
Abstract
Background
Chlorella vulgaris (ChV), a unicellular green algae has been reported to have anticancer and antioxidant effects. The aim of this study was to determine the chemopreventive effect of ChV on liver cancer induced rats by determining the level and expression of several liver tumour markers.
Methods
Male Wistar rats (200–250 g) were divided into 4 groups according to the diet given: control group (normal diet), ChV group with three different doses (50, 150 and 300 mg/kg body weight), liver cancer- induced group (choline deficient diet + 0.1% ethionine in drinking water or CDE group), and the treatment group (CDE group treated with three different doses of ChV). Rats were killed at 0, 4, 8 and 12 weeks of experiment and blood and tissue samples were taken from all groups for the determination of tumour markers expression alpha-fetoprotein (AFP), transforming growth factor-β (TGF-β), M2-pyruvate kinase (M2-PK) and specific antigen for oval cells (OV-6).
Results
Serum level of TGF-β increased significantly (p < 0.05) in CDE rats. However, ChV at all doses managed to decrease (p < 0.05) its levels to control values. Expressions of liver tumour markers AFP, TGF-β, M2-PK and OV-6 were significantly higher (p < 0.05) in tissues of CDE rats when compared to control showing an increased number of cancer cells during hepatocarcinogenesis. ChV at all doses reduced their expressions significantly (p < 0.05).
Conclusions
Chlorella vulgaris has chemopreventive effect by downregulating the expression of tumour markers M2-PK, OV-6, AFP and TGF-β, in HCC-induced rats.
Similar content being viewed by others
Background
Cancer formation is a complex process involving several stages, namely initiation, promotion and progression [1, 2]. These stages are described as a series of successive mutations that ultimately lead to malignant tumour growth [3]. Increasing evidences showed that accumulation of free radicals in the body causes a variety of biochemical and physiological abnormalities associated with cardiovascular disease, cancer and the aging process [4,5,6]. Mutation of tumour suppressor genes and activation of protooncogenes would transform normal cells into cancer cells which grow rapidly and metastasize to other parts of the body [7]. To preserve the integrity of an organism, the production of free radicals must be kept in balance by antioxidants. Antioxidants are enzymes [8] or non-enzymes mainly found in our diet that can either scavenge free radicals directly, or regulate its level [9,10,11,12].
Fortunately, the presence of most human cancers can be detected by tumour markers, which should have high specificity and few false positive for specific tumours. They must be undetectable in non-neoplastic conditions. Effective screening strategies using molecular markers for most types of cancers have saved or improved quality of life for many patients. Alpha-fetoprotein (AFP) is the most commonly used tumour marker for hepatocellular carcinoma (HCC) screening [13] whereby patients with a high AFP level indicates a bad prognosis than patients with lower AFP levels [14]. Sangiovani et al. (2001) [15] found that cirrhosis patients with elevated AFP level have higher risk of developing HCC. Increased level of AFP in the serum has also been reported to increase in chronic hepatitis B patients [16], fatty liver disease [17] and metabolic syndrome [18]. Although AFP is considered as the gold standard marker for HCC, it is however not useful in the early diagnosis of the disease, particularly AFP-negative HCC, suggesting that new biomarkers are needed [19]. Other liver tumour markers of interest are pyruvate kinase isoenzymes M2-PK, transforming growth factor – β (TGF-β), and specific antigen for oval cells, OV-6. M2-PK is highly expressed in oval cells, the precursor of liver tumour and it catalyzes the conversion of phosphoenolpyruvate to pyruvate [20]. When a normal cell transforms into a tumour cell, M2- PK expression is upregulated due to the uncommonly high anaerobic glycolysis [21], followed by its release into the blood fluid system, which enables it to be measured quantitatively [22]. TGF- β has been recently considered as a possible liver tumour marker. TGF-β1 and TGF-β1 mRNA were shown to be sensitive indicators in the diagnosis of HCC induced by HBV, with the sensitivity and specificity being 89.5 and 94.0%, respectively [23]. Oval cells were first discovered by Farber [24]. They are oval-shaped cells that are not present in the normal liver [25, 26], but their levels are increased during liver regeneration and early stage of HCC. The specific tumour marker for oval cells is OV-6 [27].
Chlorella vulgaris (ChV), a unicellular green algae, has long been used as a health supplement especially in Japan and Korea [28, 29]. ChV contains high content of nutrients including vitamins and minerals [30]. It has been shown to strengthen the immune system [31, 32], and exhibit anti- inflammatory effect [32, 33]. In addition, our laboratory has shown that ChV reduced cellular proliferation and induced apoptosis in hepatoma cell line, HepG-2 [34,35,36], and exhibited antioxidant property in hepatoma-induced rats [35] and STZ-induced diabetic rats [37, 38].
The main objective of this study is to evaluate the antitumour property of ChV in liver cancer- induced rats, by determining the level and expression of novel tumour marker AFP, and comparing with other possible markers M2-PK, OV-6 and TGF-β.
Methods
Chlorella vulgaris culture
C. vulgaris algae Beijerinck 072 obtained from UMACC (University of Malaya Algae Culture Collection) Malaysia was grown in Bold Basal Media in laboratory setting with 12 h of dark and light cycle, and harvested by centrifugation (3000 RPM, 10 min, three times at 4 °C). The pelleted algae were diluted in distilled water to arrive at concentrations of 50, 150 and 300 mg/kg body weight.
