
Abstract
Background
Peanut (Arachis hypogaea), a vital oil and food crop globally, is susceptible to web blotch which is a significant foliar disease caused by Phoma arachidicola Marasas Pauer&Boerema leading to substantial yield losses in peanut production. Calcium treatment has been found to enhance plant resistance against pathogens.
Results
This study investigates the impact of exogenous calcium on peanut resistance to web blotch and explores its mechanisms. Greenhouse experiments revealed that exogenous calcium treatment effectively enhanced resistance to peanut web blotch. Specifically, amino acid calcium and sugar alcohol calcium solutions demonstrated the best induced resistance effects, achieving reduction rates of 61.54% and 60% in Baisha1016, and 53.94% and 50% in Luhua11, respectively. All exogenous calcium treatments reduced malondialdehyde (MDA) and relative electrical conductivity (REC) levels in peanut leaves, mitigating pathogen-induced cell membrane damage. Exogenous calcium supplementation led to elevated hydrogen peroxide (H2O2) content and superoxide anion (O2∙-) production in peanut leaves, facilitating the accumulation of reactive oxygen species (ROS) crucial for plant defense responses. Amino acid calcium and sugar alcohol calcium treatments significantly boosted activities of peroxidase (POD), superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) in peanut leaves. Activation of these antioxidant enzymes effectively scavenged excess ROS, maintaining ROS balance and mitigating cellular damage.
Conclusions
In summary, exogenous calcium treatment triggered ROS production, which was subsequently eliminated by the activation of antioxidant enzymes, thereby reducing cell membrane damage and inducing defense responses against peanut web blotch.
Similar content being viewed by others


Background
Peanut (Arachis hypogaea), rich in nutrients such as proteins, vitamins, minerals, polyphenols, and flavonoids, is a crucial source of oil originating from the tropical regions of South America [1, 2]. As the world’s leading producer, China has dedicated approximately 4684 thousand hectares to peanut cultivation, yielding around 18.329 million tons in 2022 (National Bureau of Statistics data, http://www.stats.gov.cn/). However, various biotic and abiotic stresses affect peanut yield, with pests and diseases being among the major factors causing yield reduction [3]. Peanut is susceptible to a multitude of pathogens, with common diseases including peanut black spot, peanut early leaf spot, and peanut web blotch, the latter being the most severe and caused by Phoma arachidicola Marasas Pauer&Boerema [4]. Peanut web blotch primarily occurs during the mid to late stages of peanut growth, especially during the pod-setting and maturing phases, leading to significant defoliation and yield losses. It can generally result in a 10–20% reduction in peanut yield, with losses exceeding 30% in severe cases, thereby posing a serious threat to global peanut production [5]. At present, the predominant strategy for managing peanut web blotch in agricultural production involves the application of chemical fungicides. However, the long-term and frequent application of fungicides can lead to a series of severe problems, including pathogen resistance, excessive pesticide residues in agricultural products, and environmental pollution, thereby degrading peanut quality, constraining sustainable agricultural development, and affecting human health [6,7,8]. Therefore, finding practical and effective control methods that are harmless to the environment and human health has become a hot topic in peanut disease management.
Compared with traditional chemical fungicides, the use of plant immune elicitors is a more environmentally friendly and sustainable plant protection strategy. It enhances the plant’s resistance to pathogens and other stress factors by stimulating the plant’s own defense system, thereby increasing crop yield and economic benefits [9, 10]. Several studies have confirmed that many synthetic and natural chemicals can act as elicitors, such as calcium chloride (CaCl2) [11, 12], benzothiadiazole (BTH) [13, 14], salicylic acid (SA) [15, 16], methyl jasmonate (MEJA) [17, 18], β-aminobutyric acid (BABA) [19, 20], chitosan [21, 22], and silicates [23, 24]. Within this context, calcium (Ca) is recognized as a pivotal element essential for plant growth, serving as a secondary messenger in plant signaling pathways. The application of various calcium compounds is acknowledged for its efficacy in augmenting plant resistance to both biotic and abiotic stresses [25, 26].
The role of calcium in inducing disease resistance operates through enhancing plant cell membrane stability, regulating reactive oxygen species metabolism, and activating antioxidant enzyme systems. Numerous studies have demonstrated that the application of exogenous calcium significantly enhances the levels of both free and bound calcium within plants [27]. This process not only preserves the integrity of cell walls but also maintains the stability of biological membranes. For instance, immersing post-harvest pears in calcium chloride has been shown to increase the formation of calcium phosphate and calcium oxalate on the fruit peel. Such treatments help to maintain the normal structure of cell wall pectin, consequently diminishing the ability of pathogens to invade and reproduce [28,29,30]. Research by Jiang et al. indicates that exogenous calcium application increases the activity of peroxidase enzymes and ROS levels in tomato plants, reducing the incidence of tomato bacterial wilt, suggesting that calcium can regulate the balance of the reactive oxygen species metabolism system, affecting the generation and scavenging of free radicals within cells, thus regulating the biomembrane system and metabolic processes to enhance plant disease resistance [31, 32]. Studies on soybeans by Sugimoto et al. have revealed that spraying calcium formate and calcium nitrate boosts defensive enzyme activities, significantly inhibiting the occurrence of soybean stem rot, indicating that calcium, as an important signaling molecule, can improve plant disease resistance by regulating specific antioxidant enzymes, such as superoxide dismutase, catalase, peroxidase, and ascorbate peroxidase [33, 34]. Furthermore, exogenous calcium also plays a very important role in enhancing the stress resistance and improving the quality of peanuts [35,36,37]. However, compared to vegetables and crops, the mechanism of calcium-induced resistance in peanuts is still not clear.
