
Abstract
Bell pepper (Capsicum annuum L.); an important spice crop of the region is a rich source of vitamins and antioxidants having many health benefits. Many biotic and abiotic factors contribute towards growth and yield losses of this crop. Arsenic (As) toxicity is a global issue, but it is particularly critical in developing countries. The current study was designed to evaluate the efficacy of selenium (Se) in mitigating the toxic effects of As in two varieties (HSP-181 A and PS09979325) of Capsicum annuum L. Different concentrations of As (0, 50, and 100 µM) and Se (0, 5, and 10 µM) were tested using 14 days old seedlings of C. annuum L. The As stress caused a significant (P ≤ 0.001) reduction in growth, uptake of nutrients, and eco-physiological attributes in both varieties however, the response was specific. While the overproduction of osmo-protectants and antioxidants intensified the symptoms of oxidative stress. The maximum reduction in shoot length (45%), fresh weight (29%), and dry weight (36%) was observed in under 100 µM As stress. The organic acids exudation from the roots of both cultivars were significantly increased with the increase in As toxicity. The Se treatment significantly (p ≤ 0.001) improved growth, nutrient uptake, gas exchange attributes, antioxidant production, while decreased oxidative stress indicators, and As uptake in the roots and shoots of all the subjects under investigation. It is concluded from the results of this study that Se application increased photosynthetic efficiency and antioxidant activity while decreasing As levels, organic acid exudation, and oxidative stress indicators in plants. Overall, the var. PS09979325 performed better and may be a good candidate for future pepper breeding program.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
The As is a well-documented carcinogenic metalloid, its contamination across the board is hazardous for agriculture, and all kinds of living organisms worldwide [1]. It alters the morphological, biochemical, and physiological processes in plants even at low concentrations (range 20–100 µM), particularly those that are consumed by humans [2,3,4]. Various human endeavours, such as mining or naturally occurring chemical processes are increasing As concentration in soil (1–40 mg kg-1). In soil, drinking water (6–12 ng ml-1), and air (3–30 ng m-3), it can be found in four different oxidation states i.e. -III, 0, +III, and + V [5]. As (III) and As (V), as well as their interconversion, cause oxidative stress by producing reactive oxygen species (ROS). This affects the regulatory control of a variety of metabolic activities [6].
According to a recent investigation, As contamination has also been found in Hungary, Mexico, Argentina, Australia, and the United States, among other areas. It is well known that South-East Asia suffers from groundwater pollution [7]. The As may cause severe devastation to plants and animals in various parts of Pakistan [8]. A high As concentration in both surface and groundwater has been recorded in different regions of Punjab and Sindh province of Pakistan [9]. However, a number of variables, including soil type, plant species, and uptake processes, significantly impact the pace of As absorption and accumulation [10].
Vegetables are regarded as the best diet for promoting health and preventing disease because they contain essential nutrients for body repair and growth. Vegetables, with their high vitamin and mineral content, help the body maintain its alkaline reserves [11]. C. annuum L., sometimes known as the bell pepper, hot pepper, sweet pepper, and chilli, is a well-known vegetable crop. It belongs to the Solanaceae family and is consumed as a pickle, sauce, spice, and vegetable. All over the world, chilies are utilized as a spice and have also found their way into foods like beverages and medicines [12]. The As contents in vegetables grown in soils with high As extractability are more likely to be higher than soils with low As extractability [13].
Developing effective strategies to address pollution is critical because it poses a serious risk to agricultural production globally [14]. Various remediation techniques reduce As accumulation in vegetables, including soil replacement, leaching, electrokinetic remediation, phytoremediation, flood culture, organic additions, intercropping, and breeding for low As accumulating genotypes [15]. Another environmentally friendly strategy for reducing negative impacts of As is the exogenous use of ions and essential micronutrients like Se. The Se was once believed to be harmful, but it is now understood to be an essential nutrient for plants. Exogenous application of Se reduces plant damage caused by various environmental adversities either used as topically or as a root zone treatment [16]. Heavy metal absorption and translocation such as Cd, Cr, Pb, As, Cu, Hg and others is also significantly declined [17]. By preserving cell membrane stability, mineral absorption, and cellular processes, as well as minimising oxidative stress damage, the Se treatment decrease toxicity of heavy metal [18]. The objective of the current study was to examine the protective role of Se under As stress and to ascertain the potential of the Capsicum annuum L. cultivars employed in the study to withstand As stress. Because of their diverse parentage, both cultivars have genetic differences that could explain this differential behaviour towards As tolerance. The As tolerance and translocation in bell pepper under Se treatment described here will contribute to the existing body of knowledge for bell pepper and will be useful in selecting cultivars for future variety development programmes.
Materials and methods
Acid washed sand (pH 5.5-6) filled pots (40 cm in height and a diameter of 34 cm) with established volumetric field capacity 0.072 m3/m3 were used for C. annuum L. (HSP-181 A and PS09979325) seed sowing. Karam Ceramics Limited in Karachi supplied the acid-washed sand. The day/night temperature was 25 ± 1/15 ± 1 °C. Ali’s method [19] was used to prepare the nutrition solution. The plants were irrigated with Hoagland solution. Young seedlings from nursery were shifted to each sand filled pot and every pot received 500 mL of nutritional solution throughout the early stages of development, and this amount was subsequently increased to 1000 mL at alternate days. Five plants were maintained in each pot and 14 days old seedlings were subjected to different treatments. The As (50 and 100 µM) stress was maintained for two weeks then Se (5 and 10 µM) was applied using nutrient solution for two weeks and plants were harvested at four leaf stage for further analyses.
The plants were classified into the following categories
Control (nutrient solution alone), control with 5 µM Se, control with 10 µM Se, As (50 µM), As (100 µM), As (50 µM) with Se (5 µM), As (100 µM) with Se (5 µM), As (50 µM) with Se (10 µM), and As (100 µM) with Se (10 µM). Preliminary tests employing a variety of As concentrations, (0, 10, 20, 30, 40, 60, 70, 80, 90, 100 µM), were used to determine effective levels of As (unpublished data). Similarly, the Se levels (5, 10 µM) were determined in a preliminary investigation in which three-week-old bell pepper seedlings were watered using a nutrient solution containing different levels (0, 1, 3, 5, 7, 10, 12 µM) of Se. Sodium selenate (Na2SeO4) and meta-arsenate (Na2HAsO4.7H2O) were purchased from Sigma-Aldrich (USA). Completely randomized design was followed in all experiments with three replicates.
Root shoot length
After uprooting the plant from pots, roots and shoots were carefully separated. A meter rod was used to measure the root and shoot length of the harvested plants.
Root shoot fresh and dry weight
After harvesting, the root and shoot fresh weight was determined using electrical weight balance. For dry weight measurement, the samples were dried at 75 °C for 48 h using laboratory oven.
Photosynthetic pigments
A leaf sample of 0.5 g was homogenized in 10 mL of 90% acetone at 4 °C. Supernatant from each sample was obtained after centrifugation at 12,000 rpm for 5 min. Absorbance was recorded using a UV-Spectrophotometer (Hitachi U-2001, Japan) at 480, 645, and 663 nm. Calculations of photosynthetic pigments were done using [20] method.
