
Abstract
Background
Calamus rotang L. (CR) is an Indian shrub. The leaves and other organs of the plant are traditionally used in India for treatment of various diseases. The in vitro antioxidant property of the leaves extract was previously established. Thus, the current study aimed to evaluate the antioxidant and hepatoprotective effects of CR ethyl acetate extract at a dose of 350 mg/kg on CCl4 induced hepatotoxic rats through different mechanisms.
Methods
Histopathological examination of the treated rats’ group in comparison with positive and negative controls were performed. Quantitative measuring of the proinflammatory cytokines (TNF α), inflammatory regulators (Arginase, PPAR α) and the antiapoptotic protein Bcl-2 in comparison with positive and negative control groups was achieved using immunohistochemical examination. HPLC profiling of the polyphenol contents and molecular docking of the identified compounds against BH3 proapoptotic protein were correspondingly studied to evaluate the potential antiapoptotic property.
Results
The CR extract greatly protects the liver tissue through the suppression of TNF α, arginase and PPAR α induced by CCl4 as well as its enhancement of the antiapoptotic Bcl-2 protein. Fourteen polyphenols of different classes were identified in CR extract and tested via molecular docking for their potential antiapoptotic activities against BH3 protein. Naringin, rutin, 7-hydroxy flavone, and ellagic acid compounds exhibit the highest affinity and potential inhibition of pro-apoptotic protein BH3 via molecular docking study.
Conclusions
The ethyl acetate fraction of the leaves of C. rotang is rich in polyphenols that exhibited potent hepatoprotective effect on CCl4 induced hepatotoxic rats through its antioxidant, anti-inflammatory, anti-steatosis and antiapoptotic properties.
Similar content being viewed by others


Introduction
The liver is a site of metabolism and detoxification of chemicals, drugs, and harmful environmental toxicants. It protects the whole-body organs from injury by those harmful toxins. The exposure to liver toxins in food, water, air and medicines for a long time causes acute liver injury, cirrhosis, fibrosis, fatty liver and cancer. These diseases are worldwide spread nowadays. In some critical cases, urgent liver transplantation is required to save the life of patients [1].
Halogenated alkanes such as carbon tetrachloride (CCl4), chloroform and iodoform are examples of chemical toxins that have been restricted due to their high risk of hepatotoxicity [2, 3]. Liver enzymes activate the halogenated alkanes (CCl4) into highly reactive free radicals such as trichloromethyl and trichloromethyl peroxyl radicals (·CCl3 and ·CCl3O2). These radicals interact with hepatic cell proteins, fats and nucleic acids which harm the normal cellular processes such as lipid metabolism, hepatic enzyme reactions, initiation of cancer, liver fibrosis, and finally cell death [2]. Therefore, antioxidants such as plant polyphenols (flavonoids and tannins) have an important role in scavenging the free radicals (·CCl3 and ·CCl3O2) [4] and protect the hepatic cells from their liver toxicity [5, 6].
Kupffer cells are the main players in liver inflammation. They are localized macrophages in the liver and are responsible for the expression and release of cytokines upon their activation during liver injury. They exhibit markers such as M1-like macrophages or M2-like macrophages that regulate liver inflammation [7]. Several inflammatory cytokines such as chemokines, IL-6, IL-12, IL-18, and TNFα are released from Kupffer cells in response to liver injury, alcoholic liver disease, and other hepatic toxins [7]. Other bone-marrow derived macrophages, such as M1 and M2 macrophages, play a role in liver inflammation by releasing inflammatory cytokines (TNF-α, IL-1, and IL-6) [7]. After liver inflammation, hepatic macrophages such as Kupffer cells and macrophage-TGF-β1 activate myofibroblasts (hepatic stellate cells) to produce fibrosis [7].
CCl4, a liver toxin, induces Kupffer cells to release several inflammatory mediators such as TNFα, TGF-ß, and IL-1, nitric oxide, IL-10 and IL-6 which are responsible for necrosis and fibrosis of hepatic cells [8]. Polyphenols (flavonoids and tannins) are promising anti-inflammatory compounds that inhibit the release of the inflammatory mediators by several mechanisms and hence, protect the hepatic cells from necrosis and fibrosis [9].
Several plant extracts exhibited potent in vivo hepatoprotective activity against CCl4 induced hepatotoxicity. For example, the methanol and chloroform extracts as well as some isolated phenolics from Aframomum melegueta seeds exhibited hepatoprotective activity on CCl4 induced hepatotoxic rats. Suppression of inflammatory response of cytokines, apoptosis and enhancement of the antioxidant defense activity were the mode of action [10]. Lannea stuhlmannii and Lannea humilis tannins rich extracts displayed a potent hepatoprotective effect through the enhancement of the anti-apoptotic protein Bcl-2 [11]. Strawberry (Fragaria ananassa) juice showed enhancement of the anti-apoptotic Bcl-2 protein and controlled the pro-apoptotic Bax and caspase-3 proteins with a clear reducing collagen areas in hepatic tissue [12]. Phenolic extract of barley (Hordeum vulgare) exhibited a hepatoprotective effect on hepatotoxic mice induced by CCl4 [13]. The elevation of liver antioxidant enzymes and reducing the hepatocyte apoptosis were the mechanism of action of the phenolic extract of barley [13].
Calamus rotang L. (Rattan palm) a monocot climbing shrub, belongs to family Ericaceae. It is native to South-West Asia. In India, the shrub is cultivated for its’ edible fruits and medicinal importance especially in Ayurveda. The shoots are used as anthelmintic and the leaves are used for eye problems, skin diseases, pruritus and cough [14,15,16].