Calcium, as an elicitor for plant disease control, has potential applications, but its mechanism of inducing disease resistance is still unclear. To explore the potential application of calcium in resisting peanut web blotch and to further elucidate its role in the mechanisms underlying disease resistance in peanuts, this research was structured around four main objectives: (1) the inductive effect of exogenous calcium treatment on peanut web blotch; (2) the impact of exogenous calcium treatment on the permeability of peanut leaf cell membranes; (3) the effect of exogenous calcium treatment on the hydrogen peroxide content and superoxide anion production rate in peanut leaves; (4) the impact of exogenous calcium treatment on the activity of antioxidant enzymes in peanut leaves. The research findings will provide a theoretical basis for further understanding the application of calcium in the mechanism of peanut resistance to web blotch.
Methods
Peanut plants and pathogen preparation
The susceptible peanut variety Baisha1016 and the resistant variety Luhua11 (Dongya Seed) were selected as the study subjects [38]. The peanut web blotch pathogen was provided by the Plant Pathology Laboratory in the Plant Protection College of Shenyang Agricultural University. Spore propagation of the pathogen was conducted before inoculation, by cultivating on oatmeal agar (OA) medium under near-ultraviolet light at 23 ± 1 °C in an incubator for a duration of 14 days. The concentration of spore suspension was measured using a hemocytometer. The spore concentration for inoculation was adjusted to 1.0 × 105 spores/mL using sterile distilled water. The whole peanut plants were inoculated by employing an artificial spray inoculation method.
Exogenous calcium treatment and inoculation
Five calcium compounds (2000 mg/L calcium chloride, 2000 mg/L calcium nitrate, 1500-fold dilution of sorbitol calcium, 1000-fold dilution of amino acid calcium, and 1000-fold dilution of EDTA-Ca solution) were used for treatment [39]. 28 days after sowing, the peanuts were divided into 7 groups. Groups 1 and 2 were sprayed with clean water on the leaves, while groups 3, 4, 5, 6, and 7 were sprayed respectively with calcium chloride, calcium nitrate, sorbitol calcium, amino acid calcium, and EDTA-Ca solution. 48 h after calcium treatment, the prepared spore suspension was sprayed on the leaves of groups 2, 3, 4, 5, 6, and 7 for inoculation. Post-inoculation, the peanut plants within all groups were enclosed in humidity bags for 48 h to ensure consistent environmental conditions, culminating in a total of 7 distinct treatment groups. Each treatment was replicated three times, and samples were taken at 0, 12, 24, 48, 72, and 96 h post-inoculation. Peanut leaves from the same position were cut and immediately frozen in liquid nitrogen and stored at -80 °C for further analysis.
Disease index and induced resistance effect
The development of the disease condition was monitored on the 7th, 14th, and 21st days after inoculation with the peanut web blotch pathogen. The severity grading standard for peanut web blotch disease was used for scoring: 0 level: no disease spots; 1 level: disease spot area covering less than 25% of the leaf area; 2 level: disease spot area covering 26%~50% of the leaf area; 3 level: disease spot area covering more than 50% of the leaf area; 4 level: severe leaf drop or plant death [40]. The disease index and induced resistance effect were calculated using Formula 1 and Formula 2, respectively.
Malondialdehyde (MDA) content and relative electrical conductivity (REC) measurement
The Malondialdehyde (MDA) content was determined using a commercial MDA content detection kit (Solarbio). To evaluate membrane leakage, the relative electrical conductivity (REC) was measured according to Dionisio-Sese and Tobita [41]. 0.1 g of peanut leaves from different treatments were rinsed with deionized water and dried with clean filter paper, then soaked in 10 mL room temperature deionized water for 12 h to measure their conductivity (R1). The test tubes were then heated in a boiling water bath for 30 min, cooled to room temperature, and shaken well before measuring the conductivity post-boiling (R2). The relative electrical conductivity was calculated using Formula 3.
