
Abstract
Background
Type 2 diabetes mellitus (T2DM) and its related complications contribute to the high morbidity and mortality in worldwide. Skeletal muscle insulin resistance plays a critical role in the onset of T2DM due to the decreasing in the insulin-stimulated glucose uptake. T2DM is associated not only with the inherited factors but also with the noninherited factors. However, the susceptibility genes related with the two factors and the transcription factors (TF) regulating the susceptibility genes in skeletal muscle, which aggravate the development of T2DM were still ill-defined.
Methods
In the present study, the expression profiles by the array of GSE25462 were retrieved from the GEO database. GEO2R was performed to validate the susceptibility differentially expressed genes (SDEG) in skeletal muscle of T2DM. Gene Ontology (GO) analysis and The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis were conducted via The Database for Annotation, Visualization, and Integrated Discovery (DAVID). A Protein-Protein Interaction (PPI) network was performed with the STRING.
Results
With the performance of GEO2R, 229 SDEGs in skeletal muscle of T2DM were identified. The biological processes (BP) of SDEGs was enriched in the cellular response to UV-B most significantly. KEGG pathway analysis revealed that the SDEGs were most significantly enriched in glycosaminoglycan degradation. 5 hub susceptibility genes (GPR84, CALCB, GCG, PTGDR, GNG8) in the skeletal muscle of T2DM were identified. Eventually, the common transcription factors regulating the hub susceptibility genes were identified by means of the online tool PROMO.
Conclusions
Five hub susceptibility genes (GPR84, CALCB, GCG, PTGDR, GNG8) in the skeletal muscle of T2DM and the common transcription factors were identified. The outputs would provide new clues on the novel potential targets and the therapeutic strategies for treating T2DM and its related diseases.
Similar content being viewed by others


Introduction
Type 2 diabetes mellitus (T2DM) and its related complications, such as skeletal muscle atrophy, diabetic cardiomyopathy and tumors, contribute to the high morbidity and mortality in worldwide [1,2,3]. Meanwhile, during the pathophysiology of T2DM, a chronic metabolic disordered process, skeletal muscle plays a crucial role, mostly because skeletal muscle is one of the main insulin-sensitive tissues which uptakes glucose by responding to insulin stimulation via glucose transporter 4 (GLUT4). Skeletal muscle insulin resistance, which is defined as less sensitivity of skeletal muscle to normal insulin concentration, aggravates the development of T2DM [4]. Therefore, skeletal muscle is emerging as a promising therapeutic target for T2DM and its related metabolic disorders, such as adipose dysfunction and alcoholic fatty liver disease (NAFLD )[5].
Transcription factors (TFs) are a certain kind of cellular proteins that bind to the specific promoter regions of DNA, which lie upstream of the coding region in a gene to regulate the transcriptional machinery [6]. TF is emerging as an important driver in the pathophysiology of T2DM, and it would be of importance to identify the TF associated with inherited factors.
T2DM is a complex disease due to the interplay between the inherited factors and noninherited factors [7, 8], which makes it to be of great significance to identify the susceptibility differentially expressed genes (SDEG) in skeletal muscle of T2DM. To better figure out the mechanisms of SDEGs, the common TFs regulating the SDEGs are necessary to be identified simultaneously.
The SDEGs and the common TFs will be possibly emerging as the potential targets to treat T2DM. The identification of the SDEGs and TFs related with the interaction between the inheritance and the noninherited factors on the development of T2DM would provide new clues for the potential therapeutic strategies for the treatment of T2DM and its related disorders.
In the present study, to identify the SDEGs of T2DM, the datasets of the GSE25462 were retrieved from Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo/), an international public repository providing freely high-throughput microarray [9]. With the performance of the web tool GEO2R, both the differentially expressed genes (DEGs) in skeletal muscle between T2DM subjects and the normoglycemic insulin-resistant subjects with parental family history (PFH) of T2DM subjects, and the DEGs between T2DM subjects and the normoglycemic insulin resistant subjects without PFH of T2DM subjects were identified. The overlap of DEGs of the two groups was considered as the SDEGs associating with the interplay of the inheritance and the noninherited factors in skeletal muscle of T2DM. In the present study, the 5 hub susceptibility genes (GPR84, CALCB, GCG, PTGDR, GNG8) and the common transcription factors regulating the hub susceptibility genes in skeletal muscle were identified with the performance of the various bioinformatics methods. Our outputs would hold promise for the novel therapeutic strategies for the treatment of T2DM and its related diseases.
