
Abstract
Background
Chlormadinone acetate (CMA) is a derivative of progesterone and is used as an oral contraceptive. The aim of this study was to investigate the effects of CMA on odontogenic differentiation and mineralization of human dental pulp cells (hDPCs) and related signaling pathways.
Methods
Cell viability was determined by the water-soluble tetrazolium (WST)-1 assay. Odontogenic differentiation of hDPCs was evaluated by real-time polymerase chain reaction using odontogenic marker genes, such as alkaline phosphatase (ALP), osteocalcin (OCN), dentin sialophosphoprotein (DSPP), and dentin matrix protein-1 (DMP-1). Mineralization of hDPCs was evaluated by ALP staining and alizarin red staining. The extracellular signal-regulated kinase (ERK) pathway was examined by Western blot analysis.
Results
There was no statistically significant difference in cell viability between the control and CMA-treated groups. Our analysis of odontogenic marker genes indicated that CMA enhanced the expression of those genes. CMA-treated hDPCs showed increased ALP activity and formation of mineralized nodules, compared with control-treated cells. In addition, CMA stimulation resulted in phosphorylation of ERK and resulted in inhibition of downstream molecules by the ERK inhibitor U0126.
Conclusions
These findings suggest that CMA improves odontogenic differentiation and mineralization of hDPCs through the ERK signaling pathway.
Similar content being viewed by others

Background
Dental pulp is made up of loose connective tissue. However, in comparison to other types of connective tissue, dental pulp has several unique properties, such as the presence of odontoblasts, absence of histamine-releasing mast cells, tissue confinement in a hard cavity with little collateral circulation, and vascular access that is limited to the root apex [1]. Dental pulp is often subject to damage or injury, and, in most cases, dental pulp cells can differentiate to odontoblast-like cells, secreting a reparative or tertiary dentin [2, 3]. This suggests that the dentin–pulp complex has regenerative potential in response to injury.
Dental tissue has functional, developmental, and anatomical similarities to bone. The maintenance of hard tissue in the tooth or bone depends upon the stimulation of morphologically and functionally related cells, odontoblasts or osteoblasts, which are derived from the same mesenchymal stem cells [4]. Odontoblasts and osteoblasts secrete the same collagenous and non-collagenous proteins, such as osteonectin, osteocalcin (OCN), osteopontin, and bone sialoprotein [5]. Dentin matrix protein-1 (DMP-1) and dentin sialophosphoprotein (DSPP) were originally considered dentin-specific non-collagenous proteins [6], but several studies have revealed that DMP-1 and DSPP are also expressed in bones [7]. Therefore, these proteins have been used as mineralization markers for the odontogenic or osteogenic differentiation of human dental pulp cells (hDPCs).
Many studies have demonstrated that hard tissue formation is stimulated by gonadal steroid hormones, including estrogens and androgens [8]. Patients with estrogen deficiency often experience bone loss [9], and patients with postmenopausal osteoporosis show reduced bone mineral density [10]. Androgen-deficient patients suffer from reduced bone mass [11, 12]. These results suggest that sex hormones have an important role in osteogenic differentiation and bone tissue metabolism.
Chlormadinone acetate (CMA) is a derivative of naturally secreted progesterone, one of the fundamental female hormones. CMA has been used as an oral contraceptive in hormone replacement therapy, and in combination with estrogen in contraception [13]. CMA shows high contraceptive efficacy as it inhibits ovulation through suppression of endogenous gonadotropin secretion and follicular growth and maturation [14]. A recent study reported that CMA promotes osteogenic differentiation and calcium deposition in human bone marrow-derived mesenchymal stem cells (hBMSCs) [15].
To date, there has been no study examining the effects of CMA on the odontogenic differentiation of hDPCs. Thus, the aim of this study was to investigate the effects of CMA on differentiation and mineralization of hDPCs and the role of extracellular signal-regulated kinase (ERK) as a mediator of CMA-stimulated odontogenic differentiation in hDPCs. The null hypothesis was that CMA has no effect on the odontogenic differentiation and mineralization of hDPCs.
