
Abstract
Background
Common bean (Phaseolus vulgaris) is an essential crop with high economic value. The growth of this plant is sensitive to environmental stress. Heat shock factor (Hsf) is a family of antiretroviral transcription factors that regulate plant defense system against biotic and abiotic stress. To date, few studies have identified and bio-analyzed Hsfs in common bean.
Results
In this study, 30 Hsf transcription factors (PvHsf1–30) were identified from the PFAM database. The PvHsf1–30 belonged to 14 subfamilies with similar motifs, gene structure and cis-acting elements. The Hsf members in Arabidopsis, rice (Oryza sativa), maize (Zea mays) and common bean were classified into 14 subfamilies. Collinearity analysis showed that PvHsfs played a role in the regulation of responses to abiotic stress. The expression of PvHsfs varied across different tissues. Moreover, quantitative real-time PCR (qRT-PCR) revealed that most PvHsfs were differentially expressed under cold, heat, salt and heavy metal stress, indicating that PvHsfs might play different functions depending on the type of abiotic stress.
Conclusions
In this study, we identified 30 Hsf transcription factors and determined their location, motifs, gene structure, cis-elements, collinearity and expression patterns. It was found that PvHsfs regulates responses to abiotic stress in common bean. Thus, this study provides a basis for further analysis of the function of PvHsfs in the regulation of abiotic stress in common bean.
Similar content being viewed by others

Background
Common bean (Phaseolus vulgaris) is an essential legume with high economic value [1]. Seeds of common bean are rich in lectin and α-Amylase inhibitors (α-AI) which are used to synthesize pesticides [2] and raw materials for preparation of chemicals [3]. The growth of common bean is sensitive to various abiotic stress stimuli, such as salt, cold and drought [4]. Therefore, it is necessary to improve the tolerance of common bean under abiotic stress.
Heat shock transcription factor (Hsf) regulates signal transductions associated with heat stress in plants [5]. Hsfs participate in signal reception and transmission, regulation of gene expression, resistance to stress and heat tolerance in plants. Members of the Hsf family have five conserved domains [6]: DNA-binding Domain (DBD), oligomerization domain, (OD), C-terminal activation domain (CTAD), nuclear localization signals (NLS), and nuclear export signals (NES) [7]. Among them, DBD and OD domain are the most evolutionary conserved and are used to accurately identify and combine the promoters of heat shock protein (Hsp) [8]. The C-terminal activation domain contains a repressor domain: LFGV-peptide [9], which directly regulates the expression of genes in response to heat shock. The NLS and NES modulate the transnuclear transport of Hsf protein and free distribution of proteins in the nucleus and cytoplasm. Hsf can activate the heat shock protein (Hsp) thereby promote refolding, assembly, distribution and degradation of damaged proteins. In this way, it helps plants to resist abiotic stress [10].
The first Hsf gene was cloned in yeast. Since then, several mammalian Hsf genes have been identified. In plants, the first Hsf gene was cloned in tomato (Solanum lycopersicum) [6]. To date, Hsfs have been reported in several species, including Arabidopsis [11], rice (Oryza sativa) [12], soybean (Glycine max) [13], maize (Zea mays) [14] due to the continuous improvement of genome sequencing technology. The number of Hsf genes varies across plants. For instance, wheat, soybean, maize and Arabidopsis have 56, 52, 30 and 21 genes, respectively [7].
Hsf gene members are involved in the regulation of growth and development in plants. Carotenoids and chlorophyll have been shown to promote plant photosynthesis in transgenic tobacco by upregulating HsfA9 expression [15]. A total of 14 Hsfs have been identified in Citrus, all of which participate in fruit development and maturation. For instance, CrHsfB2a and CrHsfB5 were found to regulate citrate content [16]. Various stimuli of biotic and abiotic stress can upregulate expression level of Hsfs, hence influence plant response to stress. The expression of HsfA2, HsfB1, HsfA4a, HsfB2a, HsfB2b and HsfA7a in Arabidopsis has been shown to increase with temperature. However, high temperatures do not increase expression of some Hsfs [11]. Previously, 19 Hsf members, including VHsf01, VHsf05, VHsf15 and VHsf18 were found to exhibit different gene expression patterns between resistant and susceptible grape species under high-temperature stress (47 °C) and heat adaptation (38 °C). The differential expression of these genes may explain differences in heat tolerance among various grape species [17]. Different types of biotic and abiotic stresses including cold, drought, pests and diseases increase the expression of Hsf members, such as HsfB3a, HsfA6a, HsfA2a and HsfA9b in Cassava (Manihot esculenta Crantz) [18]. It has also been reported that salinity, osmotic pressure and cold stress increase the expression of HSFA6b, an ABA-mediated stress response gene in Arabidopsis, [19]. Heat, drought and salt stress similarly alters the expression level of Hsf genes in Populus euphratica [20].
In the study, 30 members of Hsf gene family were identified in common bean. These members’ phylogeny, motif, gene structure, evolutionary expansion and expression patterns of the identified genes were analyzed at the sprout stage. Our bionformatic analysis provide insights into PvHsfs and give useful information for further functional dissection of PvHsfs in common bean.
Results
Identification of Hsf members in P. vulgaris
The hmm search identified 35 protein sequences in the reference genome of P. vulgaris. Among them, 30 were found to be members of the Hsf family after removal of duplications. The 30 protein sequences were located on nine chromosomes of P. vulgaris except LG10 and LG11. The Hsf members were named based on their chromosomal location (e.g., PvHsf1-PvHsf30) (Fig. 1). PvHSF06 had the longest protein length (490) and CDS length (1470) whereas PvHSF12 had the shortest protein length (206) and CDS length (618). The isoelectric point of PvHsf members ranged from 4.82 to 9.08, and the molecular weight ranged from 23,907.29 to 54,538.32. The detailed information of PvHsf members is shown in Table S1.