Materials and methods
Microarray data archives
To identify SDEGs in skeletal muscle of T2DM, the expression profiles by array of GSE25462 were retrieved from GEO database. The GSE25462 collected 10 female skeletal muscle samples from T2DM subjects (T2DM), 25 female skeletal muscle samples from the normoglycemic insulin resistant (IR) subjects with PFH of T2DM (PFH), and 15 female skeletal muscle samples from the normoglycemic IR subjects without PFH of T2DM (nonPFH). The BMI, fasting glucose, fasting insulin in T2DM subjects were higher than the other two groups. The BMI and fasting glucose had no difference between PFH and nonPFH subjects, However, the fasting insulin in PFH subjects were higher than the PFH subjects group (Figure S1), which were described previously in the GEO database GSE25462 (http://www.ncbi.nlm.nih.gov/geo). The expression profiling of the database was based on GPL570 (Affymetrix Human Genome U133 Plus 2.0 Array) platform. The series matrix files and the data table header descriptions of the database were downloaded from the GEO database to screen and to verify the SDEGs associating with the interplay of the inheritance and the noninherited factors in skeletal muscle of T2DM.
SDEGs Identification
Both the DEGs in skeletal muscle between T2DM subjects and the normoglycemic insulin-resistant subjects with PFH of T2DM (PFH vs T2DM), and the DEGs between T2DM subjects and the normoglycemic insulin-resistant subjects without PFH of T2DM (non-PFH vs T2DM) were identified, respectively, with the performance of the web tool GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r/), an online tool designed to compare the different groups of samples [10]. The p<0.05 and |log FC (fold change) | > 1 were used as cut-off criteria and defined as a statistically significant difference. The overlap of DEGs between PFH vs T2DM and non-PFH vs T2DM was considered as the SDEGs associating with the interaction of the inheritance and the noninherited factors in skeletal muscle during the development of T2DM.
GO and pathway enrichment analyses
GO and KEGG pathway analyses of the SDEGs in skeletal muscle of T2DM were performed via The Database for Annotation, Visualization, and Integrated Discovery (DAVID 6.8, http://david.ncifcrf.gov) [11]. GO is a commonly used bioinformatic tool that provides comprehensive information on gene function of individual genomic products based on defined features. The GO analysis consists of biological processes (BP), and cellular components (CC), molecular functions (MF). KEGG is a significant database resource for understanding high-level biological functions and utilities. Gene count >2 and p < 0.05 were set as the threshold.
The PPI network Creation and the hub susceptibility gene identification
A PPI network of the SDEGs in skeletal muscle of T2DM was constructed by Search Tool for the Retrieval of Interacting Genes (STRING10.5; https://string-db.org/) with a combined score >0.4 as the cut-off point [12]. The hub susceptibility genes were identified using Cytohubba, a plug-in of Cytoscape software (Cytoscape, 3.7.1) and the significant modules in the PPI network were identified by molecular complex detection (MCODE 1.5.1), another plug-in of Cytoscape software [13, 14]. The parameters of SDEGs clustering and scoring were set as follows: MCODE score ≥4, degree cut-off = 2, node score cut-off = 0.2, max depth = 100, and k-score = 2.
The common transcription factors of the hub susceptibility genes prediction
Following the promotor sequences of the hub susceptibility genes were obtained from the UCSC Genome Browser database (http://genome.ucsc.edu) [15], the hub susceptibility gene promotors were submitted to the PROMO databases (Transfac 8.3), a freely available online tool, to predict the putative transcription factors binding motifs of each hub susceptibility genes [16]. The maximum matrix dissimilarity rate was set as 0. The overlap of the transcription factors of each hub susceptibility gene was identified as the common putative transcription factors of the hub susceptibility genes.
Statistical analysis
The statistical analyses of DEGs were done with GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r/), an online tool designed to compare the different groups of samples. The p<0.05 and |log FC (fold change) | > 1 were used as cut-off criteria and defined a statistically significant difference. Gene count >2 and p < 0.05 were set as the threshold in the GO and KEGG analysis with the performance of the DAVID database.