Hydrogen peroxide (H2O2) content determination
The content of Hydrogen Peroxide (H2O2) was determined according to the method of Rao MV et al. [42]. 1 g of peanut leaves from different treatments were ground into powder in liquid nitrogen, placed in a centrifuge tube with 2 mL of precooled acetone, and mixed well into a slurry. The tubes were then centrifuged at 10,000 g at 4 °C for 10 min. 1 mL of supernatant was taken, mixed with 3 mL of extracting agent (CCl4-CHCl3, 3:1 by volume) and 5 mL of distilled water, and mixed well. After centrifugation at 5000 g for 1 min, the upper aqueous phase was used for H2O2 measurement. 1 mL of the test liquid was added to 2 mL of working solution (Reagent A: 3.3 mmolL-1 ferrous sulfate, 3.3 mmolL-1 ammonium sulfate, 412.5 mmolL-1 sulfuric acid; Reagent B: 165 mmolL-1 xylenol orange, 165 mmolL-1 sorbitol; A:B volume ratio 1:10), incubated in a 30 °C water bath for color development for 30 min, and the absorbance at 560 nm was measured. A standard curve for H2O2 was prepared using the same method, and the H2O2 content in peanut leaves was calculated.
Superoxide anion generation rate (O2 ∙-) measurement
The generation rate of Superoxide Anion (O2∙-) was determined according to the method of Wang Aiguo et al. [43]. 1 g of peanut leaves from different treatments were ground into powder in liquid nitrogen, placed in a centrifuge tube with 4 mL of 50 mmolL-1 phosphate buffer (pH 7.8), and mixed well. The mixture was then centrifuged at 10,000 g at 4 °C for 20 min, and the supernatant was used as the test liquid for O2∙- generation. 0.5 mL of test liquid was added to 0.5 mL of 50 mmolL-1 phosphate buffer (pH 7.8) and 1 mL of 1 mmolL-1 hydroxylamine hydrochloride, mixed well, and incubated at 25 °C for 1 h. Then, 2 mL each of 17 mmolL-1 p-aminobenzenesulfonic acid and 17 mmolL-1 α-naphthylamine were added, mixed well, and incubated at 25 °C for 20 min. The absorbance at 530 nm was measured. A NO2- standard curve was prepared using the same method, and the generation rate of O2∙- in peanut leaves was calculated.
Antioxidant enzyme activity measurement
The activities of Peroxidase (POD), Catalase (CAT), Superoxide Dismutase (SOD), and Ascorbate Peroxidase (APX) were measured using commercial enzyme activity detection kits (Solarbio) according to the manufacturer’s instructions.
Data processing
The data were analyzed statistically using SPSS Statistics 26 software. One-way ANOVA and Duncan’s multiple range test were utilized for significance testing, with a significance level set at P < 0.05.
Results
Induced resistance effects of exogenous calcium treatment against peanut web blotch
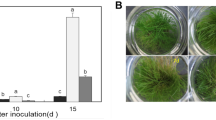
Under greenhouse cultivation conditions, foliar application of five different exogenous calcium treatments showed a certain degree of induced resistance against peanut web blotch disease (Fig. 1). The disease index of peanut web blotch post-exogenous calcium treatment was lower compared to the control, with amino acid calcium and sorbitol calcium treatments resulting in the lowest disease index (Fig. 2). 7 days post-treatment, amino acid calcium showed the best induced resistance effect, reaching 61.54% (Baisha) and 60% (Luhua), followed by sorbitol calcium and calcium nitrate at 53.94% (Baisha) and 50% (Luhua), 42.31% (Baisha) and 39.99% (Luhua) respectively, and lastly calcium chloride and EDTA-Ca at 34.61% (Baisha) and 24.99% (Luhua), 23.07% (Baisha) and 13.32% (Luhua). 14 days post-treatment, the induced resistance effects of calcium chloride, calcium nitrate, sorbitol calcium, amino acid calcium, and EDTA-Ca on peanut web blotch disease were 40.57%, 46.23%, 51.89%, 57.55%, and 32.08% (Baisha), and 38.54%, 48.95%, 52.08%, 56.25%, and 31.25% (Luhua), respectively. 21 days post-treatment, the induced resistance effects of calcium chloride, calcium nitrate, sorbitol calcium, amino acid calcium, and EDTA-Ca on peanut web blotch disease were 37.59%, 46.45%, 53.19%, 58.86%, and 22.34% (Baisha), and 42.63%, 38.94%, 48.36%, 52.05%, and 32.79% (Luhua), respectively. Among these, the overall disease resistance effect of exogenous calcium on Luhua 11 was better than that on Baisha 1016 (Fig. 3).

Exogenous calcium reduces the incidence of peanut leaf disease. a In-Vitro. b In-Vivo

Effect of foliar application of exogenous calcium on inducing resistance in peanuts. a Baisha 1016 inoculated Phoma arachidicola Disease index. b Baisha 1016 inoculated Phoma arachidicola resistance-inducing effect. c Luhua 11 inoculated Phoma arachidicola Disease index. d Luhua 11 inoculated Phoma arachidicola resistance-inducing effect. Note: Error bars represented standard deviation (n = 3); Different letters indicated significant difference at 0.05 level by Duncan; The same as below

Comparison of induced resistance effects among different peanut varieties
Effects of exogenous calcium treatments on the membrane permeability of peanut leaves
MDA content in peanut leaves without inoculation remained unchanged over time, but gradually increased after inoculation with the web blotch pathogen. Compared to the leaves inoculated post-water spray, the MDA content in leaves treated with exogenous calcium and then inoculated was consistently lower. The most significant difference was observed at 72 h post-treatment (Fig. 4ab), where the MDA content in leaves treated with calcium chloride, calcium nitrate, sorbitol calcium, amino acid calcium, and EDTA-Ca decreased by 34.04%, 36.70%, 39.89%, 40.96%, 26.60% (Baisha) and 24.84%, 29.19%, 27.95%, 29.81%, 16.77% (Luhua), respectively.