Animals, chemicals and treatment
A total of 144 male Wistar rats (200 to 250 g) were obtained from the Animal Unit, National University of Malaysia, and were lodged in polycarbonate cages in a room with controlled temperature, humidity, and light-dark cycle, housed in the animal house of the Institute for Medical Research (IMR), Malaysia. All experiments were conducted following the guidelines of National Institute of Health for the Care and Use of Laboratory Animals. The study was approved by the Animal Ethics Committee of the National University of Malaysia (Approval number: BIOK/2002/YASMIN/30-SEPTEMBER/082). All animals received adequate human care.
The rats were divided into four groups (6 rats per group): control group (normal diet), ChV group with three different doses (50, 150 and 300 mg/kg body weight), liver cancer- induced (choline deficient diet +0.1% ethionine in drinking water to induce cancer) or CDE group [31, 36, 39], and the treatment group or CDE group treated with three different doses of ChV.
The rats in the control group were given both normal diet and drinking water (normal rat chow) via ad libitum. The rats in the ChV group were administered with only ChV at three different doses (50, 150 and 300 mg/kg body weight), per day via oral gavage. The rats in CDE + ChV group were administered with CDE and ChV at 50, 150 and 300 mg/kg. The duration of the experiment was three months and the rats were sacrificed at 0, 4, 8, and 12 weeks. Animals were anesthetized for liver perfusion procedure prior to excision of the liver. Liver tissue was excised and fixed in formalin and embedded in paraffin for immunohistochemistry analysis. Blood was taken via orbital sinus prior to killing the rats at 0, 4, 8 and 12 weeks for determination of TGF-β.
Hepatic perfusion
Rats were intraperitoneally anesthetized with Zoletil 50 (0.1 ml/100 g body weight), followed by an injection of 0.2 ml of heparin (25,000 U / ml) into the inferior vena cava. The portal vein was canulated with an intravenous catheter needle (16 G IV catheter, 2:25 in.) for the perfusion. Liver was perfused with phosphate-buffered saline (PBS) for 1 min at a flow level of 10 ml/min, followed by perfusion with 4% paraformaldehyde and 0.1% glutaraldehyde (1:1) for three minutes at room temperature. The liver was then perfused again with PBS for two minutes, and rinsed with PBS. A portion of perfused liver was cut and fixed in 10% formalin followed by tissue processing and embedding in paraffin.
Determination of serum TGF-β
Blood was obtained via the orbital sinus and collected in a tube, to allow it to set for two hours before centrifugation (3000 RPM, 10 min, 4 °C). Serum obtained was stored at −80 °C. The levels of transforming growth factor-β (TGF-β) in the serum was determined by ELISA (BD Pharmingen, USA), according to the protocol by the manufacturer.
Immunohistochemistry staining for AFP, M2-PK, TGF-β and OV-6
Sequential tissue sections (3 μm) were mounted on poly-L-lysine-coated slides. Archival samples were dewaxed by gradual washings in xylene and then dehydrated in various concentrations of alcohols (100, 80, 60 and 40%). Slides then were incubated in 3% hydrogen peroxide in distilled water to quench endogenous peroxidase activity, after which slides were washed under running water. Antigen retrieval was performed by incubating slides in a preheated Colin jar containing Target Retrieval Solution (TRS) pH 9 (Dako, Glostrup, Denmark) for 20 min in a water bath with temperatures ranging from 95 °C to 99 °C. After thermal treatment, the slides were allowed to cool for 20 min at room temperature. Slides were then washed under running water for three minutes and were placed in Tris-buffered saline (TBS), pH 7.6.
Sections were then incubated for 35 min with primary monoclonal antibody: rabbit anti-human AFP (Dako, USA) at 1:200 dilution, or goat anti-human M2-PK (Biodesign, USA) at 1:600 dilution, or mouse anti-rat OV-6 (a gift from Dr. Stewart Sell, USA) at 1:400 dilution or mouse anti-human TGF-β at 1:200 dilution. Reaction products were incubated with horseradish peroxidase-conjugated secondary antibody. Diaminobenzidine (DAB) was used as a chromogenic substrate (LSAB = HRP kit, Dako) to visualize the antibody-antigen reaction. All sections were then counterstained with hematoxylin and mounted with permanent mountant DPX. Sections were visualized under light microscopy for assessment of immunoreactivity.
Human liver cancer known to be positive for AFP expression was included as positive control for AFP, cancer-induced liver tissues from previous experiment were used as positive control for both M2-PK and OV-6 immunostaining, and lesion-induced gaster tissue was used as positive control for TGF-β.
Immunoreactivity Assesment
A researcher with no knowledge of clinicopathologic data on the samples evaluated the slides in a blind fashion. Confirmation of the diagnosis was performed by a pathologist evaluating the same slides independently. Most of the slides were classified similarly by both investigators. Results were expressed in percentage of stained cells over total cells counted from ten different fields. A total of 100 stained or non-stained cells were counted from each field at a 40× objective [40].
Statistical analysis
Statistical analysis was performed with ANOVA using SPSS program ver.11.0. Results were represented as mean ± SEM with p < 0.05 considered as significant difference.
