
Abstract
Background
Somatic depression (SD) is different from non-somatic depression (NSD), and insular subregions have been associated with somatic symptoms. However, the pattern of damage in the insular subregions in SD remains unclear. The aim of this study was to use functional connectivity (FC) analyses to explore the bilateral ventral anterior insula (vAI), bilateral dorsal anterior insula (dAI), and bilateral posterior insula (PI) brain circuits in SD patients.
Methods
The study included 28 SD patients, 30 NSD patients, and 30 matched healthy control (HC) subjects. All participants underwent 3.0 T resting state functional magnetic resonance imaging. FC analyses were used to explore synchronization between insular subregions and the whole brain in the context of depression with somatic symptoms. Pearson correlation analyses were performed to assess relationships between FC values in brain regions showing significant differences and the total and factor scores on the 17-item Hamilton Rating Scale for Depression (HAMD17).
Results
Compared with the NSD group, the SD group showed significantly decreased FC between the left vAI and the right rectus gyrus, right fusiform gyrus, and right angular gyrus; between the right vAI and the right middle cingulate cortex, right precuneus, and right superior frontal gyrus; between the left dAI and the left fusiform gyrus; and between the right dAI and the left postcentral gyrus. Relative to the NSD group, the SD group exhibited increased FC between the left dAI and the left fusiform gyrus. There were no differences in FC between bilateral PI and any brain regions among the SD, NSD, and HC groups. Within the SD group, FC values between the left vAI and right rectus gyrus were positively correlated with cognitive impairment scores on the HAMD17; FC values between the right vAI and right superior frontal gyrus were positively related to the total scores and cognitive impairment scores on the HAMD17 (p < 0.05, uncorrected).
Conclusions
Aberrant FC between the anterior insula and the frontal and limbic cortices may be one possible mechanism underlying SD.
Similar content being viewed by others

Background
Major depressive disorder (MDD) is one of the most common mental disorders and is characterized by the presence of a depressive mood, loss of interest or pleasure, guilt and worthlessness, and somatic symptoms [1]. Somatic symptoms are common in patients with MDD [2]. Silverstein defined MDD patients with at least three somatic symptoms including fatigue, appetite, insomnia, unexplained breathing difficulty, poor body image, and pain, as having somatic depression (SD) [3]. Patients with SD are often misdiagnosed with other somatic diseases and repeatedly visit outpatient clinics of general hospitals [4]. Because of severe physical symptoms and poor curative effects, patients with SD are at high risk of suicidal ideation and poor quality of life [5].
Somatic symptoms are associated with greater clinical severity and lower remission rates, and pain symptoms have the greatest prognostic value [6]. Pain may be the sole complaint in SD patients who present to primary care practices and is often overlooked by clinicians [7]. The degree of pain is related to MDD severity, and is closely related to other somatic symptoms [8]. In the early stage of treatment, improvement in pain symptoms can predict higher antidepressant efficacy and a better remission rate [9]. A number of studies have reported that insomnia symptoms were independent predictors for MDD [10, 11], as well as for suicidal ideation and suicide attempts [12, 13]. Another group found a strong correlation between fatigue and depressive symptoms [14], while others showed that fatigue could be used to predict treatment efficacy and future fluctuations in patient well-being [15]. Unexplained breathing difficulties and gastrointestinal disturbances (such as poor appetite and bloating) are also prevalent in MDD, which may cause severe discomfort and result in primary health care visits. However, the pathogenesis of SD is still not fully understood, and it would be interesting and meaningful to explore the mechanisms underlying this condition.
In recent decades, researchers have used noninvasive functional neuroimaging techniques to show that SD is associated with a wide array of frontal, temporal, and parietal cortices [16,17,18]. Geng et al. found that patients with SD exhibited abnormal regional homogeneity in the frontal and temporal regions [16]. Yan et al. reported that subjects with SD exhibited abnormal amplitude of low frequency fluctuations in the right inferior temporal gyrus, left hippocampus, right inferior frontal orbital gyrus, and left thalamus [18]. Some studies have explored relationships between brain areas and somatic symptoms. For example, pain has been associated with the primary and secondary somatosensory cortices [19], the cingulate [20], and the insula [21], which belong to the so-called “pain matrix” [22]. Insomnia has been associated with impaired activity in the amygdala, thalamus, and insula [23, 24], while fatigue has been associated with impaired connectivity in the default mode network (DMN) [25] and shown to be related to decreased volumes in the prefrontal and occipital cortices [26]. Somatic symptoms include impairments in sensory, cognitive and, most importantly, affective components, which include feelings of sadness, anxiety, and depression in response to noxious stimuli [27]. Although there is no identifiable noxious stimulus in most SD patients, somatic symptoms can occur in the absence of nociceptive input in these individuals [28]. Moreover, somatic symptoms are associated with the severity of emotional symptoms [29]. Identifying which brain regions jointly regulate somatic symptoms and emotion may open new avenues into understanding the pathological mechanisms underlying SD.
The endosensory disorder hypothesis proposed by Harshaw et al. posits that the somatic symptoms of depression are related to an imbalance of internal and external perceptual functions [30]. The insula is an important brain area relevant to internal sensory functions that are responsible for processing sensory afferent information and integrating with higher cortices [31, 32]. Increasing attention has been given to the role of abnormal internal perceptual function of the insula in the pathogenesis of somatic symptoms in MDD [33,34,35]. The insula is located in the deep part of the bilateral temporal lobes and adjacent to the parietal and occipital lobes [36]. Based on the origins of the cells, the insula can be roughly divided into two subregions with functions that include sensory processing, representing feelings and emotions, motion control, body and self-awareness, risk prediction, and decision-making [37]. The anterior insula (AI) is associated with subcortical brain areas, including the anterior cingulate, ventral medial prefrontal lobe, amygdala, and ventral striatum, that integrate sensory information and participate in emotional, motivational, and cognitive functions [32, 37]. The posterior insula (PI) receives afferent information from the spinal cord and brainstem and participates in processing primary sensory information related to somatosensation and motor control [38, 39]. The AI can also be subdivided into ventral and dorsal parts: the cellular structure of the vAI is more similar to that of the marginal cortex, which may be closely related to emotion, general attention, and cognition; the AI is connected to brain areas related to the cognitive control network and may be involved more in decision-making and behaviour [40]. Zhang et al. found that weaker vAI-right orbitofrontal cortex functional connectivity (FC) strength predicted greater symptom severity [41]. Kandilarova et al. found support for the role of the right AI in the pathophysiology of MDD and found that MDD patients had significantly reduced effective connectivity strength from the AI to the dorsolateral prefrontal cortex, as well as a significant level of effective connectivity strength between the amygdala and the AI [42]. In addition, Stoyanov et al. reported that reduced effective connectivity from the AI to the dorsolateral prefrontal cortex was manifested in patients with depressive symptoms in comparison to patients with paranoid symptoms of schizophrenia [43]. These results indicated that altered effective connectivity in the AI could be a potential diagnostic biomarker to differentiate depressive symptoms [44]. Moreover, as a key node of the salience network (SN), the AI is closely related to the integration of sensory information, emotion, and cognition [45]. Thus, we suggest that abnormal FC between the insular subregions and other brain regions may lead to abnormal somatosensory perception and somatic symptoms [30]. However, to the best of our knowledge, we are not aware of any studies that have examined ventral and dorsal AI and PI FC in SD patients.
