
Abstract
Background
The endothelial nitric oxide synthase (eNOS) gene deficiency is known to cause impaired coronary vasodilating capability in animal models. In the general clinical population, the eNOS gene polymorphisms, able to affect eNOS activity, were associated with cardiometabolic risk features and prevalence of coronary artery disease (CAD).
Aim
To investigate the association of eNOS Glu298Asp gene polymorphism, cardiometabolic profile, obstructive CAD and inducible myocardial ischemia in patients with suspected stable CAD.
Methods
A total of 506 patients (314 males; mean age 62 ± 9 years) referred for suspected CAD was enrolled. Among these, 325 patients underwent stress ECG or cardiac imaging to assess the presence of inducible myocardial ischemia and 436 patients underwent non-invasive computerized tomography or invasive coronary angiography to assess the presence of obstructive CAD. Clinical characteristics and blood samples were collected for each patient.
Results
In the whole population, 49.6% of patients were homozygous for the Glu298 genotype (Glu/Glu), 40.9% heterozygotes (Glu/Asp) and 9.5% homozygous for the 298Asp genotype (Asp/Asp). Obstructive CAD was documented in 178/436 (40.8%) patients undergoing coronary angiography while myocardial ischemia in 160/325 (49.2%) patients undergoing stress testing. Patients with eNOS Asp genotype (Glu/Asp + Asp/Asp) had no significant differences in clinical risk factors and in circulating markers. Independent predictors of obstructive CAD were age, gender, obesity, and low HDL-C. Independent predictors of myocardial ischemia were gender, obesity, low HDL-C and Asp genotype. In the subpopulation in which both stress tests and coronary angiography were performed, the Asp genotype remained associated with increased myocardial ischemia risk after adjustment for obstructive CAD.
Conclusion
In this population, low-HDL cholesterol was the only cardiometabolic risk determinant of obstructive CAD. The eNOS Glu298Asp gene polymorphism was significantly associated with inducible myocardial ischemia independently of other risk factors and presence of obstructive CAD.
Similar content being viewed by others

Introduction
Coronary artery disease (CAD) is a multifactorial disease with a complex pathogenesis, which manifests clinically with myocardial ischemia syndromes and continues to be the first cause of death worldwide [1, 2], especially in low-to-middle-income countries. The pathogenesis of CAD includes the interaction of multiple non-modifiable and modifiable risk factors associated with a progressive atherosclerotic process leading to obstructive disease [3]. Ischemic manifestations are generally a consequence of obstructive CAD but are also dependent on functional abnormalities of coronary vessels [4].
Growing evidence has consistently indicated endothelium, the main actor of vascular function, as a target of many cardio-metabolic conditions associated with atherogenesis and ultimately with CAD [5]. The endothelium is a highly dynamic cell layer involved in a multitude of physiologic processes ranging from the regulation of vascular tone to the homeostasis of systemic metabolism. The endothelial function is mainly guaranteed by the production of vasoactive molecules, particularly nitric oxide (NO) by endothelial nitric oxide synthase (eNOS), which plays an atheroprotective and vasodilatory role [6].
Experimental and clinical studies have suggested endothelium as an independent determinant of both vascular dysfunction and cardiometabolic deregulation. Total eNOS knockout mice (eNOS-/-), develop either coronary vascular dysfunction and hypertension, insulin resistance, hyperlipidemia [7,8,9]. Interestingly, also mice with a partial eNOS gene deletion (eNOS ±) demonstrate coronary vascular dysfunction while developing hypertension and overt insulin resistance when fed with a high-fat diet [8, 9].
In humans, single-nucleotide polymorphisms (SNPs) of the eNOS-encoding gene have been related with both CAD [10] and systemic metabolic abnormalities [11, 12]. In particular, its Glu298Asp variant has been associated with CAD predisposition [11] and with myocardial ischemia also independently of the presence of obstructive CAD [13]. Multiple eNOS gene variants have also been associated with the cardio-metabolic features of the metabolic syndrome (MeS) in the general population [14] and in patients with cardiovascular disease [15]. This is relevant since the MeS and its components have received recently particular attention as determinant of residual risk of CAD, i.e. the risk persisting despite current medical treatment [16, 17].
Despite these multiple evidences, there are not available studies in current patient populations able to clarify whether genetically determined eNOS abnormalities and cardiometabolic risk determinants have an independent or synergistic role in CAD and myocardial ischemia manifestations. In the frame of these premises, this study aimed to investigate the specific association of the eNOS Glu298Asp polymorphism and cardiometabolic risk factors with obstructive CAD and inducible myocardial ischemia in a population of patients with suspected stable coronary disease.
Materials and methods
Study design, patient population, and ethical approval
A total of 506 consecutive patients (314 males; mean age 62 ± 9 years) referred for suspected CAD within the BIOGEN-CARE Tuscan Region Italian Study were enrolled in six cardiology units. Among these, 325 patients underwent cardiac stress test (including stress ECG and/or cardiac imaging) to assess the presence of inducible myocardial ischemia and 436 patients underwent coronary study (including non-invasive computerized tomography and/or invasive coronary angiography) to assess the presence of obstructive CAD (> 50% stenosis in at least one major coronary vessel). In 255 patients both cardiac stress tests and coronary angiography were performed. The study flow chart is presented in Fig. 1.

