
Abstract
Background
Pathogenesis and diagnostic biomarkers of aortic dissection (AD) can be categorized through the analysis of differential metabolites in serum. Analysis of differential metabolites in serum provides new methods for exploring the early diagnosis and treatment of aortic dissection.
Objectives
This study examined affected metabolic pathways to assess the diagnostic value of metabolomics biomarkers in clients with AD.
Method
The serum from 30 patients with AD and 30 healthy people was collected. The most diagnostic metabolite markers were determined using metabolomic analysis and related metabolic pathways were explored.
Results
In total, 71 differential metabolites were identified. The altered metabolic pathways included reduced phospholipid catabolism and four different metabolites considered of most diagnostic value including N2-gamma-glutamylglutamine, PC(phocholines) (20:4(5Z,8Z,11Z,14Z)/15:0), propionyl carnitine, and taurine. These four predictive metabolic biomarkers accurately classified AD patient and healthy control (HC) samples with an area under the curve (AUC) of 0.9875. Based on the value of the four different metabolites, a formula was created to calculate the risk of aortic dissection. Risk score = (N2-gamma-glutamylglutamine × -0.684) + (PC (20:4(5Z,8Z,11Z,14Z)/15:0) × 0.427) + (propionyl carnitine × 0.523) + (taurine × -1.242). An additional metabolic pathways model related to aortic dissection was explored.
Conclusion
Metabolomics can assist in investigating the metabolic disorders associated with AD and facilitate a more in-depth search for potential metabolic biomarkers.
Similar content being viewed by others
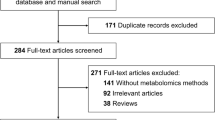
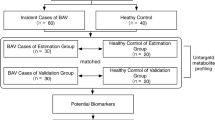

Introduction
Aortic dissection (AD) is a life-threatening condition caused by a tear in the inner layer of the aorta, which results in a separation of the layers of the aortic wall and subsequent formation of a true lumen and a false lumen [1, 2]. Current methods of diagnosing AD include the D-dimer test and contrast-enhanced computed tomography (CT) of the aorta. D-Dimers have high sensitivity, but extremely poor specificity, and enhanced CT requires expensive equipment and the rays cause potential harm to the patients [3]. The pursuit of early diagnosis and the identification of more effective therapeutic targets are crucial to ensuring the timely treatment of AD and guiding its therapy.
Metabolomics is an effective and widely used technology that has been increasingly employed to comprehensively profile disease progression [4]. Serum is frequently considered as a pool of metabolites and the analysis of serum metabolomics has been an efficient tool to identify potential metabolic biomarkers of diseases, that may improve early diagnosis, prognostic prediction, and personalized therapy [5]. Although metabolomics has made great progress in the diagnosis and treatment of diseases, most metabolomics studies mainly focus on acute myocardial infarction and coronary heart disease. There are few studies on metabolites of AD. Diagnosis of AD at an early stage is complicated and challenging. Potential metabolic biomarkers in AD patients are not clear.
This study was designed to explore the metabolites and metabolic pathways of patients with aortic dissection and search for potential biomarkers.
Methods
Patients and study design
Patients were enrolled from Shenzhen People’s Hospital between October 2019 and April 2021. Inclusion criteria were a diagnosis of acute aortic dissection, of less than two weeks duration, confirmed by aortic vascular enhanced CT. Patients were excluded with intramural hematoma of the aorta, aortic aneurysm, embolism, malignant tumor, severe infectious disease, trauma, a recent surgical procedure, and severe heart failure with left ventricular ejection fraction less than 20%. Informed consent was obtained from all patients. This study was performed under the guidance of the Helsinki Declaration and was approved by all centers.
Serum samples were collected before the aortic surgery and were immediately frozen at -80 °C for metabolomic analysis.