To examine how vAI, dAI, and PI affect the corresponding brain circuits in SD, this study used FC analyse of resting-state functional magnetic resonance imaging (rs-fMRI) data. FC measures the correlation coefficients of a single predefined region with another region [46], and these values provide novel insights into how distributed brain regions are functionally integrated in MDD patients [47]. We hypothesized that there would be different patterns of FC damage in different insular subregions. We also hypothesized that abnormal FC in insular subregions might be related to SD severity.
Methods
Study design
This study adopted a cross-sectional case-control study design. Data, which included demographic data, clinical assessments and MRI scan data, were collected from patients diagnosed with MDD and healthy controls (HCs).
Participants
Patients diagnosed with MDD were recruited from the Department of Psychiatry of the Affiliated Nanjing Brain Hospital of Nanjing Medical University between September 2011 and February 2017. The inclusion criteria for this population were as follows: (1) confirmation of the diagnosis of MDD according to DSM-IV-TR criteria [2], (2) age between 18 and 55 years old, (3) right-handed, (4) Chinese Han, (5) regardless of sex, (6) education level of junior high school or above, (7) Hamilton Rating Scale for Depression 17-item (HAMD17) [48] score > 17 on the day of scanning, and (8) 32-item hypomania checklist score < 14 [49], and total Young Mania Rating Scale (YMRS) score < 10 [50]. The exclusion criteria in this study were as follows: (1) psychopathologies other than MDD; (2) substance abuse/dependence within the previous 1 year; (3) history of neurological or systemic illness, head injury, or any other relevant medical or additional psychiatric disease; or (4) current pregnancy or breastfeeding.
The patients with MDD were divided into SD and nonsomatic depression (NSD) groups in accordance with previously described criteria [51]. MDD patients were assigned to the SD group when they had three or more of the following symptoms: sleep disturbance, eating disturbance, fatigue, headaches or pain, unexplained breathing difficulty, and poor body image. The patients who had two or fewer abovementioned somatic symptoms were assigned to the NSD group. The inclusion and exclusion criteria for the SD and NSD groups were the same as those we reported in another study [16]. Finally, the study included 28 patients with SD (mean age ± standard deviation = 37.21 ± 7.16 years old; mean education ± standard deviation = 13.36 ± 3.83 years; 15 females) and 30 patients with NSD (mean age ± standard deviation = 34.10 ± 9.91 years old; mean education ± standard deviation = 14.77 ± 2.94 years; 16 females).
Meanwhile, 30 HCs matched in age and sex (mean age ± standard deviation = 35.83 ± 8.15 years old; mean education ± standard deviation = 14.23 ± 2.47 years; 13 females) were recruited from the community with online and flier advertisements. Each HC was screened using the nonpatient version of the Structured Clinical Interview from the DSM-IV-TR [2], and none had any medical illness, neurological illness, psychiatric illness, or a family history of major psychiatric or neurological illness.
Written informed consent was obtained after a complete description of the study was given to all the participants. This study was approved by the Research Ethics Review Board of the Affiliated Nanjing Brain Hospital of Nanjing Medical University.
Clinical assessments
A homemade questionnaire was used to collect general subject information, including age, sex, years of education, age of onset of depression, number of depressive episodes, and treatment history. The HAMD17 and YMRS were used to assess depressive and manic symptom severity, respectively.
MRI scan acquisitions
Imaging data were obtained from 3-Tesla MRI scans performed at the Affiliated Nanjing Brain Hospital of Nanjing Medical University. The parameters for T1 anatomic axial imaging and echo-planar imaging were the same as those used in our previous articles [16, 18].
Data preprocessing
Analysis of the functional imaging data was performed using the Data Processing Assistant for Resting-State fMRI (DPARSF) (http://rfmri.org/DPARSF) [52]. The first six functional volumes were discarded to allow the participants to adapt to the scanner noise. Then, slice timing, head-motion correction, and spatial normalization to the Montreal Neurological Institute (MNI) space were conducted. The resampled voxel size was 3 × 3 × 3 mm3. These steps were the same as those in our previous research [18]. To ensure data reliability, data were included if subject head movement was < 2 mm with head rotation angle < 2 degrees. We removed several sources of variances that included head-motion parameters, averaged global blood oxygen level-dependent (BOLD) signals and mean BOLD signals in the ventricular and white matter regions. An estimate of head motion at each time point was calculated as the framewise displacement using six displacements from the rigid body-motion correction procedure [53]. The structural images were normalized to the structural (T1-weighted) MNI template. No participant was excluded due to excessive head motion or bad normalization. Then, the remaining data were smoothed using a 4-mm full-width at half maximum (FWHM) Gaussian kernel. Finally, linear detrending and temporal filtering (0.01–0.08 Hz) were performed to reduce low-frequency drift and high-frequency noise.
FC analysis
We used a seed-based approach to examine FC. First, we defined the bilateral vAI, bilateral dAI, and bilateral PI as regions of interest (ROIs). The centres of each ROI coordinate were derived from the MNI space and included the left vAI (− 33, 13, − 7), right vAI (32, 10, − 6), left dAI (− 38, 6, 2), right dAI (35, 7, 3), left PI (− 38, − 6, 5), and right PI (35, − 11, 6) [40]. Second, we used the centre of each ROI to draw a sphere with a 6-mm radius as an ROI. Third, we employed a voxelwise approach to calculate the correlation between the averaged time series of all voxels in the ROI seeds and the time series from the entire brain template. The correlation coefficients were defined as FC [46] and were converted to z values using Fisher’s transformation to improve normality.