Flow chart of the BIOGEN-CARE Tuscan Region Italian Study
The study was conducted according to the Declaration of Helsinki and its later amendments. The protocol was approved by the Coordinating Center Ethical Committee and all local Ethical Committees. All patients included in the study signed a written informed consent.
Clinical and laboratory characterization
For each patient, clinical and demographic characteristics including age, gender, smoking, family history of CAD, left ventricular ejection fraction (LVEF %) and medical therapy were recorded at enrollment before any diagnostic testing. Blood samples were collected from each patient for genotyping and measurements of clinical lipid and glucose parameters. Glucose, insulin, Total-cholesterol (Total-C), triglycerides (TG), low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), and Apolipoprotein A1 (ApoA1) levels (mg/dL) were evaluated using standard clinical chemistry laboratory methods. The HOMA index was calculated from the formula: HOMA index = fasting serum insulin (μU/ml) x fasting plasma glucose (FPG) (mmol/l)/22.5. The TG/HDL-C ratio was calculated as TG level divided by HDL-C levels.
The recorded non modifiable cardiovascular disease (CVD) risk factors included age, gender, family history of CVD. The modifiable CVD risk factors were defined, according to current international standards for CVD risk assessment [3] and for the definition of the MeS [18], based on patients’ clinical records plus annotated treatment and/or, when appropriate, on available clinical and biohumoral measurements. Definitions for modifiable CVD risk factors were: high systolic blood pressure (SBP) (hypertension under treatment or SBP > 130/85 mmHg), high LDL-C (hypercholesterolemia under treatment with statins or other lipid lowering drugs, or LDL-C > 115 mg/dL), previous or current tobacco smoking, high body mass index (BMI > 30 kg/m2), diabetes (treatment with hypoglycemic agents or insulin or FPG > 126 mg/dL), high FPG (diabetes or FPG > 100 mg/dL), high TG (> 150 mg/dL) and low HDL-C (< 40 mg/dL in males and < 50 mg/dL in females). The presence of MeS was defined by the presence in each patient of at least 3 of the following risk factors: high SBP, high BMI as a proxy for obesity, high FPG, high TG and low HDL-C [19].
Genetic analysis
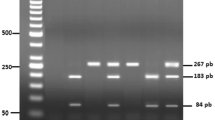
DNA was extracted from peripheral blood by using QIAamp DNA Blood Mini Kit (Qiagen, CA, USA) according to manufacturer’s instructions. The eNOS Glu298Asp polymorphism was determined by a standard polymerase chain reaction (PCR) amplification using the HotStarTaq Master Mix kit (QIAGEN Inc., Valencia, CA, USA) as follows: a first step of DNA denaturation at 95 ◦C for 15 min, 32 cycles at 94 ◦C for 30 s, 58 ◦C for 30 s and 72 ◦C for 1 min, and a final extension step at 72 ◦C for 10 min. Primers (F5’ -CATGAGGCTCAGCCCCAGAAC-3’ and R5’—AGTCAATCCCTTTGGTGCTCAC-3’) were selected from the eNOS Ensemble sequence database (Ensembl:ENSG00000164867) and designed using the free web-based application Primer3Plus (https://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi (accessed on 10 September 2021).
Statistical analysis
Continuous variables are presented as mean ± standard deviation (SD), categorical variables as relative percentages. Continuous data were first checked for normality (and ln transformed if required) before using parametric tests. Differences between two groups were compared by χ2 test for categorical variables or by two-tailed tests, unpaired Student’s t-test, for quantitative variables. Comparison of 3 means was performed by ANOVA, followed by Bonferroni post hoc test for comparison of any 2 groups. Multivariate linear regression was used to estimate the effect of cardiometabolic risk factors including age, male sex, family history of CAD, high SBP (as defined for MeS), high LDL-c (> 115mg/dL), smoking, BMI > 30 kg/m2, high FPG (as defined for MeS), low HDL-c (as defined for MeS), and high triglycerides (as defined for MeS), and eNOS Glu298Asp polymorphism (according to the dominant model Glu/Glu vs Asp) on obstructive CAD and/or inducible myocardial ischemia. A multivariable model was developed, including all variables with a p value ≤ 0.1 at univariable analysis, and then using backward and forward stepwise selections to build-up the final model. To not overfit the model with related variables, the variables were tested for collinearity. A p value < 0.05 was considered statistically significant. All statistical analyses were completed using Stata/SE 13.1 and SPSS Version 24.
Results
Clinical risk factors, metabolic profiles and eNOS genotype
The clinical risk factors, the metabolic characteristics and the eNOS genotype distribution of the whole study population and of groups defined according to the presence/absence of obstructive CAD or presence/absence of inducible myocardial ischemia are shown in Table 1.
Obstructive CAD was diagnosed in 178/436 (41%) patients undergoing coronary study. Patients with obstructive CAD were older, mainly males, had lower LVEF%, and more frequently referred typical angina. Patients with obstructive CAD, as compared with those without, had a similar traditional risk profile and received similar treatments at enrollment, but had a higher prevalence of low HDL-C (40 vs 22%, p < 0.001) and MeS (36 vs 26%, p = 0.04). Moreover, patients with obstructive CAD had lower levels of Total-C, LDL-C, HDL-C and ApoA1 as well as a higher TG/HDL-C ratio than patients without obstructive CAD.