Results
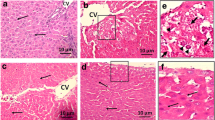
Figure 1b confirms the formation of oval cells as opposed in the control group (Fig. 1a), indicating the cellular changes expected in liver cancer rats. This result shows that rats treated with CDE is a good model for liver cancer rats.

Tissue sections showing rat liver tissues (× 400 magnification). a) Normal rat liver tissue. b) Formation of oval cells (as shown by arrows) in hepatoma-induced rats
Figure 2 clearly showed the hepatoprotective effect of ChV in reducing the elevated levels of TGF-β seen in serum of liver cancer rats (CDE). As seen from the figure, TGF-β level was significantly increased (p < 0.05) at all weeks following carcinogen (CDE) diet, compared to control. TGF-β level was significantly higher after eight and 12 weeks (p < 0.05), compared to four weeks. Treatment of the rats with ChV at all doses reduced TGF-β level at all weeks. Rats fed with ChV at 300 mg/kg body weight brought the level of TGF-β to almost control level. ChV alone did not raise the levels of TGF-β and seemed to be the same level as in the control, showing no toxicity effect to the liver.

Serum level of TGF-β in CDE rats treated with ChV. asignificant difference (p < 0.05) compared to control, bsignificant difference (p < 0.05) compared to CDE, csignificant difference (p < 0.05) compared to ChV50m dsignificant difference (p < 0.05) compared to ChV150, 1significant difference (p < 0.05) compared to Week 4
As can be seen from Fig. 3, liver cancer rats resulted in brown-stained cells (shown by arrows in the middle panel) indicating positive expressions of AFP (Fig. 3b), OV-6 (Fig. 3e), M2-PK (Fig. 3h) and TGF-β (Fig. 3k), as compared to rats fed with normal diet (Fig. 3a, d, g, and j). Liver cancer rats treated with 300 mg/kg ChV resulted in suppression of the proteins, as evidenced by the smaller amount of brown-stained cells for AFP (Fig. 3c), OV-6 (Fig. 3f), M2-PK (Fig. 3i) and TGF-β (Fig. 3l).

Immunohistochemical expressions of liver tumour markers in rats (× 400 magnification). (a) AFP in Control, (b) AFP in CDE, (c) AFP in CDE + ChV, (d) OV-6 in Control, (e) OV-6 in CDE, (f) OV-6 in CDE + ChV, (g) M2-PK in Control, (h) M2-PK in CDE, (i) M2-PK in CDE + ChV, (j) TGF-β in Control, (k) TGF-β in CDE and (l) TGF-β in CDE + ChV. Arrows indicate brown-stained positive cells. Boxed area in (b) contains oval cells
Figure 4a shows that the expression of AFP in liver tissues was significantly increased (p < 0.05) in the CDE (liver cancer) group for weeks 8 and 12, but treatment with all concentrations of ChV significantly (p < 0.05) reduced its expression. Greater reduction of AFP expression was seen at 300 mg/kg body weight (p < 0.05).

Quantitative expressions of (a) AFP, (b) OV-6, (c) M2-PK and (d) TGF-β in liver cancer tissues. asignificant difference (p < 0.05) compared to control, bsignificant difference (p < 0.05) compared to CDE, 1significant difference (p < 0.05) compared to Week 4, 2significant difference (p < 0.05) compared to Week 8
Similarly, OV-6 (Fig. 4b) was significantly (p < 0.05) expressed in CDE group, but showed no significant difference between 8 and 12 weeks of HCC induction. Treatment of the CDE group with 150 and 300 mg/kg body weight ChV significantly reduced (p < 0.05) the OV-6 expression for weeks 8 and 12. Treatment with 300 mg/kg body weight ChV for 12 weeks resulted in significant (p < 0.05) OV-6 suppression compared to week 4.
M2-PK (Fig. 4c) was significantly expressed (p < 0.05) in the CDE group, where its expression reached the highest point at 8 weeks, and reduced by more than 2% at 12 weeks. Treatment with 300 mg/kg body weight of ChV resulted in a significant reduction of M2-PK expression for all weeks of supplementation compared to CDE group. ChV at lower doses (50 and 150 mg/kg body weight) significantly (p < 0.05) reduced M2-PK expression for weeks 8 and 12.
TGF-β (Fig.4d) was also significantly expressed (p < 0.05) in increasing manner in the CDE group with time of exposure to the carcinognen. Its expression peaked at 12 weeks. Treatment with 150 and 300 mg/kg body weight ChV resulted in a significant reduction of TGF-β expression for all weeks of treatment. However a lower dose of ChV (50 mg/kg body weight) significantly reduced TGF-β expression for weeks 8 and 12 only.
Interestingly the changes of serum level of TGF-β is in concordance with its expressions in tissues (figs. 2 and 4) when treated with ChV. We have also documented the same changes in serum level of AFP in a previous report [41].
Discussion
Hepatocellularcarcinoma (HCC) affects approximately one million individuals annually worldwide, making it one of the world’s most lethal cancer [42, 43], and the most common form in adults [43]. In 2016, the American Cancer Society estimated there would be over 39,000 of new cases of primary liver cancer and intrahepatic bile duct cancer in the United States alone. Out of this staggering figure, over 27,000 people are estimated to die from these types of cancer [43]. Worldwide, the most common primary liver cancer that occur is HCC, which accounts for 70% to 90% of cases [43].