Statistical analyses
One-way analysis of variance (ANOVA) was used to compare the age, education, total illness duration, and HAMD17 scores, while chi-square tests were used to compare the sex ratio in the SD, NSD and HC groups. The disease duration and HAMD17 factor scores were compared between the SD and NSD groups using two-sample t tests (SPSS 19.0 software, IBM Corp., Armonk, NY, USA). The level of statistical significance was set at p < 0.05 (two-tailed).
Here, we took a two-step approach. First, to compare FC values among the SD, NSD and HC groups, a one-way ANOVA model was applied to six seed-based voxelwise correlation maps using rs-fMRI data analysis toolkit (REST) software [54]. We also regressed out age, sex, and years of education as covariates using regression analysis when performing ANOVA. To control for multiple comparisons, the AlphaSim correction in the REST software was used to adjust the alpha level (4-mm FWHM, individual voxel p < 0.001, and minimum cluster size of 162 mm3) [54]. There were six cluster maps, and each map showed significant FC values. Second, for each cluster map, post hoc t tests were performed to identify differences between each pair of groups. We continued to use AlphaSim correction to adjust the alpha level (individual voxel p < 0.001, and minimum cluster size of 108 mm3) with the REST software [54].
Pearson correlation analysis was conducted to explore the possible clinical relevance of the relationship between FC alterations and SD severity. We defined the regions with significant differences in FC between the SD and NSD groups as ROIs. Then, the mean z values of the ROIs were calculated for each SD patient. Finally, Pearson correlation analyses were performed to assess abnormal mean z values of ROIs, total HAMD17 scores, and five separate symptomatic factors, including anxiety, weight loss, cognitive disturbance, retardation, and sleep disturbance in HAMD17 [48]. To control for multiple comparisons, Bonferroni correction was performed to adjust the alpha level, where the threshold was set to alpha = 0.05/6.
Results
Demographics and clinical characteristics
Fifty-eight MDD patients were enrolled in this study, including 28 in the SD group and 30 in the NSD group. Thirty HC subjects were also recruited. SPSS 19.0 software (IBM Corp., Armonk, NY, USA) was used to compare sociodemographic data. There were no statistically significant differences in sex, age, or years of education among the three groups. There was no significant difference in the disease course between the two patient groups. The anxiety/somatization factor scores on the HAMD17 in the SD group were significantly higher than those in the NSD group (t = 2.359, p = 0.022). The total scores and other factor scores on the HAMD17 were not significantly different between the two patient groups (p > 0.05); see details in Table 1.
Insula-seed FC analysis
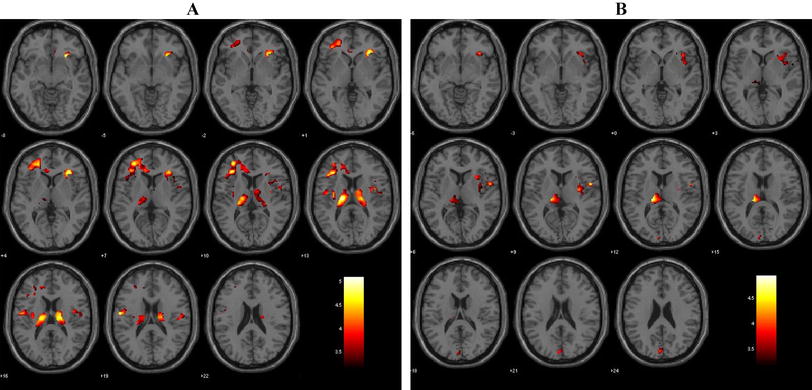
Among the three groups, there were FC differences between the left vAI and the bilateral rectus gyri, bilateral fusiform gyri, right orbital inferior frontal gyrus, and right angular gyrus (Table 2, Fig. 1A); between the right vAI and the bilateral orbital frontal gyri, right superior frontal gyrus, right middle cingulate cortex, and right precuneus (Table 3, Fig. 2A); between the left dAI and the left fusiform gyrus and left inferior frontal gyrus (Table 4, Fig. 3A); and between the right dAI and the right fusiform gyrus and left postcentral gyrus (Table 5, Fig. 4A) (p < 0.001, k > 6 voxels, p < 0.05 corrected for multiple comparisons using AlphaSim).

Brain regions showing differences in the left vAI-seed FC among the SD, NSD, and HC groups (A), between the SD and NSD groups(B), between the SD and HC groups(C), and between the NSD and HC groups(D). p < 0.05, corrected for multiple comparisons using AlphaSim correction

Brain regions showing differences in the right vAI-seed FC among the SD, NSD, and HC groups (A), between the SD and NSD groups(B), between the SD and HC groups(C), and between the NSD and HC groups(D). p < 0.05, corrected for multiple comparisons using AlphaSim correction

Brain regions showing differences in the left dAI-seed FC among the SD, NSD, and HC groups (A), between the SD and NSD groups(B), between the SD and HC groups(C), and between the NSD and HC groups(D). p < 0.05, corrected for multiple comparisons using AlphaSim correction

Brain regions showing differences in the right dAI-seed FC among the SD, NSD, and HC groups (A), between the SD and NSD groups(B), between the SD and HC groups(C), and between the NSD and HC groups(D). p < 0.05, corrected for multiple comparisons using AlphaSim correction
Compared with the NSD group, the SD group showed lower FC between the AI subregions and the frontal and limbic cortices. FC was lower between the left vAI and the right rectus, right fusiform gyrus, and right angular gyrus (Table 2, Fig. 1B); between the right vAI and the right middle cingulate cortex, right precuneus, and right superior frontal gyrus (Table 3, Fig. 2B); between the left dAI and the left fusiform gyrus (Table 4, Fig. 3B); and between the right dAI and the left postcentral gyrus (Table 5, Fig. 4B) (p < 0.001, k > 6 voxels, p < 0.05 corrected for multiple comparisons using AlphaSim).