Myocardial ischemia was documented in 160/325 (49.2%) patients undergoing stress testing. The risk profile of patients with inducible myocardial ischemia differed from that of patients without for older age, more frequent male gender and low HDL-C while high LDL-C and obesity were less frequent. They had lower levels of Total-C, LDL-C, HDL-C, as well as a lower levels of insulin and a lower HOMA index.
The genotype analysis in the whole population yielded 251 (50%) patients homozygous for the G894 allele (GG, Glu298Glu, Glu/Glu), 207 (41%) heterozygotes (GT, Glu298Asp, Glu/Asp), and 48 (9%) homozygous for the T894 allele (TT, Asp298Asp, Asp/Asp). The prevalence of the eNOS alleles satisfied the Hardy–Weinberg equilibrium law (χ2 = 0.31, p = 0.58). No difference in the genotype distribution of eNOS Glu298Asp variant was observed between patients with or without obstructive CAD. On the other hand, the eNOS Asp genotype was significantly more frequent in patients with inducible myocardial ischemia than in patients without (Table 1).
Between the clinical characteristics, only gender was significantly different in patients with different genotype with a male predominance in individuals with the mutant Asp genotype (Glu/Asp and Asp/Asp). No statistically significant differences in cardiometabolic risk factors prevalence and treatments were observed between groups. At laboratory analysis, patients with the mutant genotype showed lower levels of HDL-C, and ApoA1 as well as a higher TG/HDL-C ratio (Supplementary Table 1).
A multivariate logistic regression analysis was performed to identify risk factors (including non-modifiable and modifiable determinants) associated with obstructive CAD or inducible myocardial ischemia (Table 2).
Among non-modifiable risk factors, older age and male sex were independent predictors of obstructive CAD. The eNOS genotype was not associated with obstructive disease. Among modifiable risk factors, the MeS was not an independent determinant of obstructive CAD (Supplementary Table 2). Among MeS components only low levels of HDL-C were independently associated with obstructive CAD while obesity was inversely associated.
At multivariate logistic regression analysis for the prediction of inducible myocardial ischemia, the eNOS Asp genotype was a strong independent determinant of myocardial ischemia together with gender and low HDL-C levels while obesity remained inversely associated (Table 2). The MeS was not an independent determinant of inducible myocardial ischemia (Supplementary Table 2).
In the subgroup of patients who underwent both cardiac stress tests and coronary angiography (n = 255) the eNOS 298Asp genotype remained associated with an increased prevalence of inducible myocardial ischemia even after adjustment for the presence of obstructive CAD (OR 2.1; CI 1.2–3.7, p = 0.01) (Table 3). The clinical characteristics, risk factors and metabolic profiles were compared in the two extreme clinical presentations of this subgroup of patients (Supplementary Table 3). Patients having both obstructive CAD and inducible myocardial ischemia (CAD/MI, n = 84) were older, mainly males, with more frequent typical angina and with significantly reduced LVEF% compared with patients having neither obstructive CAD nor inducible myocardial ischemia (noCAD/noMI, n = 87). Regarding the cardiometabolic profile, CAD/MI patients had lower prevalence of obesity (12 vs 31%, p = 0.002) and of high LDL-C (39 vs 61%, p = 0.003) but a higher prevalence of low HDL-C (39 vs 10%, p < 0.001) and high TG (36 vs 22%, p = 0.014) compared to noCAD/noMI patients. Differences in laboratory data mirrored those in the cardiometabolic profile as shown in the whole population of patients with or without obstructive CAD (Table 1). The eNOS Asp genotype was more prevalent in CAD/MI compared to noCAD/noMI patients (58% vs 42%, p = 0.03).
Discussion
In this population of patients with suspected stable CAD, low HDL-C was the only cardiometabolic risk determinant independently associated with both obstructive disease and inducible myocardial ischemia. The eNOS Asp genotype was not associated with obstructive CAD but was a significant determinant of inducible myocardial ischemia independently of other risk factors, even after adjustment for presence of obstructive CAD. Obesity resulted inversely associated with both obstructive CAD and inducible myocardial ischemia.
Evidence shows that each component of the standard lipid profile including total cholesterol, HDL-C, LDL-C, and triglycerides are markers of cardiovascular risk [20] although the attention has been largely focused on total cholesterol and LDL-C levels since their substantial reduction is able to decrease the occurrence of cardiovascular events and the progression of coronary atherosclerosis [20, 21]. Nevertheless, more recently, a number of studies have shown an association between the co-occurrence of high TG and low HDL-C with a specific cardiometabolic profile, also referred to the pathophysiologic condition known as atherogenic dyslipidemia, strictly linked to cardiovascular atherosclerotic risk, but independent of LDL-C and LDL-C lowering therapies [17, 22].
More in general, low HDL-C and high TG are key components of the MeS that is defined by the presence of at least three out of five clinical features also including obesity, hypertension and hyperglycemia. The MeS has been related with increased risk of developing cardiovascular disease [19].