Chemical and sample preparation
All chemicals and solvents were of analytical or HPLC (High Performance Liquid Chromatography) grade. Water, methanol, acetonitrile, and formic acid were purchased from CNW technologies GmbH (Dusseldorf Germany). The serum was separated after − 4 °C refrigeration for 60 min, and stored at -80 °C for metabolite analysis. Samples stored at -80 C were thawed at room temperature and 200 µL of the sample was added to a 1.5 mL Eppendorf tube with 10 µL of 2-chloro-l-phenylalanine (0.3 mg/mL) dissolved in methanol as internal standard. Then, the tube was vortexed for 10 s, and a 15 µL ice-cold mixture of methanol and acetonitrile was added to the sample. The mixtures were vortexed for 1 min, ultrasonicated at an ambient temperature of 25–28 °C for 10 min, and stored at -20 °C for 30 min. The extract was centrifuged at 13,000 rpm at 4 °C for 15 min, 1.0mL of supernatant in a brown and glass vial was dried in a freeze concentration centrifugal dryer and 15 µL mixture of methanol and water were added to each sample, vortexed for 30 s, and then kept at 4 °C for 2 min. Samples were centrifuged at 1300 rpm, 4 °C for 5 min. Then, 100 µL supernatant aliquots from each tube were collected using crystal syringes, filtered through 0.22 pm microflora and transferential vials. These were stored at -80 °C for liquid chromatography-mass spectrometry (LC-MS) analysis.
An ACQUITY UHPLC system (Waters Corporation, Milford, USA) was used to analyze the metabolic profiling in both ESI positive and ESI negative ion modes. An ACQUITY UPLC BEH C18 column (1.7 μm, 2.1 × 100 mm) were employed in both positive and negative modes. The binary gradient elution system consisted of (A) water (containing 0.1% formic acid, v/v) and (B) acetonitrile (containing 0.1% formic acid, v/v) .We separated sample by following gradient: 0 min, 5% B; 2 min, 20% B; 4 min, 60% B; 11 min, 100% B; 13 min, 100% B; 13.5 min, 5% B and 14.5 min, 5%B .The flow rate was 0.4 mL/min and column temperature was 45 ℃. All the samples were kept at 4℃ during the analysis. The injection volume was 5 µL.
Parameters of mass spectrometry were as follows: Ion source temperature, 550 °C (+) and 550 °C (−); ion spray voltage, 5500 V (+) and 4500 V (−); curtain gas of 35 PSI; declustering potential, 100 V (+) and − 100 V (−); collision energy, 10 eV (+) and − 10 eV (−); and interface heater temperature, 550 °C(+) and 600 °C (−). For IDA analysis, range of m/z was set as 25–1000, the collision energy was 30 eV.
Quality control (QC) samples were prepared by pooling aliquots of all samples and injected every 10 samples throughout the analytical run to provide a set of data from which repeatability could be assessed.
Data preprocessing and statistical analysis
We analyze the LC-MS raw data by the orogenesis QI software (Waters Corporation, Milord, USA), with the parameters was set in 5 ppm (parts per million) ,10 ppm and 0.02 min for the precursor tolerance (PT), fragment tolerance (FT) and retention time (RT). According to three-dimensional data sets including m/Z, peaks RT and peak intensities. We acquired the Excel file which obtained filtered data. The internal standard was used for data QC reproducibility.
Metabolites were identified by progenesis QI Data Processing Software, based on public databases and self-built databases. We acquired combined data from combined positive and negative data, and imported it into the R ropls package. We conducted Principle component analysis (PCA) and orthogonal partial least-squares-discriminant analysis (O) PLS-DA to compare different of metabolic alterations among experimental groups. Overall contribution of each variable was ranked in the OPLS-DA model. Those variables with a VIP > 1 are considered relevant for group discrimination.
In this study, the default seven-round cross-validation was applied with one-seventh of the samples excluded from the material model inch round, to guard against overfitting.
The deferential metabolite was selected based on the combination of the statistically significant threshold of variable influence on projection (VIP) values obtained from the OPLS- DÁ model and p-values from a two-tailed Student’s test on the normalized peak areas, where metabolites with VIP values larger than 1.0 and p < 0.05 were considered a deferential metabolite.
Results
This study followed-up 44 patients who were suspected of having AD and 30 patients diagnosed with acute AD were enrolled (see Fig. 1). Sixty samples of human serum, 30 from AD patients and 30 from healthy controls (HC), were collected for LC-MS analysis. Nontargeted profiling was performed to obtain the serum metabolic characteristics as comprehensively as possible. In this study, by employing nontargeted LC-MS platforms, the metabolic profiles of ADs and HCs could be analyzed in detail and the related disordered metabolism pathway underlying AD development investigated. A novel biomarker panel was subsequently identified and validated for differentiating ADs and HCs and its clinical practicability assessed (Fig. 1).

Study design: 30 AD patients and 30 healthy controls
Baseline clinical characteristics of the AD and HC groups were compared in Table 1. Compared with HC subjects, AD patients had higher levels of heart rate and D-Dimers. There were no significant differences in age, blood pressure levels on admission, body mass index (BMI), whether hypertension and diabetes were present, cTnI, NT-pro BNP, total cholesterol, low- and high-density lipoprotein cholesterol and triglyceride levels.