Compared with the HC group, the SD and NSD groups exhibited very different alterations in AI functional coupling, with lower FC in the SD patients but higher FC in the NSD patients. The SD group showed lower FC between the left vAI with the right fusiform gyrus (Table 2, Fig. 1C), between the right vAI with the left posterior cingulate cortex and right precuneus (Table 3, Fig. 2C), between the left dAI with the left inferior frontal gyrus (Table 4, Fig. 3C), and between the right dAI with the right fusiform gyrus and left postcentral gyrus (Table 5, Fig. 4C). The NSD group showed higher FC between the left vAI with the bilateral rectus gyri and right orbital inferior frontal gyrus (Table 2, Fig. 1D), between the right vAI with the bilateral orbital frontal gyri (Table 3, Fig. 2D), between the left dAI with the left fusiform gyrus and left inferior frontal gyrus (Table 4, Fig. 3D), and between the right dAI with the right fusiform gyrus (Table 5, Fig. 4D) (p < 0.001, k > 6 voxels, p < 0.05 corrected for multiple comparisons using AlphaSim).
There were no differences in FC between the bilateral PI and any brain regions among the SD, NSD, and HC groups.
Exploratory correlational analyses between FC values and clinical data in SD patients
Our study did not find a significant correlation between FC values and any clinical characteristics in the SD patients. Some correlations failed to survive the multiple comparison correction. In the SD group, FC values between the left vAI and right rectus gyrus were positively correlated with cognitive impairment factor scores on the HAMD17 (r = 0.35, p = 0.040, uncorrected), and FC values between the right vAI and right superior frontal gyrus were positively correlated with total HAMD17 scores and cognitive impairment factor scores (r = 0.42 and p = 0.015, r = 0.43 and p = 0.013, uncorrected).
Discussion
The present study applied resting-state FC analyses to explore functional synchronization between the insula and the whole brain in MDD patients with somatic symptoms. The results showed that these subjects had decreased FC between the vAI and dAI and right frontal regions (superior frontal and rectus gyri) and several limbic brain areas (precuneus, angular gyrus, fusiform gyrus, and middle cingulate cortex). Unfortunately, subsequent Pearson correlation analyses revealed that there were no significant correlations between averaged FC values and some clinical indices. The uncorrected results showed that FC values between the left vAI and right rectus gyrus positively correlated with the cognitive impairment factor scores on the HAMD17 in the SD patients, while FC values between the right vAI and right superior frontal gyrus were positively correlated with the total and cognitive impairment factor scores on the HAMD17 in the SD patients. Patients without somatic symptoms (NSD group) showed enhanced FC between the AI and bilateral orbitofrontal cortices, rectus gyri and fusiform gyri.
The insula can be roughly divided into anterior and posterior subregions based on different cell origins and functions [31, 55]. The PI receives interoceptive signals from the spinal cord and brainstem and participates in the processing of primary sensory information such as somatosensory and motor control. The AI is connected with several regions in the frontal cortex, including the anterior cingulate cortex, medial prefrontal cortex, amygdala, and ventral striatum; it is involved in the processes of emotion, motivation, and cognition [55, 56]. The posterior-to-anterior transmission of interoceptive signals and interactions with multiple other brain regions enables conscious emotional perception of interoceptive information [31]. Previous functional neuroimaging work demonstrated a cognition - emotion - interoception division of the insula [56], dividing the AI into the ventral and dorsal subregions. The vAI has a similar cell structure as the limbic cortex, suggesting that it is more involved in emotion and attention, while the dAI is connected to frontal areas that function in cognitive control [40, 57]. With regard to large-scale networks, the insula is an important component of the salience network (SN), which is responsible for switching between the central executive network (CEN; task activated) and the DMN (task-negative activated); this structure comprehensively evaluates the importance of internal and external stimuli to affect cognitive emotional behaviour [58]. According to a recent review [37], patients with depression have abnormalities in the structure, activation, and FC of the insula, which may help guide the treatment of depression.
Our results showed decreased FC between the vAI and dAI with right frontal regions (superior frontal gyrus and rectus gyrus) in the SD group. Similar findings have been reported in previous studies. Zhang et al. investigated resting-state FC based on ventral and dorsal AI seeds; they concluded that depressive patients had increased FC between the vAI and orbitofrontal cortex, and FC values were negatively correlated with somatic symptom severity. Moreover, these FC decreases could be reversed with electroconvulsive therapy [35]. Yao et al. found that insular activity and FC between the insula and prefrontal cortex were reduced in patients with abnormal pain empathy [59]. The intervention measures improved pain empathy and increased FC between the insula and prefrontal cortex [59], suggesting that the decrease in functional coupling between these structures may be related to abnormal somatic perception, which is consistent with our observation of lowered insula-prefrontal cortex FC in depressed patients with somatic symptoms. Kim et al. demonstrated that patients with complex regional pain syndrome showed decreased FC between the AI and the dorsomedial prefrontal cortex, inferior frontal gyrus and cingulate cortex [60]. The superior frontal gyrus plays an important role in self-awareness, emotion regulation, and cognitive processing [61, 62]. The rectus gyrus is a part of the medial prefrontal cortex that mediates sensory information integration by connecting the prefrontal cortex with the hypothalamus and brainstem [63]. It was reported that deep brain stimulation therapy targeting the rectus gyrus can effectively improve refractory depression [64]. In addition, the superior frontal gyrus is a key region of the CEN [65] that participates in cognitive control and decision-making [66]. The insula is an important component of the SN. Decreased FC between the AI and superior frontal gyrus in depressed patients may reflect abnormal functional regulation between the SN and CEN, thus contributing to somatic symptoms. The trends in the correlations between FC values between the left vAI-right rectus gyrus and right vAI-right superior frontal gyrus with cognitive factors in the present study suggest that abnormal coupling of the AI and frontal cortex might induce somatic symptoms by affecting the cognition of patients with MDD. Compared with the HC group, the NSD group showed enhanced FC between the AI and the bilateral orbitofrontal cortices, rectus gyri, and fusiform gyri. The opposite patterns of AI connectivity in the two depression subgroups indicated that functional synchronicity between the AI and the abovementioned brain regions may play important roles in the somatic symptoms of depression. We speculated that functional synchronization between the insula and superior frontal gyrus and the rectus was weakened, inducing abnormal somatosensory information processing that resulted in uncomfortable somatic symptoms.
