
Li KP, Yu Y, Yuan M, Zhang CM, Rong XL, Turnbull JE, Guo J. Tian-Huang Formula, a traditional chinese medicinal prescription, improves hepatosteatosis and glucose intolerance targeting AKT-SREBP nexus in diet-induced obese rats. Evid Based Complement Alternat Med. 2021;2021:6617586.
Wang J, Wang L, Lou GH, Zeng HR, Hu J, Huang QW, Peng W, Yang XB. Coptidis rhizoma: a comprehensive review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Pharm Biol. 2019;57:193–225.
Wang T, Guo R, Zhou G, Zhou X, Kou Z, Sui F, Li C, Tang L, Wang Z. Traditional uses, botany, phytochemistry, pharmacology and toxicology of Panax notoginseng (Burk.) F.H. Chen: A review. J Ethnopharmacol. 2016;188:234–258.
Xiao S, Liu C, Chen M, Zou J, Zhang Z, Cui X, Jiang S, Shang E, Qian D, Duan J. Scutellariae radix and coptidis rhizoma ameliorate glycolipid metabolism of type 2 diabetic rats by modulating gut microbiota and its metabolites. Appl Microbiol Biotechnol. 2020;104:303–17.
Brownlee M. The pathobiology of diabetic complications: a unifying mechanism. Diabetes. 2005;54:1615–25.
Gilbert ER, Fu Z, Liu D. Development of a nongenetic mouse model of type 2 diabetes. Exp Diabetes Res. 2011;2011: 416254.
Tsai YZ, Tsai ML, Hsu LY, Ho CT, Lai CS. Tetrahydrocurcumin upregulates the adiponectin-adipor pathway and improves insulin signaling and pancreatic β-Cell function in high-fat diet/streptozotocin-induced diabetic obese mice. Nutrients. 2021;13:4488.
Park E, Lee CG, Jeong H, Yeo S, Kim JA, Jeong SY. Antiadipogenic effects of mixtures of and extracts on 3T3-L1 preadipocytes and high-fat diet-induced mice. Molecules. 2020;25:2350.
Taylor SI, Cama A, Accili D, Barbetti F, Imano E, Kadowaki H, Kadowaki T. Genetic basis of endocrine disease. 1. molecular genetics of insulin resistant diabetes mellitus. J Clin Endocrinol Metab. 1991;73:1158–1163.
Warren BE, Lou PH, Lucchinetti E, Zhang L, Clanachan AS, Affolter A, Hersberger M, Zaugg M, Lemieux H. Early mitochondrial dysfunction in glycolytic muscle, but not oxidative muscle, of the fructose-fed insulin-resistant rat. Am J Physiol Endocrinol Metab. 2014;306:E658–67.
Kim JS, Lee H, Jung CH, Lee SJ, Ha TY, Ahn J. Chicoric acid mitigates impaired insulin sensitivity by improving mitochondrial function. Biosci Biotechnol Biochem. 2018;82:1197–206.
Kitada M, Ogura Y, Monno I, Koya D. Sirtuins and type 2 diabetes: role in inflammation, oxidative stress, and mitochondrial function. Front Endocrinol (Lausanne). 2019;10:187.
Boengler K, Kosiol M, Mayr M, Schulz R, Rohrbach S. Mitochondria and ageing: role in heart, skeletal muscle and adipose tissue. J Cachexia Sarcopenia Muscle. 2017;8:349–69.
Cui W, Ma J, Wang X, Yang W, Zhang J, Ji Q. Free fatty acid induces endoplasmic reticulum stress and apoptosis of β-cells by Ca2+/calpain-2 pathways. PLoS ONE. 2013;8: e59921.
Kwong JQ, Lu X, Correll RN, Schwanekamp JA, Vagnozzi RJ, Sargent MA, York AJ, Zhang J, Bers DM, Molkentin JD. The mitochondrial calcium uniporter selectively matches metabolic output to acute contractile stress in the heart. Cell Rep. 2015;12:15–22.
Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414:813–20.
Hu L, Ding M, Tang D, Gao E, Li C, Wang K, Qi B, Qiu J, Zhao H, Chang P, Fu F, Li Y. Targeting mitochondrial dynamics by regulating Mfn2 for therapeutic intervention in diabetic cardiomyopathy. Theranostics. 2019;9:3687–706.
Suarez J, Cividini F, Scott BT, Lehmann K, Diaz-Juarez J, Diemer T, Dai A, Suarez JA, Jain M, Dillmann WH. Restoring mitochondrial calcium uniporter expression in diabetic mouse heart improves mitochondrial calcium handling and cardiac function. J Biol Chem. 2018;293:8182–95.
Glancy B, Balaban RS. Role of mitochondrial Ca2+ in the regulation of cellular energetics. Biochemistry. 2012;51:2959–73.
De-Stefani D, Raffaello A, Teardo E, Szabo I, Rizzuto R. A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature. 2011;476:336–40.
Kain V, Sawant MA, Dasgupta A, Jaiswal G, Vyas A, Padhye S, Sitasawad SL. A novel SOD mimic with a redox-modulating mn (II) complex, ML1 attenuates high glucose-induced abnormalities in intracellular Ca2+ transients and prevents cardiac cell death through restoration of mitochondrial function. Biochem Biophys Rep. 2016;5:296–304.
Acknowledgements
Not applicable.
Funding
This work was supported by National Natural Science Foundation of China (81830113 and 82074210), Basic and Applied Basic Research Foundation of Guangdong Province (2019B030302005) and a research project with the Natural Science Foundation of Guangdong Province (2020A1515010245).
Author information
Authors and Affiliations
Contributions
GJ, DSL and XLR conceived and designed the study. DSL, YRZ, ZYF and YTZ performed the experiments. YRZ, ZYF, YTZ, YH and JYP contributed to the analysis of date. YRZ, ZYF and YTZ wrote the manuscript. GJ, DSL and XLR reviewed and edited themanuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All the animal experiments were approved by the Animal Ethical Committee of Guangdong Pharmaceutical University (SPF2017310). All methods were carried out in accordance with relevant guidelines and regulations of the Animal Ethical Committee of Guangdong Pharmaceutical University. All methods were performed in accordance with ARRIVE guidelines (https://arriveguidelines.org/). All procedures complied with the IUCN Policy Statement on Research Involving Species at Risk of Extinction and the Convention on the Trade in Endangered Species of Wild Fauna and Flora.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table 1. Primers for Real-Time PCR detection.
Additional file 2:
Table 2. Primary antibodies for Western blotting assay.
Additional file 3: Supplementary Figure 5.
Effect of THF on mitochondrial energy metabolism-related protein levels in epididymal white adipose tissue of HFD/STZ-induced T2DM mice.
Additional file 4: Supplementary Figure 6.
Effect of THF on mitochondrial energy metabolism-related protein levels in 3T3-L1 adipocytes cells induced dexamethasone.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Luo, D., Zhao, Y., Fang, Z. et al. Tianhuang formula regulates adipocyte mitochondrial function by AMPK/MICU1 pathway in HFD/STZ-induced T2DM mice. BMC Complement Med Ther 23, 202 (2023). https://doi.org/10.1186/s12906-023-04009-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-023-04009-5