To reveal AD pathogenesis serum metabolites, the pairwise comparisons between the AD and HC groups were conducted. Based on the partial least squares (PLS-DA) method, the data showed significant separation between the two groups of samples (Fig. 2A) and the permutation test showed that the PLS-DA score were dependable without overfitting with R2 = 0 to 0.44 and Q2 = 0 to 0.455 (Supplementary materials, Fig. 2.)

PLS-DA score plots and metabolic pathways
The screening criteria were that the variable’s importance in projection (VIP) > 1 and a t-test where p < 0.05. Once the metabolite fulfilled this condition, it was considered a potential biomarker. The AD and HC samples were compared to identify and characterize specific metabolites and underlying metabolic pathways. A total of 1221 metabolites, including 426 downregulated and 795 upregulated metabolites, were screened by LC-MS analysis and focused on 71 differential metabolites between two groups, Visualizing the p-value, VIP, and fold change values was helpful for screening differential metabolites, as shown in Fig. 3.

Thirteen specific metabolic biomarkers for distinguishing AD from HC
13 specific metabolic biomarkers were identified for distinguishing AD from HC (Table 2). A regression algorithm [6,7,8,9], least absolute shrinkage and selection operator (LASSO), was used to further identify four predictive metabolic biomarkers, i.e., N2-gamma-Glutamylglutamine, PC (20:4(5Z,8Z,11Z,14Z)/15:0), propionyl carnitine and taurine (Fig. 4), which served as a molecular diagnostic signature of AD. The training and test sets were set according to the concentration of metabolites and these four predictive metabolic biomarkers accurately classified the AD and HC samples with an AUC of 0.9875. Based on the value of the four different metabolites, a formula was developed to calculate the risk of aortic dissection:
As the circulatory levels of taurine and N2-gamma-glutamylglutamine are decreased, whereas, those of PC and propionylcarnitine are elevated in AD patients compared to HC subjects, Therefore, we use the Ratiometric to demonstrate it’s diagnostic potential.
R1 = PC(20:4(5Z,8Z,11Z,14Z)/15:0)/taurine = 1.8701/0.4862 = 3.85.
R2 = PC(20:4(5Z,8Z,11Z,14Z)/15:0)/N2-gamma-glutamylglutamine = 1.8701/0.3725 = 5.02.
R3 = propionylcarnitine/taurine = 2.3394/0.4862 = 4.81.
R4 = propionylcarnitine/N2-gamma-glutamylglutamine = 2.3394/0.3725 = 6.28.

Diagnostic outcomes and prediction accuracies: the diagnostic outcomes are shown in the receiver-operating characteristic (ROC) curves for comparison between ADs and HCs. The prediction accuracies by the biomarkers in the training and test sets were compared between two groups
To reveal the metabolic processes of metabolites and analyze the pathways in which these metabolites were involved, 71 differential metabolites were introduced into three Kyoto encyclopedia of genes and genomes (KEGG) databases HMDB, Lipimaps, and Metlin. The results suggested that the differential metabolites after screening were enriched in these three metabolic pathways choline metabolism in cancer, neuroactive ligand-receptor interaction and glycerophospholipid metabolism (Fig. 2B). The study found that of the metabolic biomarkers, PC (20:4(5Z,8Z,11Z,14Z)/15:0) played a key role in the glycerophospholipid metabolism pathway (Fig. 5) and PC (20:4(5Z,8Z,11Z,14Z)/15:0) activated the glycerophospholipid metabolism pathway downstream of phosphitylated, including cell-cycle progression, proliferation migration angiogenesis, and actin reorganization.

Proposed mechanisms of metabolite phosphitylated on pathogenesis of AD
Discussion
This study found that a metabolomic strategy could be used to identify potential metabolic markers of AD and to explore the metabolic pathways related to its occurrence. We carried out a comprehensive metabolomic evaluation of 30 AD patients. Metabolic phenotypes revealed significant pattern differences between patients with AD and HC, suggesting that AD may involve some metabolic disturbance, such as phospholipid metabolism disorder.
Patients with AD have a higher heart rate and elevated D-dimer levels [10]. . Except for heart rate at admission and D-dimer levels, the clinical baseline data were similar between the two patient groups .