Results
Identification of the SDEGs in skeletal muscle of T2DM
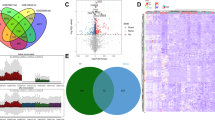
To identify the SDEGs in skeletal muscle of T2DM, relevant microarray expression profiles of GSE25462 was retrieved from GEO database. 704 DEGs were identified between T2DM subjects and the normoglycemic IR subjects with PFH of T2DM (PFH vs T2DM) (Table S1), and 767 DEGs were identified between T2DM subjects and the normoglycemic IR subjects without PFH of T2DM (non PFH vs T2DM) (Table S2). 229 SDEGs were included in the overlap of two groups of the DEGs in skeletal muscle of T2DM (Fig. 1, Table 1).

Venn diagram showing 704 DEGs were identified betweenT2DM subjects and the normoglycemic IR subjects with PFH of T2DM, and 767 DEGs were identified between T2DM subjects and the normoglycemic IR subjects without PFH of T2DM. 229 SDEGs were included in the overlap of two groups of the DEGs. A: PFH vs T2DM (DEGs: T2D subjects vs the normoglycemic IR subjects with PFH of T2D); B: nonPFH vs T2DM DEGs (T2DM subjects vs the normoglycemic IR subjects without PFH of T2DM). DEG: differentially expressed gene; C: SDEG: susceptibility differentially expressed gene
GO enrichment analysis of SDEGs in skeletal muscle of T2DM
To figure out the biological features of the SDEGs in skeletal muscle of T2DM, GO analysis was accomplished by the DAVID online tool. BP terms indicated that the SDEGs were the most significantly enriched in cellular response to UV-B and positive regulation of keratinocyte proliferation, potassium ion transmembrane transport (Fig. 2). The CC analysis showed that SDEGs were enriched in extracellular space, cell surface, extracellular region (Fig. 2,). Changes in MF of SDEGs were major enriched in zinc ion binding, heparin binding, calcium ion binding. (Fig. 2). The genes that were included GO enrichment analysis of SDEGs in skeletal muscle of T2DM were presented in Table 4.

GO enrichment result of SDEGs. Abscissa represents different adjusted p value, and ordinate represents GO terms. Different colors stand for different GO classifications. GO: Gene Ontology; SDEG: susceptibility differentially expressed gene
KEGG enrichment analysis of SDEGs in skeletal muscle of T2DM
To explore the potential mechanism of the SDEGs in skeletal muscle of T2DM, KEGG pathway analysis was performed using DAVID online tools. KEGG analysis revealed that SDEGs were mainly involved in glycosaminoglycan degradation, gastric acid secretion and pancreatic secretion (Fig. 3). The genes that were included KEGG enrichment analysis of SDEGs in skeletal muscle of T2DM were presented in Table 5.

KEGG enrichment result of SDEGs. Abscissa represents different adjusted p value, and ordinate represents KEGG terms. KEGG: Kyoto Encyclopedia of Genes and Genomes SDEG: susceptibility differentially expressed gene
Identification of the hub susceptibility genes in skeletal muscle of T2DM
To identify the most significant clusters of the SDEGs, a PPI network of SDEGs was constituted by STRING. As shown in Fig. 4, there were 176 nodes and 116 edges in the PPI network. The most significant module (score=5) was recognized by MCODE, a plug-in of Cytoscape (Fig. 5). Five hub susceptibility genes involved in SDEGs were identified, including GPR84, CALCB, GCG, PTGDR, GNG8 (Table 2).

Results of the PPI network. The PPI network was analyzed by String software. Here were 176 nodes and 116 edged in the PPI network. PPI: protein–protein interaction. The color of the string maps was based on the score value with which it interacted

The most significant module identified by MCODE (score = 5). MCODE: molecular complex detection
Identification of the common putative transcription factors of the hub susceptibility genes in skeletal muscle of T2DM
To identify the common putative transcription factors of the hub susceptibility genes in skeletal muscle of T2DM, the gene promotors were submitted to the PROMO databases (Transfac 8.3). Each hub susceptibility gene had several putative transcription factors in the PROMO databases (Table 3). The overlap of the transcription factors was identified as the common putative transcription factors of 5 hub susceptibility genes in skeletal muscle of T2DM, including ER-alpha, YY1, GR-beta, GR-alpha, C/EBP beta, TFIID. The DNA-binding sequence logs of YY1 and C/EBP in the JASPAR database were shown in Fig. 6.