Thus it is crucial to diagnose HCC early to improve the survival rate of the patients afflicted with the disease. AFP is a the gold standard tumour marker for HCC and its expression is upregulated during hepatocarcinogenesis, hence the use of AFP as a standard biomarker of liver cancer screening [13, 44]. However the specificity and sensitivity of AFP used in liver cancer screening are not satisfactory [45, 46], although it is useful in HCC surveillance in patients with cirrhosis [47]. The outcome of this research indicate the potentials of OV-6, M2-PK and TGF-β as liver tumour biomarkers besides AFP, while it attests the beneficial effect of ChV extract as an anti-cancer treatment.
OV-6 and M2-PK have been previously shown to over-express in oval cells following HCC induction by choline-deficient + ethionine (CDE) diet [26, 48, 49]. Oval cells play an important role in the development of HCC [50], where their presence is thought to be one of the first cellular changes in hepatic neoplasia, following exposure of the tissue to chemical carcinogens such as ethionine [39, 51]. Oval cells in hepatocytes are transient in nature whereby when triggered by toxic compounds or insults they will proliferate. However normal hepatocytes do not express oval cells as seen in this study and others [52, 53].
Our earlier study has shown the development of liver cancer nodules [36] and increased serum AFP in rats fed with CDE diet [41]. Here we showed that AFP is detected in the hepatocytes of liver cancer rats. High level of AFP has been found in 60–70% of patients with HCC and is associated with poor prognostic and survival rates in untreated patients [54]. AFP also correlates closely with the growth rate (number of dividing cells) and size of the tumour [55] as well as progressive elevation of alpha fetoprotein in biopsied liver samples of patients with liver cirrhosis and hepatocellular carcinoma [56]. Its re-expression in patients with HCC suggests abnormal or altered liver cell regeneration, or dedifferentiation of hepatocytes into tumour cells [57].
Lowes et al. [58] have documented three types of oval cell population; 1) primitive oval cells that are not expressing AFP, OV-6, CK-19 and π-GST, 2) oval-shaped cells resembling hepatocytes and express AFP but not OV-6, CK-19 and π-GST and 3) oval-shaped cells that resemble duct cells expressing OV-6, CK-19 and π-GST but not AFP. However, in this study, the OV-6 was expressed in the cytoplasm of oval-shaped cells resembling hepatocytes and bile duct cells. These results are supported by studies stating that oval cells are multipotent, where they are able to differentiate into hepatocytes [59] and bile ducts [60]. The increased number of oval cells in CDE rats was depicted by the increase in the expression of OV-6, which is consistent with other studies that observed the increasing number of oval cells is directly proportional to the severity of the disease [25, 58, 61]. Expression of OV-6, which is a specific marker for the presence of oval cells, has been reported in the early stages of liver regeneration in human and animals, either due to injury or inhibition of hepatocyte replication [52, 62]. These cells play an important role in the development of HCC [27]. The presence of oval cell population is deemed to be the first cellular changes that occur in neoplastic liver, following intrusion of toxic substances into the liver, in particular carcinogens such as ethionine [58, 61, 63].
In rats supplemented with the carcinogen N-nitrosomospholine (NNM) and CDE, M2-PK expression can be observed in the cytoplasm of oval cells found in the liver tissue [64,65,66]. However, not all oval cells are M2-PK-positive. The outcome depends on the fate of the oval cells; either to differentiate into a hepatocyte or a bile duct cell [59, 60]. In this study, the level of M2-PK expression was elevated in liver cancer-induced rats as compared to control, plausibly due to the increase in respiratory rate [21], following the transformation of the cell into a cancerous state [20, 64]. However M2-PK may not be selective biomarker for liver cancer since its level in the blood has been observed to be raised in other types of cancer such as lung [67], breast [68], cervical [69], esophagus and gastro [21] cancers.
Antioxidants, especially those derived from plant sources are reported to prevent carcinogenesis through the suppression of cell proliferation, stimulation of apoptosis and scavenging free radicals [28]. Our previous study has successfully shown that ChV (300 mg/kg body weight) significantly reduced the percentage of CDE-induced preneoplastic liver nodules (ranging in size from 0.1 to 0.5 cm) from 100% to 17% [36].
ChV has anti-proliferative as well as apoptosis effects against HepG2 liver cancer cells [34, 36, 70]. In addition, Sulaiman et al. [35] showed that ChV treatment resulted in the decline of superoxide dismutase and catalase activity levels, increased level of glutathione peroxidase activity, and the reduction of malondialdehyde in rats supplemented with CDE diet [35]. They suggested that ChV exerted its chemopreventive effect by replacing or compensating for endogenous antioxidant enzyme activity and inhibiting lipid peroxidation. The study also pointed out that free radicals generated by carcinogenesis were scavenged by ChV, which led to the reduction of oxidative stress, thus reducing the formation of cancer cells.