Analysis of metabolic pathways enriched by 71 different metabolites showed that the different metabolites were involved in five pathways, including choline metabolism in cancer, neuroactive ligand-receptor interaction, glycerophospholipid metabolism, sphingolipid metabolism, and ether lipid metabolism. KEGG analysis showed oxidative stress and lipid transport and metabolism were significantly implicated.
Compared to HCs, patients with AD had down-regulated N2-gamma-glutamylglutamine(γ-glu) and taurine, both of which play a key role in amino acid and fat metabolism. especially taurine. There is evidence that it affects mitochondrial bioenergetics, counteracts lipid peroxidation and even increases cellular antioxidant defense in response to inflammation [11].
The patients with AD also had up-regulated propionyl carnitine and PC(20:4(5Z,8Z,11Z,14Z)/15:0). Propionyl carnitine is a free radical that can produce positive effects on endothelial function and protect endothelia from oxidative stress [12]. The increase in propionyl carnitine may be related to the repair of endometria after aortic dissection tear.
The PC(20:4(5Z,8Z,11Z,14Z)/15:0) is the precursor of phospholipids, which can decholine to produce phospholipids through a series of metabolic reactions in the body. The phospholipid metabolism disorder may be closely related to the occurrence of AD.
The diagnosis of acute AD relies on imaging such as vascular enhanced CT [13]. For community hospitals that lack relevant equipment, this is not conducive for the early diagnosis of acute aortic dissection, so finding a high-sensitivity metabolite of AD would be important for rescuing these patients.
This study identified and validated the signature consisting of four differential metabolites known to be related to AD pathogenesis that can accurately distinguish AD patients from healthy controls. Despite limitations such as the related small sample size, evidence showed that metabolites may be adopted as markers for the diagnosis of AD.
Aortic dissection results from a tear in the intimal layer of the aorta, leading to the separation of the layers of the aortic wall [14]. . While Extracellular matrix degradation and inflammation may contribute to the occurrence of the disease, the precise trigger of aortic dissections is still unknown [2]. Within the metabolic pathways enriched with differential metabolites, this study constructed a simple metabolic pathway model, wherein choline promoted the increase of phospholipids through transport proteins and second messengers. Phospholipids activated downstream pathways including cell-cycle progression, proliferation migration angiogenesis and actin reorganization, However, PC(20:4(5Z,8Z,11Z,14Z)/15:0) controls and regulates other pathways by negative feedback.
Previous studies considered that AD would activate cell-cycle progression [15, 16] and this is consistent with these results. The inflammatory response is deemed crucial for AD [17,18,19], inducing the expression of secretory molecules, including cytokines and extracellular matrix (ECM) molecules in smooth muscle cells (SMCs), thereby enhancing proliferative capacity [20]. Inflammatory cells, such as lymphocytes and macrophages, secrete multiple inflammatory cytokines to promote vascular adhesion molecule expression [21], eventually leading to proliferation migration angiogenesis. Inflammatory cells also contribute to the apoptosis of SMCs in the aortic artery and lead to medial degradation, exacerbating intimal tears [22].
Actin is an important protein, playing a critical role in many cellular functions and the interaction of actin with myosin forms the basis of muscle contraction [23]. In human aortic smooth muscle cells, smooth muscle α-actin (α-SMA) participated in filamentous actin formation and its expression can facilitate stress fiber formation and cell contraction in human aortic smooth muscle cells [24]. During the development of AD, smooth muscle cells may switch occurrence of phenotype [25],lead to actin reorganization and cause smooth muscle cell apoptosis [25], eventually causing the occurrence and progression of aortic dissection.
This study had some limitations. It failed to functionally verify the differential metabolites further. The sample size was small and could be further expanded in the future.
Conclusion
In conclusion, we employed a metabolomics strategy to characterize metabolite signatures and their potential mechanisms associated with AD. Our results demonstrate that N2-gamma-Glutamylglutamine(γ-glu), taurine, propionyl carnitine, and PC (20:4(5Z,8Z,11Z,14Z)/15:0) were identified as critical metabolites. They may become markers for diagnosing aortic dissection. Moreover, phospholipids play a key role in the phosphorylated metabolite, leading to cell-cycle progression, proliferation, migration, angiogenesis, and actin reorganization. This evaluation enhances the understanding of AD pathogenesis.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Abbreviations
- AD:
-
Aortic Dissection
- CT:
-
Computed Tomography
- HPLC:
-
High Performance Liquid Chromatography
- LC-MS:
-
Liquid Chromatography-Mass Spectrometry
- QC:
-
Quality control
- RT:
-
Retention Time
- PCA:
-
Principal Component Analysis
- LC-MS:
-
Liquid Chromatography-Mass Spectrometry
- (O) PLS-DA:
-
Orthogonal Partial Least-Squares-Discriminant Analysis
- HC:
-
Healthy Controls
- BMI:
-
Body Mass Index
- SMCs:
-
Smooth Muscle Cells
- ECM:
-
Extracellular Matrix
- α-SMA:
-
Smooth Muscle α-actin
References
Silaschi M, Byrne J, Wendler O. Aortic dissection: medical, interventional and surgical management. Heart. 2017;103:78–87.