We also found decreased FC between the AI (especially the ventral AI) and several limbic brain regions (precuneus, angular gyrus, fusiform gyrus and middle cingulate cortex). These results were consistent with the aforementioned different FC patterns of the vAI and dAI with other brain regions and confirm the close interactions between the vAI and the limbic system. Considering that the limbic system is closely related to emotion and attention, aberrant FC between the vAI and limbic cortex may influence somatic and visceral sensory processes, as well as autonomic regulation of the heart and gastrointestinal tract. Avery et al. reported decreased activity in the dorsal mid-insula cortex (dmIC) during a task requiring attention to visceral interoceptive sensations, as well as increased resting-state FC between the dmIC and limbic brain regions that was positively correlated with depression severity and somatic symptoms [67]. On the other hand, the precuneus, superior marginal gyrus, and angular gyrus are important components of the DMN that are involved in emotional cognition and executive functions such as motivation, memory, and self-referential activity. The insula is a core node of the SN. Decreased functional coupling between the AI and the abovementioned limbic areas probably indicates inadequate regulation between the SN and DMN [68], leading to the presentation of somatic symptoms in MDD.
There were no differences in FC between the bilateral PI and any other brain regions in the depressive patients with or without somatic symptoms and HCs. The PI is responsible for the preliminary processing of sensory information, and is a critical hub for central integration and processing of painful stimuli [69]. Studies have found that the PI was associated with interoceptive awareness and relieved patients from the sensory and affective burden of chronic pain. Moreover, disruption functional connectivity of right PI were found in MDD [69,70,71]. Our results suggest that the somatic symptoms of patients with MDD mainly involve abnormal cognitive regulation and not primary sensory information processing [72]. Regardless of the important role of the PI in the primary processing of interoceptive signals, fMRI results in previous studies have been inconsistent. Gu et al. claimed that activity in the bilateral AI but not the PI was aberrant during tasks assessing cognitive demand and pain stimulus valence. Importantly, these cognitive demand and stimulus valence tasks resulted in a significant interaction in the AI and regions of the frontoparietal network [73]. Conversely, decreased FC between both the AI and PI and the frontal and cingulate cortices was reported in patients with pain syndrome [60]. Such discrepancies may be due to different diseases and task paradigms. The obvious difference in FC of the AI and PI underscores the importance of researching insular subregions.
Limitations
There are some limitations that should be considered in the present study. First, we defined seeds with population-level but not individual-level atlases, which may have concealed actual individual variations in brain functioning. In addition, as a real-world study, medication was not restricted before admission to this study, and a number of patients were prescribed antidepressants. Although electroconvulsive therapy was reported to reverse abnormal insular FC [35], it is unclear whether medication influenced insular FC in the present study. Therefore, we used drugs as covariates to reduce the impact of drugs on result. Additionally, the correlation between FC values and clinical scale scores failed to survive after corrections for multiple comparisons. This may have been due to the small sample size, which limited the generalization of our findings. Finally, we used the FC method to calculate FC values of the insula to the whole brain. However, unlike the effective connectivity method, FC has no directionality [44]. Thus, future studies could utilize an effective connectivity approach in a larger sample size of medication-free patients to further explore causality as it relates to FC.
Conclusion
In summary, the present study revealed aberrant functional synchronization between the AI and frontal and limbic cortices. These changes indicate abnormal interactions between the three main large-scale brain functional networks (SN, DMN, and CEN) that might underlie somatic symptoms in SD by affecting the cognitive and emotional processing of interoceptive information.
Availability of data and materials
The datasets generated during the current study are not publicly available due to the subjects’ privacy, but are available from the corresponding author on reasonable request.
Abbreviations
- MDD:
-
Major depressive disorder
- SD:
-
Somatic depression
- NSD:
-
Non-somatic depression
- FC:
-
Functional Connectivity
- vAI:
-
Ventral anterior insula
- dAI:
-
Dorsal anterior insula
- PI:
-
Posterior insula
- dmIC:
-
dorsal mid-insula cortex
- HAMD17 :
-
17-item Hamilton Rating Scale for Depression
- DMN:
-
Default mode network
- SN:
-
Salience network
- YMRS:
-
Young Mania Rating Scale
- HCs:
-
Healthy controls
- DSM-IV-TR:
-
Statistical Manual of Mental Disorders, 4th Edition, Text Revision
- DPARSF:
-
Data Processing Assistant for Resting-State fMRI
- MNI:
-
Montreal Neurological Institute
- BOLD:
-
Blood oxygen-level dependent
- FWHM:
-
Full-width at half maximum
- ROIs:
-
Regions of interest
- REST:
-
Rs-fMRI data analysis toolkit
References
Otte C, Gold SM, Penninx BW, Pariante CM, Etkin A, Fava M, et al. Major depressive disorder. Nat Rev Dis Primers. 2016;2:16065. https://doi.org/10.1038/nrdp.2016.65.
APA. Diagnostic and statistical manual of mental disorders. 4th ed; 2000.
Silverstein B. Gender difference in the prevalence of clinical depression: the role played by depression associated with somatic symptoms. Am J Psychiatry. 1999;156(3):480–2.
Walker J, Burke K, Wanat M, Fisher R, Fielding J, Mulick A, et al. The prevalence of depression in general hospital inpatients: a systematic review and meta-analysis of interview-based studies. Psychol Med. 2018;48(14):2285–98. https://doi.org/10.1017/S0033291718000624.
Fang X, Zhang C, Wu Z, Peng D, Xia W, Xu J, et al. The association between somatic symptoms and suicidal ideation in Chinese first-episode major depressive disorder. J Affect Disord. 2019;245:17–21. https://doi.org/10.1016/j.jad.2018.10.110.
Novick D, Montgomery WS, Aguado J, Peng X, Brugnoli R, Haro JM. Which somatic symptoms are associated with an unfavorable course in Chinese patients with major depressive disorder? Asia Pac Psychiatry. 2015;7(4):427–35. https://doi.org/10.1111/appy.12189.
Zis P, Daskalaki A, Bountouni I, Sykioti P, Varrassi G, Paladini A. Depression and chronic pain in the elderly: links and management challenges. Clin Interv Aging. 2017;12:709–20. https://doi.org/10.2147/CIA.S113576.
