
Abstract
Background
The climatic changes crossing the world menace the green life through limitation of water availability. The goal of this study was to determine whether Moringa oleifera Lam. trees cultivated under Tunisian arid climate, retain their tolerance ability to tolerate accentuated environmental stress factors such as drought and salinity. For this reason, the seeds of M. oleifera tree planted in Bouhedma Park (Tunisian arid area), were collected, germinated, and grown in the research area at the National Institute of Research in Rural Engineering, Waters and Forests (INRGREF) of Tunis (Tunisia). The three years aged trees were exposed to four water-holding capacities (25, 50, 75, and 100%) for 60 days to realise this work.
Results
Growth change was traduced by the reduction of several biometric parameters and fluorescence (Fv/Fm) under severe water restriction (25 and 50%). Whereas roots presented miraculous development in length face to the decrease of water availability (25 and 50%) in their rhizospheres. The sensitivity to drought-induced membrane damage (Malondialdehyde (MDA) content) and reactive oxygen species (ROS) liberation (hydrogen peroxide (H2O2) content) was highly correlated with ROS antiradical scavenging (ferric reducing antioxidant power (FRAP) and (2, 2’-diphenyl-1-picrylhydrazyle (DPPH)), phenolic components and osmolytes accumulation. The drought stress tolerance of M. oleifera trees was associated with a dramatic stimulation of superoxide dismutase (SOD), catalase (CAT), glutathione reductase (GR), ascorbate peroxidase (APX), and glutathione peroxidase (GPX) activities.
Conclusion
Based on the several strategies adopted, integrated M. oleifera can grow under drought stress as accentuated adverse environmental condition imposed by climate change.
Similar content being viewed by others

Introduction
The climatic changes sweeping the world and menace the green life requires urgent attention and the prevention of many disasters, such as the heightened hazard of limited water availability [1]. This horrifying truth that global droughts are increasing over the different regions of world, is driving researchers to focus on finding practical and effective solutions to overcome the intense problematic. In fact, water unavailability is the main limiting factor of plants growth and a total disruption of the several physiological processes causing early senescence and development reduction [2]. As consequence, the hazard effects on field crops largely affected food security [3].
Moringa, together with its 13 species, is one of the most well-known plants studied in science. However, Moringa oleifera Lam. is the most studied species, owing to its significant benefits. For the first time, it is crucial to emphasise Moringa’s extraordinary ability to endure drought stress [4]. In addition, Ashutosh et al. [5] in their published update study explain several significant features of this species in detail. Thanks to its safety property for human consumption, the Moringa tree could be considered as promising natural ingredient food product aiming to improve the overall nutritional characteristics [6]. Moringa oleifera known as the miracle tree or tree of life, the main representative of the order Moringa, it belongs to family Moringaceae and it is also commonly known as horseradish tree or drum stick. Moringa has both medicinal and nutritional values and uses with essential minerals, amino acids and vitamins [7]. Especially, leaves were largely used because of their high tenor of proteins, essential amino acids, dietary fibres, vitamins, and minerals, while being low in fat, sugars, and total starch protein and total dietary fibre amounts [8].
Moringa is referenced for its diverse therapeutic properties, for example antipyretic, antipathy to diabetes, calming, anticancer and antiulcer [9]. Seeds oils, which are rich in bioactive compounds, were used as anti-inflammatory, nephroprotective, antioxidant, anti-diabetic, anti-cancer, and anti-obesity [10]. In the agronomic sector, Moringa can be used as biofertilizer reducing the costs of chemical fertilizer application and save the environment. Several studies realised by Yap et al. [11], Yaseen and Takacs-Hajos [12], Chuene et al. [13] showed that Moringa used as fertilizer, improve crops productivity of Foeniculum vulgare, Mill, Solanum lycopersicum, Silybum marianum, and Lactuca sativa under normal and stress conditions.
Face to the environmental horrible problem, many plant species tolerate drought stress and survive under limited water availability. Moringa is one of the miraculous plants that incorporate several ecophysiological mechanisms to persist face this abiotic stress factor [14, 15]. Under stress conditions, plants present several strategies to overcome the negative induced effect produced through production of large range of chemicals [16]. Secondary metabolites were generated to support the internal changes caused by stress factors. More precisely, they play an antioxidant role through hydrogen donation from phenolic groups to quenching of free radicals [17]. Drought stress which becoming more erratic, caused significant changes in nutritional quality and biochemical compounds such as phenolic acids [18].
Alternatively, in stressed plants, the build-up of appropriate organic solutes (betaines, soluble sugar, proline, and organic acids) is observed to preserve cellular homeostasis, shield enzymes, and scavenge reactive oxygen species [19]. Reactive oxygen species (ROS) generation is mainly attributed to the negative effect of stress conditions. To endure this oxidative damage and to improve their antioxidant potential, plants cells modulate several enzymatic responses. Antioxidant enzymes (superoxide dismutase (SOD), catalase (CAT), Ascorbate peroxidase (APX) and glutathione reductase (GR) peroxidase (POD) were stimulated under abiotic stress such as, salinity and drought. In fact, numerous studies [15, 20] showed a positive correlation between abiotic stress factors, bioactive molecules accumulation such as polyphenols and the enzymatic antioxidant activities enhancement in Moringa oleifera in order to mitigate the unfavourable conditions of growth.
To address the lack of water availability effects, this study was conducted to: (1) assess the morphophysiological parameters, growth responses, secondary metabolites, osmoregulators, and enzymatic and non-enzymatic antioxidant changes in Moringa oleifera trees. (2) Use various multivariate analysis approaches to verify and validate the relationships between morphophysiological parameters and biochemical responses.
Materials and methods
Plant material and growth conditions
Moringa oleifera Lam. seeds collected from Bouhedma National Park (Tunisian center, east) in 2018, were cultivated in the research area at the National Institute of Research in Rural Engineering, Waters and Forests (INRGREF) of Tunis (Tunisia) (latitude: 36° 50ʹ North, longitude: 10° 14ʹ East, altitude: 3 m). The resulting three years old trees were transferred to pots filled with 9 kg of soil (mixture of sand and loamy soil (1/4)). The experiment was conducted for 60 days (July-August 2021) in a completely randomized design with different field capacities (25, 50, 75, and 100%) and three replicates. The weather in July and August 2021 in the experimental location was characterized, respectively by an average of 28 and 29 °C, in the early morning, 34 and 36 °C at midday, and 41 and 45 °C at the end of the day. The temperature was recorded at 41 and 45 °C in July and August, respectively (https://www.historique-meteo.net).