Estimation of enzymatic antioxidants
A leaf sample weighing 0.5 g was taken, ground in liquid nitrogen, and then mixed with EDTA (0.5 mM) and NaCl (0.2 mM) in 5 mL of sodium phosphate buffer (50 mM, pH 7.00). The resulting homogenate underwent a centrifugation process at 12,000 × rpm for 15 min at 4 °C. The activity of the enzymes, superoxide dismutase (SOD) and peroxidase (POD) was assessed using the collected supernatant.
A reaction mixture of 3 mL containing 50 mM sodium phosphate buffer (pH 7.0), 55 mM NBT (nitro blue tetrazolium), 10 mM methionine and 100 µL enzyme extract prepared to calculate the SOD activity. Then guaiacol was employed as the substrate to measure POD activity according to methodology reported [21]. The absorbance of SOD and POD was noted using a UV-visible spectrophotometer (Hitachi U-2001, Japan) at 560 and 470 nm, respectively.
For CAT activity was estimated by methodology reported by Hatch [22] by maintaining H2O2 concentration in a reaction mixture as reported, and absorbance changes were measured at 240 nm. A 100 µL of enzyme extract, 100 µL of 250 mM H2O2, 2.5 mL of 50 mM phosphate buffer, and 2 mM EDTA (pH 7.0) made up the reaction mixture. The activity of APX was assessed using the method described by Asada and Takahashi [23]. A 100 µL of enzyme extract, 100 µL of 7.0 mM ascorbate, 100 µL of 250 mM H2O2, and 2.5 mL of 25 mM potassium phosphate buffer with 2 mM EDTA (pH 7.00) made up the reaction mixture. At 290 nm, the absorbance was measured. The unit of measurement for enzyme activity was mg-1 protein.
Non-enzymatic antioxidants and oxidative stress indicators estimation
An ethanol extract was made to assess non-enzymatic antioxidants and osmolytes. After being homogenized in 10 mL of 80% ethanol, a 50 g sample of air-dried leaves was filtered through Whatman filter paper No. 41. Once more extracting the filtrate residues, the filtrate was mixed to create a final amount of 20 mL.
Estimation of phenolic contents
Julkunen-Tiitto’s [24] method was used with little modification for estimation of total phenolic contents. In this procedure, 2 ml of 80% acetone was used to homogenize 0.5 g of fresh leaf material. The homogenate was centrifuged at 12,000 × g for 10 min. After separating the supernatant (100 l), 1 ml of the Folin-Ciocalteu phenol reagent and 2 ml of distilled water were added. Following this, 3 ml of 20% N2CO3 was added before the final volume of 10 ml of distilled water was added. It was vigorously Shaked. To measure absorbance at 750 nm, a UV-vis spectrophotometer (Hitachi U-2001, Japan) was used.
Determination of anthocyanin content
Using methods described by Hodges & Nozzolillo, anthocyanin content was measured. In this protocol, Fresh leaves (0.1 g) are ground in 2 ml of acidic methanol (Methanol + 1% HCL). After completion of a homogenous mixture of samples, transferred it to the test tubes. Then tubes were boiled at 100 oC for 30 min. Centrifugation at 12,000 × sample (g) for 15 min. Absorbance was observed using a spectrophotometer at 536 nm and 600 nm [25].
Determination of flavonoids content
The flavonoid contents were determined by the method of Karadeniz [26]. The 0.1 g sample of fresh plant leaves grinding was performed in 80% acetone. Add 0.3 ml of 5% NaNO2 and 3 ml of distilled water to the clean plant leaf extract. The reaction mixture was then mixed, and it was left to reset for five minutes at room temperature. After that, add 0.6 ml of 10% AlCl3 and 2 ml of a solution of 1 M NaOH to the reaction mixture. distilled water was added to the mixture to get the volume up to 10 milliliters. Uv-Vis spectrophotometer was used to measure absorbance at 510 nm (Hitachi U-2001, Japan).
Reducing sugars estimation
Dinitro alicyclic acid (DNS) reagent was used for the determination of reducing sugars. For the preparation of reagent Wood and Bhat (1988) method was used [27].
Proline contents
Proline contents were estimated by following procedure [28]. A sample of 0.2 g fresh leaves was grinded in 8 mL of 30% aqueous sulphosalicylic acid. A new test tube was filled with 1 mL of supernatant after the lysate had been centrifuged for 10 min at 12,000 × rpm. This tube was filled with 1 mL of acidic ninhydrin and 1 mL of glacial acetic acid, heated for 10 min at 100 ℃, and then cooled in the ice bath. Following the addition of 5 mL of toluene, the test tube was vortexed for 30 s and then cooled. To measure the absorbance, a UV-visible spectrophotometer was used at 520 nm (UV-2550; Shimadzu, Kyoto, Japan). Using a standard curve, the amount of proline was determined and expressed in µmol g-1 FW.
MDA estimation
Using a modified version of procedure, the degree of lipid peroxidation, the MDA content was calculated [29]. In 20 mL of 50 mM phosphate buffer with a pH of 7.8, A frozen leaf sample of 2 g was homogenized. The resultant homogenate was centrifuged at 4 ℃ and 12,000 × rpm for 10 min. Supernatant was collected and mixed 5 mL supernatant with 1.5 mL of 0.5% TBA. The resultant mixture was chilled in an ice bath after being heated for 20 min at 95 ℃ in water bath. Using a spectrophotometer, the supernatant absorbance was recorded at 532, 600, and 440 nm wavelengths (UV-2550; Shimadzu, Kyoto, Japan). The MDA content was estimated as follows and expressed as nmol g-1 FW.
H2O2 content estimation
For H2O2 estimation, in 20% H2SO4, 3 mL of the leaf extract mixture and 1 mL of titanium sulphate were added. For 10 min, this reaction mixture was centrifuged at 8000 rpm. Supernatant was employed at a wavelength of 410 nm to measure absorbance. The H2O2 concentration was calculated by an extinction coefficient of 0.28-1 cm-1 and was expressed as µmolg-1 FW [30].
Electrolyte leakage
Electrolyte leakage (EL) was determined following [31] procedure using a fully stretched top second leaf. 10 mL of distilled water and the leaves were put in a test tube after being uniformly chopped into 5 mm pieces. The first electrical conductivity (EC1) was determined after two hours of incubation. The samples were autoclaved at 121 ℃ for 15 min before being chilled to 27 ℃ for evaluating their final electrical conductivity (EC2).
Gas exchange characteristics
The net photosynthetic rate (Pn), stomatal conductance (gs), and transpiration rate (E) of the second youngest leaf from the top of each plant were measured using the photosynthesis measuring-system CI-340 portable infrared gas analyzer (Analytical Development Company, Hoddesdon, USA). All these parameters were recorded from 9:00 to 11:00 AM. Atmospheric pressure was approximately 99.9 kPa, water vapors pressure ranged from 6.0 to 8.9 mbar, maximum PAR at leaf surface was up to 1711 molm-2 s-1, leaf temperature was between 28.4 and 32.4 ℃, ambient temperature was between 22.4 and 27.9 ℃. Three plants from each replicate of each treatment were used to measure the said parameters.
Water use efficiency
The formula for calculating water use efficiency was (Pn/E), where Pn = net photosynthetic rate.