Few biological studies on C. rotang (CR) were previously reported. For example, the aqueous extract of leaves showed immunoadjuvant activity against hepatitis B surface antigen [17]. The in vitro antioxidant study on fruits and leaves methanol extracts revealed that the leaves have more polyphenolics than the fruits [18]. The methanol extract of seeds displayed in vivo antidiabetic and hypoglycemic activities [19]. The phytochemical study on C. rotang rhizome led to isolation of ( +) –afzelechin, β-sitosterol and β-sitosterol glucoside [20]. No chemical study was previously reported on the leaves, fruits and seeds. The current study aimed to determine the total phenolics, total flavonoids, antioxidant property and in vivo hepatoprotective activity of the CR extract on CCl4 induced hepatotoxic rats. HPLC chemical characterization of the polyphenols (flavonoids and phenolics) in CR extract was performed. The binding ability of the identified compounds to the pro-apoptotic Bcl-2: Bim (BH3) protein by molecular docking was studied.
Material and methods
Solvents and chemicals
The solvents used in this study for extraction (methanol) and chromatography (methanol and acetonitrile) were HPLC solvents (Thermo Fisher Scientific, Pittsburgh, USA). The chemicals, such as Folin-Ciocalteu’s phenol reagent, gallic acid, quercetin, AlCl3, DPPH, and ascorbic acid, used in the determination of phenolic and flavonoid contents and antioxidant activity were purchased from Sigma (Sigma-Aldrich, Burlington, MA, USA). Hematoxylin and eosin kits and toluidine blue dye used in the histopathological study were bought from Abcam (Cambridge, UK). The following antibodies: anti-TNFα antibody (sc-52746), anti-PPARα antibody (sc-398394), anti-arginase (sc-271430), and anti-Bcl-2 antibody (sc-7382) used in the immunohistochemistry study were purchased from Santa Cruz Biotechnology (Texas, USA).
Plant materials
The leaves of C. rotang L. were used in the current study. They were collected in October 2018 from Aswan Botanical Garden, Aswan, Egypt. The plant was kindly identified by Dr. Amr M. M. Mahmoud, Director of Aswan Botanical Garden, Hort. Res. Institute, Agriculture centre, Egypt. Voucher specimen of the plant leaves was placed in the herbarium of the Pharmacognosy Department, Faculty of Pharmacy, Assiut University, Egypt (Voucher no. A20220906).
Extraction and fractionation
A hundred grams of dried powdered leaves were macerated in methanol (500 mL) at room temperature (25 °C) with shaking for 24 h. The extraction process was repeated twice until exhaustion. The methanol extract was combined and concentrated under reduced pressure using rotavapor. The fractionation of the dried methanol extract (10 g) was performed by suspending it in 250 mL of 10% methanol in water using a 1L separating funnel. Liquid–liquid extraction to fractionate the aqueous extract using dichloromethane and ethyl acetate solvents (250 mL × 3 for each solvent) was performed. The dichloromethane and ethyl acetate fractions were collected and concentrated at reduced pressure to yield dichloromethane extract (3 g), ethyl acetate extract (2 g), and aqueous extract (5 g). The preliminary screening test for flavonoids demonstrated that the ethyl acetate fraction is the richest flavonoid fraction.
Total phenolic contents
The total phenolic content of CR extract was determined according to the method previously reported [21]. Briefly, gallic acid was used as a standard phenolic compound. Serial dilution of 100 ppm gallic acid solution was prepared (100–10 ppm) in distilled water. Folin-Ciocalteu’s phenol reagent (0.2 mL) was mixed with all gallic acid solutions and tested CR ethyl acetate extract (1:1). One mL of sodium carbonate and 1.6 mL distilled water were added to the mixtures after 5 min. All tested and standard mixtures were kept on dark for 30 min. After that, the samples were centrifuged, and their colorimetric absorbance was measured in triplicates at 750 nm using UV2000 spectrophotometer (Ray Wild Limited company, L569 Gottingen, Germany). Standard curve of gallic acid was obtained (Fig. S3) and the total phenolic content of CR extract GAE mg/g dry weight was calculated as a mean of triplicate measurements ± SD.
Total flavonoid contents
The total flavonoid content of CR extract was determined according to the method previously reported [22]. Serial dilution of 100 ppm quercetin solution was prepared (100–10 ppm) in methanol. AlCl3 2% reagent (0.6 mL) was mixed with all quercetin solutions and tested CR ethyl acetate extract (1:1). All tested and standard mixtures were kept for 60 min at room temperature. After that, the samples were measured for their colorimetric absorbance in triplicates at 415 nm using UV2000 spectrophotometer (Ray Wild Limited company, L569 Gottingen, Germany) Standard curve of quercetin was obtained (Fig. S4) and the total flavonoid content of CR extract QE mg/g dry weight was calculated as a mean of triplicate measurements ± SD.
Antioxidant DPPH assay
The capacity of CR extract to scavenge free radicals was calculated by DPPH previously reported method [23]. DPPH stock solution of 200 µM, tested extract samples (60—1000 µg/mL ethanol) and ascorbic acid reference samples (60—1000 µg/mL ethanol) were prepared. The antioxidant reaction was performed by adding 300 µL DPPH solution to 300 µL of each tested or control sample on dark for 30 min at room temperature. The colorimetric absorbance was measured at 517 nm. The percentage antioxidant activity was calculated by the following equation SA% = (A0—As/A0) × 100 (A0 = Absorbance of DPPH solution in ethanol, As = Absorbance of CR extract and DPPH).