Growth parameters
The height of the plant, diameter of the principal stem, and leaves parameters (Fresh weight, dry weight, area, length, width, and perimeter) were determined at t0 (3 years aged trees), t1 (after 15 days), t2 (after 30 days), t3 (after 45 days) and t4 (after 60 days), in plants grown under different drought stress conditions. The different leaf parameters were measured using a planimeter (model CI-202, USA). At the end of the experimental period, five separate plants subjected to the different field capacities were used to determine leaves, stems, and roots’ fresh weight, dry weight, length, diameter, and fluorescence measurements. Dry weight was determined following drying in an oven at 60 °C for 72 h.
Relative growth rate (RGR, mg. g− 1. d− 1) was determined following equation:
w0 and t0 are the shoot weight and time at the beginning of the experiment, and w4 and t4 were shoot weight and time at the end of the experiment, respectively.
Net assimilation rate, (NAR, g. m–2. d–1) was calculated following the following equation:
w0 and w4 are the dry weight of biomass; dt per unit time; LA0 and LA4 are the total leaf area and t0 and t4 are the time at the beginning and the end of the experiment, respectively.
Relative leaf area expansion rate, (RGRA, mm− 2.d− 1) was determined following the equation:
LA0 and LA4 are the total leaves area; t0 and t4 are the time at the beginning and the end of the experiment, respectively.
The relative growth rate of the principal stem in height (RGRh, cm. cm–1.d–1) was calculated according to the following formula:
h0 and h4 are the stem height; t0 and t4 are the time at the beginning and end of the experiment, respectively.
Determination of relative chlorophyll content and fluorescence measurements
The relative chlorophyll concentration (SPAD) was evaluated at the end of treatments using the chlorophyll meter (model SPAD-502, Minolta, Japan). The measure of the maximum quantum efficiency of PSII photochemistry (Fv/Fm) was assessed in the same leaves after 30 min. of dark adaptation, using modulated chlorophyll fluorometer (OS1p, Opti-Sciences Inc, Hudson, USA).
Determination of MDA and H2O2 contents
The lipid peroxidation level of foliar tissues was determined as MDA (Malondialdehyde). To do, concentration of thiobarbituric acid-reacting substances (TBARS) was measured spectrophotometrically at 535 nm, as described by Alia et al. [21].
The content of hydrogen peroxide (H2O2) in leaves was determined based on the modified method of Patterson et al. [22]. Colorimetric method (λ = 508) was used to determine H2O2 content, using H2O2 as a standard.
Antioxidant activity: FRAP (ferric reducing antioxidant power) and DPPH (2, 2’- diphenyl-1- picrylhydrazyle) radical-scavenging activity assays
Hinneburg [23] method was used to realise FRAP test. Leaves extracts were incubated (At 50 °C for 30 min) in presence of phosphate buffered saline PBS and K3Fe (CN)6. After that, Trichloric acid (10% TCA) and FeCL3 (0.1%) were added, respectively in which the reduction of ferric ion (Fe3+)-ligand complex to the brightly blue ferrous (Fe2+) complex by antioxidants in an acidic solution. 30 min. later, optical density was determined at 700 nm. Results were expressed in mmol ascorbic acid equivalent per gram of dry extract (mmol AAE/g).
DPPH anti-radical activity was carried out according to Parejo et al. [24] proceed. Each foliar extract (at different concentrations) was incubated with DPPH reactive for 30 min. To determine DPPH reduction, the absorbance of the DPPH solution as well as samples were measured at 517 nm. These optical densities were used for determination of sample concentration needed to neutralize 50% of radicals (IC50).
Total phenolic, total flavonoids, and condensed tannins contents quantification
Extraction of secondary metabolites was realised by maceration (methanol, 70%) for 48 h. After filtration on Whatman paper No.1, the filtrates obtained were evaporated at using a rotary evaporator at 60 °C. The residues of this filtrate were dried in an oven for 48 h at 45 °C to obtain the extracts dry.
Total phenolic content was measured using the Folin–Ciocalteu method described by Li et al. [25]. Total phenolic content was expressed as gallic acid mg. g− 1 dry weight (mg GAE.g− 1). Flavonoid content was carried out according to the method of Dehpeur et al. [26]. Contents were expressed in mg quercetin equivalent per gram of dry weight (mg Qu. g− 1). Total condensed tannins were estimated according to Ba et al. [27]. Contents were expressed in mg catechol equivalent/g dry weight (mg CAT. g− 1).
Proline and soluble sugars contents determination
Proline analysis was performed according to the protocol described by Bates [28] and expressed as µmol. g− 1 DW. Each extract was added to glacial acetic acid and Ninhydrin reagent. The mixture was placed in water bath at 100 °C for 30 min. After that, toluene was added, and the absorbance was determined spectrophotometrically at 520 nm.
Total soluble sugars were fixed according to Dubois et al. [29] method. Contents were estimated by Anthrone reagent and defined as µmol. g− 1 DW. The mixture was placed in water bath (100 °C) for 10 min. The absorbance was spectrophotometrically measured at 630 nm.
Antioxidant enzymatic extraction and assays
Enzyme extractions were carried out at 4 °C. The fresh plant tissues were powdered in liquid nitrogen and extracted at a ratio of 1:3 (w/v) fresh weight. Protein content was determined by Bradford’s method [30] using bovine serum albumin (BSA) as the standard. SOD; superoxide dismutase (EC: 1.15.1.1) activity was determined by the photochemical method according to Beyer and Fridovich [31] (1987). Catalase (CAT; EC: 1.11.1.6) activity assay was recorded following the method of Cakmak [32]. Ascorbate peroxidase (APX; EC: 1.11.1.11) activity was measured according to Nakao and Asada [33] by monitoring the rate of ascorbate oxidation at 290 nm (E = 2.8 mM− 1 cm− 1). Glutathione reductase (GR; EC: 1.6.4.2) activity was determined by monitoring the GSH-dependent oxidation of NADPH at 340 nm according to Rao et al. [34]. Peroxidase (GPX; EC: 1.11.1.9) activity was measured by the increase in absorbance at 470 nm caused by guaiacol oxidation according to Decleire et al. [35].