Nutrient analysis
Digestion
For sample digestion, the Wolf (1982) method was used, and 5 mL of concentrated H2SO4 and 0.5 g of dried, material was introduced to the digestion tube. The extract was used to measure potassium (K), calcium (Ca), and phosphorous (P) after being filtered.
Estimation of K+
Using a flame photometer (Jenway PFP 7), potassium (K+) was measured.
Determination of P
A spectrophotometer was used to measure the amount of phosphorus [32].
Calcium (ca) and magnesium (mg) determination
Using an atomic absorption spectrophotometer (Hitachi Z-2000), calcium (Ca) and magnesium (Mg) concentrations were determined.
Root organic acid estimation
Using high-performance liquid chromatography (HPLC), organic acids were measured (PerkinElmer, MA, USA). The procedure developed by [33] was used to analyze the root organic acids. Briefly, three plants each, from different treatments were washed with tap water and transferred to a rhizo box-like system containing 200 g autoclaved humid soil. A nylon net was used in the rhizoboxes to avoid root penetration into the soil. After 36 h, plants were taken out of the rhizoboxes and rinsed with10 ml of double distilled water for 30s. The collected exudates were passed through 0.45 mm syringe filters. (BioRad USA) and poured into 15 ml falcon tubes. For organic acid analysis, samples were mixed with 0.01 M NaOH. The root exudates were stored at -80 °C for further analysis.
As level estimation in roots and shoots
To estimate the amount of As present in the root and shoot, the obtained samples were dried in an oven (Esco OFA-32-8 Singapore) at 70 ℃ for 24 h while ash was produced in a muffle furnace at 550 ℃ for 20 h. It received a treatment of 17.5% (v/v) H2O2 and 31% (m/v) HNO3 at 70 ℃ for two hrs before being given distilled water. To estimate As contents, an atomic absorption spectrophotometer (Hitachi Z-2000) was employed.
Statistical analysis
Using CoStat software version 6.303, a three-way analysis of variance (ANOVA) was conducted using the LSD test at p < 0.05 to measure differences in variables among the various treatments. The LSD test was then used to compare treatment means. Using R Studio, the principal component analysis and Pearson’s correlation were also computed.
Results
Different biotic and abiotic stresses tend to reduce plant growth and metabolism. Different metabolic changes take place when plants encounter stress conditions, and these alter the expression of different genes relative to production of different chemicals beneficial for the plant’s health. Stress reduced the plant growth by reducing root and shoot length significantly as reported in the results. It also increased the H2O2 and MDA content which is indicator that the plant is undergoing stress. Different types of root exudates were released to reduce metal uptake shows the plant capability to defend themselves against the stress conditions. But plant’s simple own defense mechanism can’t mitigate the stress conditions that’s why Se have been reported to increase the secondary metabolite production like flavonoids and anthocyanins which have been reported to increase plants defense mechanism. Similar results are presented in next sections showing increased plant defense under Se application.
Growth parameters
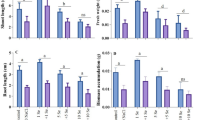
The findings of this work demonstrated that As toxicity significantly (p < 0.001) reduced growth attributes of both bell pepper varieties. Decrease in growth parameter was observed which might be due to many different defense mechanisms like ROS scavenging and osmolyte disturbance. The most notable reduction in shoot length (27%) fresh weight (13%) and dry weight (18%) was recorded in var. HSP-181 A under 50 µM As stress as compared to control. Under 100 µM As stress, the same cultivar experienced a maximum reduction in shoot length (45%), fresh weight (29%), and dry weight (36%). The var. PS09979325 showed the most minimal drop in shoot length (8 and 18%), fresh weight (7 and 12%), and dry weight (6 and 10%), respectively, under 50 µM and 100 µM As stress.
Under control conditions, the shoot length, shoot dry weight, and shoot fresh weight of cv. HSP-181 A increased up to 13, 7, 14, 25, 23, and 36%, respectively, following the application of Se (5 µM and 10 µM) in nutrition media. While greater increases in shoot length (5 and 33%), shoot fresh weight (7 and 25%), and shoot dry weight (8 and 26%) were observed in cv. HSP-181 A by Se application (5 µM) under As stress (50 µM, 100 µM), and maximum increases in shoot length (14%) were observed in variety HSP-181 A by Se application (10 M) under As stress (Fig. 1).

Effect of Se on growth attributes of bell pepper (Capsicum annuum L.) varieties under As stress. Shoot length (a), root length (b), shoot fresh weight (c), shoot dry weight (d), root fresh weight (e) and root dry weight (f). T0 = Control, T1 = control + Se (5µM), T2 = control + Se (10µM), T3 = As (50µM), T4 = As (50µM) + Se (5µM), T5 = As (50µM) + Se (10µM), T6 = As (100µM), T7 = As (100µM) + Se (5µM), T8 = As (100µM) + Se (10µ)
Under As stress, both vars. of bell pepper experienced significant (p < 0.01) reductions in root length, dry weight, and fresh weight. However, the response varied between the varieties. As compared with control, the var. HSP-181 A experienced the greatest decrease in root length (27%), fresh weight (18%), and dried root weight (25%). When plants were treated with 100 µM As, maximum drop in root length (45%), fresh weight (44%), and dry weight (36%) was observed. Under untreated control, the Se application (5 µM and µ10 M) enhanced root length (13% and 25%), root fresh weight (25 and 52%), and root dry weight (4 and 28%), respectively. When applied to HSP-181 A under As (50 µM and 100 µM) stress, 5 µM Se increased root length (5 and 33%), fresh weight (5 and 27%), and dry weight (6 and 29%), respectively. In the same cultivar, 10 µM Se also enhanced root length (14 and 34%), fresh weight (11 and 37%), and dry weight (21 and 32%), respectively, under As (50 and 100 µM) stress. So, based on our findings, plants produced more biomass when Se was applied under As stress (Fig. 1).
Pigments and photosynthetic parameters concentrations
Figure 2 depicts the effects of As stress with or without the application of Se on the contents of Chl a, b, Total Chl., and Car in both vars. of bell pepper. Under As (50 µM) stress, Chl a (27%), b (20%), T. Chl (24%) and Car. (17%) contents decreased significantly (p < 0.01) in the leaves of the HSP-181 A. However, at 100 µM As stress, the var. HSP-181 A showed maximum drop in Chl a (47%) Chl b (35%) Total Chl (42%) and Car (32%). Furthermore, by applying Se 5 µM under control conditions, Chl a, b, Total Chl, and Car levels increased up to 3, 5, 4, and 7%, respectively in this variety. While application of 10 µM Se increased these contents up to 15, 30, 22, and 56% in the same variety under same conditions. When Se was applied (5 µM) under As stress of 50 µM and 100 µM, a greater increase in Chl a (5 and 31%), Chl b (7 and 24%), T. Chl (6 and 28%), and carotenoids (5 and 19%) was observed in the var. HSP-181 A (Fig. 2).