HPLC analysis of CR ethyl acetate extract
Detection of phenolic compounds
Twenty-five microliter of CR was injected in HPLC-Agilent 1100 with UV/Vis detector (Agilent technology, USA). The phenolic compounds were separated on C18 column (125 mm × 4.60 mm, 5 µm) using gradient solvent elution. Two solvents were used as mobile phase for separation of phenolics, solvent A: acetic acid in water (1:25) and solvent B: 100% methanol. The gradient solvent method was started with 100% of solvent A for 3 min, followed by 50% solvent B for 5 min, 80% solvent B for next 2 min and finally 50% solvent B for 5 min. Phenolic compounds were detected at 280 nm. They were detected using Agilent ChemStation software (Agilent technology, USA) and were identified by using authentic samples.
Detection of flavonoids compounds
Twenty-five microliter of CR was injected in HPLC-Agilent 1100 with UV/Vis detector (Agilent technology, USA). The phenolic compounds were separated on C18 column (250 mm × 4.60 mm, 5 µm) using isocratic solvent elution in 15 min. Two solvents were used as mobile phase for separation of flavonoids, solvent A: formic acid in water (1%) and solvent B: 100% acetonitrile. Seventy % of solvent B was used as isocratic eluted mobile solvent of the experiment. The detector wavelength was 320 nm. The identification of flavonoids was performed by using authentic samples.
Animals and experimental design
Eighteen Wistar albino rats weighting 180 to 250 g (aged 6 weeks) were obtained from Animal Housing Center, Faculty of Medicine, Assiut University. They were housed, provided feed and water ad libitum under standard conditions for 7 days before experiments begun. This experiment was carried out in accordance with relevant guidelines and regulations. It was approved by the Institutional Animal Care and Use Committee of the Faculty of Pharmacy, Assiut University (Approved No. S27-22). The rats were divided into three groups; control, CCl4 intoxicated and CR extract treated groups. CCl4 was chosen to intoxicate the hepatic cells in the positive and treated groups based on a previously published protocol of CCl4 induced hepatotoxicity in rats [24,25,26,27]. The rats of treated (CR extract) group were supplied orally by 0.3 mL of 350 mg/kg CR extract once a daily for 21 days [28]. The dose of ethyl acetate extract was determined in the range of the previous reported doses of hepatoprotective medicinal plants rich in flavonoids and phenolics [29, 30]. The rats of other groups were supplied orally by 0.3 mL of saline once a daily for 21 days. The rats of all groups except the control were supplied orally with 0.25 mL of 1% CCl4 in olive oil per day from day 10. The rats were fasted for 4 h and the animals killed at the end of the corresponding experimental periods. The animals were anesthetized by pentobarbital (50 mg/kg, i.p.). After the rat loss the reflexes, the rat’s thoracic cages had been incised and transcardially perfused with normal saline followed by 4% paraformaldehyde fixative. Liver specimens were obtained after whole-body perfusion of experimental rats with 4% paraformaldehyde. All methods are reported in accordance with ARRIVE guidelines (https://arriveguidelines.org).
Histopathological examination
The dissected samples were processed and stained with hematoxylin & eosin and Crossmon’s trichrome (Fig. S5) according to the standard protocols [31]. Histopathological studies were performed using an Olympus CX 41RF light microscope (Olympus Corporation, Tokyo, Japan). Also, specimens from liver were used for semithin sections and stained with toluidine blue [32].
Immunohistochemical examination
Immunohistochemistry staining was performed according to the previous reported strudy [33]. The sections were incubated overnight at 4 °C with the following antibodies: anti-TNFα antibody (sc-52746), anti-PPARα antibody (sc-398394), anti-arginase (sc-271430) and anti-Bcl-2 antibody (sc-7382). Immunohistochemical staining was evaluated by Olympus CX 41RF light microscope (Olympus Corporation, Tokyo, Japan).
Quantitative analysis of TNFα, Arginase I, BCL2, PPARα immunostaining
The morphometric studies carried out on the immunohistochemical images of the liver of all groups using Image-J software. The measurements were done according to the previous study [34].
Statistical analysis
The data of immunohistochemical analysis were summarized in figures using “GraphPad Software” (Version 6.05, International Scientific Community) to compare between different variables in CCl4, CR extract treated animals in relation to control group. Differences were considered significant if P < 0.05 (*). All data were statistically analyzed using Tukey’s test.
Molecular docking
Molecular docking was conducted by using Autodock [35] vina 1.5.6 software to assess the binding affinities of the selected polyphenols with antiapoptotic protein Bcl-2: (BH3). The crystal structure of BH3 with its inhibitor with PDB ID: 4B4S was retrieved from Protein Data Bank [36]. The interaction residues of the binding site of BH3 that was used for the docking study were identified based on the previous literature [11, 36]. The ligand and all the water molecules were removed from the crystal structure. The polar hydrogen atoms were added and pdbqt file was generated by using AutoDock tools-1.5.6 [37]. The site of the grid box was set at -8.425, 9.62, and -2.654 Å (for × , y and z) by means of a grid of 40, 40, and 40 points (for × , y and z). The binding affinities of polyphenols with Bcl-2 were predicted based on the average of the lowest energy of docking. Chimera [38] 1.12 software was used to analyze and visualize the best-scored conformation.