Demyttenaere K, Reed C, Quail D, Bauer M, Dantchev N, Montejo AL, et al. Presence and predictors of pain in depression: results from the FINDER study. J Affect Disord. 2010;125(1–3):53–60. https://doi.org/10.1016/j.jad.2010.02.106.
Schneider E, Linden M, Weigmann H, Wagner T, Quail D, Hundemer HP, et al. Early reduction in painful physical symptoms is associated with improvements in long-term depression outcomes in patients treated with duloxetine. BMC Psychiatry. 2011;11:150. https://doi.org/10.1186/1471-244X-11-150.
Riemann D, Voderholzer U. Primary insomnia: a risk factor to develop depression? J Affect Disord. 2003;76(1–3):255–9. https://doi.org/10.1016/s0165-0327(02)00072-1.
Hertenstein E, Feige B, Gmeiner T, Kienzler C, Spiegelhalder K, Johann A, et al. Insomnia as a predictor of mental disorders: a systematic review and meta-analysis. Sleep Med Rev. 2019;43:96–105. https://doi.org/10.1016/j.smrv.2018.10.006.
Wang X, Cheng S, Xu H. Systematic review and meta-analysis of the relationship between sleep disorders and suicidal behaviour in patients with depression. BMC Psychiatry. 2019;19(1):303. https://doi.org/10.1186/s12888-019-2302-5.
Harris LM, Huang X, Linthicum KP, Bryen CP, Ribeiro JD. Sleep disturbances as risk factors for suicidal thoughts and behaviours: a meta-analysis of longitudinal studies. Sci Rep. 2020;10(1):13888. https://doi.org/10.1038/s41598-020-70866-6.
Wilson N, Lee JJ, Bei B. Postpartum fatigue and depression: a systematic review and meta-analysis. J Affect Disord. 2019;246:224–33. https://doi.org/10.1016/j.jad.2018.12.032.
Lee CH, Giuliani F. The role of inflammation in depression and fatigue. Front Immunol. 2019;10:1696. https://doi.org/10.3389/fimmu.2019.01696.
Geng J, Yan R, Shi J, Chen Y, Mo Z, Shao J, et al. Altered regional homogeneity in patients with somatic depression: a resting-state fMRI study. J Affect Disord. 2019;246:498–505. https://doi.org/10.1016/j.jad.2018.12.066.
Yao Z, Yan R, Wei M, Tang H, Qin J, Lu Q. Gender differences in brain activity and the relationship between brain activity and differences in prevalence rates between male and female major depressive disorder patients: a resting-state fMRI study. Clin Neurophysiol. 2014;125(11):2232–9. https://doi.org/10.1016/j.clinph.2014.03.006.
Yan R, Tao S, Liu H, Chen Y, Shi J, Yang Y, et al. Abnormal alterations of regional spontaneous neuronal activity in inferior frontal orbital gyrus and corresponding brain circuit alterations: a resting-state fMRI study in somatic depression. Front Psychiatry. 2019;10:267. https://doi.org/10.3389/fpsyt.2019.00267.
Finnerup NB, Kuner R, Jensen TS. Neuropathic pain: from mechanisms to treatment. Physiol Rev. 2021;101(1):259–301. https://doi.org/10.1152/physrev.00045.2019.
Smith ML, Asada N, Malenka RC. Anterior cingulate inputs to nucleus accumbens control the social transfer of pain and analgesia. Science. 2021;371(6525):153–9. https://doi.org/10.1126/science.abe3040.
Liu MG, Zhuo M. Loss of long-term depression in the insular cortex after tail amputation in adult mice. Mol Pain. 2014;10:1. https://doi.org/10.1186/1744-8069-10-1.
Legrain V, Iannetti GD, Plaghki L, Mouraux A. The pain matrix reloaded: a salience detection system for the body. Prog Neurobiol. 2011;93(1):111–24. https://doi.org/10.1016/j.pneurobio.2010.10.005.
Bagherzadeh-Azbari S, Khazaie H, Zarei M, Spiegelhalder K, Walter M, Leerssen J, et al. Neuroimaging insights into the link between depression and insomnia: a systematic review. J Affect Disord. 2019;258:133–43. https://doi.org/10.1016/j.jad.2019.07.089.
Gong L, Yu S, Xu R, Liu D, Dai X, Wang Z, et al. The abnormal reward network associated with insomnia severity and depression in chronic insomnia disorder. Brain Imaging Behav. 2021;15(2):1033–42. https://doi.org/10.1007/s11682-020-00310-w.
Saltiel PF, Silvershein DI. Major depressive disorder: mechanism-based prescribing for personalized medicine. Neuropsychiatr Dis Treat. 2015;11:875–88. https://doi.org/10.2147/NDT.S73261.
Tang LW, Zheng H, Chen L, Zhou SY, Huang WJ, Li Y, et al. Gray matter volumes in patients with chronic fatigue syndrome. Evid Based Complement Alternat Med. 2015;2015:380615. https://doi.org/10.1155/2015/380615.
Doan L, Manders T, Wang J. Neuroplasticity underlying the comorbidity of pain and depression. Neural Plast. 2015;2015:504691. https://doi.org/10.1155/2015/504691.
Wang VC, Mullally WJ. Pain neurology. Am J Med. 2020;133(3):273–80. https://doi.org/10.1016/j.amjmed.2019.07.029.
IsHak WW, Wen RY, Naghdechi L, Vanle B, Dang J, Knosp M, et al. Pain and depression: a systematic review. Harv Rev Psychiatry. 2018;26(6):352–63. https://doi.org/10.1097/HRP.0000000000000198.
Harshaw C. Interoceptive dysfunction: toward an integrated framework for understanding somatic and affective disturbance in depression. Psychol Bull. 2015;141(2):311–63. https://doi.org/10.1037/a0038101.
Namkung H, Kim SH, Sawa A. The insula: an underestimated brain area in clinical neuroscience, psychiatry, and neurology. Trends Neurosci. 2017;40(4):200–7. https://doi.org/10.1016/j.tins.2017.02.002.
Motomura K, Terasawa Y, Natsume A, Iijima K, Chalise L, Sugiura J, et al. Anterior insular cortex stimulation and its effects on emotion recognition. Brain Struct Funct. 2019;224(6):2167–81. https://doi.org/10.1007/s00429-019-01895-9.