Methods
Cell isolation and culture
Freshly extracted third molars from healthy patients were obtained from the Department of Oral and Maxillofacial Surgery, Chonnam National University Dental Hospital (Gwangju, Republic of Korea). Teeth were split under sterile conditions and pulp tissue was minced and plated in a 100-mm culture plate (Nunc, Roskilde, Denmark). Cells were cultured in α-minimum essential medium (α-MEM; Gibco) supplemented with 10% fetal bovine serum (FBS; Gibco, Invitrogen), 100 U/mL penicillin and 100 mg/mL streptomycin (Gibco, Invitrogen) in a humidified atmosphere of 5% CO2 at 37 °C. Cells passaged 3 to 5 times were used in this study. For mineralization experiments, cells were cultured in odontogenic induction media (OIM) with 50 μg/mL ascorbic acid (Sigma-Aldrich, St Louis, MO, USA) and 10 mmol/L β-glycerophosphate (Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
Cell viability assay
Cells were seeded in 96-well culture plates at a density of 1 × 104 cells per well. Then, the cells were exposed to 0, 0.01, 0.1, 1, or 10 μM CMA (Sigma-Aldrich) for 48 h. Cell viability was examined using an EZ-Cytox cell viability assay kit (Daeil Lab Service, Seoul, Korea) according to the manufacturer’s recommendations. Briefly, 10 μL Ez-Cytox (tetrazolium salt) was added to the medium, and the cells were incubated at 37 °C for 3 h. Absorbance was measured at 420 nm using a spectrophotometer (VERSAmax multiplate reader; Molecular Devices, Sunnyvale, CA, USA).
Quantitative real-time polymerase chain reaction (PCR)
Cells were seeded in 6-well culture plates at a density of 2 × 105 cells per well. The cells were exposed to differentiation medium containing 0, 0.1, 1, or 10 μM CMA for 2, 5 and 7 days. Total RNA was isolated with the TRIzol reagent (Gibco, Invitrogen) according to the manufacturer’s instructions. cDNA was synthesized using the Maxime RT PreMix Kit (iNtRON Biotech, Seongnam, Korea). Quantitative real-time PCR was conducted using the QuantiTect SYBR Green PCR Kit (Qiagen, Valencia, CA, USA) in triplicate in a Rotor- Gene 6000 (Corbett Research, Sydney, Australia). The primer sequences are detailed in Table 1. All quantified values were normalized to endogenous β-actin. The data for gene expression were analyzed by the ΔΔCt method as described previously [16].
ALP staining
Cells were seeded in 24-well culture plates at a density of 2 × 104 cells per well with differentiation medium containing 0, 0.1, 1, and 10 μM CMA with or without pretreatment with ERK inhibitor (U0126) for 7 days. Cultured hDPCs in OIM were used as a positive control. After 7 days, the samples were washed with PBS and fixed with 70% ice-cold ethanol, rinsed 3 times with deionized water, and then treated with 300 μL of ALP staining solution (1-Step NBT/BCIP Solution; Thermo Fisher Scientific Inc, Rockford, IL, USA) for 15 min. ALP staining was photographed using an Officejet pro L7580 scanner (HP, Palo Alto, CA, USA). For quantitative analysis, the stains were extracted with 10% (w/v) cetylpyridium chloride in 10 mmol/L sodium phosphate (pH = 7.0) for 15 min. ALP staining was quantified by measuring absorbance at 540 nm using a spectrophotometer (VERSAmax Multiplate Reader).
Alizarin red staining
For alizarin red staining, hDPCs were cultured as mentioned with ALP staining. After 14 days, hDPCs were stained with 2% alizarin red stain solution (LIFELINE Cell Tech, Frederick, MD, USA) for 20 min and washed five times with sterile water. Alizarin red staining was photographed and staining density was quantified using an image analyzing program (Image J; National Institutes of Health, Bethesda, MD, USA).