Results
Determination of total phenolics, total flavonoids and antioxidant properties of C. rotang L. ethyl acetate
The total phenolics content of C. rotang ethyl acetate extract was determined by using gallic acid as a reference compound in Folin-Ciocalteu assay [21]. It contains 804 ± 0.18 mg Gallic Acid Equivalent (GAE)/100 g dry weight as shown in Fig. S1A. The quantity of flavonoids in C. rotang ethyl acetate fraction (Fig. S1A) was determined as 1760 ± 0.69 mg Quercetin Equivalent (QE)/100 g dry weight using quercetin as a reference compound in Aluminum chloride colorimetric assay [22]. The CR extract displayed potent DPPH scavenging activity with IC50 = 0.12 mg/mL using ascorbic acid as a reference compound in the DPPH assay (Fig. S1B.).
HPLC analysis of C. rotang L. leaves ethyl acetate extract
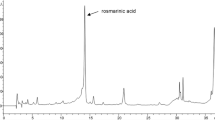
CR leaves extract was analyzed by using HPLC–UV for detection of flavonoids and phenolics. Authentic compounds were used to identify the components of CR extract. Fourteen phenolics and flavonoids were detected as shown in HPLC chromatograms (Fig. S2). All the identified compounds are listed in Table 1 and Fig. 1.

Chemical structures of phenolics and flavonoids detected by HPLC analysis of C. rotang L. ethyl acetate extract
Histopathological findings
The liver in the control group exhibited the normal appearance of hepatic lobules with hexagonal hepatocytes, sinusoidal spaces, and central vein (Fig. 2A & D). However, in the CCl4 treated group, the normal hepatic architecture was distorted; hepatocytes showed vacuolated cytoplasm, the central and portal veins were dilated and congested with blood (Fig. 2B). Also, there was a mononuclear cellular infiltration and steatosis (Fig. 2B & E). CR ethyl acetate extract group exhibited potent protection of the hepatic tissue from the histopathological changes caused by CCl4 (Fig. 2C & F). There was no or minor vacuolation of the hepatocytes, no sinusoidal dilation or the central and portal veins with slight dilation and congestion were recorded (Fig. 2C & F).

Photomicrograph of a liver sections stained with H & E (A-C) and semithin sections stained by Toluidine blue (D-F). A, D showing the normal architecture of hexagonal liver lobules with apparently intact hepatocyte, central vein (CV), and blood sinusoids (S). B, E Liver section of rat treated with CCl4, showing distortion of the normal hepatocyte’s appearance with vacuolated cytoplasm, congested portal (PV) and central veins, as well as dilated blood sinusoids, mononuclear cellular infiltration (arrow) and steatosis (arrow heads). C, F Liver section of rat treated with CR extract group, showing the hepatocytes appearance improved with lesser and smaller vacuoles
Immunohistochemical findings
The Kupffer cells (resident macrophage) and T-cells in the liver produces proinflammatory cytokines like TNF α [39]. TNF α strongly expressed in hepatocytes, endothelium sinusoids and Kupffer cells in CCl4 treated group in relative to the normal and CR treated groups (Fig. 3A-D). Arginase is a manganese-containing enzyme, it catalyzes the last stage in the urea cycle, that is responsible for conversion of arginine to urea [40]. Arginase strongly immunoreacted in hepatocytes of the CCl4 treated group in relative to the normal and CR treated groups (Fig. 3E-H). Additionally, Bcl-2 is an anti-apoptotic marker, showed a significant increase in hepatocytes immunostaining of the normal and CR treated groups in comparison with CCl4 treated group (Fig. 3I-L). The peroxisome proliferator-activated receptors alpha (PPARs α) are transcription factor responsible for the catabolism of the fatty acids in liver [41]. In the CCl4 treated group the immunostaining of PPAR α was significantly higher in relation to the normal and CR treated groups (Fig. 3M-P).

Immunohistochemical staining: A Normal hepatocytes showing a weak immunoreactivity against TNF α. B CCl4 treated groups, showing strongest immunoreactivity in Kupffer cells and endothelium of the central vein. C CR treated group, is showing lesser immunoreactivity. D Staining quantification of TNF α using image j software. E Normal hepatocyte is showing a weak immunoreactivity against Arginase I. F CCl4 treated group, the cytoplasm of the hepatocytes (red squares) exhibiting strong reactivity within the hepatocyte’s cytoplasm and Kupffer cells (short arrows). G: The hepatocytes show lesser immunoreactivity in CR ethyl acetate extract. H Staining quantification of Arginase I, using image j software. I Increasing in the number of the Bcl-2 positive hepatocytes (arrows). J CCl4 treated group, is showing decrease in Bcl-2 positive cells. K: CR ethyl acetate extract treated groups. L Staining quantification of Bcl-2 using image j software. M Normal group, the cytoplasm of the periportal hepatocytes is showing a staining against PPAR α (arrows). N CCl4 treated group, is showing an increase in PPAR α positive cells. O CR ethyl acetate extract treated group. P Staining quantification of PPAR α using image j software. The CR extract treated groups is compared to control and CCl4 groups using Tukey test (n = 3). *P ≤ 0.05
Molecular modelling study
Naringin, rutin, 7-hydroxy flavone, and ellagic acid exhibited the highest binding affinities with Bcl-2: Bim (BH3) among all the studied polyphenols with docking energy of binding -6.88 ± 0.06, -6.70 ± 0.19, -6.42 ± 0.31 and -6.57 ± 0.26 kcal/mol respectively. All the binding affinities of the polyphenols are listed in Table S1. Naringin formed two hydrogen bonds with the side chains of SER40 and TYR73 at the binding site of Bcl-2: Bim (BH3) (Fig. 4A). Rutin formed a unique hydrogen bonds pattern with the binding site of Bcl-2: Bim (BH3), it formed three hydrogen bonds with SER40 and one more with TYR73. Also, its aromatic ring formed aryl–aryl interactions with the aromatic ring of PHE163 (Fig. 4B). Hydroxy flavone formed two hydrogen bonds with the side chains of SER40 and TYR73 (Fig. 4C). Ellagic acid formed two hydrogen bonds with SER40 and one more hydrogen bond with TYR73 (Fig. 4D).