Nienaber CA, Clough RE, Sakalihasan N, Suzuki T, Gibbs R, Mussa F, Jenkins MP, Thompson MM, Evangelista A, Yeh JS, Cheshire N. Rosendahl U and Pepper J. Aortic dissection. Nat Rev Dis Primers. 2016;2:16053.
Pape LA, Awais M, Woznicki EM, Suzuki T, Trimarchi S, Evangelista A, Myrmel T, Larsen M, Harris KM, Greason K, Di Eusanio M, Bossone E, Montgomery DG, Eagle KA, Nienaber CA, Isselbacher EM, O’Gara P, Presentation. Diagnosis, and outcomes of Acute Aortic dissection: 17-Year trends from the International Registry of Acute Aortic Dissection. J Am Coll Cardiol. 2015;66:350–8.
Sabatine MS, Liu E, Morrow DA, Heller E, McCarroll R, Wiegand R, Berriz GF, Roth FP, Gerszten RE. Metabolomic identification of novel biomarkers of myocardial ischemia. Circulation. 2005;112:3868–75.
Tang WH, Wang Z, Cho L, Brennan DM, Hazen SL. Diminished global arginine bioavailability and increased arginine catabolism as metabolic profile of increased cardiovascular risk. J Am Coll Cardiol. 2009;53:2061–7.
Liu X, Zheng X, Wang J, Zhang N, Leung KS, Ye X, Cheng L. A long non-coding RNA signature for diagnostic prediction of sepsis upon ICU admission. Clin Translational Med. 2020;10:e123.
Yang Y, Zhang Y, Li S, Zheng X, Wong MH, Leung KS, Cheng L. A robust and generalizable Immune-Related signature for Sepsis Diagnostics. IEEE/ACM Trans Comput Biol Bioinf. 2022;19:3246–54.
Xu C, Li W, Li T, Yuan J, Pang X, Liu T, Liang B, Cheng L, Sun X, Dong S. Iron metabolism-related genes reveal predictive value of acute coronary syndrome. Front Pharmacol. 2022;13:1040845.
Liu S, Zhao W, Liu X, Cheng L. Metagenomic analysis of the gut microbiome in atherosclerosis patients identify cross-cohort microbial signatures and potential therapeutic target. FASEB Journal: Official Publication Federation Am Soc Experimental Biology. 2020;34:14166–81.
Nazerian P, Mueller C, Soeiro AM, Leidel BA, Salvadeo SAT, Giachino F, Vanni S, Grimm K, Oliveira MT Jr., Pivetta E, Lupia E, Grifoni S, Morello F. Diagnostic accuracy of the aortic dissection detection risk score plus D-Dimer for Acute aortic syndromes: the ADvISED prospective Multicenter Study. Circulation. 2018;137:250–8.
Seidel U, Huebbe P, Rimbach G, Taurine. A Regulator of Cellular Redox Homeostasis and skeletal muscle function. Mol Nutr Food Res. 2019;63:e1800569.
Andreozzi GM. Propionyl l-carnitine: intermittent claudication and peripheral arterial disease. Expert Opin Pharmacother. 2009;10:2697–707.
Nienaber CA, Clough RE. Management of acute aortic dissection. Lancet. 2015;385:800–11.
Hagan PG, Nienaber CA, Isselbacher EM, Bruckman D, Karavite DJ, Russman PL, Evangelista A, Fattori R, Suzuki T, Oh JK, Moore AG, Malouf JF, Pape LA, Gaca C, Sechtem U, Lenferink S, Deutsch HJ, Diedrichs H, Marcos J, Llovet A, Gilon D, Das SK, Armstrong WF, Deeb GM, Eagle KA. The International Registry of Acute Aortic Dissection (IRAD): new insights into an old disease. JAMA. 2000;283:897–903.