Western blot analysis
After hDPCs were exposured to OIM containing 1 μM CMA, cell lysates were prepared by solubilizing the cells with 1 mL of PBS-TDS (PBS, 1% Triton X-100, 0.05% sodium deoxycholate, 0.01% SDS, 0.5 μg/ml leupeptin, 1 mM EDTA, 1 μg/ml pepstatin, 0.2 mM PMSF) for 15 min on ice. The lysates were centrifuged at 12,000 rpm for 10 min to remove cell membranes, and protein concentrations were determined with a BCA assay kit (Sigma-Aldrich) using bovine serum albumin (BSA) as a reference point. Samples containing equal amounts of protein were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose transfer membranes (Bio-Rad, Hercules, USA) for 2 h at 100 V. The membranes were blocked with 5% non-fat dry milk in PBS-T (PBS, 0.1% Tween 20) for 1 h at room temperature. Then, the membranes were incubated with anti-ERK and anti-phospho-ERK (Cell signaling, Denver, MA, USA) for 1 h with the primary antibodies, which were diluted 1:1000 in PBS. Then, horseradish peroxidase (HRP)-conjugated secondary antibodies, anti-mouse IgG or anti-rabbit IgG (Sigma-Aldrich), were used at a 1:5000 dilution for 1 h at room temperature. After three washes, chemiluminescent HRP (Millipore Corporation, Billerica, MA, USA) was applied for 30 to 60 s, and luminescence was detected with a Chemiluminescence Imaging System (Ez-capture; Atto, Tokyo, Japan).
Statistical analysis
Experiment was performed at least twice which consisted of triplicate independent test. One-way ANOVA was performed and Tukey’s test was used for post hoc analysis. The SPSS 18.0 software program was used for all analysis (SPSS, Chicago, IL, USA). Differences were considered significant at p < .05.
Results
Cytotoxicity of CMA
To evaluate cellular viability following treatment with CMA, the WST-1 assay was performed (Fig. 1). Cell viability was not inhibited by CMA. There was no statistically significant difference between the untreated group and the CMA treated groups (p > .05).

Effect of CMA on cell viability as measured by the WST-1 assay. Results are expressed as relative cell viability (percentage of control). There was no statistically significant difference between the groups (p > .05.)
Effects of CMA on odontogenic differentiation
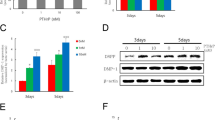
To investigate the effect of CMA on odontogenic differentiation in hDPCs, we assessed the levels of mRNA expression of odontogenic marker genes by quantitative real-time PCR (Fig. 2). The mRNA levels of the ALP gene increased significantly after 7 days of 10 μM CMA treatment. OCN mRNA levels increased significantly after treatment with 1 and 10 μM CMA at 5 days. DSPP and DMP-1 gene expression increased significantly with treatment of 10 μM CMA for 5 days (p < .05).

Expression profiles of ALP, OCN, DSPP, and DMP-1 during odontogenic differentiation by CMA in hDPCs, as measured by quantitative real-time PCR. The relative expression levels of a ALP, b OCN, c DSPP, and d DMP-1 genes were normalized to that of a housekeeping gene (β-actin). *Significant differences compared with the control (p < .05)
Effects of CMA on mineralization
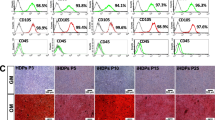
To investigate whether CMA regulates mineralization in hDPCs, we examined ALP activity by ALP staining and calcium nodule deposition by Alizarin red staining (Fig. 3). CMA up-regulated ALP activity at concentrations of 0.1 and 1 μM (Fig. 3a), and increased matrix mineralization at 0.1, 1, and 10 μM (Fig. 3b). These values were significantly different when compared with OIM alone (p < .05).

ALP activity evaluated by ALP staining and calcium nodule deposition evaluated by Alizarin red staining during odontogenic differentiation by CMA in hDPCs. a CMA increased ALP activity at concentrations of 0.1 and 1 uM. b CMA increased mineralized nodule formation at concentrations of 0.1, 1, and 10 uM. Quantification of ALP (c) and Alizarin red staining (d). OIM, odontogenic induction media.*Significant differences compared with cells treated with OIM alone (p < .05)
Effects of CMA on ERK signaling pathway in hDPCs
To investigate the signaling pathways involved in CMA-mediated odontogenic differentiation of hDPCs, we examined the phosphorylation of ERK by Western blot. CMA treatment increased the phosphorylation of ERK within 10 min (Fig. 4a). U0126 is a selective ERK inhibitor. ERK phosphorylation following exposure to 1 μM CMA was inhibited by U0126 in dose-dependent manner (Fig. 4b). Furthermore, 10 μM U0126 significantly decreased CMA-induced ALP activity (Fig. 4c and d). These results suggest that CMA increases ERK phosphorylation.