Effect of Se on pigments and photosynthesis attributes of bell pepper (Capsicum annuum L.) varieties under As stress. Chlorophyll a (a), chlorophyll b (b), total chlorophyll (c), carotenoids (d), net photosynthetic rate (e), stomatal conductance (f), transpiration rate (g) and water use efficiency (h). T0 = Control, T1 = control + Se (5µM), T2 = control + Se (10µM), T3 = As (50µM), T4 = As (50µM) + Se (5µM), T5 = As (50µM) + Se (10µM), T6 = As (100µM), T7 = As (100µM) + Se (5µM), T8 = As (100µM) + Se (10µM)
Under As stress, gaseous exchange attributes were significantly decreased (p < 0.01) as compared with control. The var. HSP-181 A under As stress (50 µM) demonstrated reductions in Pn (23%), gs (60%), E (30%), and WUE (7%). The greatest decreases in Pn (33%), gs (80%), E (48%), and WUE (16%) were observed when plants were exposed to nutritional solution containing 100 µM As. Additionally, under control conditions, with Se application of 5 µM, the var. HSP-181 A exhibited an increase in Pn, gs, E and WUE values up to 19, 17, 9, and 10% while 10 µM increased these attributes up to 35, 38, 28, and 28 respectively. With 5 µM Se application under As stress (50 and 100 µM), var. HSP-181 A showed an increase in Pn (14 and 16%), WUE (1 and 12%), gs (18 and 51%), and E (15 and 10%), respectively. Additionally, 10 µM Se treatment increased Pn (21 and 28%), gs (37 and 70%), E (20 and 33%), and WUE (9 and 17%) under 50 µM and 100 µM As stress, respectively (Fig. 2).
Antioxidants, osmolytes and stress indicators profile
In the present investigation, enzymatic antioxidant activity increased significantly (p < 0.01) under As exposure in both bell pepper varieties. When var. HSP-181 A was exposed to As stress (50 µM), the greatest increase in SOD (26%), POD (53%), CAT (51%), and APX (68%) was recorded as compared to As non-treated plants. But under 100 µM As stress, var. HSP-181 A showed the highest increases in SOD (43%), POD (61%), CAT (57%), and APX (77%). In addition, SOD, POD, CAT and APX in cv. HSP-181 A increased under control conditions by Se application (5 µM and 10 µM) up to 3, 39, 28, 42% and 12, 67, 69, 62%, respectively. At the same time, a greater increase in SOD (51 and 60%), POD (53 and 58%), CAT (51 and 57%), and APX (71 and 81%) was recorded in var. HSP-181 A by 5 µM Se application under As stress (50 and 100 µM) as well as SOD (77 and 67%), POD (55 and 61%), CAT (52 and 59%) and APX (74 and 86%) were increased in var. HSP-181 A with Se application (10 µM) under As 50 and 100 µM stress (Fig. 3).

Effect of Se on enzymatic antioxidants of bell pepper (Capsicum annuum L.) varieties under As stress. SOD (a), POD (b), CAT (c) and APX (d). T0 = Control, T1 = control + Se (5µM), T2 = control + Se (10µM), T3 = As (50µM), T4 = As (50µM) + Se (5µM), T5 = As (50µM) + Se (10µM), T6 = As (100µM), T7 = As (100µM) + Se (5µM), T8 = As (100µM) + Se (10µM)
The As stress significantly (p < 0.01) enhanced the phenolics, flavonoids, MDA, and anthocyanin contents in both bell pepper varieties. The maximum accumulation of phenolics (62%), flavonoids (65%), MDA (6%), and anthocyanin (52%) contents was recorded in var. HSP-181 A under 50 µM As stress. However, under 100 µM As stress, a maximum increase in phenolics (73%), flavonoids (71%), MDA (27%), and anthocyanin (72%) contents was observed in var. HSP-181 A. In addition, maximum increase in phenolics, flavonoids, and anthocyanin contents of var. HSP-181 A was recorded under 10 µM Se applications. MDA content decreased with 5 and 10 µM Se application by 16 and 29% respectively under control condition. At the same time, more increases in phenolics (72 and 60%), flavonoids (53 and 78%), and anthocyanin (51 and 77%) were recorded in var. HSP-181 A with Se application (5 µM) under As stress (50 and 100 µM) and more decrease in MDA contents were observed in HSP-181 A with Se application (5 µM) under As stress (50 and 100 µM) by 8 and 18% respectively. Moreover, a maximum increase in phenolics (79 and 67%), flavonoids (58 and 81%), and anthocyanin (55 and 80%) contents were observed, more decrease in MDA contents (12 and 25%) were observed in var. HSP-181 A with Se application (10 µM) under As (50 µM and 100 µM) stress, respectively (Fig. 4).

Effect of Se on non-enzymatic antioxidants and oxidative damage indicators of bell pepper (Capsicum annuum L.) varieties under As stress. Phenolics (a), flavonoids (b), anthocyanin (c) reducing sugar (d), proline (e), MDA (f), H2O2 (g) and electrolyte leakage (h). T0 = Control, T1 = control + Se (5µM), T2 = control + Se (10µM), T3 = As (50µM), T4 = As (50µM) + Se (5µM), T5 = As (50µM) + Se (10µM), T6 = As (100µM), T7 = As (100µM) + Se (5µM), T8 = As (100µM) + Se (10µM)
Ion concentrations
In the present study Ca2+, Mg2+, K+ and P were decreased significantly (p < 0.001) in both bell pepper varieties under stress. The maximum adverse effect of As (50 µM) was exhibited by var. HSP-181 A in Ca2+ (28%), Mg2+ (29%), K+ (16%) and P (23%) contents under control condition. Maximum reduction in Ca2+ (46%), Mg2+ (46%) K+ (53%) and P (37%) were observed in var. HSP-181 A under 100 µM As stress. In addition, Ca2+, Mg2+, K+, P in var. HSP-181 A increased under control conditions with Se (5 and 10µM) application up to 17, 20, 9, and 7%, and 35, 38, 38, and 23%, respectively. At the same time, a higher increase in Ca2+ (13% and 9%), Mg2+ (13 and 34%) K+ (2 and 36%) and P (3 and 25%) was recorded in var. HSP-181 A with Se application (5 µM) under As stress (50 and 100µM) level as well as Ca2+ (18 and 31%), Mg2+ (17 and 40%), K+ (6 and 49%), and P (11 and 32%) was increased maximum in var. HSP-181 A with Se (10 µM) application under 50 and 100 µM As stress (Fig. 5).

Effect of Se on ionic contents and organic acids of bell pepper (Capsicum annuum L.) varieties under As stress. Calcium (a), magnesium (b), potassium (c), phosphorus (d), malic acid (e), citric acid (f), acetic acid (g) and oxalic acid (h). T0 = Control, T1 = control + Se (5µM), T2 = control + Se (10µM), T3 = As (50µM), T4 = As (50µM) + Se (5µM), T5 = As (50µM) + Se (10µM), T6 = As (100µM), T7 = As (100µM) + Se (5µM), T8 = As (100µM) + Se (10µM)
Root exudates profile
Analysis of variance revealed that malic acid, citric acid, acetic acid, and oxalic acid significantly (p < 0.01) increased. The adverse effect of As (50 µM) was recorded in var. HSP-181 A for malic acid (38%), citric acid (21%), acetic acid (23%), and oxalic acid (32%) contents increment as compared to As non-treated plants. While under 100 µM As stress, maximum increment in malic acid (40%), citric acid (35%), acetic acid (46%), and oxalic acid (38%) was observed in var. HSP-181 A as compared to control. A maximum increase in malic acid (5 and 50%), citric acid (14 and 27%), acetic acid (17 and 37%), and oxalic acid (12 and 28%) was observed in var. HSP-181 A with Se application (10 µM) under 50 µM and 100 µM As stress, respectively (Fig. 5).