Statistical analysis
All data were subjected to one way ANOVA analysis based on Tukey’s HSD test (IBM SPSS Statistics 23) to determine the significant differences between means of treatments at a probability level of 0.05. All data were evaluated for normality using Shapiro-Wilk at the 0.05 level prior to statistical analysis, and if needed, the data were transformed. A randomised complete block design with three replications was used to gather the data. Every parameter under study was given its mean (± standard deviation). For the graphical representation of morpho-physiological characteristics, the JMP® ver. 17.2.0 cell plot was employed (SAS Institute Inc., Cary, NC, USA, 2022–2023). Principal Component Analysis (PCA) was used to examine the contribution of the combination of morpho-physiological, biochemical, and metabolic characteristics to better summarize the data and comprehend the conclusions. For additional analysis, components with an Eigen value larger than one were retained. Scatter plot matrix with heatmap correlation circles of 29 quantitative traits were calculated. The similarity matrix was produced by employing the hierarchical co-clustering (dendrogram) utilizing the ward method and a heatmap based on all the data. These studies were performed with JMP® version 17.2.0.
Results and discussion
Morpho-physiological traits
Abiotic stresses such as drought, affect negatively every growth stage of many crops by reducing leaf production, photosynthesis, and yield [36]. Cell plot results of 15 quantitative morpho-physiological traits showed tremendous variations of M. oleifera trees under different field capacity regimes (Fig. 1). An analogous trend in the root length of Moringa plants was noted. Under the extreme field capacity (25% FC), all the quantitative traits showed the lowest values, which marked in green, in different tissues (stem, root and leaves) except root length parameter which recorded the highest value (remarked as red). Also, under 50% FC, all M. oleifera parameters showed intermediate color and values except stem fresh weight and root length parameters obtain had values near maximum and remarked as faint red color. In the contrast, under 75 and 100% showed the highest values of all parameters which marked with red color and root length (remarked with green) showed the contrary value of the tested parameters of 25% FC as illustrated in Fig. (1).

Cell plot of 15 morpho-physiological traits of stem, root, leaf parameters, relative chlorophyll content and fluorescence measurements (Fv/Fm efficiency) of Moringa oleifera trees submitted to different field capacities (25, 50, 75, and 100%). Red color assumes the highest value, while the green color assumes the lowest ones
On the other hand, data showed a reduction of the shoot and roots’ fresh and dry weights (Supplementary Fig. 1), principal stem height, and diameter under accentuating situation of water stress (25% FC). In contrast, a dramatic enhancement of root length was noted in trees submitted to 25% FC (Supplementary Fig. 1G). The impact of water unavailability on M. oleifera growth and development was reflected by other morphological parameter changes. When low water availability was applied in a culture medium (25%), the shorter plant height and less stem diameter were recorded (Supplementary Fig. 1E and F).
This regression of height and diameter was the results of cells elongation, turgor, and volume reduction as described by Heng et al. [37] and Ximeng et al. [38]. More that, slower organs growth under stress condition is a strategy adopted to acclimate to the stress conditions [38]. In contrast, when the water amount was very limited (25% FC), roots are longer compared to those in control plants (Supplementary Fig. 1G). This responsible absorption water organ showed a more developed expansion and proliferation attributed to the necessity to search and acquisition of water in the soil under low osmotic pressure. Also, the morphological and traits changes of roots, explain the Moringa tree orientation to a more water and nutrients conservative strategy.
Far, no senescence symptoms were observed in all our trees subjected to the experimental protocol. For this reason, we have measured the maximum quantum efficiency of PSII (Fv/Fm) in Moringa leaves at the end of the experiment period. We found that this parameter was reduced by water lack (Supplementary Fig. 1H). In fact, PSII efficiency was under regulation of the quantum yield (Fv/Fm) to predicts plant development under various environmental conditions [39]. It is true that the decrease of PSII efficiency caused by abiotic stress resulted in photoinhibition and cause photodamage [14, 40]. However, in another way, the diminution of Fv/Fm is another benefit response presented by plants to minimize photosynthetic activity under stress factor [39].
Crop plants exhibit a variety of drought stress adaptation and acclimatization mechanisms, ranging from ostensibly straightforward physiological or morphological characteristics that function as critical stress tolerance [41]. To encompass, we can suggest that the origin of tolerance capacity to water stress of M. oleifera, is identified by the maintenance of clear photosynthetic assimilation at the level of the leaves, monitoring height of shoot part, and enhancement of roots elongation.
Growth response
Several key indices were determined as an aid to understanding the growth response of M. oleifera trees face to the different water-holding capacities applied during the experiment period. In the beginning, relative growth rate (RGR) was determined, because it is useful for analyzing whole plant response to the abiotic stress factor fixed in this study (Fig. 2). Under severe water unavailability (25% FC), RGR is in order of 0.016 mg. g− 1d− 1, which showed 70% decline compared with the positive control (100%) (Supplementary Fig. 2A, E). Secondly, these results showed that the relative leaf expansion (RGRA) (Fig. 2B, E) was considerably reduced, especially in plants subjected to water restriction (25% FC) by − 71.78%. This may explain, first, the drop in net biomass gains which presents the result of the assimilation of CO2 by the leaves minus the respiratory loss by the whole plant. Indeed, the net CO2 assimilation rate (NAR) was found to be reduced by (0.01 and 0.004 g− 1. d− 1) which represented the declining percentage of 62.7 and 83.2, respectively when the plants were grown at field capacities of 50 and 25% (Fig. 2C, E). For further evidence, it was also found that the relative growth rate in height (RGRh) of the principal stem was inhibited by 51.33% from 0.004 mm− 1. d− 1 (control condition) to 0.002 mm− 1. d− 1 under severe drought conditions (25% FC) (Fig. 2D, E). The link between growth modifications and interior leaf biological (cell division and expansion) and physiological (photosynthesis) processes under conditions of water deprivation can help us to understand these findings. Numerous authors have correctly pointed out that leaves primarily regulate a plant’s ability to withstand drought [38].