The role of ChV as anticancer agents can be seen clearly in this study. Based on immunohistochemistry results, AFP, OV-6, M2-PK and TGF-β were undetectable in the liver tissue of rats supplemented with ChV, thus implying its non-toxic nature. Interestingly, our study showed that treatment of CDE rats with 300 mg/kg body weight ChV as early as four weeks was adequate to suppress AFP expression, while the lowest dose (50 mg/kg body weight) managed to suppress AFP expression after prolonged supplementation for 12 weeks. The efficacy of ChV is also observed with OV-6 expression where prolonged supplementation with ChV for 12 weeks suppressed its expression. ChV was also able to reduce the expression of M2-PK, as was also seen with AFP and TGF-β in liver cancer induced rats. The actual mechanism of ChV as an antioxidant and anticancer agent has yet to be elucidated. This study documented the reduction of oval cells formation by ChV supplementation in the liver cancer-induced rats. ChV contains a variety of antioxidants such as ascorbic acid, tocopherols and reduced glutathione [71], and has the potential as an anticancer agent which can restrict and suppress the growth of initiated clonal cell populations into foci or preneoplastic nodules and HCC [72, 73, 74].
Conclusions
This study documented the reduction of oval cells formation as well as the expression of tumour markers M2-PK, OV-6, AFP and TGF-β in HCC induced rats supplemented with Chlorella vulgaris (ChV). Based on this study as well as from our previous studies [36, 38, 41], we postulate that the chemopreventive mechanism of ChV, which is rich in antioxidants, is by scavenging ROS found high in tumour cells, as well as inducing antiproliferative and apoptosis effect resulting in the reduction of neoplastic nodules as reflected by the reduction in tumour markers M2-PK, OV-6, AFP and TGF-β.
Abbreviations
- π-GST:
-
π-glutathione
- ANOVA:
-
Analysis of variance
- CK-19:
-
Cytokeratin-19
- ELISA:
-
Enzyme-linked immunosorbent assay
- Min:
-
Minute
- RPM:
-
Revolutions per minute
- SEM:
-
Standard error of mean
References
Dapito DH, Mencin A, Gwak G-Y, et al. Promotion of hepatocellular carcinoma by the intestinal microbiota and TLR4. Cancer Cell. 2012;21(4):504–16.
Marquardt JU, Seo D, Andersen JB, et al. Sequential transcriptome analysis of human liver cancer indicates late stage acquisition of malignant traits. J Hepatol. 2014;60(2):346–53.
Ross JB, Huh D, Noble LB, Tavazoie SF. Identification of molecular determinants of primary and metastatic tumour re-initiation in breast cancer. Nat Cell Biol. 2015;17(5):651–64.
Madole MB, Bachewar NP, Aiyar CM. Study of oxidants and antioxidants in patients of acute myocardial infarction. Adv Biomed Res. 2015;4:241–50.
Grigorescu R, Gruia MI, Nacea V, Nitu C, Negoita V, et al. The evaluation of non-enzymatic antioxidants effects in limiting tumor- associated oxidative stress, in a tumor rat model. J Med Life. 2015;8(4):513–6.
Diolez P, Bourdel-Marchasson I, Calmettes G, et al. Hypothesis on skeletal muscle aging: mitochondrial adenine nucleotide translocator decreases reactive oxygen species production while preserving coupling efficiency. Front Physiol. 2015;6:1–9.
Orozco-Morales M, Sánchez-García FJ, Golán-Cancela I, et al. RB mutation and RAS overexpression induce resistance to NK cell-mediated cytotoxicity in glioma cells. Cancer Cell Int. 2015;15(1):57.
Russo M, Cocco S, Secondo A, et al. Cigarette smoke condensate causes a decrease of the gene expression of cu-Zn superoxide dismutase, Mn superoxide dismutase, glutathione peroxidase, catalase, and free radical-induced cell injury in SH-SY5Y human neuroblastoma cells. Neurotox Res. 2011;19(1):49–54.
Packer L, Smith JR. Extension of the lifespan of cultured normal human diploid cells by vitamin E. PNAS. 1974;71(12):4763–7.
Ting Y, Stocker R, Dawes IW. The lipophilic antioxidants α -tocopherol and coenzyme Q10 reduce the replicative lifespan of Saccharomyces cerevisiae. Free Radic Biol Med. 2010;49(2):237–44.
Fang Y-Z, Yang S, Free WG. Radicals, antioxidants, and nutrition. Nutrition. 2002;18(10):872–9.
Kim J, Park E, Ha H, Jo C, Lee W, et al. Resveratrol-loaded Nanoparticles Induce Antioxidant Activity against Oxidative Stress. Asian Australas. J. Anim. Sci. 2016;29:288–98.
Yi X, Yu S, Bao Y. Alpha-fetoprotein-L3 in hepatocellular carcinoma: a meta-analysis. Clin Chim Acta. 2013;425:212–20.
Li D, Mallory T, Satomura S. AFP-L3: a new generation of tumor marker for hepatocellular carcinoma. Clin Chim Acta. 2001;313(1–2):15–9.
Sangiovanni A, Colombo E, Radaelli F, et al. Hepatocyte proliferation and risk of hepatocellular carcinoma in cirrhotic patients. Am J Gastroenterol. 2001;96(5):1575–80.