Palotai M, Cavallari M, Koubiyr I, Morales PA, Nazeri A, Healy BC, et al. Microstructural fronto-striatal and temporo-insular alterations are associated with fatigue in patients with multiple sclerosis independent of white matter lesion load and depression. Mult Scler. 2020;26(13):1708–18. https://doi.org/10.1177/1352458519869185.
Sprengelmeyer R, Steele JD, Mwangi B, Kumar P, Christmas D, Milders M, et al. The insular cortex and the neuroanatomy of major depression. J Affect Disord. 2011;133(1–2):120–7. https://doi.org/10.1016/j.jad.2011.04.004.
Zhang T, Bai T, Xie W, Wei Q, Lv H, Wang A, et al. Abnormal connectivity of anterior-insular subdivisions and relationship with somatic symptom in depressive patients. Brain Imaging Behav. 2021;15(4):1760–8. https://doi.org/10.1007/s11682-020-00371-x.
Nieuwenhuys R. The insular cortex: a review. Prog Brain Res. 2012;195:123–63. https://doi.org/10.1016/B978-0-444-53860-4.00007-6.
Gogolla N. The insular cortex. Curr Biol. 2017;27(12):R580–6. https://doi.org/10.1016/j.cub.2017.05.010.
Zucker NL, Kragel PA, Wagner HR, Keeling L, Mayer E, Wang J, et al. The clinical significance of posterior insular volume in adolescent anorexia nervosa. Psychosom Med. 2017;79(9):1025–35. https://doi.org/10.1097/PSY.0000000000000510.
Namkung H, Kim SH, Sawa A. The insula: an underestimated brain area in clinical neuroscience, psychiatry, and neurology: (trends in neuroscience 40, 200-207, 2017). Trends Neurosci. 2018;41(8):551–4. https://doi.org/10.1016/j.tins.2018.05.004.
Deen B, Pitskel NB, Pelphrey KA. Three systems of insular functional connectivity identified with cluster analysis. Cereb Cortex. 2011;21(7):1498–506. https://doi.org/10.1093/cercor/bhq186.
Zhang T, Bai T, Xie W, Wei Q, Lv H, Wang A, et al. Abnormal connectivity of anterior-insular subdivisions and relationship with somatic symptom in depressive patients. Brain Imaging Behav. 2020. https://doi.org/10.1007/s11682-020-00371-x.
Kandilarova S, Stoyanov D, Kostianev S, Specht K. Altered resting state effective connectivity of anterior insula in depression. Front Psychiatry. 2018;9:83. https://doi.org/10.3389/fpsyt.2018.00083.
Stoyanov D, Aryutova K, Kandilarova S, Paunova R, Arabadzhiev Z, Todeva-Radneva A, et al. Diagnostic task specific activations in functional MRI and aberrant connectivity of insula with middle frontal gyrus can inform the differential diagnosis of psychosis. Diagnostics (Basel). 2021;11(1). https://doi.org/10.3390/diagnostics11010095.
Todeva-Radneva A, Paunova R, Kandilarova S, St SD. The value of neuroimaging techniques in the translation and Transdiagnostic validation of psychiatric diagnoses - selective review. Curr Top Med Chem. 2020;20(7):540–53. https://doi.org/10.2174/1568026620666200131095328.
Manoliu A, Riedl V, Zherdin A, Mühlau M, Schwerthöffer D, Scherr M, et al. Aberrant dependence of default mode/central executive network interactions on anterior insular salience network activity in schizophrenia. Schizophr Bull. 2014;40(2):428–37. https://doi.org/10.1093/schbul/sbt037.
Friston KJ. Functional and effective connectivity: a review. Brain Connect. 2011;1(1):13–36. https://doi.org/10.1089/brain.2011.0008.
Greicius MD, Flores BH, Menon V, Glover GH, Solvason HB, Kenna H, et al. Resting-state functional connectivity in major depression: abnormally increased contributions from subgenual cingulate cortex and thalamus. Biol Psychiatry. 2007;62(5):429–37. https://doi.org/10.1016/j.biopsych.2006.09.020.
Hamilton M. Development of a rating scale for primary depressive illness. Br J Soc Clin Psychol. 1967;6(4):278–96.
Angst J, Adolfsson R, Benazzi F, Gamma A, Hantouche E, Meyer TD, et al. The HCL-32: towards a self-assessment tool for hypomanic symptoms in outpatients. J Affect Disord. 2005;88(2):217–33. https://doi.org/10.1016/j.jad.2005.05.011.
Young RC, Biggs JT, Ziegler VE, Meyer DA. A rating scale for mania: reliability, validity and sensitivity. Br J Psychiatry. 1978;133:429–35. https://doi.org/10.1192/bjp.133.5.429.
Silverstein B, Patel P. Poor response to antidepressant medication of patients with depression accompanied by somatic symptomatology in the STAR*D study. Psychiatry Res. 2011;187(1–2):121–4. https://doi.org/10.1016/j.psychres.2010.12.026.
Chao-Gan Y, Yu-Feng Z. DPARSF: a MATLAB toolbox for “pipeline” data analysis of resting-state fMRI. Front Syst Neurosci. 2010;4:13. https://doi.org/10.3389/fnsys.2010.00013.
Power JD, Barnes KA, Snyder AZ, Schlaggar BL, Petersen SE. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage. 2012;59(3):2142–54. https://doi.org/10.1016/j.neuroimage.2011.10.018.
Song XW, Dong ZY, Long XY, Li SF, Zuo XN, Zhu CZ, et al. REST: a toolkit for resting-state functional magnetic resonance imaging data processing. Plos One. 2011;6(9):e25031. https://doi.org/10.1371/journal.pone.0025031.
Shura RD, Hurley RA, Taber KH. Insular cortex: structural and functional neuroanatomy. J Neuropsychiatry Clin Neurosci. 2014;26(4):276–82. https://doi.org/10.1176/appi.neuropsych.260401.
Nomi JS, Farrant K, Damaraju E, Rachakonda S, Calhoun VD, Uddin LQ. Dynamic functional network connectivity reveals unique and overlapping profiles of insula subdivisions. Hum Brain Mapp. 2016;37(5):1770–87. https://doi.org/10.1002/hbm.23135.
Nomi JS, Schettini E, Broce I, Dick AS, Uddin LQ. Structural connections of functionally defined human insular subdivisions. Cereb Cortex. 2018;28(10):3445–56. https://doi.org/10.1093/cercor/bhx211.