Following inoculation with the web blotch pathogen, the REC of peanut leaves gradually increased over time. At 72 h post-treatment, compared to water spray followed by inoculation, foliar application of exogenous calcium before inoculation significantly lowered the REC, showcasing decreases of 35.72%, 38.36%, 37.34%, 40.49%, 30.52% (Baisha) and 39.50%, 37.89%, 35.50%, 37.20%, 27.52% (Luhua), respectively (Fig. 4cd).

Effect of exogenous calcium treatments on the MDA content of peanut leaves. a Baisha 1016 inoculated Phoma arachidicola.b Luhua 11 inoculated Phoma arachidicol. Effect of exogenous calcium treatments on the REC content of peanut leaves. c Baisha 1016 inoculated Phoma arachidicola.d Luhua 11 inoculated Phoma arachidicol
Impact of exogenous calcium treatment on hydrogen peroxide content and superoxide anion production rate in peanut leaves
Under calcium chloride and calcium nitrate treatments, the hydrogen peroxide (H2O2) content in peanut leaves rapidly increased after inoculation with the web blotch pathogen, peaking at 12 h, sharply decreasing after 24 h, and returning to baseline levels by 72 h. For sorbitol calcium, amino acid calcium, and EDTA-Ca treatments, the H2O2 content gradually increased over 0 to 48 h, peaked at 48 h, and then slowly decreased. This indicates that exogenous calcium treatments enhance the accumulation of H2O2 in peanut leaves, thus inducing an efficient and rapid response to infection by the web blotch pathogen (Fig. 5ab).
Following inoculation with the web blotch pathogen, all treatments influenced the rate of superoxide anion (O2∙-) production in peanut leaves to some extent. As shown in Fig. 4, foliar application of calcium chloride and calcium nitrate resulted in a rapid increase in the production rate of O2∙- within 0 to 12 h, peaking at 12 h before quickly declining. For leaves treated with amino acid calcium and sorbitol calcium, the production rate of O2∙- gradually rose over 0 to 24 h, peaking at 24 h, with amino acid calcium treatment leading to the fastest production rate. This demonstrates that exogenous calcium treatments can induce bursts of O2∙- in peanut leaves, enhancing resistance against the web blotch pathogen (Fig. 5cd).

Effect of exogenous calcium treatments on the H2O2 content of peanut leaves. a Baisha 1016 inoculated Phoma arachidicola.b Luhua 11 inoculated Phoma arachidicol. Effect of exogenous calcium treatments on the O2∙- content of peanut leaves. c Baisha 1016 inoculated Phoma arachidicola.d Luhua 11 inoculated Phoma arachidicol
Effects of exogenous calcium treatment on antioxidative enzyme activities in peanut leaves
Following inoculation with the web blotch pathogen and exogenous calcium treatment, peroxidase (POD) activity in leaves of different peanut varieties was significantly enhanced (Fig. 6ab). In leaves inoculated post-water spray, POD activity rapidly rose within 0 to 48 h, peaking at 48 h before immediately dropping back to baseline; for leaves treated with calcium chloride and calcium nitrate and then inoculated, POD activity peaked at 24 h; leaves treated with sorbitol calcium and amino acid calcium and then inoculated saw a peak in POD activity at 48 h; with EDTA-Ca treatment, no distinct peak period was observed, with the highest POD activity recorded between 24 and 72 h.
Under exogenous calcium treatment, catalase (CAT) activity in peanut leaves was affected to varying degrees (Fig. 6cd). 24 h post-treatment, leaves treated with calcium chloride and calcium nitrate and then inoculated with the web blotch pathogen reached peak CAT activity levels, which then gradually declined; forty-eight hours post-treatment, leaves treated with sorbitol calcium and amino acid calcium and then inoculated reached peak CAT activity levels, which then gradually declined. Under EDTA-Ca treatment, CAT activity in peanut leaves continuously increased between 24 and 72 h without a clear peak.
Similarly, superoxide dismutase (SOD) activity was significantly enhanced in leaves of different peanut varieties following inoculation with the web blotch pathogen and exogenous calcium treatment (Fig. 6ef). In leaves inoculated post-water spray, SOD activity rapidly rose within 0 to 48 h, peaking at 48 h before quickly declining to baseline levels; leaves treated with calcium chloride and calcium nitrate and then inoculated reached peak SOD activity at 24 h; leaves treated with sorbitol calcium and amino acid calcium and then inoculated peaked in SOD activity at 48 h; with EDTA-Ca treatment, no distinct peak period was observed, with the highest SOD activity recorded between 24 and 72 h.