Wang L, Zhang J, Fu W, Guo D, Jiang J, Wang Y. Association of smooth muscle cell phenotypes with extracellular matrix disorders in thoracic aortic dissection. J Vasc Surg. 2012;56:1698–709. 1709.e1.
Müller BT, Modlich O, Prisack HB, Bojar H, Schipke JD, Goecke T, Feindt P, Petzold T, Gams E, Müller W, Hort W, Sandmann W. Gene expression profiles in the acutely dissected human aorta. Eur J Vasc Endovasc Surg. 2002;24:356–64.
Son BK, Sawaki D, Tomida S, Fujita D, Aizawa K, Aoki H, Akishita M, Manabe I, Komuro I, Friedman SL, Nagai R. And Suzuki T. Granulocyte macrophage colony-stimulating factor is required for aortic dissection/intramural haematoma. Nat Commun. 2015;6:6994.
Anzai A, Shimoda M, Endo J, Kohno T, Katsumata Y, Matsuhashi T, Yamamoto T, Ito K, Yan X, Shirakawa K, Shimizu-Hirota R, Yamada Y, Ueha S, Shinmura K, Okada Y, Fukuda K, Sano M. Adventitial CXCL1/G-CSF expression in response to acute aortic dissection triggers local neutrophil recruitment and activation leading to aortic rupture. Circul Res. 2015;116:612–23.
Tieu BC, Lee C, Sun H, Lejeune W, Recinos A 3rd, Ju X, Spratt H, Guo DC, Milewicz D, Tilton RG, Brasier AR. An adventitial IL-6/MCP1 amplification loop accelerates macrophage-mediated vascular inflammation leading to aortic dissection in mice. J Clin Investig. 2009;119:3637–51.
Hayashi-Hori M, Aoki H, Matsukuma M, Majima R, Hashimoto Y, Ito S, Hirakata S, Nishida N, Furusho A, Ohno-Urabe S, Fukumoto Y. Therapeutic effect of rapamycin on aortic dissection in mice. Int J Mol Sci. 2020;21.
Lindholt JS, Shi GP. Chronic inflammation, immune response, and infection in abdominal aortic aneurysms. Eur J Vasc Endovasc Surg. 2006;31:453–63.
Luo F, Zhou XL, Li JJ, Hui RT. Inflammatory response is associated with aortic dissection. Ageing Res Rev. 2009;8:31–5.
Dominguez R, Holmes KC. Actin structure and function. Annual Rev Biophys. 2011;40:169–86.
Sun Y, Yang Z, Zheng B, Zhang XH, Zhang ML, Zhao XS, Zhao HY, Suzuki T, Wen JK. A Novel Regulatory mechanism of smooth muscle α-Actin expression by NRG-1/circACTA2/miR-548f-5p Axis. Circul Res. 2017;121:628–35.
Clément M, Chappell J, Raffort J, Lareyre F, Vandestienne M, Taylor AL, Finigan A, Harrison J, Bennett MR, Bruneval P, Taleb S. Jørgensen HF and Mallat Z. Vascular smooth muscle cell plasticity and autophagy in dissecting aortic aneurysms. Arterioscler Thromb Vasc Biol. 2019;39:1149–59.
Acknowledgements
We thank the Department of Cardiology, Shenzhen Cardiovascular Minimally Invasive Medical Engineering Technology Research and Development Center, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology) data.
Funding
This study was supported by National Natural Science Foundation of China (grant No.82000058 and 32370711).
Author information
Authors and Affiliations
Contributions
Yun Gong , Tangzhiming Li Qiyun Liu, Xiaoyu Wang, Zixian Deng conceptualized and designed the study, carried out the initial analyses, drafted the initial manuscript and reviewed and revised the manuscript; Huadong Liu, Biao Yu, Lixin Cheng critically reviewed and revised the manuscript; and all authors approved the final manuscript as submitted.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The research ethics committee of the Shenzhen People’s Hospital.in China approved this study and waived informed consent. All the participants agreed with this study. The written consents were taken from patients before participation. The study was carried out in accordance with Helsinki Declaration. All methods were carried out in accordance with the declaration of Helsinki.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gong, Y., Li, T., Liu, Q. et al. Analysis of differential metabolites in serum metabolomics of patients with aortic dissection. BMC Cardiovasc Disord 24, 226 (2024). https://doi.org/10.1186/s12872-024-03798-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12872-024-03798-y