3D-docking poses of Naringin (1A), Rutin (1B), Hydroxy flavone (1C), and Ellagic acid (1D) docked to Bcl-2: Bim (BH3). The binding affinities of Naringin, rutin, 7-hydroxy flavone, and ellagic acid with Bcl-2: Bim (BH3) are -6.88 ± 0.06, -6.70 ± 0.19, -6.42 ± 0.31 and -6.57 ± 0.26 kcal/mol respectively
Discussion
The CR leaf extract displayed potent DPPH scavenging activity, with IC50 = 0.12 mg/mL. The DPPH scavenging of the CR increased with concentration. The higher the concentration of the CR extract, the greater the scavenging of DPPH radicals (Fig. S1).
The most natural plant components that are responsible for antioxidants are flavonoids and phenolics [42, 43]. The first quantitative determination of flavonoids and phenolics in the CR leaf extract was reported in the current study. The total phenolic content of the CR extract was 804 ± 0.18 mg GAE/100 g, and the total flavonoid content was 1760 ± 0.69 mg QE/100 g (Fig. S1). Therefore, CR is a rich source of flavonoids and phenolics in comparison with the previously reported data on antioxidant plants [44,45,46]. The HPLC characterization of phenolics in CR extract led to the identification of 6 simple phenolic acids; gallic acid, ellagic acid, syringic acid, p-coumaric acid, caffeic acid and ferulic acid in addition to one phenolic compound (pyrogallol) as shown in Fig. 1. Additionally, seven flavonoid compounds of different classes; flavonols (Rutin, quercetin, and kaempferol), flavones (Apigenin, 7-OH flavone and myricetin) and a flavanone (Naringin) were identified (Fig. 1). The detection of compounds was achieved by using Agilent ChemStation software (Agilent technology, USA). Authentic compounds were used to identify and quantify flavonoids and phenolics in CR extract. This study is the first to report the polyphenol composition of CR leaf extract.
The histopathological and immunohistochemical studies proved that CR extract greatly protects the liver tissue from the steato-cirrhotic effect of CCl4. The exposure of the rats to CCl4 causes hepatocytes vacuolation, and induces hepatic inflammation and fibrosis as previously reported [47]. The CR extract significantly decreased the collagen deposition, and suppressed the inflammatory cellular infiltrations. Tumor necrosis factor α (TNF α) is a one of cytokines produced mainly by macrophages, that stimulates cellular response such as production of the inflammatory mediators and cell death [48]. In current study, the CCl4 treated group showed a significant increase in TNF α in comparison to CR treated group. This was attributed to the role of CR extract as an anti-inflammatory agent. Arginine is a substrate for the synthesis of urea and nitric oxide, arginase has a catalytic activity which converts arginine to urea [40]. In the present work, the arginase showed a significant decrease in CR treated group in comparison to CCl4, and its elevation in the CCl4, intoxicated group occurred as a result of the up regulation of TNF α [49].
The accumulation of reactive oxygen species (ROS) within the cells along with a decrease in the cellular antioxidants provokes mitochondrial damage [50]. This led to the release of the apoptogenic factors (Cytochrome c) through the mitochondrial membrane. Bcl-2 is an anti-apoptotic member which inhibits or decreases apoptosis [51]. The CR extract exhibited a significantly increase in Bcl-2 positive hepatocytes which indicates its role in the protection of the hepatocytes from the programed cell death. Additionally, our results showed a significant increase in PPAR α in the CCl4 intoxicated group due to the elevation of intercellular lipid accumulation. PPAR α mainly controls the lipid metabolism and regulates the inflammatory response in liver [41]. In this regard, the increase in the number of PPAR α positive cells is due to the excess of the inflammatory cell’s infiltration after the exposure to CCl4 [52]. PPARα levels decreased in the CR extract group [53]. Thus, CR ethyl acetate extract has a prospective anti-steatosis activity.
Molecular docking is a powerful approach that has been employed in drug discovery studies to screen the binding affinities of the drugs with its possible target proteins [54]. Therefore, we used this approach to study the inhibitory role of the identified polyphenols on Bcl2: Bim (BH3) as a crucial pro-apoptotic protein. Interestingly, naringin, rutin, hydroxy flavone, and ellagic acid showed a promising binding affinity with Bcl2: Bim (BH3). These high binding affinities are attributed to the unique structure of polyphenols that combine the aromatic rings and the substituted polar groups that allow them to form various non-covalent interactions with the target proteins. These findings suggested the antiapoptotic role of polyphenols through the inhibition of BH3 protein.
Conclusion
The ethyl acetate extract of C. rotang L. displayed significant in vivo hepatoprotective effect on CCl4 induced hepatotoxic rats. The current study confirms its potent antioxidant, anti-inflammatory and anti-steatosis activities through the suppression of TNF α, arginase and PPAR α induced by CCl4. The antiapoptotic property of the extract was through the enhancement of Bcl-2 antiapoptotic protein. DPPH antioxidant activity, total phenolic contents, total flavonoid contents and HPLC analysis revealed that C. rotang L. ethyl acetate extract is a rich source of antioxidant and anti-inflammatory metabolites. Fourteen phenolic and flavonoid compounds were detected. Their docking on proapoptotic protein Bcl-2 (BH3) led to the detection of naringin, rutin, 7-hydroxy flavone and ellagic acid as the highest affinity compounds to BH3.