The signaling pathway of CMA-stimulated odontogenic differentiation in hDPCs, as determined by Western blot analysis and ALP staining. a Phosphorylation of ERK increased within 10 min and gradually decreased in a time-dependent manner. b ERK phosphorylation-dependent CMA expression was suppressed in a dose-dependent manner with U0126, an ERK inhibitor. c 10 uM U0126 significantly decreased CMA-induced odontogenesis. d Quantification of ALP staining by densitometry. *Significant differences compared with the control (p < .05)
Discussion
CMA is a derivative of 17-hydroxyprogesternoe, which is chlorinated in position 6 and has an α-acetoxy-group at position 17. It was first synthesized in 1961 and has been used as an oral contraceptive. Previous studies suggested that CMA has strong progestogenic activity and moderate anti-androgenic properties [13]. However, no previous studies have investigated the effect of CMA on odontogenic differentiation of hDPCs. Only one report suggested that CMA promotes osteogenic differentiation and mineralization through stimulation of the differentiation of hBMSCs into mature osteoblasts through the ERK signaling pathway [15]. Because DPCs and BMSCs are very similar [17], we hypothesized that CMA plays a significant role in odontogenic differentiation in hDPCs.
In this study, ALP, OCN, DSPP, and DMP-1 were selected as specific odontoblast markers. ALP, which plays an important role in mineral deposition, is considered an early marker of osteo/odontogenic differentiation [18]. OCN, which regulates the mineral phase of bone and dentin, and DSPP, which is involved in the dentin mineralization process, are markers of late stages of osteo/odontogenic differentiation [19, 20]. DMP-1, which is predominantly expressed in odontoblasts, is required during the early and late stages of osteo/odontogenic differentiation [21]. In our study, we observed upregulation of odontogenic markers in the CMA-treated group after 5 or 7 days. Our results are consistent with those of previous reports involving hBMSCs [15].
We next investigated whether CMA regulates mineralization in hDPCs by ALP staining and Alizarin red staining. During early odontogenic differentiation, ALP can be used as a marker of specific proteins associated with pulp cells [22]. Alizarin red is an early-stage marker of matrix mineralization, which is a crucial step in the formation of calcified extracellular matrix in odontogenic differentiation [23]. Therefore, ALP activity and alizarin red staining can indirectly represent the mineralization capacity and differentiation ability of cells. In the present study, CMA-stimulated hDPCs showed increased ALP activity and formation of mineralized nodules. These results indicate that CMA is able to promote the mineralization and differentiation of hDPCs.
Mitogen-activated protein kinases (MAPKs) are serine/threonine kinases that are an essential component of many physiological processes, such as cell growth, proliferation, differentiation, and apoptosis [24]. The MAPK family is comprised of three actors, extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 protein kinase [25]. We postulated that ERK may be involved in CMA-induced differentiation because the ERK pathway has been reported to contribute to odontogenic differentiation in hDPCs [26, 27]. Our results showed that CMA increased ERK phosphorylation within 10 min. In addition, a specific antagonist of ERK, U0126, inhibited CMA-induced phosphorylation of ERK and mineralization. These results suggest that the ERK pathway is a regulator of CMA-induced odontogenic differentiation in hDPCs.
As yet, there has been no study on the effects of progesterone, which is a component of CMA, on human dental pulp. Previous study only reported that the progesterone receptor is present in human dental pulp [28]. Progesterone plays an important role in the periodontium and bone. Progesterone alters periodontal ligament fibroblast metabolism and increases vascular permeability in periodontal tissue [29], and it activates bone metabolism and inhibits bone resorption [30]. Based on these results, we expected that progesterone would have an effect on dental pulp cells, including periodontium and bone. In this study, CMA was shown to promote odontogenic differentiation of hDPCs, as evidenced by the formation of mineralized nodules, the induction of ALP phenotypes, and upregulation of odontogenic markers. For the clinical usage, CMA should be used in local application because there might be some side effect when used in systemic application. The topical application of CMA on the exposed pulp tissue may be considerable. In this respect, if CMA is delivered to the pulp-dentin complex properly, CMA may provide a therapeutic effect for the regeneration of dental pulp tissues.
Conclusion
Collectively, the current study demonstrated that CMA increased odontogenic differentiation markers and mineralization nodule formation, and these are regulated by the ERK signaling pathway. These results suggest that CMA may play a role in regeneration of pulp-dentin complex.