One of the major results in our population was that the MeS was not an independent determinant of obstructive CAD while low HDL-C was its only component that increased the risk of obstructive disease.
This finding expands the discussion around the role of HDL-C in contributing to the global atherosclerotic risk. The proposed effect of low HDL-C in predisposing to obstructive CAD is due to the anti-atherosclerotic properties of HDL mainly involving the so called “reverse cholesterol transport” mechanism [23], by which excess cholesterol is removed from the vessels wall and is delivered to the liver where it is metabolized. The efficiency of this mechanism is not only related with the abundance of HDL-C particles but also with their functionality. For example, there is recent evidence that high levels of TG rich lipoproteins cause not only a decrease in HDL lipoproteins, due to enhanced catabolism, but also some structural changes [24]. The alteration of HDL composition is able to decrease the anti-atherosclerotic properties of these particles involving not only their contribution to the reverse cholesterol transport but also their anti-inflammatory actions [25]. In the present study, high levels of TG were not per-se independent determinants of obstructive CAD risk but patients with obstructive CAD had significantly increased TG/HDL-C ratio as documented in other studies [17]. While the potential role of TG rich lipoproteins has been recently posed to great attention as a new direct determinant of residual atherosclerotic risk, the independent role of HDL-C is debated also due to the apparent inefficacy of treatments developed so far to increase HDL-C levels [26]. In our, as in previous studies, there are confounding factors which could influence HDL-C levels and their possible pathogenetic effects such as the use of lipid modifying medications. This area definitely needs further investigations.
Another important result of the present study was that low levels of HDL-C were predictors of both obstructive CAD and inducible MI together with the eNOS 298Asp SNP, suggesting a pathophysiologic interaction between the properties of HDL and a genetically determined primary endothelial dysfunction such that associated with eNOS Glu298Asp polymorphism [27]. While the association of low HDL-C with inducible myocardial ischemia in this population appeared mainly driven by its association with obstructive CAD, the link between the eNOS genetic variant and myocardila ischemia resulted independent.
Classically, the substrate of ischemic heart disease is mainly attributed to the presence of obstructive CAD although growing evidence demonstrates that this is not the only determining factor. Indeed, vessel obstruction is not always present in patients with angina [27, 28] and, on the other hand, myocardial ischemia may affect patients without obstructive CAD. Myocardial ischemia is a complex process involving more coronary districts from larger epicardial arteries to coronary microvessels. A “primitive” (genetic) endothelial dysfunction might represent a shared pathophysiological mechanism that, affecting both large and micro-vessels may determine myocardial ischemia [27]. The endothelium is a highly active metabolic and endocrine organ producing a multitude of different molecules, including NO which exerts a main role in both vascular and microvascular homeostasis [29].
In endothelium, NO is mainly produced by eNOS starting from L-arginine, multiple cofactors, and prosthetic groups. Nitric oxide plays an atheroprotective effect able to counteract harmful conditions (oxidative stress, inflammation, and platelet dysfunction) that trigger vascular damage in large epicardial arteries but, it has also a major role at the microcirculation level in the regulation of adequate coronary blood to be supplied to the myocardium [30].
We previously showed that mice with a partial or total deletion of eNOS gene had an impairment of coronary vasodilating capability (as documented by higher coronary resistance and lower percent decrease during Ach infusion) compared with wild types [9].
In humans, genetic variants of eNOS gene have been associated with cardiovascular disease including ischemic heart disease and its clinical manifestation. In particular, the point mutation G894T in the coding sequence of the eNOS that modifies the glutamic acid in aspartic acid at codon 298 (Glu298Asp) modifies the primary structure of the protein and has the potential to alter one or more functional properties of the enzyme. Two different studies have shown the eNOS protein containing Asp at position 298 is subject to selective proteolytic cleavage in endothelial cells and vascular tissues leading to a decreased NO synthesis [31, 32]. Two other reports showed contrasting results [33, 34]. However, it has been demonstrated that the eNOS G894T SNP affects the enzyme localization to caveolar membrane harming the eNOS signaling in response to shear stress [35]. In cardiovascular genetic association studies, the Glu298Asp variant has been associated with the development of myocardial infarction in very young individuals, whose coronary arteries are characterized by a very small atheromatic burden [36]. Additionally, this genetic variant was an independent risk factor for early STEMI presentation in young (under 45 years) patients, emphasizing the pivotal role of genetic susceptibility to endothelial dysfunction [37]. Additionally, the Glu298Asp polymorphism in the eNOS gene has been associated with coronary spasm in patients with variant angina, independently of the degree of atherosclerotic burden [38]. Notably, it had been shown that the T(-786)C promoter polymorphism and its interaction with exon 7 Glu298Asp affect endothelium-dependent vasodilation in human forearm microcirculation [39]. As a matter of fact, similarly to our present results, in a meta-analysis study, Luo et al. showed that the eNOS Glu298Asp polymorphism was associated with myocardial ischemia, also when corrected for the traditional cardiovascular risk factors [40]. More recently, in a population of patients with suspected acute coronary syndrome, the eNOS 298Asp SNP was an independent predictor of ischemic heart disease and clinical presentation of acute coronary syndrome [13]. The findings of our study confirm and expand all these previous reports showing a strong association of endothelial genetic predisposition with inducible myocardial ischemia independently of the presence of epicardial arteries obstructive disease.