Availability of data and materials
Correspondence and requests for materials should be addressed to A.M.Z. and F.M.A.
Change history
20 February 2023
A Correction to this paper has been published: https://doi.org/10.1186/s12906-023-03881-5
Abbreviations
- HPLC:
-
High Performance Liquid Chromatography
- Bcl-2:
-
Beta cell lymphoma 2
- TNFα:
-
Tumer Necrosis Factor Alpha
- DPPH:
-
2,2-Diphenyl-1-picrylhydrazyl
- PPARα:
-
Peroxisome Proliferator-Activated Receptor Alpha
- °C:
-
Celsius temperature degree
- mg/kg:
-
Milligram/kilogram
- nm:
-
nanometer
- UV/Vis:
-
Ultraviolet/Visible
- GAE:
-
Gallic acid Equivalent
- QE:
-
Quercetin Equivalent
- SD:
-
Standard Deviation
- CR:
-
Calamus rotang
- µM:
-
Micromolar
- µl:
-
Microliter
- ppm:
-
Part per million
- g:
-
Gram
- CCl4 :
-
Carbon Tetra Chloride
- Bcl-2:BH3:
-
Beta cell lymphoma 2 Homology 3
- PDB ID: 4B4S:
-
Crystal structure of a pro-survival Bcl-2:Bim BH3 complex
References
Bischoff K, Mukai M, Ramaiah SK. Liver toxicity. In: Veterinary toxicology. edn. London: Elsevier. 2018. p. 239–57.
Weber LW, Boll M, Stampfl A. Hepatotoxicity and mechanism of action of haloalkanes: carbon tetrachloride as a toxicological model. Crit Rev Toxicol. 2003;33(2):105–36.
Rahmouni F, Badraoui R, Ben-Nasr H, Bardakci F, Elkahoui S, Siddiqui AJ, Saeed M, Snoussi M, Saoudi M, Rebai T. Pharmacokinetics and Therapeutic Potential of Teucrium polium against Liver Damage Associated Hepatotoxicity and Oxidative Injury in Rats: Computational, Biochemical and Histological Studies. Life. 2022;12(7):1092.
Rahmouni F, Saoudi M, Amri N, El-Feki A, Rebai T, Badraoui R. Protective effect of Teucrium polium on carbon tetrachloride induced genotoxicity and oxidative stress in rats. Arch Physiol Biochem. 2018;124(1):1–9.
Wei E, Zhang S, Zhai J, Wu S, Wang G. The evaluation of hepatoprotective effects of flavonoids from Scorzonera austriaca Wild against CCl4-induced acute liver injury in vitro and in vivo. Drug Chem Toxicol. 2022;45(3):1284–94.
Ghosh A, Sarmah P, Patel H, Mukerjee N, Mishra R, Alkahtani S, Varma RS, Baishya D. Nonlinear molecular dynamics of quercetin in Gynocardia odorata and Diospyros malabarica fruits: Its mechanistic role in hepatoprotection. PLoS One. 2022;17(3):e0263917.
Koyama Y, Brenner DA. Liver inflammation and fibrosis. J Clin Investig. 2017;127(1):55–64.
Popović D, Kocić G, Katić V, Zarubica A, Janković Veličković L, Ničković VP, Jović A, Veljković A, Petrović V, Rakić V. Anthocyanins protect hepatocytes against CCl4-induced acute liver injury in rats by inhibiting pro-inflammatory mediators, polyamine catabolism, lipocalin-2, and excessive proliferation of Kupffer cells. Antioxidants. 2019;8(10):451.
Maleki SJ, Crespo JF, Cabanillas B. Anti-inflammatory effects of flavonoids. Food Chem. 2019;299:125124.
El-Halawany AM, Dine RSE, El Sayed NS, Hattori MJ. Protective effect of Aframomum melegueta phenolics against CCl4-induced rat hepatocytes damage; role of apoptosis and pro-inflammatory cytokines inhibition. Science Rep. 2014;4(1):1–9.
Sobeh M, Mahmoud MF, Hasan RA, Abdelfattah MA, Sabry OM, Ghareeb MA, El-Shazly AM, Wink MJ. Tannin-rich extracts from Lannea stuhlmannii and Lannea humilis (Anacardiaceae) exhibit hepatoprotective activities in vivo via enhancement of the anti-apoptotic protein Bcl-2. Science Rep. 2018;8(1):1–16.
Hamed SS, Al-Yhya NA, El-Khadragy MF, Al-Olayan EM, Alajmi RA, Hassan ZK, Hassan SB, AbdelMoneim AE. The protective properties of the strawberry (Fragaria ananassa) against carbon tetrachloride-induced hepatotoxicity in rats mediated by anti-apoptotic and upregulation of antioxidant genes expression effects. Front Physiol. 2016;7:325.
Quan M, Li Q, Zhao P, Tian CJ. Chemical composition and hepatoprotective effect of free phenolic extract from barley during malting process. Sci Rep. 2018;8(1):1–9.
Basumatary S, Ahmed M, Deka S. Some medicinal plant leaves used by Boro (tribal) people of Goalpara district, Assam. 2004.
Khare CP. Indian medicinal plants: an illustrated dictionary: Springer Science & Business Media. 2008.