Following inoculation with the web blotch pathogen and exogenous calcium treatment, ascorbate peroxidase (APX) activity was significantly enhanced in leaves of different peanut varieties (Fig. 6gh). Leaves treated with calcium chloride and calcium nitrate and then inoculated experienced a sharp rise in APX activity between 12 and 24 h, peaking at 24 h before quickly declining, but remained consistently higher than APX activity in leaves inoculated post-water spray and untreated; leaves treated with sorbitol calcium and amino acid calcium and then inoculated reached peak APX activity at 48 h, which then slowly declined; under EDTA-Ca treatment, no distinct peak period was observed, with the highest APX activity recorded between 48 and 96 h.

Effect of exogenous calcium treatments on the peroxidase system activities of peanut leaves. a POD Baisha 1016 inoculated Phoma arachidicola.b POD Luhua 11 inoculated Phoma arachidicol.c CAT Baisha 1016 inoculated Phoma arachidicola.d CAT Luhua 11 inoculated Phoma arachidicol.e SOD Baisha 1016 inoculated Phoma arachidicola.f SOD Luhua 11 inoculated Phoma arachidicol. g APX Baisha 1016 inoculated Phoma arachidicola.h APX Luhua 11 inoculated Phoma arachidicol
Discussion
The results of this experiment indicate that the application of five exogenous calcium treatments exhibited varying levels of induced resistance against peanut web blotch. Among these treatments, amino acid calcium was the most effective, inducing resistance in Baisha1016 and Luhua11 varieties by 61.54% and 60%, respectively. This was followed by sorbitol calcium and nitrate calcium, with induction effects of 53.94% and 50%, 42.31% and 39.99%, respectively, while calcium chloride and EDTA-Ca had the least effects, at 34.61% and 24.99%, 23.07% and 13.32%. Additionally, exogenous calcium treatments reduced the MDA content and REC in peanut leaves, enhanced the accumulation of H2O2 and O2∙-, and increased the activities of POD, CAT, SOD, and APX. This suggests that exogenous calcium treatments can induce peanut’s defense against web blotch through various pathways, including reducing cell membrane permeability, triggering ROS bursts, and activating antioxidant enzymes, thereby enhancing resistance to the disease.
Previous research results have also confirmed the role of calcium in inducing disease resistance. For instance, studies by Paula Júnior et al. have shown that spraying CaCl2 and CaSiO3 on bean leaves significantly reduces the disease index of bean plants infected with Sclerotinia sclerotiorum [44]; Yoon et al. found that treatments with different calcium compounds significantly reduce the susceptibility of peppers to Botrytis cinerea [45]. Thus, the use of calcium as an elicitor for controlling peanut diseases holds broad prospects.
The cell membrane is considered the first line of defense for plants against environmental stresses. When plants are subjected to various harmful stresses, the cell membrane gets damaged, significantly increasing its permeability. MDA content and REC are two main indicators of cell membrane permeability [46, 47]. Calcium treatments can stabilize plant epidermal cell membranes by reducing the leaf MDA content and REC, thereby mitigating pathogen invasion and proliferation. In contrast, a deficiency in calcium increases membrane permeability, allowing low molecular compounds (e.g., sugars, amino acids) to seep into the extraplastidial matrix of leaves and stem tissues, facilitating pathogen invasion and reproduction [48]. Exogenous calcium treatment enhances the stability of leaf epidermal cell membranes, activates the plant’s defense response to the web blotch pathogen, and effectively reduces the severity of peanut web blotch.
ROS are key defenders against pathogen invasion, either by directly killing pathogens or by slowing their entry into plant tissues. Under stress conditions, ROS such as H2O2 and O2∙- are produced. These strong oxidizing agents can rapidly attack and cause damage to various biomolecules. Both H2O2 and O2∙- can interact with polyunsaturated lipids in cell membranes, generating lipid peroxides that can lead to the destruction of biological membranes [49]. The application of exogenous calcium can induce an increase in H2O2 content and enhance the rate of O2∙- production in peanut leaves. This indicates that under pathogen inoculation conditions, exogenous calcium-treated peanut leaves can rapidly accumulate more ROS, a process considered a key step in inducing further defense responses in plants. However, excessive ROS accumulation can lead to oxidative stress and ultimately cell death [49]. Plants have evolved many mechanisms to eliminate excess ROS throughout their evolutionary history, among which antioxidant enzymes are primary ROS scavengers, crucial for plant stress resistance [50, 51]. Exogenous calcium treatment significantly increased the activity of antioxidant enzymes, including POD, CAT, SOD, and APX, in peanut leaves, effectively eliminating excess ROS and maintaining ROS balance [50]. Overall, exogenous calcium treatment can induce peanut plant resistance to web blotch pathogen infection by regulating ROS metabolism to accumulate ROS and by activating antioxidant enzymes to eliminate excess ROS.