Finally, in our population, obesity resulted inversely associated with both obstructive CAD and inducible myocardial ischemia. Obesity, a complex, multifactorial condition characterized by abnormal or excessive accumulation of body fats is increasing progressively in recent decades worldwide and it is one of the most significant global health challenges [41]. It is generally recognized as a risk factor for CAD mainly through its influence on the development and severity of comorbidities including hypertension, dyslipidemia, and glucose intolerance or diabetes [42]. Nevertheless, once CAD has developed, overweight or obese individuals have lower mortality than normal-weight people, the so-called “obesity paradox” [43]. The mechanism of the obesity paradox remains mainly unknown although it has been related with variable individual production of adipokines able to potentially neutralize the action of circulating pro-inflammatory cytokines involved in the atherosclerotic process [44]. In addition, clinical and experimental evidence indicates that obesity may be an independent risk factor for coronary microvascular dysfunction, possibly contributing to inducible myocardial ischemia, [45]. Cortigiani et al. recently demonstrated that obesity exerted a “paradoxical” protective effect in patients with stress-induced ischemia and/ or coronary microvascular dysfunction in a population of patients with known or suspected CAD [46]. In the present study obesity was not a significant predictor of obstructive CAD and/or inducible myocardial ischemia and our results suggest a possible protective role at least in this population.
Clinical implication
From a clinical point of view, the results of this study, whether confirmed in larger populations, highlight the possible role of endothelial genetic dysfunction in predisposing to inducible myocardial ischemia independently of the presence of epicardial arteries obstructive atherosclerotic disease. The underlying genetic and molecular pathways beyond myocardial ischemia in the absence of obstructive CAD are complex and deserve continuing mechanistic research. The study of genetically induced endothelial dysfunction may represent a major aspect in the comprehension of pathophysiological mechanisms governing myocardial ischemia independently (or jointly) to traditional risk factors. Interestingly in the present study we have shown that low HDL-C is among the modifiable cardiometabolic risk determinant the only one which is independently associated with both obstructive CAD and inducible myocardial ischemia. The possible synergistic actions between endothelial genetic dysfunction and HDL in determining the pathogenesis and the clinical manifestations of ischemic heart disease remain to be investigated. Confirming the major impact of a defective eNOS gene on myocardial ischemia induction could also pave the way for the research of novel new targeted drug strategies. Moreover, the identification of specific genetic and cardiometabolic profiles will allow recognizing patients at high risk for obstructive CAD and myocardial ischemia which might be fundamental for their prognostic stratification and correct management.
Study limitations
This study has several limitations. The study population was relatively small and included mainly patients from the Tuscany Region of Italy and, definitely, with the same Caucasian ethnicity. There are dissimilarities in the frequencies of eNOS Glu298Asp variants in different races thus, it is unknown whether our findings may be extended to a different population of patients with similar clinical features. Then, we investigated only a single SNP of the eNOS gene although previous studies suggested that there is linkage disequilibrium among the most clinically relevant eNOS SNPs. It is conceivable that other SNPs in eNOS gene as well as in other genes affecting endothelial function including also genes linked to lipoprotein metabolism can interact each other’s. Moreover, our population lacks information on waist circumference, which is a critical component of the MeS definition of central obesity. Finally, our patients were enrolled based on referral for suspected stable CAD and patients with previous disease were excluded. Accordingly, the present population is at relatively low risk and these results cannot be extended to populations with known CAD or at higher risk.
Conclusions
In patients with suspected stable CAD, the eNOS Glu298Asp gene polymorphism was a risk factor for inducible myocardial ischemia independent of the presence of obstructive coronary lesions and of other established risk determinants. In this population, among additional cardiometabolic risk factors, only low HDL-C was independently associated with obstructive CAD and, together with the eNOS Glu298Asp gene polymorphism, with inducible myocardial ischemia. In Fig. 2 a diagrammatic representation of possible mechanisms underlying the association of eNOS Glu298Asp polymorphism and low levels of HDL-C with inducible myocardial ischemia and obstructive CAD is presented. Further studies are needed to explore the role of eNOS genetic variants and their interaction with a specific cardiometabolic profile in ischemic heart disease.