Our result revealed that As in root and shoot increased substantially (p < 0.01) in both bell pepper varieties under As treatment. A maximum increment in As levels in root and shoot were observed in var. HSP-181 A under 100 µM As stress. In addition, As contents in root and shoot increased under control conditions with Se application (5 and 10 µM) up to 5, 32, and 13, 28%, respectively. While the greater decrease in As contents of root and shoot were recorded in var. HSP-181 A with Se application (5 µM) under both levels of As stress. A maximum reduction in As contents of root/shoot was recorded in var. HSP-181 A with 10 µM Se application under 50 and 100 µM As stress (Table 1).
Plant development, biomass, photosynthetic pigments, gaseous exchange characteristics, antioxidants, root exudates, and mineral intake were correlated, according to PCA analysis. Figure 6 describes that Mg, WUE, P, Ca, K, RDW, RFW, SDW, SFW, SL, RL, Chl. a, Chl. b, T. Chl, Car, Elec, and H2O2 negatively correlate with MDA, Ma, As R, Oxa, Ace, As S, Cit, POD, CAT, APX, SOD, Anth, FL, RS, Pro, and Phe. However, Mg, WUE, and P are closely positively correlated. Oxa, As S, and Ace are closely positively correlated. POD, CAT, and Anth are also closely positively correlated. Among the extracted components, the significant contribution of Dim 1 (66.5%) was followed by Dim 2 (13.8%), with a cumulative donation of 80.3%. Pearson’s correlation shows the relationship between plant growth, biomass, photosynthetic pigments, gaseous exchange attributes, antioxidants, root Exudates, and mineral intake (Fig. 6). MDA has a highly negative correlation with SL, SFW, SDW, RL, RFW, RDW, Chl. b, Chl. a, T. Chl, Car, RS, Pn, C, E, WUE, Ca, Mg, K, P strongly correlates with SOD, POD, CAT, Pro, Elec, H2O2, and Anth. Ma, Cit, Ace, As R, and As S has a highly negative correlation with SL, SFW, SDW, RL, RFW, RDW, and Chl. b, Chl. a, T. Chl, Car, Pn, C, E, WUE, Ca, Mg, K, P, and it has a highly positive correlation with MDA. POD and CAT negatively correlate with SL, SFW, SDW, RL, RFW, RDW, and Chl. b, Chl. a, T. Chl, Car, Pn, C, E, WUE, Ca, Mg, K, P strongly correlates positively with SOD, CAT, APX, Phe, FL, and Anth.

Principal component and correlation analysis of the all studied attributes of bell pepper (Capsicum annuum L.) varieties under As stress. Different abbreviations used in this figure are as follow: RL, root length; SL, shoot length; SFW, shoot fresh weight; SDW, shoot dry weight; RFW, root fresh weight; RDW, root dry weight; Chl. a, chlorophyll a; Chl. b, chlorophyll b; T. Chl, total chlorophyll; Car, carotenoids; SOD, superoxide dismutase; POD, peroxidase activity ; CAT, catalase ; APX, ascorbate peroxidase; MDA, malondialdehyde content; Anth, anthocyanin, FL, flavonoids; Phe, phenolics; H2O2, hydrogen peroxide; Pro, proline, Elec, electrolyte leakage; RS, reducing sugar; Ca, calcium; Mg, magnesium; K, potassium; P, phosphorus; Ma, malic acid; Cit, citric acid; Ace, acetic acid, Oxa, oxalic acid; As R, arsenic contents in roots; As S, arsenic contents in shoots
Discussion
Like all other heavy/toxic metals, when discharged into the environment As is hazardous to living things being incorporated and accumulated through food chain [34, 35]. The first tissue to come into contact with As from polluted soil is the root system. It inhibits root development and extension [36] by delaying or preventing cell development and biomass creation as it passes through shoots and may significantly limit plant growth [37]. Numerous chemicals are used to combat heavy metal toxicity. Due to its metabolic functions in the living system, Se application has become one of the most efficient methods for reducing heavy metal toxicity [38]. By using it, heavy metals are restricted from being absorbed by roots and passed on to shoots. Its supplementation is expected to increase the quantity of pectin as well as the cell wall thickness enhancing plant growth [39]. In the present study root and shoot biomass was significantly reduced in both bell pepper varieties under As stress although the observed biomass reduction was dose and cultivar specific. The var. HSP-181 A was found to be more vulnerable to As stress while Se application significantly restored the development attributes in both varieties under investigation.
A considerable decrease in the production of chlorophyll and carotenoids has been recorded in the present study possibly due to disagreement between the adaptive alterations in photosystems II and I under elevated As stress as has been reported previously [40]. Another explanation for the decreased pigment level could be that the As stress prevents the absorption of N and Mg, two crucial elements of the chlorophyll molecule [41]. In the current study, the same reduced uptake behaviour was observed for both Mg and N, resulting in reduced chlorophyll content. As a result of severe As stress, the external supply of Se protects chloroplasts and aids in the maintenance of photosynthetic pigments. The findings of the current investigation showed that Se application considerably boosted the bell pepper’s pigment content as previously reported in Brassica juncea [42]. The As is absorbed by plants and interacts with different metabolic pathways to produce peroxides thus increasing ROS levels in plant cells. According to [38], ROS production creates enzymatic antioxidants under stress conditions. These antioxidants are regarded as the first line of defence since they scavenge ROS from various cellular compartments [43]. In contrast to As-stressed plants, the current study showed that Se treatments boosted SOD, POD, CAT, and APX activity in bell pepper and similar findings has been reported in past [44]. The SOD oversees neutralising ROS and changing O2 into H2O2. Later, utilising ascorbate, POD and CAT scavenge the H2O2 produced by dismutation and transform it into H2O and O2 [45]. The enzyme APX, which is a part of the (ASA)-Glu cycle, inhibits the buildup of H2O2 in tissue by converting it to H2O. Several enzymes may function individually or in groups via crosstalk, as may be appropriate in different plant species, to prevent oxidative damage and toxicity inside plants [42].
In response to As stress, the anthocyanin contents increased. Anthocyanin synthesis under stress may result from glutathione-S-transferase activation, whereas violaxanthin, a xanthophyll, serves as a precursor for the manufacture of abscisic acid, It is also connected with the protection of oxidative stress [46]. Numerous investigations have shown that the enzyme’s gene expression in anthocyanin synthesis pathways that scavenge reactive oxygen species is closely related to anthocyanin accumulation by Se administration [47]. The flavonoids content increased during the early stages of As stress; it is also regarded as a polyphenolic molecule with antioxidant properties. As stress was followed by an increase in radical scavenging, demonstrating the beneficial function flavonoids play in removing free radicals [48]. Phenolics have hydroxyl and carboxyl groups with metal-binding properties. Plant phenolics also act as antioxidants. The high phenolic content in this study may have performed a strong antioxidant effect. According to previous studies, use of Se in plants under Stress boosted the activity of enzymes involved in the phenylpropanoid production pathway [49, 50] and same is thought to be happened in the present study however, the varietal differences may be due to variation in level of genes expression for candidate enzymes of the metabolic process.