Conclusions
In the context of sustainable agriculture, disease control strategies are increasingly considering the impact on the ecological environment and the protection of biodiversity among other issues. The use of exogenous inducers as an alternative to chemical fungicides for plant disease control presents a better option. Exogenous calcium as an inducer in the control of peanut diseases can reduce the impact of chemical fungicides on the environment, agricultural products, and human health. This experiment show that exogenous calcium has a certain inducing resistance effect on peanut web blotch. Exogenous calcium treatment can induce peanut to produce a defensive response through various pathways such as reducing the permeability of the cell membrane, triggering ROS bursts, and activating the activity of the antioxidant enzyme system, thereby improving the disease resistance of peanuts. This experiment provides a theoretical basis for controlling peanut diseases with exogenous calcium and offers reference significance for the prevention and control of diseases in other crops. However, the specific mechanisms of exogenous calcium treatment still require further research, including the analysis of regulatory networks of related genes and calcium signaling pathways. Moreover, in practical applications, it is necessary to further consider the optimal conditions and application methods for exogenous calcium treatments to maximize plant disease resistance.
Data availability
The author confirms that all data generated or analyzed during this study are included in this published article.
References
Bertioli DJ, Cannon SB, Froenicke L, Huang G, Farmer AD, Cannon EK, Liu X, Gao D, Clevenger J, Dash S, Ren L, Moretzsohn MC, Shirasawa K, Huang W, Vidigal B, Abernathy B, Chu Y, Niederhuth CE, Umale P, Araújo AC, Kozik A, Kim KD, Burow MD, Varshney RK, Wang X, Zhang X, Barkley N, Guimarães PM, Isobe S, Guo B, Liao B, Stalker HT, Schmitz RJ, Scheffler BE, Leal-Bertioli SC, Xun X, Jackson SA, Michelmore R, Ozias-Akins P. The genome sequences of Arachis duranensis and Arachis Ipaensis, the diploid ancestors of cultivated peanut. Nat Genet. 2016;48(4):438–46.
Song Q, Liu Y, Pang J, Yong JWH, Chen Y, Bai C, Gille C, Shi Q, Wu D, Han X, Li T, Siddique KHM, Lambers H. Supplementary calcium restores peanut (Arachis hypogaea) growth and photosynthetic capacity under low nocturnal temperature. Front Plant Sci. 2020;10:1637.
Moretzsohn MC, Dos Santos JF, Moraes ARA, Custódio AR, Michelotto MD, Mahrajan N, Leal-Bertioli SCM, Godoy IJ, Bertioli DJ. Marker-assisted introgression of wild chromosome segments conferring resistance to fungal foliar diseases into peanut (Arachis hypogaea L). Front Plant Sci. 2023;14:1139361.
Sun Z, Cheng Y, Qi F, Zhang M, Tian M, Wang J, Wu X, Huang B, Dong W, Zhang X. Zhang Z Resistance of peanut to web blotch caused by Phoma arachidicola is related to papillae formation and the hypersensitive response.Plant Pathology. 2022;71:1921–31.
ZHOU R, CUI J, FU J, XU Z. Effect of mixed infection of Cercospora arachidicola and Phoma arachidicola on their infection probability and latent periods. Scientia Agricultura Sinica. 2015;48(21):4264–71.
Luo M, Dang P, Bausher MG, Holbrook CC, Lee RD, Lynch RE, Guo BZ. Identification of transcripts involved in resistance responses to Leaf Spot Disease caused by Cercosporidium Personatum in Peanut (Arachis hypogaea). Phytopathology. 2005;95(4):381–7.
Woodward JE, Brenneman TB, Kemerait RC, Culbreath AK, Smith NB. Management of peanut diseases with reduced input fungicide programs in fields with varying levels of disease risk. Crop Prot. 2010;29(3):222–29.
Culbreath AK, Kemerait RC Jr, Brenneman TB, Cantonwine EG, Rucker KS. Effect of In-Furrow application of Fluopyram on Leaf Spot diseases of Peanut. Plant Dis. 2021;105(9):2374–9.
EL-Hendawy S, Shaban W, Sakagami JI. Does treating faba bean seeds with chemical inducers simultaneously increase chocolate spot disease resistance and yield under field conditions? Turk J Agric for. 2014;34(6):475–85.
QIU D. Progress and Prospect of Plant Immunity Inducer. J Agricultural Sci Technol. 2014, 16(1): 39–45.
Chandra S, Chakraborty N, Chakraborty A, Rai R, Bera B. Abiotic elicitor-mediated improvement of innate immunity in Camellia sinensis. J Plant Growth Regul. 2014;33(4):849–59.
Ayón-Reyna LE, González-Robles A, Rendón-Maldonado JG, Báez-Flores ME, López-López ME, Vega-Garcia MO. Application of a hydrothermal-calcium chloride treatment to inhibit postharvest anthracnose development in papaya. Postharvest Biol Technol. 2017;124:85–90.
Suo Y, Leung DWM. BTH-induced accumulation of extracellular proteins and blackspot disease in rose. Biol Plant. 2002;45(2):273–9.
Lin J, Gong D, Zhu S, Zhang L, Zhang L. Expression of PPO and POD genes and contents of polyphenolic compounds in harvested mango fruits in relation to Benzothiadiazole-induced defense against anthracnose. Sci Hort. 2011;130(1):85–9.