Scatter plot matrix of variation in the cumulative average of final growth rates (%): Relative growth rate (RGR mg. g− 1.d− 1), Relative leaf area expansion rate, (RGRA mm− 2.d− 1), Relative growth rate of the principal stem in height (RGRh, cm. cm–1.d–1) and Net assimilation rate, (NAR g. m–2.d–1) of M. oleifera trees submitted to different field capacities (25, 50, 75 and 100%)
To confirm this previous suggestion, leaf dimensions, which are the major driving variables for photosynthetic machinery and whole-plant growth, were followed and measured for 60 days of applied drought stress. Table 1 demonstrated that there was a non-significant difference in the various leaf measurements between 75% and 100% FC at P ≤ 0.05 level. Whereas, compared to the control condition, a 25% FC provoked a net reduction of these parameters at the four times (t1, t2, t3 and t4) set for taking the measurements. We found that, after 60 days (t4), extreme field capacity (25% FC) induced reduction of leaf fresh (-47.07%) and dry weights (-37.34%), area (-53.4%), length (-67.34%), width (-43.93%), and perimeter (-51.77%). These leaves’ morphological adjustments generated by the water constraint condition, resulted in growth performance and adaptability to this stress factor [37, 39, 40]. The limitation in leaves expansion resulting in stomatal closure is an adaptative tolerance mechanism included in photosynthetic machinery protection under such environmental condition.
Antioxidant activity, membrane lipid peroxidation, secondary metabolites and osmolytes contents
The ability of antioxidants to scavenge free radicals is measured by the 2,2 diphenyl-b picrylhydrazyl (DPPH) radical scavenging activity assay [42]. It was revealed that FRAP and DPPH assays are often used to evaluate the ability of the extract to scavenge free radicals produced because of the induced-oxidative stress biological damage [15]. The compound’s ability to stop b-carotene from oxidising was determined by Chitiyo et al. [43] using the b-carotene-linoleic acid model system, which demonstrated a higher antioxidant capacity in M. oleifera (75% FWC) under mild stress. Our agreed data showed that the highest total antioxidant activity (FRAP and DPPH) and radical scavenging activity corresponding to the lowest IC50 value was marked in leaves extract derived from plants grown under the reduced capacity field (25% FC) (Table 2). Furthermore, these same plants demonstrated the greatest ability to scavenge ROS. The variability observed in the relationship between polyphenolic compounds and antioxidant capacities as indicated by the different antioxidant measurements may be attributed to the fact that biological activity is influenced by the elevated concentration of total secondary metabolites (phenolic compounds) [44].
More than that, a positive relationship was observed between the FRAP and DPPH activities increase, and MDA and H2O2 accumulation at the different water holding capacities. Increasing drought stress levels carries the antioxidative potential for chain-breaking inhibition of lipid peroxidation scavenging as previously suggested by Nasraoui et al. [45] and Abdul Waheb et al. [19]. On the other hand, the high antioxidant capacity detected may be attributed to the cooperative effects of antioxidant activity (DPPH) and high total phenolic, flavonoids, and tannins contents (Table 2).
According to McMahon et al. [46], condensed tannins are a product of flavonoid polymers. Popovic et al. [47] found that drought-induced slightly greater amounts of condensed tannins were found in poplar trees. Moreover, transgenic poplar lines with high condensed tannin content showed decreased levels of hydrogen peroxide, photosystem II damage, and leaf death [48]. This makes sense since it demonstrates a clear link between high tannin content and antioxidant capacity.
Furthermore, phenolic compounds contents of edible plants may be considered an inducement of their antioxidant capacity as showed by several researchers [13, 15,16,17], phenolics compounds have been shown to act as a photoprotector by blocking the stimulation of chlorophyll under conditions of moisture stress by decreasing light absorption [49]. The second harvest of Moringa oleifera under extreme stress had the highest concentrations of total phenolic compounds (35% FWC) and proanthocyanidins (50%) for flavonoids [43].
Osmotic stress and the other different abiotic stress factors (salinity, metallic) induce the synthesis and accumulation of osmoregulator molecules such as proline amino acid and soluble sugar [50]. It is largely demonstrated that proline participates in reinforcement of the antioxidant system scavenging induced ROS accumulation [19]. According to our data, Chitiyo et al. [43] found that in drought stressed Moringa proline and soluble sugar contents increased with the severity of moisture stress in all plant parts (root, stem and leaf), with the leaves under severe stress (35% FWC) recording the highest concentration. A strong correlation of these compounds’ accumulation and higher radical scavenging activity against FRAP and DPPH radical were detected confirming their role in antioxidant defense potential of Moringa [15, 51]. This corroborates the results of the present investigation, which demonstrated a positive correlation between antioxidant activity, secondary metabolites and osmolytes amounts. In addition to having the highest concentrations of condensed tannins and total phenolic compounds which are known to rise with an elevation in phenolic concentration in the severely stressed (25% FC) plants also showed the highest proline and soluble sugar tenors ferric reducing capability. This suggests that M. oleifera’s antioxidant properties are enhanced by moisture stress. By raising the amounts of antioxidant substances such as total phenols, and flavonoids, drought stress improved the therapeutic potentials of Moringa. In fact, free radicals can be chelated by total phenolics and flavonoids before they damage cells [52]. Briefly, M. oleifera drought tolerance is achieved through metabolite pattern alteration, which ensures good cellular activities.
Antioxidant enzyme activity
The crucial role in the reduction of reactive oxygen species generation and resistance to oxidative damage by activation of a defensive antioxidant system in stressed plants is largely confirmed in other studies [44, 50]. The stimulation of antioxidant enzymes (SOD and CAT) to react more strongly to freshwater deficiency stress was triggered by the first water deficit cycle in control stressed plants. These findings suggest that these plants are best protected in the peroxisome, where CAT breaks down H2O2 produced by photorespiration [53].
Accordingly, our results (Fig. 3A-E) demonstrated that superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR), and glutathione peroxidase (GPX) activities were significantly stimulated. These increases were 76.15%, 66.74%, 43.26%, 79.30 and 22.2%, respectively in Moringa leaves stressed with 25% FC (Fig. 3F). Similar activations by stresses factors such as drought and salinity were demonstrated in other several studies [15, 51]. However, the astounding equilibrium between ROS production and antioxidative enzyme system activation reduces the detrimental effects of oxidative stress and increases the powerful ROS detoxifying capacity of the Moringa tree. This biochemical approach is an important tool for resisting, defending, and surviving in hard climate circumstances, such as abiotic stress.