Schematic representation of the possible mechanisms underlying the association of eNOS Glu298Asp polymorphism and low levels of HDL with inducible myocardial ischemia and obstructive CAD. The endothelial cells produce a multitude of different molecules, including nitric oxide (NO) which is mainly generated by endothelial nitric oxide synthase (eNOS) starting from L-arginine. The eNOS Glu298Asp polymorphism has been shown to modify the primary structure of the enzyme leading to a decreased NO synthesis causing reduced macrovascular and microvascular vasodilation (Tesauro M et al. Proc Natl Acad Sci U S A. 2000; Persu A et al. Hum Mol Genet 2002) which may trigger inducible myocardial ischemia. Reduced NO availability also limits its antiatherogenic properties, i.e., its capability to counteract conditions such as oxidative stress, and inflammation. On the other hand, an alteration of either the abundance and/or composition of HDL, is able to decrease the anti-atherosclerotic properties of these particles involving not only their contribution to the reverse cholesterol transport (RCT) but also their anti-inflammatory actions. HDL is involved in RCT by scavenging cholesterol from foam cells in atherosclerotic plaque. Briefly, ApoA-I initiates cholesterol efflux by binding to adenosine triphosphate-binding cassette transporter A1 (ABCA1), with further uptake through the ATP-binding cassette transporter G1 (ABCG1) and scavenger receptor class B type I (SR-BI). Cholesterol, esterified by lecithin-cholesterol acyl transferase (LCAT), is included into the lipid core of mature HDL, and then transported to the liver for excretion. HDL also promotes endothelial function by stimulating eNOS via shingosine-1-phosphate (S1P), and ApoA-I (Vecoli C et el. J Cell Biochem 2011). Moreover, HDL inhibits the expression of recruitment factors for inflammatory cells such as intercellular adhesion molecule-1 (ICAM-1), and vascular cell adhesion molecule (VCAM-1), stimulated by tumor necrosis factor-α (TNF-α) which is released by foam cells. Dotted Arrows denote reduced effects while dotted T bars denote reduced inhibitory effects
Availability of data and materials
The data and materials underlying this article will be shared on reasonable request to the corresponding authors.
Abbreviations
- CAD:
-
Coronary artery disease
- MI:
-
Myocardial ischemia
- HDL-C:
-
HDL cholesterol
- eNOS:
-
Endothelial nitric oxide synthase
- SBP:
-
Systolic blood pressure
- Total-C:
-
Total cholesterol
- LDL-C:
-
LDL cholesterol
- BMI:
-
Body mass index
- FPG:
-
Fasting plasma glucose;
- TG:
-
Triglycerides
- ApoA1:
-
Apolipoprotein 1
References
Moran AE, Forouzanfar MH, Roth G, Mensah GA, Ezzati M, Flaxman A. Christopher J L Murray, Mohsen Naghavi The global burden of ischemic heart disease in 1990 and 2010: the global burden of disease 2010 study. Circulation. 2014;129:1493–501.
GBD; 2015 Mortality and Causes of Death Collaborators. Global, regional and national life expectancy, all-cause mortality and cause-specific mortality for 249 causes of death, 1980–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. 2016;388:1459–544.
Vaduganathan M, Mensah GA, Turco JV, Fuster V, Roth GA. The Global Burden of Cardiovascular Diseases and Risk: A Compass for Future Health. J Am Coll Cardiol. 2022;80:2361–71.
Crea F, Montone RA, Rinaldi R. Pathophysiology of Coronary Microvascular Dysfunction. Circ J. 2022;86:1319–28.
Poredos P, Poredos AV, Gregoric I. Endothelial Dysfunction and Its Clinical Implications. Angiology. 2021;72:604–15.
Palmer RM, Ashton DS, Moncada S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature. 1988;333:664–6.
Duplain H, Burcelin R, Sartori C, Cook S, Egli M, Lepori M, Vollenweider P, Pedrazzini T, Nicod P, Thorens B, et al. Insulin resistance, hyperlipidemia, and hypertension in mice lacking endothelial nitric oxide synthase. Circulation. 2001;104:342–5.
Cook S, Hugli O, Egli M, Menard B, Thalmann S, Sartori C, Perrin C, Nicod P, Thorens B, Vollenweider P, et al. Partial gene deletion of endothelial nitric oxide synthase predisposes to exaggerated high-fat diet-induced insulin resistance and arterial hypertension. Diabetes. 2004;53:2067–72.
Vecoli C, Novelli M, Pippa A, Giacopelli D, Beffy P, Masiello P, L’Abbate A, Neglia D. Partial deletion of eNOS gene causes hyperinsulinemic state, unbalance of cardiac insulin signaling pathways and coronary dysfunction independently of high fat diet. PLoS ONE. 2014;9:e104156–66.
Yu J, Wu X, Ni J, Zhang J. Relationship between common eNOS gene polymorphisms and predisposition to coronary artery disease: Evidence from a meta-analysis of 155 published association studies. Genomics. 2020;112(3):2452–8.
Monti LD, Barlassina C, Citterio L, Galluccio E, Berzuini C, Setola E, Valsecchi G, Lucotti P, Pozza G, Bernardinelli L, Casari G, Piatti P. Endothelial nitric oxide synthase polymorphisms are associated with type 2 diabetes and the insulin resistance syndrome. Diabetes. 2003;52:1270–5.
Gonzalez-Sanchez JL, Martinez-Larrad MT, Saez ME, Zabena C, Martinez-Calatrava MJ, Serrano-Rios M. Endothelial nitric oxide synthase haplotypes are associated with features of metabolic syndrome. Clin Chem. 2007;53:91–7.
Severino P, D’Amato A, Prosperi S, Magnocavallo M, Mariani MV, Netti L, Birtolo LI, De Orchi P, Chimenti C, Maestrini V, et al. Potential Role of eNOS Genetic Variants in Ischemic Heart Disease Susceptibility and Clinical Presentation. J Cardiovasc Dev Dis. 2021;8:116.