Abbreviations
- ALP:
-
Alkaline phosphatase
- CMA:
-
Chlormadinone acetate
- DMP:
-
Dentin matrix protein
- DSPP:
-
Dentin sialophosphoprotein
- ERK:
-
Extracellular signal-regulated kinase
- FBS:
-
Fetal bovine serum
- GM:
-
Growth media
- hBMSC:
-
Human bone marrow-derived mesenchymal stem cells
- hDPC:
-
Human dental pulp cell
- HRP:
-
Horseradish peroxidase
- IRB:
-
Institutional review board
- JNK:
-
c-Jun N-terminal kinase
- MAPK:
-
Mitogen-activated protein kinases
- MEM:
-
Minimum essential medium
- OCN:
-
Osteocalcin
- OIM:
-
Odontogenic induction media
- PBS:
-
Phosphate-buffered saline
- WST:
-
Water-soluble tetrazolium
References
Seltzer S, Bender IB. The dental pulp: biologic considerations in dental procedures. Baltimore: Lippincott Williams & Wilkins; 1984.
Cooper PR, Takahashi Y, Graham LW, Simon S, Imazato S, Smith AJ. Inflammation–regeneration interplay in the dentine–pulp complex. J Dent. 2010;38:687–97.
Gronthos S, Mankani M, Brahim J, Robey PG, Shi S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci. 2000;97:13625–30.
Dale JB, Sarich SL, Bretz TM, Hatton JF, Zachow RJ. Hormonal regulation of androgen receptor messenger ribonucleic acid expression in human tooth pulp. J Dent Res. 2002;81:360–5.
Butler WT. Dentin matrix proteins and dentinogenesis. Connect Tissue Res. 1995;33:59–65.
Bègue‐Kirn C, Ruch JV, Ridall AL, Butler WT. Comparative analysis of mouse DSP and DPP expression in odontoblasts, preameloblasts, and experimentally induced odontoblast‐like cells. Eur J Oral Sci. 1998;106:254–9.
Qin C, Brunn JC, Cadena E, Ridall A, Tsujigiwa H, Nagatsuka H, et al. The expression of dentin sialophosphoprotein gene in bone. J Dent Res. 2002;81:392–4.
Bland R. Steroid hormone receptor expression and action in bone. Clin Sci. 2000;98:217–40.
Riggs BL, Khosla S, Melton III LJ. Sex steroids and the construction and conservation of the adult skeleton. Endocr Rev. 2002;23:279–302.
Nakamura T, Imai Y, Matsumoto T, Sato S, Takeuchi K, Igarashi K, et al. Estrogen prevents bone loss via estrogen receptor α and induction of Fas ligand in osteoclasts. Cell. 2007;130:811–23.
Orwoll ES. Androgens as anabolic agents for bone. Trends Endocrinol Metab. 1996;7:77–84.
Raisz LG, Wiita B, Artis A, Bowen A, Schwartz S, Trahiotis M, et al. Comparison of the effects of estrogen alone and estrogen plus androgen on biochemical markers of bone formation and resorption in postmenopausal women. J Clin Endocrinol Metab. 1996;81:37–43.
Druckmann R. Profile of the progesterone derivative chlormadinone acetate—Pharmocodynamic properties and therapeutic applications. Contraception. 2009;79:272–81.
Bouchard P. Chlormadinone acetate (CMA) in oral contraception–a new opportunity. Eur J Contracept Reprod Health Care. 2005;10:7–11.
Kim JM, Lee JE, Ryu SH, Suh PG. Chlormadinone acetate promotes osteoblast differentiation of human mesenchymal stem cells through the ERK signaling pathway. Eur J Pharmacol. 2014;726:1–8.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− (Delta Delta C(T)) method. Methods. 2001;25:402–8.
Ito K, Yamada Y, Nakamura S, Ueda M. Osteogenic potential of effective bone engineering using dental pulp stem cells, bone marrow stem cells, and periosteal cells for osseointegration of dental implants. Int J Oral Maxillofac Implants. 2011;26:947–54.
Garimella R, Bi X, Anderson HC, Camacho NP. Nature of phosphate substrate as a major determinant of mineral type formed in matrix vesicle-mediated in vitro mineralization: an FTIR imaging study. Bone. 2006;38:811–7.