Rethy P, Singh B, Kagyung R, Gajurel P. Ethnobotanical studies of Dehang-Debang Biosphere Reserve of Arunachal Prad esh with special reference to Memba tribe. 2010.
Gupta A, Shaikh AC, Chaphalkar SR. Aqueous extract of Calamus rotang as a novel immunoadjuvant enhances both humoral and cell mediated immune response. J Herbmed Pharmacol. 2016;6(1):43–8.
Shil A, Himel MK, Khair A, Alam MN, Uddin A. In vitro antioxidant activity of the methanolic extracts of leaf and fruit of Calamus rotang Linn. J Exp Biosci. 2012;3(2):33–6.
Ripa F, Dash P, Podder A. Anti-diarrheal and hypoglycemic activities of methanol extract of Calamus rotang L. seed in rat. Res J Pharmacognosy. 2016;3(2):33–40.
Saraswathy A, Vidhya B. Afzelechin from the rhizome of Calamus rotang Linn. Indian Drugs. 2012;49:49–50.
Zhang A, Zeng L, Bo H, Hardie WJ. Sulphite-corrected, non-phenolic and phenolic antioxidant capacities of fruit wines profiled by differential Folin-Ciocalteu assay. Int J Food Sci Technol. 2022;57(2):1259–72.
Chen X, Wang H, Huang X, Xia S, Chen C, Nie Q, Nie S. Efficient enrichment of total flavonoids from kale (Brassica oleracea L. var. acephala L.) extracts by NKA-9 resin and antioxidant activities of flavonoids extract in vitro. Food Chemistry. 2022;374:131508.
Isildak Ö, Yildiz I, Genc N. A new potentiometric PVC membrane sensor for the determination of DPPH radical scavenging activity of plant extracts. Food Chem. 2022;373:131420.
Barakat H, Alkabeer IA, Aljutaily T, Almujaydil MS, Algheshairy RM, Alhomaid RM, et al. Phenolics and volatile compounds of fennel (Foeniculum vulgare) seeds and their sprouts prevent oxidative DNA damage and ameliorates CCl(4)-induced hepatotoxicity and oxidative stress in rats. Antioxidants (Basel). 2022;11(12):2318-1–22.
Mazani M, Rezagholizadeh L, Shamsi S, Mahdavifard S, Ojarudi M, Salimnejad R, Salimi A. Protection of CCl(4)-induced hepatic and renal damage by linalool. Drug Chem Toxicol. 2022;45(3):963–71.
Shakya AK, Sharma N, Bhadauria M, Nirala SK, Shrivastava S, Shukla S. Ameliorative impact of herbal formulation -Majoon-Dabeed-ul-ward and Sharbat-e-Deenar against CCl(4) induced liver toxicity via regulation of antioxidant enzymes and oxidative stress. Toxicol Res (Camb). 2022;11(5):819–30.
Sarhadi Kholari F, Dehpour AR, Nourbakhsh M, Doustimotlagh AH, Bagherieh M, Golestani A. Erythrocytes Membrane Alterations Reflecting Liver Damage in CCI4-Induced Cirrhotic Rats: The Ameliorative Effect of Naltrexone. Acta Med Iran. 2016;54(10):631–9.
Quan M, Li Q, Zhao P, Tian C. Chemical composition and hepatoprotective effect of free phenolic extract from barley during malting process. Sci Rep. 2018;8(1):4460.
Ripa FA, Dash PR, Podder AK. Anti-diarrheal and hypoglycemic activities of methanol extract of Calamus rotang L. seed in rat. Res J Pharmacognosy. 2016;3(2):33–40.
Arman M, Chowdhury KAA, Bari M, Khan MF, Huq MM, Haque M, et al. Hepatoprotective potential of selected medicinally important herbs: evidence from ethnomedicinal, toxicological and pharmacological evaluations. Phytochemistry Rev. 2022;21:1–24.
Bancroft JD, Stevens Ai. Theory and practice of histological techniques. 2nd ed: Churchill Livingston; 1982. p. 338439.
Suvarna KS, Layton C, Bancroft JD. Bancroft’s theory and practice of histological techniques E-Book. London: Elsevier Health Sciences; 2018.
Hussein MT, Abdel-Maksoud FM. Structural Investigation of Epididymal Microvasculature and Its Relation to Telocytes and Immune Cells in Camel. Microsc Microanal. 2020;26(5):1024–34.
Mustafa FEA, Abdel-Maksoud FM, Hassan AHS, Mokhtar DM. Melatonin induces a stimulatory action on the scrotal skin components of Soay ram in the non-breeding season. Sci Rep. 2020;10(1):10154.
Xue Q, Liu X, Russell P, Li J, Pan W, Fu J, Zhang AJE, Safety E. Evaluation of the binding performance of flavonoids to estrogen receptor alpha by Autodock. Autodock Vina and Surflex-Dock. 2022;233:113323.
Rautureau G, Yabal M, Yang H, Huang D, Kvansakul M, Hinds MJ. The restricted binding repertoire of Bcl-B leaves Bim as the universal BH3-only prosurvival Bcl-2 protein antagonist. Cell Death Dis. 2012;3(12):e443–e443.
Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(2):455–61.
Butt SS, Badshah Y, Shabbir M, Rafiq M. Molecular docking using chimera and autodock vina software for nonbioinformaticians. JMIR Bioinform Biotechnol. 2020;1(1):e14232.
Hoffmann R, Grewe M, Estler H-C, Schulze-Specking A, Decker K. Regulation of tumor necrosis factor-α-mRNA synthesis and distribution of tumor necrosis factor-α-mRNA synthesizing cells in rat liver during experimental endotoxemia. J Hepatol. 1994;20(1):122–8.