The findings of the present study revealed that as As concentration increased in various plant tissues, oxidative damage as indicated by MDA level, also increased. As a result, Se accumulation’s impact on oxidative damage because of which oxidative stress increased. The supply of Se thereby mitigated oxidative harm because the MDA level in the Se application was equivalent to that of the non-stressed plants. In previous study [51] it has been found that Se can potentially deactivate ROS produced by As toxicity in oilseed rape.
The increase in the concentration of As in the solution of nutrients resulted in reduction of gaseous exchange attributes. However, the main effect of As stress is that it hinders the electron transport chain (ETC), reduces the photosynthesis rate, and causes ultrastructure disruption of chloroplast [52]. As metal reduce produce secondary metabolite which destroy the PS-II and PS-I structure and reduce the plant growth and defense mechanism [53]. As inhibits key enzymes of ETC like cytochrome c oxidase and thus disrupting plant metabolism [53]. One of the initial coping mechanisms of stressed plants is closure of stomata, which reduces leaf transpiration and, as a result, the regular CO2 flow into the carboxylation site. The principal factor decreasing photosynthesis in plants grown under stress is stomatal conductance [54]. Additionally, In the respiratory chain, Se increased respiration and electron transport, which improved chlorophyll biosynthesis. By boosting chlorophyll content and preserving chloroplast ultrastructure, the Se treatment may help stressed plants recover their ability to photosynthesize [55]. In the current investigation, the exact same response was noted in both varieties of bell pepper.
Leaf Ca2+, Mg2+, K+, and P contents in bell pepper varieties were decreased with increasing As stress level in our study because stress reduced mineral nutrient retention, as is evident in previous study [56]. Such consequences could result from increased stress accumulation, which displaces mineral nutrients from binding sites, limiting their translocation and uptake [57]. Under stress, for example, a decrease in K+ concentration may be related to xylem obstruction and suppression of root growth of plant [58]. A similar result shows that the As stress lowered plant Ca2+ levels resulting in poor rice growth [58]. Metalloid stress reduced uptake of Mg2+ in tomato shoots and roots, decreasing chlorophyll production [59]. For instance, [39] reported the beneficial effect of Se treatment on ion homeostasis in Chinese cabbage under heavy metal stress.
The As significantly boosted the root exudation of citric, acetic, maleic, and oxalic acids in the current study. The metabolism of root cells, which is related to the buildup and release of different organic acids, is disrupted by toxic chemicals. As observed in Solanum nigrum L. the current study showed the exudation of various organic acids from stressed plant roots, which may be crucial in lowering As toxicity through metal detoxification, nutrient stabilisation, and improved plant growth [33]. According to prior research on poplar roots under metal stress, organic acid exudation occurs via anion channels [60], and their release ought to be balanced by the efflux of cations/protons [61].
Applying Se under As stress increased the metal content of plant organs, increased mineral uptake, and impacted the exudation of organic acids from plant roots [62]. Despite the genetic variations among the species, our study revealed the same results in bell pepper varieties under investigation. This study found that 100 µM As caused greater As accumulation in the roots. Ions are transported across the cell membrane by transporters, which are unique proteins. Only a small part of the total ions reaches the plant root. The majority of such ions are physically takes in by the negative-charged -COO compartment, which oversees cell wall surface absorption. This compartment prevents ions from entering the cell in the plant shoot [63]. The As competes for transport channel and gets stored in different non-essential parts disturbing the osmotic conditions in the plant. It disrupts normal metabolic machinery and competitively reduce the other ions concentrations as reflected by the results presented in the study [64]. The competition might be between specific and non-specific transporters contributing towards the stress symptoms appearance [65]. These substances build up in cellular vacuoles and become more concentrated, which inhibits them from moving to the shoot. Because of this, the level of this element has a greater impact on the root than the shoot [66]. All these components are accumulated in the root and shoot, Se decreased the deleterious impact of As stress [63].
Conclusions
The As had a significant impact on plant mineral uptake, biomass accumulation, photosynthesis, and the antioxidant profile of both cultivars (HSP-181 A and PS09979325). The As stress had significantly reduced the physicochemical and biochemical parameters by changing secondary metabolism, disrupting the normal osmotic balance, reducing ions exchange, uptake and translocating. The Se application proved to have positive effect on reducing stress conditions caused by As stress and improved plant growth and all physiological and biochemical parameters. According to the findings of the present study, it is proposed that var. PS09979325 responded more effectively overall under As stress as compared to var. HSP-181 A. The organic acid exudations, oxidative stress markers, and As levels in plant organs also increased as the toxicity level did. The Se supplementation, however, promoted greater plant growth and biomass, decreased the production of ROS, maintained minerals, and decreased the concentration of As in plant organs. Different industrial effluents are used for irrigation purposes because these crops are cultivated in suburban regions of the big cities. Contaminated water is used for irrigation of vegetables grown near urban regions and it poses a threat to plants’ health and tends to modulate gene expression as well as metabolite production. So, it is essential to know the varietal response against different heavy metals including As present in industrial wastewater. The var. PS09979325 showed a more effective response against As toxicity, which might be due to its genetic makeup. Based on these observations this var. may be considered as potential candidate for future variety development program. Further molecular studies are required to elaborate on its distinguished behavior of As toxicity tolerance. Additionally, after Se supplementation, balanced organic acid exudation confers typical activities of metabolites in both kinds even under As stress. Thus, to fully comprehend the cellular and molecular mechanisms of As toxicity avoidance/tolerance, long-term field research should be conducted to draw parallels between plant/crop root exudations, metal stress, Se fertigation regimes, nutrient uptake, and plant biomass accumulation.
Data availability
All data generated or analysed during this study are included in this published article.
References
Zemanová V, Pavlíková D, Hnilička F, Pavlík M. Arsenic Toxicity-Induced physiological and metabolic changes in the shoots of Pteris cretica and Spinacia oleracea. 2021.
Chattopadhyay A, Singh AP, Kasote D, Sen I, Regina A. Effect of Phosphorus Application on Arsenic Species Accumulation and Co-Deposition of Polyphenols in Rice Grain: Phyto and Food Safety Evaluation. Plants 2021, Vol 10, Page 281. 2021; 10:281.
Kofroňová M, Hrdinová A, Mašková P, Tremlová J, Soudek P, Petrová Š, et al. Multi-component Antioxidative System and Robust Carbohydrate Status, the Essence of Plant Arsenic Tolerance. Antioxid (Basel). 2020;9:283.
Praveen A, Pandey A, Gupta M. Protective role of nitric oxide on nitrogen-thiol metabolism and amino acids profiling during arsenic exposure in Oryza sativa L. Ecotoxicology. 2020;29:825–36.
Solórzano E, Corpas FJ, González-Gordo S, Palma JM. Reactive oxygen species (ROS) metabolism and nitric oxide (NO) content in roots and shoots of Rice (Oryza sativa L.) plants under Arsenic-Induced stress. Agronomy. 2020;10:1014.