Zhang W, Walboomers XF, van Osch GJ, van den Dolder J, Jansen JA. Hard tissue formation in a porous HA/TCP ceramic scaffold loaded with stromal cells derived from dental pulp and bone marrow. Tissue Eng Part A. 2008;14:285–94.
Bakopoulou A, Leyhausen G, Volk J, Tsiftsoglou A, Garefis P, Koidis P. Comparative analysis of in vitro osteo/odontogenic differentiation potential of human dental pulp stem cells (DPSCs) and stem cells from the apical papilla (SCAP). Arch Oral Biol. 2011;56:709–21.
Narayanan K, Gajjeraman S, Ramachandran A, Hao J, George A. Dentin matrix protein 1 regulates dentin sialophosphoprotein gene transcription during early odontoblast differentiation. J Biol Chem. 2006;281:19064–71.
Goldberg M, Smith AJ. Cells and extracellular matrices of dentin and pulp: a biological basis for repair and tissue engineering. Crit Rev Oral Biol Med. 2004;15:13–27.
Magne D, Bluteau G, Lopez-Cazaux S, Weiss P, Pilet P, Ritchie HH. Development of an odontoblast in vitro model to study dentin mineralization. Connect Tissue Res. 2004;45:101–8.
Neary JT. MAPK cascades in cell growth and death. Physiology. 1997;12:286–93.
Johnson GL, Lapadat R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science. 2002;298:1911–2.
Zhao X, He W, Song Z, Tong Z, Li S, Ni L. Mineral trioxide aggregate promotes odontoblastic differentiation via mitogen-activated protein kinase pathway in human dental pulp stem cells. Mol Biol Rep. 2012;39:215–20.
Ye L, Peng L, Tan H, Zhou X. HGF enhanced proliferation and differentiation of dental pulp cells. J Endod. 2006;32:736–41.
Whitaker SB, Singh BB, Weller RN, Bath KR, Loushine RJ. Sex hormone receptor status of the dental pulp and lesions of pulpal origin. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 1999;87:233–7.
Nanba H, Nomura Y, Kinoshita M, Shimizu H, Ono K, Goto H, et al. Periodontal tissues and sex hormones. Effects of sex hormones on metabolism of fibroblasts derived from periodontal ligament. Nihon Shishubyo Gakkai kaishi. 1989;31:166–75.
Gallagher JC, Kable WT, Goldgar D. Effect of progestin therapy on cortical and trabecular bone: comparison with estrogen. Am J Med. 1991;90:171–8.
Acknowledgments
None.
Funding
This study was financially supported by Chonnam National University, 2016 and a grant (CRI 16041–1) Chonnam National University Hospital Biomedical Research Institute and National Research Foundation of Korea (NRF) grant funded by the Korea government (MEST) (No. 2011–0030121).
Availability of data and materials
The datasets generated or analysed during the current study available from the corresponding author on reasonable request.
Authors’ contribution
Conception and design of study: KSM, YCH, and WMO. Acquisition and interpretation of data: INH, HSC, and JTK. Performing most of the experiments: SMK and BNL. Writing of the manuscript: SMK, BNL, KSM, and YCH. All authors approved the manuscript for publication. Kim SM and Lee BN contributed equally to this work as first authors. Min KS and Hwang YC contributed equally to this work as corresponding authors.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All procedures were conducted with informed consent, and the experimental protocol was approved by the Committee of Ethics of the Chonnam National University Dental Hospital (IRB#: CNUDH-2016-009).
The same dentist performed all extraction procedures using the standard surgical technique. Briefly, patients’ complete medical history was reviewed, identifying their age, gender, and oral hygiene status. The anesthetic used was 2% lidocaine with 1:100,000 epinephrine. Then, a straight elevator and molar extraction forceps were used to luxate the tooth and finish the extraction, respectively. Additionally, patients received a follow-up evaluation from the same clinician.
All patients were aware of the purpose of the present study and provided written consent of their participation.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Se-Min Kim and Bin-Na Lee are First authors.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Kim, SM., Lee, BN., Koh, JT. et al. The effect of chlormadinone acetate on odontogenic differentiation of human dental pulp cells: in vitro study. BMC Oral Health 17, 89 (2017). https://doi.org/10.1186/s12903-017-0379-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12903-017-0379-0