Liu Y, Lin B, Zeng D, et al. Alpha-fetoprotein level as a biomarker of liver fibrosis status : a cross-sectional study of 619 consecutive patients with chronic hepatitis B. BMC Gastroenterol. 2014;14:145–52.
Xu P, Xu C, Wan X, et al. Association between serum alpha-fetoprotein levels and fatty liver disease : a cross-sectional study. World J Gastroenterol. 2014;20(33):11865–70.
Chen Y, Zhao Y, Feng L, Zhang J, Zhang J, Feng G. Association between alpha-fetoprotein and metabolic syndrome in a Chinese asymptomatic population : a cross-sectional study. Lipids Health Dis. 2016;15:85–93. 1–9
Yousuf F, Ding W, Richardson P, Palmer D, Patanwala I, Cross T. PWE-135 does Afp predict survival in patients with hepatocellular carcinoma (hcc)? Gut. 2014;63(Suppl 1):A184. https://doi.org/10.1136/gutjnl-2014-307263.395.
Mazurek S, Drexler HC. A, Troppmair J, Eigenbrodt E, Rapp UR. Regulation of pyruvate kinase type M2 by a-Raf: a possible glycolytic stop or go mechanism. Anticancer Res. 2007;27(6B):3963–71.
Koss K, Harrison RF, Gregory J, Darnton SJ, Anderson MR, Jankowski JAZ. The metabolic marker tumour pyruvate kinase type M2 (tumour M2-PK) shows increased expression along the metaplasia-dysplasia-adenocarcinoma sequence in Barrett’s oesophagus. J Clin Pathol. 2004;57(11):1156–9.
Tonus C, Neupert G, Witzel K. The faecal tumour M2-PK screening test for invasive & pre-invasive colorectal cancer: estimated specificity & results as a function of age for a study population of 4854 volunteers. NOWOTWORY Journal of Oncology. 2009;59(2):32–7.
Dong ZZ, Yao DF, Yao M, et al. Clinical impact of plasma TGF-beta1 and circulating TGF-beta1 mRNA in diagnosis of hepatocellular carcinoma. Hepatobiliary Pancreat Dis Int. 2008;7:288–95.
Farber E. Ethionine Carcinogenesis. Adv Cancer Res. 1963;7:383–474.
Lin C-H, Liu C-H, Tsai H-L, Wang J-Y, Tsai H-P, Chai C-Y. Expression of OV-6 in primary colorectal cancer and rectal cancer with preoperative chemoradiotherapy: a clinicopathological study. Histopathology. 2013;2:742–51.
Oishi N, Wang XW. Novel therapeutic strategies for targeting liver cancer stem cells. Int J Biol Sci. 2011;7:517–35.
Yang W, Yan H-Y, Chen L, et al. Wnt/β-catenin signaling contributes to activation of normal and tumorigenic liver progenitor cells. Cancer Res. 2008;68:4287–95.
Lee SH, Kang HJ, Lee H-J, Kang M-H, Park YK. Six-week supplementation with Chlorella has favorable impact on antioxidant status in Korean male smokers. Nutrition. 2010;26(2):175–83.
Safi C, Zebib B, Merah O, Pontalier P-Y, Vaca-Garcia C. Morphology, composition, production, processing and applications of Chlorella vulgaris: a review. Renew Sust Energ Rev. 2014;35:265–78.
Yusof YAM, Basari JMH, Mukti NA, Sabuddin R, Muda AR, Sulaiman S, Makpol S, Ngah WZW. Fatty acids composition of microalgae Chlorella vulgaris can be modulated by varying carbon dioxide concentration in outdoor culture. African J of Biotech. 2013;10(62):13536-13542.
Hasegawa T, Yoshikai Y, Okuda M, Nomoto K. Accelerated restoration of the leukocyte number and augmented resistance against Eschericia coli in cyclophosphamide-treated rats orally administered with a hot water extract of Chlorella vulgaris. Int J Immunopharmacol. 1990;12(8):883–91.
Hasegawa T, Kimura Y, Hiromatsu K, et al. Effect of hot water extract of Chlorella vulgaris on cytokine expression patterns in mice with murine acquired immunodeficiency syndrome after infection with Listeria monocytogenes. Immunopharmacology. 1997;35(3):273–82.
Park J-Y, Cho H-Y, Kim J-K, et al. Chlorella dichloromethane extract ameliorates NO production and iNOS expression through the down-regulation of NF kappa B activity mediated by suppressed oxidative stress in RAW 264.7 macrophages. Clin Chim Acta. 2005;351(1–2):185–96.
Saad SM, Yusof YAM, Zurinah W, Ngah W. Comparison between locally produced Chlorella vulgaris and Chlorella vulgaris from Japan on proliferation and apoptosis of liver cancer cell line, HepG2. Malaysian Journal of Biochemistry and Molecular Biology. 2006;13:32–6.
Sulaiman S, Shamaan NA, Ngah WZWN, Yusof YAM. Chemopreventive effect of Chlorella vulgaris in choline deficient diet and ethionine induced liver carcinogenesis in rats. Int J Cancer Res. 2006;2(3):234–41.
Azamai ESM, Sulaiman S, Habib SHM, Lee LM, Das S, Hamid NAA, Ngah WZW, Yusof YAM. Chlorella vulgaris triggers apoptosis in hepatocarcinogenesis-induced rats. J Zhejiang Univ Sci B. 2009;10:14–21.