Relative (A, B, C, D, E) and percentage (F) changes of antioxidant enzyme activity: superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR), and glutathione peroxidase (GPX) activities in M. oleifera leaves submitted to different field capacities (25, 50 and 75%) compared to control condition (100%). Data represented as mean with error bars having different significant letters according to Tukey’s HSD test
Inter-correlation among all data of M. oleifera trees under different field capacity regimes: multivariate analysis
Multivariate analysis approaches can be utilized to investigate correlations, categorization, and parameter prediction within complicated data sets since the results are more realistic, meaningful, and accurate [54].
Hierarchical co-clustering with heatmap
Multivariate analysis based on hierarchical co-cluster analysis using Ward’s method was performed to identify the distribution of the M. oleifera trees due to differences in morpho-physiological, biochemical and all combined data under different field capacities (25, 50, 75, and 100%) into two distinct clusters as represented in Fig. 4. Correlograms were based on the correlation coefficients of morphological and/or metabolic measurements. The red color indicated the positive correlation between measurements, whereas the blue color assumed the negative one.
In the presence of outlying observations in the dataset, the resilient hierarchical co-cluster (RHCOC) strategy generates a significantly lower clustering error rate than standard hierarchical clustering algorithms [55].
As shown in Fig. 4A, the hierarchical co-cluster dendrogram based on morpho-physiological parameters differences is divided into two clusters. The first cluster had M. oleifera trees under 25 and 50% FC and the second one is included M. oleifera trees under 75 and 100% FC. Under 75 and 100% FC, red positive correlation was found between all parameters except root length parameter (remarked as blue). Low water availability was favourable for the growth of Moringa plants. The plants displayed reduced height when they were fully capable of retaining water. In fact, reduced osmotic pressure during stressful situations, which causes root proliferation in quest of water and nutrients, may be the cause of the increase in root length at 70 and 40% water holding capacity [14].
At the same trend, the hierarchical co-cluster dendrogram based on biochemical and metabolic parameters differences is divided into two clusters. The first cluster had M. oleifera trees under 25% FC and the second one is divided into two sub-clusters. The sub-cluster I included M. oleifera trees under 75 and 100% FC and the other one contained 50% FC. Under 75 and 100% FC, DPPH is negatively correlated (remarked blue) with all other parameters (remarked as red) as illustrated in Fig. 4).
A two-way co-cluster matrix using hierarchical co-clustering dendrogram and heatmap; row clusters were obtained at field capacity level, whereas the column cluster were recorded at trait or marker of all combined data of M. oleifera trees submitted to different field capacities (Fig. 4C). The co- cluster was consisted of 4 rows (treatments) and 29 column clusters (traits). At the row clustering (FC level), it is divided to two clusters. The first cluster had M. oleifera trees under 25% FC and the second one is classified into subclusters I and II. The subcluster I had M. oleifera trees under 50% FC and the other subcluster II included M. oleifera trees under 75 and 100% FC. At the column clustering (parameters), it is divided into two major clusters. The first cluster had the following parameters: root length, H2O2, CAT, APX, MDA, phenols, FRAP, proline, sugars, SOD, GR, flavonoids, and tannins. On the other hand, the second cluster had stem, root, leaf fresh and dry weight, stem diameter and height, leaf length, width, foliar area, chlorophyll content, Fv/Fm and DPPH. Sun et al. [56] reported progressive drought stress generally caused an elevated in activity of SOD, POD, CAT, and APX enzymes that coordinate ROS concentration.

Multivariate analysis based on hierarchical dendrogram and heatmap correlation between (A) morpho-physiological parameters, (B) biochemical parameters and enzymatic and non-enzymatic antioxidants, (C) Two-way clustering using hierarchical co-clustering dendrogram and heatmap; row clusters were obtained at field capacity level, whereas the column cluster were recorded at trait or marker of all combined data of morpho-physiological, biochemical parameters and enzymatic and non-enzymatic antioxidants) of M. oleifera trees submitted to different field capacities (25, 50, 75 and 100%). Correlation levels are colored by red for positive correlation and blue for negative one
Scatter plot matrix with correlations circles
Scatter plot matrix with heatmap correlation circles of 29 quantitative traits (morphological, physiological, and metabolic measurements) of M. oleifera trees submitted to different field capacities (25, 50, 75, and 100%). The scatter plot matrix showed the density ellipses in each individual scatter plot and the red circles contain about 95% of the data as represented in (Fig. 5). Red color assumed the positive correlation, blue colour assumed the negative correlation, while the size of circles indicated the significance. In this investigation, root length was negatively correlated with all morpho-physiological parameters, but it was positively correlated to all biochemical and metabolic parameters except DPPH. Highly positive correlation was detected among root length, MDA, H2O2, phenols and FRAP.

Scatter plot matrix with heatmap correlation circles of all combined data (morphological, physiological, and metabolic measurements) of M. oleifera trees submitted to different field capacities (25, 50, 75 and 100%). Red color indicates positive correlation, blue colour indicates the negative correlation, while the size of circles indicates the significance (see scale at the above right corner)
On contrary, DPPH was positively correlated to all morpho-physiological parameters except root length and was negatively correlated with all biochemical to FRAP, MDA and phenols. In the same trend, MDA was strongly positive correlated with H2O2, proline, phenols and FRAP. Also, Fv/Fm showed the maximum positive correlation to chlorophyll content, and it also correlated positively with all leaf parameters. The adaptive response of Moringa to drought stress caused by strong accumulation by endogenous proline and an increase in the total polyphenol and radical scavenging activity. A strong correlation of these compounds’ accumulation and higher radical scavenging activity against FRAP and DPPH radical were detected confirming their role in antioxidant defense potential of Moringa [15, 51].