Kobeasy MI, Elbeltagi HS, Elshazly MA, Khattab EA. Induction of resistance in Arachis hypogaea L. against Peanut Mottle virus by nitric oxide and salicylic acid. Physiol Mol Plant Pathol. 2011;76(2):112–8.
Tang Y, Kuang J, Wang F, Chen L, Hong K, Xiao Y, Xie H, Lu W, Chen J. Molecular characterization of PR, and WRKY, genes during SA- and MeJA-induced resistance against Colletotrichum musae in banana fruit. Postharvest Biol Technol. 2013;79(1):62–8.
Jiang L, Jin P, Wang L, Yu X, Wang H, Zheng Y. Methyl jasmonate primes defense responses against Botrytis Cinerea and reduces disease development in harvested table grapes. Sci Hort. 2015;192:218–23.
Pan L, Chen X, Xu W, Fan S, Wan T, Zhang J, Cai Y. Methyl jasmonate induces postharvest disease resistance to decay caused by Alternaria alternata in sweet cherry fruit. Sci Hort. 2022;292:110624.
Chavan V, Bhargava S, Kamble A. Temporal modulation of oxidant and antioxidative responses in Brassica carinata during β-aminobutyric acid-induced resistance against Alternaria brassicae. Physiol Mol Plant Pathol. 2013;83(5):35–9.
Siegrist J, Orober M, Buchenauer H. β-Aminobutyric acid-mediated enhancement of resistance in tobacco to tobacco mosaic virus depends on the accumulation of salicylic acid. Physiol Mol Plant Pathol. 2000;56(3):95–106.
Nandeeshkumar P, Sudisha J, Ramachandra KK, Prakash H, Niranjana SR, Shekar S. Chitosan induced resistance to downy mildew in sunflower caused by Plasmopara Halstedii. Physiol Mol Plant Pathol. 2008;72(4):188–94.
Bhaskara Reddy MV, Arul J, Angers P, Couture L. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearum and improves seed quality. J Agric Food Chem. 1999;47(3):1208–16.
Van Bockhaven J, Spíchal L, Novák O, Strnad M, Asano T, Kikuchi S, Höfte M, De Vleesschauwer D. Silicon induces resistance to the brown spot fungus Cochliobolus miyabeanus by preventing the pathogen from hijacking the rice ethylene pathway. New Phytol. 2015;206(2):761–73.
Najihah NI, Hanafi MM, Idris AS, Hakim MA. Silicon treatment in oil palms confers resistance to basal stem rot disease caused by Ganoderma boninense. Crop Prot. 2015;67:151–9.
Sugimoto T, Aino M, Sugimoto M, Watanabe K. Reduction of Phytophthora stem rot disease on soybeans by the application of CaCl2 and ca(NO3)2. J Phytopathol. 2005;153(9):536–43.
Yang S, Wang F, Guo F, Meng JJ, Li XG, Wan SB. Calcium contributes to photoprotection and repair of photosystem II in peanut leaves during heat and high irradiance. J Integr Plant Biol. 2015;57(5):486–95.
Silveira AC, Aguayo E, Chisari M, Artés F. Calcium salts and heat treatment for quality retention of fresh-cut ‘Galia’ melon. Postharvest Biol Technol. 2011;62:77–84.
Fang WANG, Zhen-lun LI, Yan-li CHEN, Shui-ying YANG, Yi XU. Recent advances on inhibition mechanisms of calcium on Plant Diseases[J]. Biotechnol Bull. 2017;33(2):1–7.
Ortiz A, Graell J, Lara I. Preharvest calcium applications inhibit some cell wall-modifying enzyme activities and delay cell wall disassembly at commercial harvest of Fuji Kiku-8 apples[J]. Volume 62. Postharvest Biology & Technology; 2011. pp. 161–7. 2.
Jianfeng Liu P, Tang, Peng L. Effects of Calcium Dipping after Harvest on Content of Calcium in different forms and physio-chemical characteristics of pear [J]. J Huazhong Agricultural Univ, 2004,23 (5).
Jiang JF, Li JG, Dong YH. Effect of calcium nutrition on resistance of tomato against bacterial wilt induced by Ralstonia solanacearum[J]. Eur J Plant Pathol. 2013;136(3):547–55.
Cheour F. Calcium delays the postharvest ripening and related membrane lipid-changes-of-tomato[J]. Nutrition & Food Science; 2015.
Sugimoto T, Watanabe K, Yoshida S, Aino M, Furiki M, Shiono M, Matoh T, Biggs AR. Field Application of Calcium to Reduce Phytophthora Stem rot of soybean, and calcium distribution in plants. Plant Dis. 2010;94(7):812–9.
Li L, Zou Y. Induction of disease resistance by salicylic acid and calcium ion against Botrytis Cinerea in tomato (Lycopersicon esculentum)[J]. Emirates J Food Agric. 2016;29(1):1.
Song Q, Zhang S, Bai C, Shi Q, Wu D, Liu Y, Han X, Li T, Yong JWH. Exogenous Ca2 + priming can improve peanut photosynthetic carbon fixation and pod yield under early sowing scenarios in the field. Front Plant Sci. 2022;13:1004721.