Imamura A, Takahashi R, Murakami R, Kataoka H, Cheng XW, Numaguchi Y, Murohara T, Okumura K. The effects of endothelial nitric oxide synthase gene polymorphisms on endothelial function and metabolic risk factors in healthy subjects: the significance of plasma adiponectin levels. Eur J Endocrinol. 2008;158:189–95.
Vecoli C, Andreassi MG, Liga R, Colombo MG, Coceani M, Carpeggiani C, L’Abbate A, Neglia D. T(-786)→C polymorphism of the endothelial nitric oxide synthase gene is associated with insulin resistance in patients with ischemic or non ischemic cardiomyopathy. BMC Med Genet. 2012;13:92.
Guembe MJ, Fernandez-Lazaro CI, Sayon-Orea C, Toledo E, Moreno-Iribas C, for the RIVANA Study Investigators. Risk for cardiovascular disease associated with metabolic syndrome and its components: a 13-year prospective study in the RIVANA cohort. Cardiovasc Diabetol. 2020;19:195.
Caselli C, De Caterina R, Smit JM, Campolo J, El Mahdiui M, Ragusa R, Clemente A, Sampietro T, Clerico A, Liga R, et al. EVINCI and SMARTool. Triglycerides and low HDL cholesterol predict coronary heart disease risk in patients with stable angina Sci Rep. 2021;11:20714.
Grundy SM, Cleeman JI, Daniels SR, Donato KA, Eckel RH, Franklin BA, Gordon DJ, Krauss RM, Savage PJ, Smith SC Jr, et al. American Heart Association; National Heart, Lung, and Blood Institute. Diagnosis and management of the metabolic syndrome: an American Heart Association/National Heart, Lung, and Blood Institute Scientific statement. Circulation. 2005;112:2735–52.
Alshammary AF, Alharbi KK, Alshehri NJ, Vennu V, Ali KI. Metabolic Syndrome and Coronary Artery Disease Risk: A Meta-Analysis of Observational Studies. Int J Environ Res Public Health. 2021;18:1773.
Catapano AL, Graham I, De Backer G, Wiklund O, Chapman MJ, Drexel H, Hoes AW, Jennings CS, Landmesser U, Pedersen TR, et al. ESC/EAS guidelines for the management of dyslipidaemias. Eur Heart J. 2016;37:2999–3058.
Knuuti J, WijnsW SA, Capodanno D, Barbato E, Funck-Brentano C, Prescott E, Storey RF, Deaton C, Cuisset T, et al. ESC Guidelines for the diagnosis and management of chronic coronary syndromes: The Task Force for the diagnosis and management of chronic coronary syndromes of the European Society of Cardiology (ESC). Eur Heart J. 2019;2020(41):407–77.
Chen Z, Chen G, Qin H, Cai Z, Huang J, Chen H, Wu W, Chen Z, Wu S, Chen Y. Higher triglyceride to high-density lipoprotein cholesterol ratio increases cardiovascular risk: 10-year prospective study in a cohort of Chinese adults. J Diabetes Investig. 2020;11:475–81.
Pérez-Méndez Ó, Pacheco HG, Martínez-Sánchez C, Franco M. HDL-cholesterol in coronary artery disease risk: Function or structure? Clin Chim Acta. 2014;429:111–22.
Di Giorgi N, Michelucci E, Smit JM, Scholte AJHA, El Mahdiui M, Knuuti J, Buechel RR, Teresinska A, Pizzi MN, Roque A, et al. A specific plasma lipid signature associated with high triglycerides and low HDL cholesterol identifies residual CAD risk in patients with chronic coronary syndrome. Atherosclerosis. 2021;339:1–11.
Pirillo A, Catapano AL, Norata GD. Biological Consequences of Dysfunctional HDL. Curr Med Chem. 2019;26:1644–64.
von Eckardstein A, Nordestgaard BG, Remaley AT, Catapano AL. High-density lipoprotein revisited: biological functions and clinical relevance. Eur Heart J. 2023;44:1394–407.
Alexander Y, Osto E, Schmidt-Trucksäss A, Shechter M, Trifunovic D, Duncker DJ, Aboyans V, Bäck M, Badimon L, Cosentino F, et al. Endothelial function in cardiovascular medicine: A consensus paper of the European Society of Cardiology Working Groups on Atherosclerosis and Vascular Biology, Aorta and Peripheral Vascular Diseases, Coronary Pathophysiology and Microcirculation, and Thrombosis. Cardiovasc Res. 2021;117:29–42.
Kunadian V, Chieffo A, Camici PG, Berry C, Escaned J, Maas AHEM, et al. An EAPCI Expert Consensus Document on Ischaemia with Non-Obstructive Coronary Arteries in Collaboration with European Society of Cardiology Working Group on Coronary Pathophysiology & Microcirculation Endorsed by Coronary Vasomotor Disorders International Study Group. EuroIntervention. 2021;16:1049–69.
Padro T, Manfrini O, Bugiardini R, Canty J, Cenko E, De Luca G, Duncker DJ, Eringa EC, Koller A, Tousoulis D, et al. ESC Working Group on Coronary Pathophysiology and Microcirculation position paper on ‘coronary microvascular dysfunction in cardiovascular disease. Cardiovasc Res. 2020;116:741–55.
Camici PG, Crea F. Coronary microvascular dysfunction. N Engl J Med. 2007;356:830–40.