Wu G, Morris SM Jr. Arginine metabolism: nitric oxide and beyond. Biochem J. 1998;336(Pt 1):1–17.
Orfila C, Lepert JC, Alric L, Carrera G, Béraud M, Pipy B. Immunohistochemical distribution of activated nuclear factor kappaB and peroxisome proliferator-activated receptors in carbon tetrachloride-induced chronic liver injury in rats. Histochem Cell Biol. 2005;123(6):585–93.
Barhé TA, Tchouya GRF. Comparative study of the anti-oxidant activity of the total polyphenols extracted from Hibiscus Sabdariffa L., Glycine max L. Merr., yellow tea and red wine through reaction with DPPH free radicals. Arab J Chem. 2016;9(1):1–8.
Mishra K, Ojha H, Chaudhury NK. Estimation of antiradical properties of antioxidants using DPPH assay: A critical review and results. Food Chem. 2012;130(4):1036–43.
Lin J-Y, Tang C-Y. Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem. 2007;101(1):140–7.
Bajalan I, Mohammadi M, Alaei M, Pirbalouti AG. Total phenolic and flavonoid contents and antioxidant activity of extracts from different populations of lavandin. Ind Crops Prod. 2016;87:255–60.
Aryal S, Baniya MK, Danekhu K, Kunwar P, Gurung R, Koirala N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants. 2019;8(4):96.
Fu Y, Zheng S, Lin J, Ryerse J, Chen A. Curcumin protects the rat liver from CCl4-caused injury and fibrogenesis by attenuating oxidative stress and suppressing inflammation. Mol Pharmacol. 2008;73(2):399–409.
Wajant H, Pfizenmaier K, Scheurich P. Tumor necrosis factor signaling. Cell Death Differ. 2003;10(1):45–65.
Hesse M, Modolell M, La Flamme AC, Schito M, Fuentes JM, Cheever AW, Pearce EJ, Wynn TA. Differential regulation of nitric oxide synthase-2 and arginase-1 by type 1/type 2 cytokines in vivo: granulomatous pathology is shaped by the pattern of L-arginine metabolism. J Immunol. 2001;167(11):6533–44.
Sobeh M, Mahmoud MF, Hasan RA, Abdelfattah MAO, Sabry OM, Ghareeb MA, El-Shazly AM, Wink M. Tannin-rich extracts from Lannea stuhlmannii and Lannea humilis (Anacardiaceae) exhibit hepatoprotective activities in vivo via enhancement of the anti-apoptotic protein Bcl-2. Sci Rep. 2018;8(1):9343.
Billard C. Design of novel BH3 mimetics for the treatment of chronic lymphocytic leukemia. Leukemia. 2012;26(9):2032–8.
Rahmouni F, Badraoui R, Amri N, Elleuch A, El-Feki A, Rebai T, Saoudi M. Hepatotoxicity and nephrotoxicity in rats induced by carbon tetrachloride and the protective effects of Teucrium polium and vitamin C. Toxicol Mech Methods. 2019;29(5):313–21.
Moller D, Berger J. Role of PPARs in the regulation of obesity-related insulin sensitivity and inflammation. Int J Obes Relat Metab Disord. 2003;27(3):S17–21.
Meng X-Y, Zhang H-X, Mezei M, Cui M. Molecular docking: a powerful approach for structure-based drug discovery. Curr Comput Aided Drug Des. 2011;7(2):146–57.
Acknowledgements
The authors prompt their sincere thanks to Dr. Raof Sultan, Department of Botany and Microbiology, Faculty of Science, Assiut University, Assiut, Egypt for his contribution in the UV spectroscopic measurements of the extract.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
W.S.A:. Extraction and fractionation of the plant and participated in the data analysis. F.M.A: Performing the histopathological and immunohistochemical studies. A.M.S.: Performing the molecular docking study.I.A.M.A, M.A.M: participate in data analysis. A.M.Z. Responsible for the aim of the study, selection of the plant, interpretation of the data and writing the manuscript. F.M.A., A.M.S., I.A.M.A, M.A.M and A.M.Z. reviewed the manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Authors animal experiment was approved by the Institutional Animal Care and Use Committee of the Faculty of Pharmacy, Assiut University (Approved No. S27-22). All methods are reported in accordance with ARRIVE guidelines (https://arriveguidelines.org). This experiment was carried out in accordance with relevant guidelines and regulations.
Plant ethics—The plant was kindly identified by Dr. Amr M. M. Mahmoud, Director of Aswan Botanical Garden, Hort. Res. Institute, Agriculture Centre, Egypt. Voucher specimen of the plant leaves was placed in the herbarium of the Pharmacognosy Department, Faculty of Pharmacy, Assiut University, Egypt (Voucher no. A20220906). This experiment was carried out in accordance with relevant guidelines and regulations. Permission to collect plant material was obtained from Aswan Botanical Garden, Hort. Res. Institute, Agriculture Centre, Aswan, Egypt.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: it was noted that due to a typesetting error Dr. Walaa S. Anwar and Dr. Makboul A. Makboul were incorrectly affiliated to Merit University. The correct affiliation for these authors should be the Assiut University.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Anwar, W.S., Abdel-maksoud, F.M., Sayed, A.M. et al. Potent hepatoprotective activity of common rattan (Calamus rotang L.) leaf extract and its molecular mechanism. BMC Complement Med Ther 23, 24 (2023). https://doi.org/10.1186/s12906-023-03853-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-023-03853-9