The DNA-binding sequence log of YY1, C/EBP beta in the JASPAR database
Discussion
The interplay of the diabetic inheritance and noninherited factors plays a critical role during the onset and pathophysiology of T2DM. In the present study, to better understand the hub susceptibility genes of the interaction of diabetic inheritance and noninherited factors in skeletal muscle on the onset and development of T2DM, bioinformatics analysis was performed.
In the present study, 229 SDEGs were identified in skeletal muscle of T2DM. The BP terms of GO analysis indicated that the SDEGs were significantly enriched in positive regulation of keratinocyte proliferation term. Although keratinocyte proliferation has been reported to be implicated in the development of T2DM [17], it was the first time to demonstrate that the genes of positive regulation of keratinocyte proliferation were involved in diabetic skeletal muscle. In the present study, the genes related with keratinocyte proliferation included FGF7, TP63, REG3G. Patrie et al. demonstrated that FGF7 was the ligands for the newt KGFR (keratinocyte growth factor receptor), which binded FGF7 with high affinity and mediates signaling in skeletal muscle myoblasts [18]. And Klasan et al. indicated that Reg3G gene played a major role in regenerating skeletal muscle [19]. Meanwhile, the CC terms indicated FGF7 and REG3G were found in the extracellular region term. Maybe we could hypothesize that FGF7 and REG3G could be derived from skeletal muscle cells and were transported to the keratinocyte to regulate the proliferation of the keratinocyte. The output would provide clues to explore the mechanism of diabetic keratitis.
Additionally, KEGG enrichment analysis of the SDEGs showed that these SDEGs were the most significantly mapped in the degradation of glycosaminoglycan. Glycosaminoglycan degradation could induce the decrease of proteoglycans, which were chemically diverse macromolecules to be associated with hyperglycemic conditions [20]. Yuan et al. demonstrated that glycosaminoglycan could improve insulin resistance and T2DM via enhancing liver SOD and GSH-Px activity [21]. The further connection between skeletal muscle pathology and glycosaminoglycan breakdown need to be further studied.
Based on the PPI network and the most significant module, five hub susceptibility genes in skeletal muscle of T2DM were identified in the network of SDEGs, including GPR84, CALCB, GCG, PTGDR, GNG8. GPR84 has been reported that it was highly expressed in skeletal muscle and adipose tissue [22]. GPR84 has been demonstrated to be involved in the regulation of energy metabolism mediated by the secretion of insulin and inflammatory responses related to insulin resistance [23]. Therefore, GPR84 might play a crucial role during the development of skeletal muscle insulin resistance. Although Chen and his colleagues reported that CALCB was a calcitonin gene-related peptide which was associated with the initial events triggered in T1DM [24], up to date to our knowledge, there is no more report on CALCB related with insulin resistance and T2DM. GCG has been demonstrated to be implicated in the development of T2DM [25]. In the present study, we revealed that GCG alteration in the development of T2DM was regulated by both the inheritance and the noninherited factors. The previous studies on PTGDR were focused as an asthma susceptibility gene [26, 27]. Interestingly, metformin, which improves insulin resistance and metabolic function, has been demonstrated to play the protective role on asthma [28]. Therefore, the further study on the effect of metformin on PTGDR is urgent to be performed to identify whether PTGDR is the crosstalk target of T2DM and asthma. To the best of our knowledge, except that GNG8 has not been reported to be implicated in the onset and development of T2DM, very few literatures reported any function of GNG8. It is of great significance to reveal that GNG8 was associated with the process of T2DM with the bioinformatics tool for the first time.
To better understand the roles of the hub susceptibility genes in skeletal muscle of T2DM, the TFs of each hub SDEGs were predicted. Furthermore, the common TFs of the five hub SDEGs, including ER-alpha, YY1, GR-beta, GR-alpha, C/EBP beta were predicted. All the putative TFs have been reported to be implicated in diabetes and its related diseases, controlling skeletal muscle metabolism, ameliorating diabetic nephropathy pathology, modulating human pancreas development, and leading to the improvement of metabolic health and insulin sensitivity [29,30,31]. However, there were no previous reports on the correlation between the above five hub susceptibility genes and the putative TFs.
In the present study, the limitation focused on the absence of validation of expression on mRNA lever and the absence of further study on the exact correlation and the related mechanism.
Conclusion
In the present study, five susceptibility hub genes and their related common TFs in skeletal muscle were identified to be related to the interplay of the inheritance and the noninherited factors on the T2DM. Furthermore, it is the first time that two genes, PTGDR and GNG8, were identified to be implicated in the development of T2DM. However, a further urgent study needs to be performed in the future to clarify the exact mechanisms of the hub susceptibility genes in skeletal muscle on the onset and development of T2DM.