Aizzat O, Yap SW, Sopiah H, Madiha MM, Hazreen M, et al. Modulation of oxidative stress by Chlorella vulgaris in streptozotocin (STZ) induced diabetic Sprague-Dawley rats. Adv Med Sci. 2010;55:281–8.
Aliahmat NS, Noor MRM, Yusof WJW, Makpol S, Ngah WZW, Yusof YAM. Antioxidant enzyme activity and malondialdehyde levels can be modulated by Piper betle, tocotrienol rich fraction and Chlorella vulgaris in aging C57BL/6 mice. Clinics. 2012;67:1447–54.
Akhurst B, Croager EJ, Farley-Roche CA, et al. A modified choline-deficient, ethionine-supplemented diet protocol effectively induces oval cells in mouse liver. Hepatology. 2001;34(3):519–22.
Magalhães GM, Terra EM, Vasconcelos RDO, Bandarra MDB, et al. Immunodetection of cells with a CD44+/CD24- phenotype in canine mammary neoplasms. BMC Vet Res. 2013;9:205.
Yusof YAM, Basri JMH, Mokti NA, Saad SM, Sulaiman, S, Yahaya NS, Azamai ESM, Makpol S. Chlorella vulgaris: A miracle plant?. Modeling, Simulation and Applied Optimization (ICMSAO), 2011 4th International Conference on. 2011. pp. 1–6.
Zhao Y, Ju Q, Li G. Tumor markers for hepatocellular carcinoma. Mol. Clin Oncol. 2013;1:593–8.
American Cancer Society, “Cancer Facts & Figures 2016,” Cancer Facts Fig. 2016. 2016. pp. 1–9.
Zhang K, Song P, Gao J, Li G, Zhao X, Zhang S. Perspectives on a combined test of multi serum biomarkers in China: towards screening for and diagnosing hepatocellular carcinoma at an earlier stage. Drug Discov Ther. 2014;8:102–9.
Roberts LR. HCC in focus biomarkers for hepatocellular carcinoma. Gastroenterology & Hepatology. 2016;12:252–5.
Jang ES, Jeong S, Kim J, Choi YS, Leissner P, Brechot X. Diagnostic performance of alpha- fetoprotein, protein induced by vitamin K absence, osteopontin, Dickkopf-1 and its combinations for hepatocellular carcinoma. PLoS One. 2016;11(3):e0151069.
Chang TS, Wu Y, Tung S, Wei K, Hsieh Y, et al. Alpha-fetoprotein measurement benefits hepatocellular carcinoma surveillance in patients with cirrhosis. Am J Gastroenterol. 2015;110:836–44.
Lowes KN, Brennan BA, Yeoh GC, Olynyk JK. Oval cell numbers in human chronic liver diseases are directly related to disease severity. Am J Pathol. 1999;154:537–41.
Yang W, Wang C, Lin Y, Liu Q, et al. OV6+ tumor-initiating cells contribute to tumor progression and invasion in human hepatocellular carcinoma. J Hepatol. 2012;57:613–20.
Fang C, Gong J, Zhang W. Function of oval cells in hepatocellular carcinoma in rats. World J Gastroenterol. 2004;10:2482–7.
Dumble ML, Croager E, Yeoh GCT, Quail EA. Generation and characterization of p53 null transformed hepatic progenitor cells: oval cells give rise to hepatocellular carcinoma. Carcinogenesis. 2002;23:435–45.
Ma X, Qiu DK, Peng YS. Immunohistochemical study of hepatic oval cells in human chronic viral hepatitis. World J Gastroenterol. 2001;7:238–42.
Xiao J, Jin X, Ruck P, Adam A, Kaiserling E. Hepatic progenitor cells in human liver cirrhosis: immunohistochemical, electron microscopic and immunofluorencence confocal microscopic findings. World J Gastroenterol. 2004;10:1208–11.
Tsuji M, Kashihara T, Terada N, Mori H. An immunohistochemical study of hepatic atypical adenomatous hyperplasia, hepatocellular carcinoma, and cholangiocarcinoma with alpha-fetoprotein, carcinoembryonic antigen, CA19-9, epithelial membrane antigen, and cytokeratins 18 and 19. Pathol Int. 1999;49:310–7.
Sato Y, Nakata K, Shima M, Ishii N, Koji T, Taketa K, et al. Early recognition of hepatocellular carcinoma based on altered profiles of alpha-fetoprotein. N Engl J Med. 1993;328:1802–6.
Arrieta O, Cacho B, Morales-espinosa D, Ruelas-villavicencio A, Flores-estrada D, Hernández- N. The progressive elevation of alpha fetoprotein for the diagnosis of hepatocellular carcinoma in patients with liver cirrhosis. BMC Cancer. 2007;7:28–36.
Sell S. Alpha-fetoprotein (AFP), stem cells, and cancer: how study of the production of AFP during chemical hepatocarcinogenesis led to reaffirmation of the stem cell theory of cancer. Tumour Biol. 2008;29(3):161–80.
Lowes KN, Croager EJ, Olynyk JK, Abraham LJ, Yeoh GCT. Oval cell-mediated liver regeneration: role of cytokines and growth factors. J Gastroenterol Hepatol. 2003;18:4–12.