Concerning the enzymatic antioxidants, all enzymes were positively correlated with each other. CAT was highly positive correlate to APX and H2O2 in the present study. Plants that are tolerant to drought may display modifications in their cellular metabolism as a defense against oxidative damage. The principal enzymes that eliminate H2O2 from leaves are SOD, CAT and APX. While both CAT and APX enzymes work together, APX is more probable to bind to H2O2 than CAT does. The moderate stressed plants displayed elevated enzymes activities (SOD, CAT, and APX) at the first maximum stress; only the SOD activity persisted as elevated at the second maximum stress. Peroxisomes and chloroplasts, the primary sites of action of APX, may have provided enhanced protection because of this treatment [53].
Principal component analysis (PCA) biplot
Principal component analysis (PCA) is used to analyze and deconstruct large, complex datasets. The only PCAs deemed significant were those with eigenvalues larger than one. The item diversity was analyzed in terms of quality features using the PCA technique, and the objects were grouped based on the similarity hierarchy [57]. PCA-biplot is one of the most efficient multivariate techniques for evaluating trait interaction and genotypic performance, and it is widely used to investigate trait association in various agricultural plants [58].
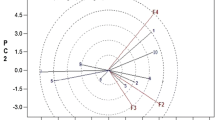
In a PCA scatter plot, M. oleifera trees submitted to different field capacities (25, 50, 75, and 100%) described by the first 2 PCs per cluster. The first component PC1 was recorded as 96% and the second component PC2 as 2.94% of the total variation. The studied treated M. oleifera trees were also grouped into three clusters: cluster I (green group) had treated M. oleifera trees subjected to 75 and 100% FC, cluster II (blue group) had M. oleifera trees subjected to 25% FC and cluster III (red group) had M. oleifera trees subjected to 50% FC which were distinct and well supported as illustrated in Fig. 6A.
In the system of the two first components, length of vector and cosine of angle were used for discrimination of treated M. oleifera trees. The small angle between these vectors: stem dry weight, foliar area, root dry weight, leaves fresh weight, stem fresh weight, Fv/Fm proves a significant strong positive correlation between these traits and were found as the most effective parameters in differentiation between M. oleifera trees under 75 and 100% FC. On the other hand, flavonoids, tannins, sugars, CAT, SOD, GR were found as the most effective parameters in differentiation between M. oleifera trees under 25% FC. Flavonoids and tannins had an acute angle which indicated highly positive correlation to each other. In the same manner, GR with SOD and GPX and proline with phenolic compounds Fig. 6 B. The multivariate analysis confirmed the correlation data reported before.
Previous studies have successfully evaluated drought tolerance indicators for wheat [59] and cotton [60] by combining principal component analysis and/or regression analysis. Also, Wang et al. [61] examined 12 physiological indicators related to wheat drought tolerance, including photosynthetic characteristics (Pn, Gs, Fv/Fm, qP, NPQ, and qN), SOD activity, and MDA content, as well as osmoregulatory substances (Pro, SS, HXKs, and GLC), were selected to further screen physiological key indicators to evaluate drought tolerance in wheat. In maize, Balbaa et al. [62] used discriminative analyses such as PCA to support their methodology as an unambiguous differential approach, indicating that chlorophyll content and transpiration rate traits were significant on maize inbred line performance under stress conditions, while other remained traits were most discriminatory under normal conditions.
Conclusion
Water stress is on the rise these days due to changes in the global climate, which is a major issue that is detrimental to plant cover. Moringa oleifera trees demonstrated the ability to tolerate this type of abiotic stress, responding in a number of ways that were responsive to the water-deficit state. To withstand the conditions of drought stress, root morphological modifications compensated the limitation of tree shoot development. First and foremost, it is important to emphasise that M. oleifera’s ability to adjust to the water unavailability conditions (25, 50, 75 and 100% field capacities, it was subjected to was evidenced by its regression of growth, the dimensions of its aerial organs and the elongation of its roots. Furthermore, we have found important connections between the activation of the defence antioxidant system (superoxide dismutase (SOD), catalase (CAT), glutathione reductase (GR), ascorbate peroxidase (APX) and glutathione peroxidase (GPX) enzymes), hyper-accumulation of secondary metabolites (phenolic compounds, total flavonoids and condensed tannins) and osmoregulators (proline and soluble sugars), and osmotic stress brought on by a lack of water. A powerful antioxidant radical scavenger (FRAP and DPPH) that was enhanced by shortage water was another important component of stress adversity adoption. Furthermore, M. oleifera shows potential as an ecological solution for agroforestry systems, particularly in dry locations where water scarcity is an issue. Further research can explore the ability of different Moringa parts to stimulate plant growth under varied water stress scenarios.

Principal component analysis (PCA), (A) the PCA scatter plot, and (B) biplot illustrating the distribution of different field capacity regimes on M. oleifera trees, on PC1 and PC2 components based on the analysis of all combined data (morphological, physiological, and metabolic measurements). Blue, red and green dashed rings showed three different clusters. The dots were M. oleifera trees under different regimes of FC, and the vectors (red arrows) were parameters. The abbreviations were previously mentioned in the earlier figures
Data availability
Data is provided within the manuscript or supplemetary information files.
References
Rocha J, Quintela A, Serpa D, et al. Water yield and biomass production for on a eucalypt-dominated Mediterranean catchment under different climate scenarios. J Res. 2023;34:1263–78.
Yujun H, Yuechao W, Dongmei Z, Hong G, Ying S, et al. Planting models and deficit irrigation strategies to improve radiation use efficiency, dry matter translocation and winter wheat productivity under semi-arid regions. J Plant Phys. 2023;280:10.
Yadav B, Jogawat A, Rahman MS, Narayan OP. Secondary metabolites in the drought stress tolerance of crop plants: a review. Gene Rep. 2021;23:101.
Azam S, Nouman W, Ubaid-ur Rehman U, Ahmed T, Gull M. Shaheen. Adaptability of Moringa oleifera Lam. Under different water holding capacities. South Afr J Bot. 2019;11:123.
Ashutosh P, Malvika P, Madan MG, Pushpa K, Yashumati R, Vivek J, Aaushi P, Anil AC. Moringa oleifera: an updated comprehensive review of its pharmacological activities, ethnomedicinal, phytopharmaceutical formulation, clinical, phytochemical, and toxicological aspects. Int J Mol Sci. 2023;24:2098.
Sultana S. Nutritional and functional properties of Moringa oleifera. Metabolism Open. 2020;8:2589.