Availability of data and materials
The datasets of the GSE25462 analysed during the current study were retrieved from Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo/). All methods were carried out in accordance with relevant guidelines and regulations.
Abbreviations
- T2DM:
-
Type 2 Diabetes Mellitus
- IR:
-
Insulin resistant
- DEG:
-
Differentially expressed genes
- SDEG:
-
Susceptibility differentially expressed genes
- TF:
-
Transcription factors
- PFH:
-
Parental family history
- GO:
-
Gene Ontology
- KEGG:
-
The Kyoto Encyclopedia of Genes and Genomes
- DAVID:
-
The Database for Annotation, Visualization, and Integrated Discovery
- BP:
-
Biological processes
- CC:
-
cellular components
- MF:
-
Molecular functions
- PPI:
-
Protein-Protein Interaction
References
Hirata Y, Nomura K, Senga Y, Okada Y, Kobayashi K, Okamoto S, et al. Hyperglycemia induces skeletal muscle atrophy via a WWP1/KLF15 axis. JCI Insight. 2019;4(4).
Borghetti G, von Lewinski D, Eaton DM, Sourij H, Houser SR, Wallner M. Diabetic Cardiomyopathy: Current and Future Therapies, Beyond Glycemic Control. Front Physiol. 2018;9:1514.
Andersen DK, Korc M, Petersen GM, Eibl G, Li D, Rickels MR, et al. Diabetes, Pancreatogenic Diabetes, and Pancreatic Cancer. Diabetes. 2017;66(5):1103–10.
DeFronzo RA, Tripathy D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care. 2009;32(Suppl 2):S157–63.
da Silva Rosa SC, Nayak N, Caymo AM, Gordon JW. Mechanisms of muscle insulin resistance and the cross-talk with liver and adipose tissue. Physiol Rep. 2020;8(19):e14607.
Desvergne B, Michalik L, Wahli W. Transcriptional regulation of metabolism. Physiol Rev. 2006;86(2):465–514.
Kelly GS. Insulin resistance: lifestyle and nutritional interventions. Altern Med Rev. 2000;5(2):109–32.
Ardisson Korat AV, Willett WC, Hu FB. Diet, lifestyle, and genetic risk factors for type 2 diabetes: a review from the Nurses' Health Study, Nurses' Health Study 2, and Health Professionals' Follow-up Study. Curr Nutr Rep. 2014;3(4):345–54.
Clough E, Barrett T. The Gene Expression Omnibus Database. Methods Mol Biol. 2016;1418:93–110.
T. Barrett, S.E. Wilhite, P. Ledoux, C. Evangelista, I.F. Kim, M. Tomashevsky, K.A. Marshall, K.H. Phillippy, P.M. Sherman, M. Holko, A. Yefanov, H. Lee, N. Zhang, C.L. Robertson, N. Serova, S. Davis, A. Soboleva, NCBI GEO: archive for functional genomics data sets--update, Nucleic Acids Res 41(Database issue) (2013) D991-D995.
W. Huang da, B.T. Sherman, R.A. Lempicki, Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists, Nucleic Acids Res 37(1) (2009) 1-13.
Franceschini A, Szklarczyk D, Frankild S, Kuhn M, Simonovic M, Roth A, et al. STRING v9.1: protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013;41(Database issue):D808–15.
Chin CH, Chen SH, Wu HH, Ho CW, Ko MT, Lin CY. cytoHubba: identifying hub objects and sub-networks from complex interactome. BMC Syst Biol. 2014;8(Suppl 4):S11.
Bandettini WP, Kellman P, Mancini C, Booker OJ, Vasu S, Leung SW, et al. MultiContrast Delayed Enhancement (MCODE) improves detection of subendocardial myocardial infarction by late gadolinium enhancement cardiovascular magnetic resonance: a clinical validation study. J Cardiovasc Magn Reson. 2012;14:83.
Meyer LR, Zweig AS, Hinrichs AS, Karolchik D, Kuhn RM, Wong M, et al. The UCSC Genome Browser database: extensions and updates 2013. Nucleic Acids Res. 2013;41(Database issue):D64–9.
Messeguer X, Escudero R, Farre D, Nunez O, Martinez J, Alba MM. PROMO: detection of known transcription regulatory elements using species-tailored searches. Bioinformatics. 2002;18(2):333–4.