Kuijk EW, Rasmussen S, Blokzijl F, Huch M, Gehart H, et al. Generation and characterization of rat liver stem cell lines and their engraftment in a rat model of liver failure. Sci Rep. 2016;6:22154.
Chen Y, Chen H, Chien C, Wu S, Ho Y, et al. Contribution of mature hepatocytes to biliary regeneration in rats with acute and chronic biliary injury. PLoS One. 2015;10:1–17.
Roskams T, Yang S, Koteish A, Durnez A, DeVos R, et al. Oxidative stress and oval cell accumulation in mice and humans with alcoholic and nonalcoholic fatty liver disease. Am J Pathol. 2003;163:1301–11.
Roskams T, De Vos R, Van Eyken P, Myazaki H, et al. Hepatic OV-6 expression in human liver disease and rat experiments: evidence for hepatic progenitor cells in man. J Hepatol. 1998;29:455–63.
Wang P, Cong M, Liu T, Yang A, Sun G, Zhang D et al. The characteristics variation of hepatic progenitors after TGF-β1-induced transition and EGF-induced reversion. Stem Cells International. 2016;2016:10. Article ID 6304385. https://doi.org/10.1155/2016/6304385.
Hacker HJ, Steinberg P, Bannasch P. Pyruvate kinase isoenzyme shift from L-type to M2 -type is a late event in hepatocarcinogenesis induced in rats by a choline- deficient/DL-ethionine-supplemented diet. Carcinogenesis. 1998;19:99–107.
Steinberg P, Klingelhöffer A, Schäfer A, Wüst G, Weisse G, Oesch F, Eigenbrodt E. Expression of pyruvate kinase M2 in preneoplastic hepatic foci of N-nitrosomorpholine-treated rats. Virchows Arch. 1999;434:99–107.
Croager EJ, Smith PG, Yeoh GCT. Ethanol interactions with a choline-deficient, ethionine- supplemented feeding regime potentiate pre-neoplastic cellular alterations in rat liver. Carcinogenesis. 2002;23:1685–93.
Schneider J, Neu K, Velcovsky HG, Morr H, Eigenbrodt E. Tumor M2-pyruvate kinase in the follow-up of inoperable lung cancer patients: a pilot study. Cancer Lett. 2003;193:91–8.
Luftner D, Mesterham J, Akrivakis C, Geppert R, Petrides PE, et al. Tumor type M2 pyruvate kinase expression in advanced breast cancer. Anticancer Res. 2000;6D:5077–82.
Landt S, Jeschke S, Koeninger A, Thomas A, Heusner T, et al. Tumor-specific correlation of tumor M2 pyruvate kinase in pre-invasive, invasive and recurrent cervical cancer. Anticancer Res. 2010;30:375–81.
Yusof YAM, Saad SM, Makpol S, Shamaan NA, Ngah WZW. Hot water extract of Chlorella vulgaris induced DNA damage and apoptosis. Clinics. 2010;65:1371–7.
Kumar RR, Rao PH, Subramanian VV, Sivasubramanian V. Enzymatic and non-enzymatic antioxidant potentials of Chlorella vulgaris grown in effluent of a confectionery industry. J Food Sci Technol. 2014;51:322–8.
Morimoto T, Nagatsu A, Murakami N, Sakakibara J, Tokuda H, Nishino H, Iwashima A. Anti-tumour-promoting glyceroglycolipids from the green alga, Chlorella vulgaris. Phytochemistry. 1005;40:1433–7.
Abdel-Baky HH, Gouda EM, El-Behairy AN. El-BarotyGS. Chemoprevention of penzo[a]pyrene-induced carcinogen and lipid peroxidation in mice by lipophilic algae extracts (phycotene). J Med Sci. 2002;2:185–93.
Tsuda H, Sata M, Ijuuin H, Kumabe T, Uchida M, et al. A novel strategy for remission induction and maintenance in cancer therapy. Oncol Rep. 2002;9:65–8.
Funding
This study was funded by the research and Development Grant from the Malaysian Ministry of Science, Technology and Innovation (IRPA 06–02–02-0023-PR0008/09–08).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
Study conception, design and supervision of experimental workflow: YAMY and WZWN. Acquisition, analysis and interpretation of data: SS and SMS. Drafting of manuscript: KTA. Technical assistance in drafting of manuscript: HAD. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
All experiments were conducted following the guidelines of National Institute of Health for the Care and Use of Laboratory Animals. The study was approved by the Animal Ethics Committee of the National University of Malaysia (Approval number: BIOK/2002/YASMIN/30-SEPTEMBER/082). All animals received adequate human care.
Consent for publication
Not applicable.
Competing interests
The authors declare that there is no competing of interest in our part in the publication of this article. The authors have no connection with any or any financial company.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Tajul Arifin, K., Sulaiman, S., Md Saad, S. et al. Elevation of tumour markers TGF-β, M2-PK, OV-6 and AFP in hepatocellular carcinoma (HCC)-induced rats and their suppression by microalgae Chlorella vulgaris . BMC Cancer 17, 879 (2017). https://doi.org/10.1186/s12885-017-3883-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-017-3883-3