Batool S, Khan S, Basra SM, Hussain M, Saddiq MS, Iqbal S, et al. Impact of Natural and Synthetic Plant Stimulants on Moringa seedlings grown under low-temperature conditions. Inte Lett Nat Sci. 2019;76:50.
Páramo-Calderón DE, Aparicio-Saguilán A, Aguirre-Cruz A, Carrillo-Ahumada J, Hernández-Uribe JP, Acevedo-Tello S, Torruco-Uco JG. Tortilla added with Moringa oleífera flour: physicochemical, texture properties and antioxidant capacity. Food Sci Techn. 2019;100:409.
Yosvany DD, Ramón PR, Danger TG. Biological activities of Moringa seeds oil. Multiple Biological activities of unconventional seed oils. Academic; 2022. pp. 55–75.
Khan RU, Khan A, Naz S, Ullah Q, Laudadio V, et al. Potential applications of Moringa oleifera in poultry health and production as alternative to antibiotics: a review. Antibio. 2021;10:1540.
Yap YK, El-Sherif F, Habib ES, Khattab S. Moringa oleifera leaf extract enhanced growth, yield, and Silybin content while mitigating salt-induced adverse effects on the growth of Silybum marianum. Agro. 2021;11:1.
Yaseen AA, Takacs-Hajos M. Evaluation of moringa (Moringa oleifera Lam.) Leaf extract on bioactive compounds of lettuce (Lactuca sativa L.) grown under glasshouse environment. J King Saud Uni Sci. 2022;34:101.
Chuene VM, Bonga LN, Alen M, Isa B, Olaniyi AF. Assessing the usefulness of Moringa oleifera Leaf Extract as a Biostimulant to supplement synthetic fertilizers. Plants. 2022;11:2217.
Geber GG, Hussein MB, Amsalu GR. Physiological responses of Moringa (Moringa Stnopetala L.) seedlings under greenhouse conditions, Southern Ethiopia. Asi J Biotech. 2020;12:97.
Azeem M, Kulsoom P, Muhammad Q, Athar M, Talha J, Haji M, et al. Salinity stress improves antioxidant potential by modulating physio-biochemical responses in Moringa oleifera Lam. Sci Rep. 2023;13:2895.
Seleiman MF, Al-Shuaibani N, Ali N, Akmal M, Alotaibi M, et al. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants. 2021;10:259.
Hasanuzzaman M. Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antiox. 2020;9:68.
Nouman W, Basra SMA, Yasmeen A, Gull T, Hussain SB, Zubair M, Gul R. Seed priming improves the emergence potential, growth and antioxidant system of Moringa oleifera under saline conditions. Plant Grow Reg. 2014;73:267.
Abdul Wahab K, Golamreza A, Muhammad HS, Naer A, Saqib U, Wadood S, Sahar M, Ghulam Y, Crina CM, Romina AM. Plants’ physio-biochemical and phyto-hormonal responses to alleviate the adverse effects of drought stress: a comprehensive review. Plants. 2022;11:1.
Peiman Z, Ewald S. Reactive oxygen species, antioxidant responses and implications from a Microbial Modulation Perspective. Biol (Basel). 2022;11:155.
Alia KVSK, Prasad P, Pardha SP. Effect of zinc on free radicals and proline in Brassica and Cajanus. Phytochem. 1995;42:45.
Patterson BDA, MacRae E, Ferguson EA. Estimation of hydrogen peroxide in plant 387 extracts using titanium (IV). Anal Bioch. 1984;139:487.
Hinneburg I, Damien DHJ, Hiltunen R. Antioxidant activities of extracts from selected culinary herbs and spices. Food Chem. 2006;97:122.
Parejo I, Francesc V, Jaume B, Alfredo RR, Nadine F, et al. Comparison between the radical scavenging activity and antioxidant activity of six distilled and no distilled Mediterranean herbs and aromatic plants. J Agr Food Chem. 2002;50:6882.
Ainswort EA, Gillespie KM. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat Proto. 2007; 2(4):875-7. https://doi.org/10.1038/nprot.2007.102. PMID: 17446889.
Dehpour AA, Ibrahimzadeh MA, Fazel NS, Mohammad NS. Antioxydant activity of the methanol extract of Ferula Assafoetida and its essential oil composition. Grasas Aceites. 2009;60:405.
Ba K, Tine E, Destain J, Cisse N, Thonart P. Étude comparative des composés phénoliques, du pouvoir antioxydant de différentes variétés de sorgho sénégalais et des enzymes amylolytiques de leur malt. Biotech Agro Soc Environ. 2010;14:131.
Bates LS, Walden RP, Tears ID. Rapid termination of free proline for water stress studies. Plant Soil. 1973;39:205.
Dubois M, Gilles KA, Hamilton JK, Reders PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Bioch. 1956;28:350.
Bradford MM. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein–dye binding. Anal Bioch. 1976;72:248.
Beyer WF, Fridovich Y. Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem. 1987;16:559.
Cakmak I, Strbac D, Marshner H. Activities of hydrogen peroxide-scavenging enzymes in germinating wheat seeds. J Exp Bot. 1993;44:127.
Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Phys. 1981;22:679.
Rao MV, Paliyath G, Ormrod DP. Ultraviolet-B- and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Phys. 1996;110:25.
Decleire MM, Honorze YP, Van Roey GV. Activité Des peroxydases, catalases et glycolase oxydases après traitement avec divers herbicides. Weed Res. 1982;285.
David-Rogeat N, Broadley MR, Stavridou E. Solanum crops with a focus on the African eggplant: the potential of biostimulants to enhance stress tolerance. Int J Plant Soil Sci. 2023;35(18):1425–51.
Heng Z, Yang Z, Jian-Kang Z. Thriving under stress: how plants balance growth and the stress response. Develop Cell. 2020;55:529.
Ximeng L, Jingting B, Jin W, Chris B, David T. Antecedent drought condition effects responses of plant physiology and growth to drought and post-drought recovery. Front Glob Change. 2021;4:1.
Ayyaz A, Amir M, Umer S, Iqbal M, Bano H, Gul HS et al. Melatonin induced changes in photosynthetic efficiency as probed by OJIP associated with improved chromium stress tolerance in canola (Brassica napus L.), Heliyon. 2020; 6:1.