Spravchikov N, Sizyakov G, Gartsbein M, Accili D, Tennenbaum T, Wertheimer E. Glucose effects on skin keratinocytes: implications for diabetes skin complications. Diabetes. 2001;50(7):1627–35.
Patrie KM, Kudla AJ, Olwin BB, Chiu IM. Conservation of ligand specificity between the mammalian and amphibian fibroblast growth factor receptors. J Biol Chem. 1995;270(48):29018–24.
Klasan GS, Ivanac D, Erzen DJ, Picard A, Takasawa S, Peharec S, et al. Reg3G gene expression in regenerating skeletal muscle and corresponding nerve. Muscle Nerve. 2014;49(1):61–8.
Hiebert LM, Han J, Mandal AK. Glycosaminoglycans, hyperglycemia, and disease. Antioxid Redox Signal. 2014;21(7):1032–43.
Yuan C, Liu P, Han X, Cui Q. Hypoglycemic effects of glycosaminoglycan from Urechis unicinctus in diabetic mice. J Med Food. 2015;18(2):190–4.
Montgomery MK, Osborne B, Brandon AE, O'Reilly L, Fiveash CE, Brown SHJ, et al. Regulation of mitochondrial metabolism in murine skeletal muscle by the medium-chain fatty acid receptor Gpr84. FASEB J. 2019;33(11):12264–76.
Puengel T, De Vos S, Hundertmark J, Kohlhepp M, Guldiken N, Pujuguet P, et al. The Medium-Chain Fatty Acid Receptor GPR84 Mediates Myeloid Cell Infiltration Promoting Steatohepatitis and Fibrosis. J Clin Med. 2020;9(4).
Chen Y, Lin Y, Wang J, Guo X, Guo Y, Dong F, et al. The serum concentration of betaCGRP is novel marker for type 1 diabetes. Heliyon. 2020;6(1):e03223.
Zhang L, Zhang M, Wang JJ, Wang CJ, Ren YC, Wang BY, et al. Association of TCF7L2 and GCG Gene Variants with Insulin Secretion, Insulin Resistance, and Obesity in New-onset Diabetes. Biomed Environ Sci. 2016;29(11):814–7.
Lee YH, Choi SJ, Ji JD, Song GG. PTGDR polymorphisms and susceptibility to asthma: a meta-analysis. Mol Biol Rep. 2013;40(3):2195–203.
Garcia-Solaesa V, Sanz-Lozano C, Padron-Morales J, Hernandez-Hernandez L, Garcia-Sanchez A, Rivera-Reigada ML, et al. The prostaglandin D2 receptor (PTGDR) gene in asthma and allergic diseases. Allergol Immunopathol (Madr). 2014;42(1):64–8.
Li CY, Erickson SR, Wu CH. Metformin use and asthma outcomes among patients with concurrent asthma and diabetes. Respirology. 2016;21(7):1210–8.
Hevener AL, Ribas V, Moore TM, Zhou Z. The Impact of Skeletal Muscle ERalpha on Mitochondrial Function and Metabolic Health. Endocrinology. 2020;161(2).
Gao P, Li L, Yang L, Gui D, Zhang J, Han J, et al. Yin Yang 1 protein ameliorates diabetic nephropathy pathology through transcriptional repression of TGFbeta1. Sci Transl Med. 2019;11(510).
Phan-Hug F, Guimiot F, Lelievre V, Delezoide AL, Czernichow P, Breant B, et al. Potential role of glucocorticoid signaling in the formation of pancreatic islets in the human fetus. Pediatr Res. 2008;64(4):346–51.
Acknowledgements
Not applicable.
Funding
This work was supported by National Natural Science Foundation of China [Grant No. 81870550; 82170843; 81471858].
Author information
Authors and Affiliations
Contributions
JK and YZ drafted the protocol. JK and XH, YZ performed the searches, analyzed the data. CW and YZ drafted the manuscript. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This research does not directly contain any material obtained from animals or humans. All data used in this study were extracted from public databases.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ke, J., Hu, X., Wang, C. et al. Identification of the hub susceptibility genes and related common transcription factors in the skeletal muscle of Type 2 Diabetes Mellitus. BMC Endocr Disord 22, 276 (2022). https://doi.org/10.1186/s12902-022-01195-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12902-022-01195-0