Chromosomal location of PvHsf members. The nine ribbons represent nine chromosomes containing the PvHsf members. The blue lines show the number of genes
Evolutionary analysis of PvHsfs
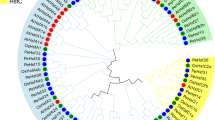
MEGAX software was used to analyze the protein sequence of PvHsf members using the Maximum Likelihoodphy function and jtt + g + i model predicted by MEGA. The 14 subgroups were divided based on the analysis result of PvHsf members (Fig. 2). In these 14 subgroups, the subgroup VI, VII and VIII had only one PvHsf member. The subgroup XIV had the highest number of PvHsf members (4).

Evolutionary analysis of PvHsf members. Different colors show different subgroups
Motifs and gene structure of PvHsfs
Using MEME tool, 10 motifs of PvHsf members were identified (indicated using squares with different colors) and the sequence of each motif is shown in Fig. S1. The location of squares represents the location of each motif (Fig. 3A and B). Motifs- 1, 2, 3 and 5 were identified in all subgroups of PvHsf. Subgroup I, II, III and IV had similar motifs which were found in different locations. Motif-10 was not found in subgroup V while motif-9 was only found in subgroup IX and X. Motif-7 was found in subgroup XIV whereas motif-8 was present in subgroup XI, XII, XIII and XIV. Each subgroup PvHsf members had similar motifs. The structures of exons and introns of PvHsfs were determined using the GSDS (Fig. 3C). Each subgroup members had a conserved Hsf domain (pink box), and the location of exon and intron were similar in each subgroup.

The motifs and gene structure of PvHsf members. (A) Evolutionary relationships among PvHsfs. Different colored shades represent different subgroups (I-XIV). (B) The 10 motifs are represented by different colors. (C) The gene structure of PvHsfs. The pink, yellow and green squares represent Hsf structure, CDS and UTR respectively. The black line represents intron
Evolutionary analysis of four species
In this study, members of PvHsf genes were identified in P. vulgaris whereas members of Hsf genes in plants, including monocotyledonous (O. sativa and Z. mays) and dicotyledonous (Arabidopsis) were retrieved from published research [11, 12, 14]. Also, the jtt + g + i model was the best model to find the evolutionary relationship between Hsf members in four species, which predicted by MEGAX. The four species Hsf members were also devided into 14 subgroups while subgroup III and XI had not monocot members; 20 motifs were identified from the four species Hsf members in motif analysis (Fig. S2), although the order of motifs had changed compared with PvHsfs, the structure of motifs was similar. Motif-1, 2 and 3 encoded DBD domain while motif-9 encoded -LFGV- motif (C-terminal activation domain); Most Hsf members also had two exons, which the results of gene structure was similar with PvHsfs; The results of evolution, gene structure and motif analyses revealed that members of Hsf in each subgroup had similar motifs and gene structure (Fig. 4).