Wu D, Liu Y, Pang J, Yong JWH, Chen Y, Bai C, Han X, Liu X, Sun Z, Zhang S, Sheng J, Li T, Siddique KHM, Lambers H. Exogenous calcium alleviates Nocturnal Chilling-Induced Feedback inhibition of photosynthesis by improving Sink demand in peanut (Arachis hypogaea). Front Plant Sci. 2020;11:607029.
Li T, Wei Q, Sun W, Tan H, Cui Y, Han C, Zhang H, Zeng F, Huang M, Yan D. Spraying sorbitol-chelated calcium affected foliar calcium absorption and promoted the yield of peanut (Arachis hypogaea L). Front Plant Sci. 2022;13:1075488.
Fu JF, Cui HC, Zhou RJ, et al. Identification of resistance of major peanut cultivars (lines) to early leaf spot and web blotch in Liaoning Province. Plant Prot. 2015;41(01):171–3.
Luo KS, Wu XM, Luo Y, et al. The effects of two types of exogenous calcium on the resistance of prickly pear to top rot and fruit quality. South China Fruits. 2023;52(02):191–6.
Yuan HX, Sun BJ, Li HL, et al. Identification of resistance to leaf spot in peanut cultivars (lines) [J]. J Henan Agricultural Sci. 2004;12:35–8.
Dionisio-Sese ML, Tobita S. Antioxidant responses of rice seedlings to salinity stress [J]. Plant Sci. 1998;135(1):1–9.
Rao MV, Paliyath G, Ormrod DP, Murr DP, Watkins CB. Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2-metabolizing enzymes. Salicylic acid-mediated oxidative damage requires H2O2. Plant Physiol. 1997;115(1):137–49.
Wang AG, Luo GH. Quantitative relation between the reaction of hydroxylamine and superoxide anion radicals in plants [J]. Plant Physiol Commun. 1990;84(15):2895–8.
Paula Júnior TJ, Vieira RF, Teixeira H, et al. Foliar application of calcium chloride and calcium silicate decreases white mold intensity on dry beans[J]. Trop Plant Pathol. 2009;34(3):171–4.
Yoon CS, Yeoung YR, Kim BS. The suppressives effects of calcium compounds against Botrytis Cinerea in paprika[J]. Korean J Hortic Sci Technol. 2010;28(6):1072–7.
Dai H, Wei S, Twardowska I, Han R, Xu L. Hyperaccumulating potential of Bidens pilosa L. for cd and elucidation of its translocation behavior based on cell membrane permeability. Environ Sci Pollut Res Int. 2017;24(29):23161–7.
Qi C, Lin X, Li S, Liu L, Wang Z, Li Y, Bai R, Xie Q, Zhang N, Ren S, Zhao B, Li X, Fan S, Guo YD. SoHSC70 positively regulates thermotolerance by alleviating cell membrane damage, reducing ROS accumulation, and improving activities of antioxidant enzymes. Plant Sci. 2019;283:385–95.
Muhamad Tajudin NS, Musa MH, Abu Seman I, et al. Determination and mapping of calcium and magnesium contents using geostatistical techniques in oil palm plantation related to basal stem rot disease[J]. Songklanakarin J Sci Technol. 2016;38(1):23–30.
Mittler R, Zandalinas SI, Fichman Y, Van Breusegem F. Reactive oxygen species signalling in plant stress responses. Nat Rev Mol Cell Biol. 2022;23(10):663–79.
Dvořák P, Krasylenko Y, Zeiner A, Šamaj J, Takáč T. Signaling toward reactive oxygen species-scavenging enzymes in plants. Front Plant Sci. 2021;11:618835.
Pan L, Zhao X, Chen M, Fu Y, Xiang M, Chen J. Effect of exogenous methyl jasmonate treatment on disease resistance of postharvest kiwifruit. Food Chem. 2020;305:125483.
Acknowledgements
The project was supported by the Natural Science Foundation of Liaoning Province (2022-MS-264).
Funding
The research was supported by the Natural Science Foundation of Liaoning Province (2022-MS-264).
Author information
Authors and Affiliations
Contributions
Lanshuo Yan: Experimentation, Data Curation, Statistical analysis, Writing and Drafting. Sichao Liu: Experimentation, Data Curation. Ruoxin Li: Experimentation, Data Curation. Zibo Li: Methodology, Statistical analysis. Jingzi Piao: Research Design, Statistical analysis, Writing and Review. Rujun Zhou: Research Design, Review and Editing.
Corresponding authors
Ethics declarations
Ethics approval, guidelines and consent to participate
The source of the plant material used in the study was purchased. The authors confirmed that all methods were performed in accordance with the relevant regulations/legislation.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yan, L., Liu, S., Li, R. et al. Calcium enhanced the resistance against Phoma arachidicola by improving cell membrane stability and regulating reactive oxygen species metabolism in peanut. BMC Plant Biol 24, 501 (2024). https://doi.org/10.1186/s12870-024-05222-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-05222-1