Menon V, Uddin LQ. Saliency, switching, attention and control: a network model of insula function. Brain Struct Funct. 2010;214(5–6):655–67. https://doi.org/10.1007/s00429-010-0262-0.
Yao S, Becker B, Geng Y, Zhao Z, Xu X, Zhao W, et al. Voluntary control of anterior insula and its functional connections is feedback-independent and increases pain empathy. Neuroimage. 2016;130:230–40. https://doi.org/10.1016/j.neuroimage.2016.02.035.
Kim JH, Choi SH, Jang JH, Lee DH, Lee KJ, Lee WJ, et al. Impaired insula functional connectivity associated with persistent pain perception in patients with complex regional pain syndrome. Plos One. 2017;12(7):e180479. https://doi.org/10.1371/journal.pone.0180479.
Goldberg II, Harel M, Malach R. When the brain loses its self: prefrontal inactivation during sensorimotor processing. Neuron. 2006;50(2):329–39. https://doi.org/10.1016/j.neuron.2006.03.015.
Guo W, Liu F, Chen J, Gao K, Xue Z, Xu X, et al. Abnormal neural activity of brain regions in treatment-resistant and treatment-sensitive major depressive disorder: a resting-state fMRI study. J Psychiatr Res. 2012;46(10):1366–73. https://doi.org/10.1016/j.jpsychires.2012.07.003.
Ongur D, Price JL. The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cereb Cortex. 2000;10(3):206–19.
Accolla EA, Aust S, Merkl A, Schneider G, Kühn AA, Bajbouj M, et al. Deep brain stimulation of the posterior gyrus rectus region for treatment resistant depression. J Affect Disorders. 2016;194:33–7. https://doi.org/10.1016/j.jad.2016.01.022.
Habas C, Kamdar N, Nguyen D, Prater K, Beckmann CF, Menon V, et al. Distinct cerebellar contributions to intrinsic connectivity networks. J Neurosci. 2009;29(26):8586–94. https://doi.org/10.1523/JNEUROSCI.1868-09.2009.
Castellanos FX, Proal E. Large-scale brain systems in ADHD: beyond the prefrontal-striatal model. Trends Cogn Sci. 2012;16(1):17–26. https://doi.org/10.1016/j.tics.2011.11.007.
Avery JA, Drevets WC, Moseman SE, Bodurka J, Barcalow JC, Simmons WK. Major depressive disorder is associated with abnormal interoceptive activity and functional connectivity in the insula. Biol Psychiatry. 2014;76(3):258–66. https://doi.org/10.1016/j.biopsych.2013.11.027.
Su Q, Yao D, Jiang M, Liu F, Long L, Dai Y, et al. Decreased interhemispheric functional connectivity in insula and angular gyrus/supramarginal gyrus: significant findings in first-episode, drug-naive somatization disorder. Psychiatry Res Neuroimaging. 2016;248:48–54. https://doi.org/10.1016/j.pscychresns.2016.01.008.
Bergeron D, Obaid S, Fournier-Gosselin MP, Bouthillier A, Nguyen DK. Deep brain stimulation of the posterior insula in chronic pain: a theoretical framework. Brain Sci. 2021;11(5). https://doi.org/10.3390/brainsci11050639.
Kuehn E, Mueller K, Lohmann G, Schuetz-Bosbach S. Interoceptive awareness changes the posterior insula functional connectivity profile. Brain Struct Funct. 2016;221(3):1555–71. https://doi.org/10.1007/s00429-015-0989-8.
Wang W, Zhornitsky S, Li CS, Le TM, Joormann J, Li CR. Social anxiety, posterior insula activation, and autonomic response during self-initiated action in a Cyberball game. J Affect Disord. 2019;255:158–67. https://doi.org/10.1016/j.jad.2019.05.046.
Hu L, Xiao M, Ai M, Wang W, Chen J, Tan Z, et al. Disruption of resting-state functional connectivity of right posterior insula in adolescents and young adults with major depressive disorder. J Affect Disord. 2019;257:23–30. https://doi.org/10.1016/j.jad.2019.06.057.
Gu X, Liu X, Van Dam NT, Hof PR, Fan J. Cognition-emotion integration in the anterior insular cortex. Cereb Cortex. 2013;23(1):20–7. https://doi.org/10.1093/cercor/bhr367.
Acknowledgements
The authors thank all the participants for their time and efforts. We also thank the Xiaoqing Wang and Xiaoxiao Wu for their help in data collection.
Funding
This study was supported by grants of the National Natural Science Foundation of China (81871066); the Jiangsu Provincial key research and development program (BE2018609, BE2019675); the Jiangsu Provincial Medical Innovation Team of the Project of Invigorating Health Care through Science, Technology and Education (CXTDC2016004); Jiangsu Provincial Medical Youth Talent of the Project of Invigorating Health Care through Science, Technology and Education (QNRC2016050); the Key Project supported by Medical Science and Technology development Foundation, Jiangsu Commission of Health (K2019011); Humanities and social sciences research fund project of the ministry of education 17YJA740023.
Author information
Authors and Affiliations
Contributions
ZhiJian Yao, JiaBo Shi and Qing Lu designed the study and wrote the protocol, and revised the paper. Rui Yan and JiTing Geng managed the literature research, analyses and wroted the first draft of the manuscript. Rui Yan and JiTing Geng contributed equally to this work. YingHong Huang, HaoWen Zou, and Xumiao Wang collected experimental data, and prepared Table 1. Yi Xia and Shuai Zhao analyzed the results, and made the Tables 2, 3, 4 and 5. ZhiLu Chen and Yu Chen analyzed the results, and prepared Figs. 1, 2, 3 and 4. All authors reviewed the manuscript, and have approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Medical Ethics Committee of the Affiliated Brain Hospital of Nanjing Medical University (No.2011-KY026). All the research procedures involving human participants followed the Declaration of Helsinki in 1964 and its subsequent amendments. Written informed consent was obtained from all participants and a parent.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yan, R., Geng, J.T., Huang, Y.H. et al. Aberrant functional connectivity in insular subregions in somatic depression: a resting-state fMRI study. BMC Psychiatry 22, 146 (2022). https://doi.org/10.1186/s12888-022-03795-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12888-022-03795-5