Tesauro M, Thompson WC, Rogliani P, Qi L, Chaudhary PP, Moss J. Intracellular processing of endothelial nitric oxide synthase isoforms associated with differences in severity of cardiopulmonary diseases: cleavage of proteins with aspartate vs. glutamate at position 298. Proc Natl Acad Sci U S A. 2000;6:2832–5.
Persu A, Stoenoiu MS, Messiaen T, Davila S, Robino C, El-Khattabi O, Mourad M, Horie S, Feron O, Balligand JL, et al. Modifier effect of eNOS in autosomal dominant polycystic kidney disease. Hum Mol Genet. 2002;11:229–41.
Fairchild TA, Fulton D, Fontana JT, Gratton JP, McCabe TJ, Sessa WC. Acidic hydrolysis as a mechanism for the cleavage of the Glu (298)/asp variant of human endothelial nitric-oxide synthase. J Biol Chem. 2001;276:26674–9.
McDonald DM, Alp NJ, Channon KM. Functional comparison of the endothelial nitric oxide synthase Glu298Asp polymorphic variants in human endothelial cells. Pharmacogenetics. 2004;14:831–9.
Joshi MS, Mineo C, Shaul PW, Bauer JA. Biochemical consequences of the NOS3 Glu298Asp variation in human endothelium: altered caveolar localization and impaired response to shear. FASEB J. 2007;21:2655–63.
Zigra AM, Rallidis LS, Anastasiou G, Merkouri E, Gialeraki A. eNOS gene variants and the risk of premature myocardial infarction. Dis Markers. 2013;34(431–6):37.
Isordia-Salas I, Leaños-Miranda A, Borrayo-Sánchez G. The Glu298ASP polymorphism of the endothelial nitric oxide synthase gene is associated with premature ST elevation myocardial infarction in Mexican population. Clin Chim Acta. 2010;411:553–7.
Chang K, Baek SH, Seung KB, Kim PJ, Ihm SH, Chae JS, Kim JH, Hong SJ, Choi KB. The Glu298Asp polymorphism in the endothelial nitric oxide synthase gene is strongly associated with coronary spasm. Coron Artery Dis. 2003;14:293–9.
Rossi GP, Taddei S, Virdis A, Cavallin M, Ghiadoni L, Favilla S, Versari D, Sudano I, Pessina AC, Salvetti A. The T-786C and Glu298Asp polymorphisms of the endothelial nitric oxide gene affect the forearm blood flow responses of Caucasian hypertensive patients. J Am Coll Cardiol. 2003;41:938–45.
Luo JQ, Wen JG, Zhou HH, Chen XP, Zhang W. Endothelial nitric oxide synthase gene G894T polymorphism and myocardial infarction: A meta-analysis of 34 studies involving 21,068 subjects. PLoS ONE. 2014;9: e87196.
Poirier P, Giles T, Bray G, Hong Y, Stern JS, Pi-Sunyer FX, Eckel RH. Obesity and cardiovascular disease: pathophysiology, evaluation, and effect of weight loss. Circulation. 2006;113:898–918.
Azab M, Al-Shudifat AE, Johannessen A, Al-Shdaifat A, Agraib LM, Tayyem RF. Are risk factors for coronary artery disease different in persons with and without obesity? Metab Syndr Relat Disord. 2018;16:440–5.
Mandviwala T, Khalid U, Deswal A. Obesity and cardiovascular disease:a risk factor or a risk marker? Curr Atheroscl Rep. 2016;18:21.
De Schutter A, Kachur S, Lavie CJ, Boddepalli RS, Patel DA, Milani RV. The impact of inflammation on the obesity paradox in coronary heart disease. Int J Obes. 2016;40:1730–5.
Sorop O, Olver TD, van de Wouw J, Heinonen I, van Duin RW, Duncker DJ, Merkus D. The microcirculation: a key player in obesity-associated cardiovascular disease. Cardiovasc Res. 2017;113:1035–45.
Cortigiani L, Haberka M, Ciampi Q, Bovenzi F, Villari B, Picano E. The obesity paradox in the stress echo lab: fat is better for hearts with ischemia or coronary microvascular dysfunction. Int J Obes (Lond). 2021;45:308–15.
Funding
This study was supported by a Grant of the “Regional Health Research Program 2009” from the Regione Toscana, Italy.
Author information
Authors and Affiliations
Contributions
CV, and DN contributed to the conception and design of the work. CV, CC, MM, GT, CP, SM, and NB contributed to the acquisition, analysis, and interpretation of data for the work. CV drafted the manuscript. RP, SV, FB, AN, LG, AC, ME, and DN contributed to the discussion and critically revised the manuscript. All gave final approval and agreed to be accountable for all aspects of work ensuring integrity and accuracy.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The protocol was approved by the Coordinating Center Ethical Committee and all local Ethical Committees. All patients included in the study signed a written informed consent.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Vecoli, C., Caselli, C., Modena, M. et al. Low HDL cholesterol and the eNOS Glu298Asp polymorphism are associated with inducible myocardial ischemia in patients with suspected stable coronary artery disease. BMC Cardiovasc Disord 24, 176 (2024). https://doi.org/10.1186/s12872-024-03846-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12872-024-03846-7