Fayiga A, Geoderma US-. 2016 undefined. Arsenic hyperaccumulating fern: Implications for remediation of arsenic contaminated soils. ElsevierAO Fayiga, UK SahaGeoderma, 2016•Elsevier.
Richards L, Casanueva-Marenco MJ, Magnone D, Sovann P, Dongen B, Polya D. Contrasting Sorption Behaviours Affecting Groundwater Arsenic Concentration in Kandal Province, Cambodia. Geosci Front. 2019;10.
Rabbani U, Mahar G, Siddique A, Fatmi Z. Risk assessment for arsenic-contaminated groundwater along River Indus in Pakistan. Environ Geochem Health. 2017;39:179–90.
Shah Saqib An, Waseem A, Khan Af, Mahmood Q, Khan A, Habib A et al. Arsenic bioremediation by low-cost materials derived from Blue Pine (Pinus wallichiana) and Walnut (Juglans regia). Arsenic bioremediation by low-cost materials derived from Blue Pine (Pinus wallichiana) and Walnut (Juglans regia). 2013; 51:88–94.
Ali H, Khan E, Sajad MA. Phytoremediation of heavy metals—concepts and applications. Chemosphere. 2013;91:869–81.
Nwaobiala CU, Ogbonna MO, editors. Adoption determinants and profitability analysis of okra farming in Aninri Local Government Area (LGA) of Enugu State, Nigeria. DISCOURSE JOURNAL OF AGRICULTURE AND FOOD SCIENCES.
Khan MTI, Qamar Ali QA, Muhammad Ashfaq MA, Muhammad Waseem MW. Economic analysis of open field chilli (Capsicum annuum L.) production in Punjab, Pakistan. 2017.
Rodríguez-Ruiz M, Aparicio-Chacón MV, Palma JM, Corpas FJ. Arsenate disrupts ion balance, sulfur and nitric oxide metabolisms in roots and leaves of pea (Pisum sativum L.) plants. Environ Exp Bot. 2019;161:143–56.
Thakur S, Choudhary S, Majeed A, Singh A, Bhardwaj P. Insights into the molecular mechanism of Arsenic Phytoremediation. J Plant Growth Regul. 2020;39:532–43.
Pezzarossa B, Remorini D, Gentile ML, Massai R. Effects of foliar and fruit addition of sodium selenate on selenium accumulation and fruit quality. J Sci Food Agric. 2012;92:781–6.
Feng R, Wei C, Tu S. The role of selenium in protecting plants against abiotic stresses. Environ Exp Bot. 2013;87:58–68.
Tran TAT, Zhou F, Yang W, Wang M, Dinh QT, Wang D, et al. Detoxification of mercury in soil by selenite and related mechanisms. Ecotoxicol Environ Saf. 2018;159:77–84.
Wang C, Rong H, Zhang X, Shi W, Hong X, Liu W, et al. Effects and mechanisms of foliar application of silicon and selenium composite sols on diminishing cadmium and lead translocation and affiliated physiological and biochemical responses in hybrid rice (Oryza sativa L.) exposed to cadmium and lead. Chemosphere. 2020;251:126347.
Ali S, Chaudhary A, Rizwan M, Anwar HT, Adrees M, Farid M, et al. Alleviation of chromium toxicity by glycinebetaine is related to elevated antioxidant enzymes and suppressed chromium uptake and oxidative stress in wheat (Triticum aestivum L). Environ Sci Pollut Res Int. 2015;22:10669–78.
Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949;24:1–15.
Sakharov IY, Ardila GB. Variations of peroxidase activity in cocoa (Theobroma cacao L.) beans during their ripening, fermentation and drying. Food Chem. 1999;65:51–4.
Hatch MD, Glasziou KT. Sugar Accumulation Cycle in Sugar Cane. II. Relationship of invertase activity to Sugar Content & Growth Rate in Storage tissue of plants grown in controlled environments. Plant Physiol. 1963;38:344–8.
Asada K. Production and scavenging of active oxygen in chloroplasts. Mol Biology free Radical Scavenging Syst. 1992;173–92.
Julkunen-Tiitto R. Phenolic constituents in the leaves of Northern willows: methods for the analysis of certain phenolics. J Agric Food Chem. 1985;33:213–7.
Hodges DM, Nozzolillo C. Anthocyanin and anthocyanoplast content of cruciferous seedlings subjected to mineral nutrient deficiencies. J Plant Physiol. 1996;147:749–54.
Karadeniz F, Burdurlu HS, Koca N, Soyer Y. Antioxidant activity of selected fruits and vegetables grown in Turkey. Turkish J Agric Forestry. 2005;29:297–303.
DuBois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric Method for Determination of Sugars and related substances. Anal Chem. 1956;28:350–6.
Bates LS, Waldren RP, Teare ID. Rapid determination of free proline for water-stress studies. Plant Soil. 1973;39:205–7.
Heath RL, Packer L. Photoperoxidation in isolated chloroplasts: I. kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys. 1968;125:189–98.
Jana S, Choudhuri MA. Glycolate metabolism of three submersed aquatic angiosperms: effect of heavy metals. Aquat Bot. 1981;11:67–77.
Dionisio-Sese ML, Tobita S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998;135:1–9.
Jackson ML. Soil chemical analysis constable. London: Ltd Co; 1958. p. 498.
UdDin I, Bano A, Masood S. Chromium toxicity tolerance of Solanum nigrum L. and Parthenium hysterophorus L. plants with reference to ion pattern, antioxidation activity and root exudation. Ecotoxicol Environ Saf. 2015;113:271–8.
Zvobgo G, Lwalaba JLW, Sagonda T, Mapodzeke JM, Muhammad N, Shamsi IH, et al. Alleviation of arsenic toxicity by phosphate is associated with its regulation of detoxification, defense, and transport gene expression in barley. J Integr Agric. 2019;18:381–94.
Karakaya O, Uygulamalı S, Üniversitesi B, Ateş U, Çelik SM, Faizy AH. Biochemical properties and antimicrobial and antioxidant activity of blackberry growing naturally in Kelkit valley. researchgate.net. 2022; 2022:2687–3818.
Kumar V, Vogelsang L, Schmidt RR, Sharma SS, Seidel T, Dietz K-J. Remodeling of root growth under combined arsenic and hypoxia stress is linked to nutrient deprivation. Front Plant Sci. 2020;11:569687.
Alam MZ, Hoque MA, Ahammed GJ, McGee R, Carpenter-Boggs L. Arsenic accumulation in lentil (Lens culinaris) genotypes and risk associated with the consumption of grains. Sci Rep. 2019;9:9431.
Wang C, Rong H, Zhang X, Shi W, Hong X, Liu W, et al. Effects and mechanisms of foliar application of silicon and selenium composite sols on diminishing cadmium and lead translocation and affiliated physiological and biochemical responses in hybrid rice (Oryza sativa L.) exposed to cadmium and lead. Chemosphere. 2020;251:126347.
Zhao Y, Hu C, Wang X, Qing X, Wang P, Zhang Y, et al. Selenium alleviated chromium stress in Chinese cabbage (Brassica campestris L. Ssp. Pekinensis) by regulating root morphology and metal element uptake. Ecotoxicol Environ Saf. 2019;173:314–21.