Gessler A, Bottero A, Marshall J, Arend M. The way back: recovery of trees from drought and its implication for acclimation. New Phyt. 2020;228:1704.
Bhargava S, Sawant K. Drought stress adaptation: metabolic adjustment and regulation of gene expression. Plant Breed. 2013;132(1):21–32.
Atoui AK, Mansouri A, Boskou G, Kefalas P. Tea and herbal infusions: their antioxidant activity and phenolic profile. Food Chem. 2005;8927.
Chitiyo ST, Ncube B, Ndhlala AR, Tsvuura Z. Biochemical responses of Moringa oleifera Lam. Plants to graded moisture deficit. South Af J Bot. 2021;138:41.
Dania SO, Akpansubi P, Eghagara OO. Comparative effects of different fertilizer sources on the growth and nutrient content of moringa (Moringa oleifera) seedling in a greenhouse trial. Adv Agric. 2014;1:22.
Nasraoui HA, Gouia H. ROS scavenging and NH4+ nitrogen fertilizer roles in alleviation of Cd-induced oxidative stress in Arabidopsis thaliana. Russ J Plant Phys. 2019;66:495.
McMahon LR, McAllister TA, Berg BP, Majak W, Acharya SN, et al. A review of the effects of forage condensed tannins on ruminal fermentation and bloat in grazing cattle. Canad J Plant Sci. 2000;80:469.
Popovic BM, Stajner D, Zdero-Pavlovi CR, Tumbas-Saponjac V, Canadanovic-Brunet J, Orlovic S. Water stress induces changes in polyphenol profile and antioxidant capacity in poplar plants (Populus spp). Plant Phys Bioch. 2016;105:242.
Gourlay G, Constabel CP. Condensed tannins are inducible antioxidants and protect hybrid poplar against oxidative stress. Tree Phys. 2019;39:345.
Nogues S, Baker NR. Effects of drought on photosynthesis in Mediterranean plants grown under enhanced UV-B radiation. J Exp Bot. 2000;51:1309.
Osman AR, El-Naggar HM. Enhancing salinity stress tolerance and phenylalanine ammonia lyasegene activity using osmolytes in Moringa seedling production. Ann Agri Sci. 2022;67:127.
Bekka S, Tayeb-Hammani K, Boucekkine I, Aissiou M, Djazouli ZE. Adaptation strategies of Moringa oleifera under drought and salinity stresses. Ukr J Eco. 2022;12:8.
Santos AFS, Argolo ACC, Paiva PMG, Coelho LCBB. Antioxidant activity of Moringa oleifera tissue extracts. Phytoth Res. 2012;26:366.
Foyer CH, Noctor G. Oxygen processing in photosynthesis: regulation and signaling. New Phytol. 2000;146:359.
Böhm K, Smidt E, Tintner J. Application of Multivariate Data Analyses in Waste Management. In Multivariate Analysis in Management, Engineering and the Sciences; de Freitas, L.V., de Freitas, A.P.B.R., Eds.; IntechOpen: London, UK, 2013.
Hasan MN, Badsha MB, Mollah MNH. Robust hierarchical co-clustering to explore toxicogenomic biomarkers and their Regulatory doses of Chemical compounds. Bio Rxiv. 2020; 094946.
Sun J, Gu J, Zeng J, Han S, Song A, Chen F, Chen S. Changes in leaf morphology,antioxidant activity and photosynthesis capacity in two different drought-tolerant cultivars of chrysanthemum during and after water stress. Sci Hortic. 2013;161:249–58.
Gregorczy kA, Smagacz J, Stankowski S, Fiejtek A. Assessment of relations between the technological characteristics of winter wheat. Acta Sci Pol Agric. 2008;7:65.
Aslam M, Maqbool MA, Zaman QU, Shahid M, Akhtar MA, Rana AS. Comparison of different tolerance indices and PCA biplot analysis for assessment of salinity tolerance in lentil (Lens culinaris) genotypes. Int J Agric Biol. 2017;19:470.
Omar AA, Heikal YM, Zayed EM, Shamseldin SAM, Salama YE, et al. Conferring of Drought and heat stress tolerance in wheat (Triticum aestivum L.) Genotypes and their response to Selenium nanoparticles Application. Nanomat. 2023;13(6):998. https://doi.org/10.3390/nano13060998.
Sun F, Chen Q, Chen Q, Jiang M, Gao W, Qu Y. Screening of Key Drought Tolerance Indices for Cotton at the flowering and Boll setting Stage using the dimension reduction method. Front. Plant Sci. 2021;12:619.
Wang J, Zhang X, Han Z, Feng H, Wang, et al. Analysis of physiological indicators Associated with Drought Tolerance in Wheat under Drought and Re-watering conditions. Antioxid. 2022;11(11):2266. https://doi.org/10.3390/antiox11112266.
Balbaa MG, Osman HT, Kandil EE, Javed T, Lamlom SF, Ali HM, Kalaji HM, Wróbel J, Telesiñski A, Brysiewicz A, Ghareeb RY, Abdelsalam NR, Abdelghany AM. Determination of morpho-physiological and yield traits of maize inbred lines (Zea mays L.) under optimal and drought stress conditions. Front Plant Sci. 2022;13:959203.
Acknowledgements
The authors would like to express their gratitude to Forest Ecology Laboratory, National Research Institute in Rural Engineering, Water and Forestry, University of Carthage, Tunisia. Additionally, the authors are thankful for the Botany Department, Faculty of Science, Mansoura University, Egypt, for providing the necessary facilities.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
A. N. H. performed the supervision, writing, review, methodology and editing. R. A. F. H. and A.M. performed the methodology, writing editing. Y. M. H. performed the statistical analysis, data visualization and writing editing. A.Y. performed the review and the validation of the manuscript. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
We declare that the plant materials for experiments were gathered and studied in accordance with the Ethical Rules applicable to BMC Plant Biology.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hajaji, A.N., Heikal, Y.M., Hamouda, R.A.E.F. et al. Multivariate investigation of Moringa oleifera morpho-physiological and biochemical traits under various water regimes. BMC Plant Biol 24, 505 (2024). https://doi.org/10.1186/s12870-024-05040-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-05040-5