Shah Fahad Sönmez, Osman S, Shah W, Depeng W, Chao. Kātibī M ʻAdnān. et al Heavy Met Stress Plants De?F Responses. 2021;October:57–82.
Handa N, Kohli SK, Sharma A, Thukral AK, Bhardwaj R, Alyemeni MN, et al. Selenium ameliorates chromium toxicity through modifications in pigment system, antioxidative capacity, osmotic system, and metal chelators in Brassica juncea seedlings. South Afr J Bot. 2018;119:1–10.
Ahmed B, Dwivedi S, Abdin MZ, Azam A, Al-Shaeri M, Khan MS, et al. Mitochondrial and chromosomal damage induced by oxidative stress in Zn2 + ions, ZnO-bulk and ZnO-NPs treated Allium cepa roots. Sci Rep. 2017;7:40685.
Ahmad P, Alyemeni MN, Al-Huqail AA, Alqahtani MA, Wijaya L, Ashraf M, et al. Zinc oxide nanoparticles application alleviates arsenic (as) toxicity in soybean plants by restricting the uptake of as and modulating key biochemical attributes, antioxidant enzymes, ascorbate-glutathione cycle and glyoxalase system. Plants. 2020;9:825.
Skalickova S, Milosavljevic V, Cihalova K, Horky P, Richtera L, Adam V. Selenium nanoparticles as a nutritional supplement. Nutrition. 2017;33:83–90.
Das K, Roychoudhury A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front Environ Sci. 2014;2:53.
Choudhury FK, Rivero RM, Blumwald E, Mittler R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017;90:856–67.
Liu D, Li H, Wang Y, Ying Z, Bian Z, Zhu W et al. How exogenous selenium affects anthocyanin Accumulation and Biosynthesis-related gene expression in Purple Lettuce. Pol J Environ Stud. 2017;26.
Sachdev S, Ansari SA, Ansari MI, Fujita M, Hasanuzzaman M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, Vol 10, Page 277. 2021; 10:277.
Padhi EMT, Liu R, Hernandez M, Tsao R, Ramdath DD. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J Funct Foods. 2017;38:602–11.
Skrypnik L, Styran T, Savina T, Golubkina N. Effect of selenium application and growth stage at harvest on hydrophilic and lipophilic antioxidants in lamb’s lettuce (Valerianella locusta L. Laterr). Plants. 2021;10:2733.
Wu Z, Yin X, Bañuelos GS, Lin Z-Q, Liu Y, Li M, et al. Indications of selenium protection against cadmium and lead toxicity in oilseed rape (Brassica napus L). Front Plant Sci. 2016;7:1875.
Albornoz F, Lieth JH, González-Fuentes JA. Effect of different day and night nutrient solution concentrations on growth, photosynthesis, and leaf NO3-content of aeroponically grown lettuce. Chil J Agric Res. 2014;74:240–5.
Singh R, Singh S, Parihar P, Singh VP, Prasad SM. Arsenic contamination, consequences and remediation techniques: a review. Ecotoxicol Environ Saf. 2015;112:247–70.
Jiang Y, Ding X, Zhang D, Deng Q, Yu C-L, Zhou S, et al. Soil salinity increases the tolerance of excessive sulfur fumigation stress in tomato plants. Environ Exp Bot. 2017;133:70–7.
Liu H, Xiao C, Qiu T, Deng J, Cheng H, Cong X et al. Selenium Regulates Antioxidant, Photosynthesis, and Cell Permeability in Plants under Various Abiotic Stresses: A Review. Plants 2023, Vol 12, Page 44. 2022; 12:44.
Liu X, Feng H, Fu J, Chen Y, Liu Y, Ma LQ. Arsenic-induced nutrient uptake in As-hyperaccumulator Pteris vittata and their potential role to enhance plant growth. Chemosphere. 2018;198:425–31.
Gupta P, Kumar V, Usmani Z, Rani R, Chandra A, Gupta VK. A comparative evaluation towards the potential of Klebsiella sp. and Enterobacter sp. in plant growth promotion, oxidative stress tolerance and chromium uptake in Helianthus annuus (L). J Hazard Mater. 2019;377:391–8.
Srivastava D, Tiwari M, Dutta P, Singh P, Chawda K, Kumari M et al. Chromium Stress in Plants: Toxicity, Tolerance and Phytoremediation. Sustainability 2021, Vol 13, Page 4629. 2021; 13:4629.
Hayat S, Khalique G, Irfan M, Wani AS, Tripathi BN, Ahmad A. Physiological changes induced by chromium stress in plants: an overview. Protoplasma. 2012;249:599–611.
Zhu XF, Zheng C, Hu YT, Jiang TAO, Liu YU, Dong NY, et al. Cadmium-induced oxalate secretion from root apex is associated with cadmium exclusion and resistance in Lycopersicon Esulentum. Plant Cell Environ. 2011;34:1055–64.
Magdziak Z, Gąsecka M, Waliszewska B, Zborowska M, Mocek A, Cichy WJ, et al. The influence of environmental condition on the creation of organic compounds in Pinus sylvestris L. Rhizosphere, roots and needles. Trees - Struct Function. 2021;35:441–57.
Guo X, Ji Q, Rizwan M, Li H, Li D, Chen G. Effects of biochar and foliar application of selenium on the uptake and subcellular distribution of chromium in Ipomoea aquatica in chromium-polluted soils. Ecotoxicol Environ Saf. 2020;206:111184.
Azizi I, Esmaielpour B, Fatemi H. Effect of foliar application of selenium on morphological and physiological indices of savory (Satureja hortensis) under cadmium stress. Food Sci Nutr. 2020;8:6539–49.
Gulz PA, Gupta SK, Schulin R. Arsenic accumulation of common plants from contaminated soils. Plant Soil. 2005;272:337–47.
Jayakumar M, Surendran U, Raja P, Kumar A, Senapathi V. A review of heavy metals accumulation pathways, sources and management in soils. Arab J Geosci 2021. 2021;14:20.
Ramos I, Esteban E, Lucena JJ, Gárate A. Cadmium uptake and subcellular distribution in plants of Lactuca sp. Cd–Mn interaction. Plant Sci. 2002;162:761–7.
Acknowledgements
The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University for funding this work through large group Research Project under grant number RGP2/486/45.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
MN, ES, AY: Conceptualization, Data curation, Writing – original draft, Formal analysis, Investigation, Methodology; MRK, MHS, AIA, AMEO, FMAK, MAA, SE,, TM: Formal analysis, Validation, Visualization Resources, Writing – review & editing; BA: Data Curation, Formal analysis, Software, Visualization, Writing – original draft, Writing – review & editing. All authors contributed significantly, have read and agreed to the published version of the manuscript.”
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Capsicum annuum L. (HSP-181A and PS09979325) seeds taken from the Ayub Agricultural Research Institute, Faisalabad, Pakistan. All the experiments were performed in accordance with relevant guidelines and regulations”.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Nawaz, M., Shahzadi, E., Yaseen, A. et al. Selenium improved arsenic toxicity tolerance in two bell pepper (Capsicum annuum L.) varieties by modulating growth, ion uptake, photosynthesis, and antioxidant profile. BMC Plant Biol 24, 799 (2024). https://doi.org/10.1186/s12870-024-05509-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-05509-3