Evolutionary analysis of Hsf members of four species (P. vulgaris, O. sativa, Z. mays and Arabidopsis). Fourteen different colored shadows represent different subgroups. The outer ring indicates the motifs of Hsf members and the inner ring shows the gene structure of Hsf members. Black circles show the PvHsf members
Cis-elements analysis of PvHsfs
Using plantCARE, 10 cis-elements were identified in PvHsfs. These cis-elements found to be involved in the regulation of hormone responsiveness, environmental stress and germination (Table S2). Elements marked in red such as ARBE, TCA-element, TATC-box, P-box, TGA-element and AuxRR-core were hormone-related elements; Three elements marked in blue (LTR, ARE, MBS) were stress-related elements whereas CAT-box was the germination-related element. These results showed that PvHsfs might regulate hormone responsiveness, environmental stress and germination (Fig. 5).

The cis-elements of PvHsfs. A The evolutionary relationships among PvHsfs. The different colored shades represent different subgroups. B The cis-elements of PvHsfs. The red models represent elements associated with hormone responsiveness. The blue models represent elements associated with stress-inducibility. The orange models represent elements involved in germination
Collinearity and Ka/Ks
PvHsfs had 16 pairs of collinearity. PvHsf22 and PvHsf27 exhibited the most pairs (3 pairs) (Fig. 6A). Compared to Arabidopsis (1) and rice (1), PvHsfs in soybeans had more collinear genes (36), indicating a close association between the common bean and soybean (Fig. 6B). KA/KS analysis revealed that there were no two pairs of PvHsfs (Table S3), indicating that PvHsfs eliminated harmful mutations, while maintaining the protein (purity selection).

Collinearity analysis of PvHsfs. A Collinearity among PvHsf members in common bean (P. vulgaris). The PvHsfs marked in red had collinearity while those marked in black lacked collinearity. The middle two rings represent the gene density of each chromosome. Gray background line represents collinear background. Green lines represent collinear relationships among PvHsf members. B Collinearity of PvHsf members in soybean, rice and Arabidopsis. Red, orange, blue and green boxes represent the LGs of common bean, soybean, rice and Arabidopsis respectively. Gray lines indicate the total collinearity background. Orange, blue and green lines show the collinearity of PvHsfs with genes of soybean, rice and Arabidopsis
In Silico tissue-specific expression analysis
Tissue-specific expressions of PvHsfs were obtained from the phytozome database. The expression levels of PvHsfs in flowers, flower buds, young pods, leaves, green mature pods, stems and roots were as shown in Fig. 7. All PvHsfs members were specifically expressed in different tissues, with various PvHsfs members exhibiting marked variations in expression in different tissues. For instance, compared to other tissues, PvHsf02 was highly expressed in green mature pods; Compared with other different tissues, PvHsf05 was highly expressed in roots and pods higher than other tissues; PvHsf03 was highly expressed in flowers, stems and young pods; PvHsf07 levels were suppressed in flowers, flower buds and young pods while PvHsf28 levels were elevated in leaves, stems and roots.

The tissue-specific expression analysis of PvHsfs. A Schematic illustration of different tissues of common bean (P. vulgaris). B A cluster map of each PvHsf. Red indicates high gene expression level of PvHsfs while blue shows low gene expression level of PvHsfs
Tissue-specific expression analysis at the sprout stage
The expression of PvHsfs in cotyledon, hypocotyl and radicle at the sprout stage was also analyzed. The expression levels of PvHsfs were specific in the cotyledon, hypocotyl and radicle. At the sprouting stage, expression levels were relatively low in the hypocotyl, relative to cotyledons and radicles (Fig. 8). Some PvHsfs, including PvHsf01, PvHsf03, PvHsf09, PvHsf17, PvHsf21 and PvHsf22 were highly expressed in the cotyledons while some PvHsfs, including PvHsf05, PvHsf10, PvHsf16 and PvHsf24 were highly expressed in both cotyledons and radicles. These results imply that cotyledons and radicles are potential target tissues for studies on PvHsfs at the sprouting stage.

The expression of PvHsfs in cotyledon, hypocotyl and radicle at the sprout stage. Green, yellow and brown squares indicate the relative mRNA expression of PvHsfs in cotyledon, hypocotyl and radicle of common bean (P. vulgaris)
Stress-associated expression levels
QRT-PCR was used to assess the expressions of PvHsfs under heat and cold stress conditions. Stress dysregulated the expression levels of PvHsfs, with specific characteristic expressions under different stressors. Under heat stress, PvHsfs levels, apart from PvHsf15 levels, were markedly elevated under heat stress except PvHsf15 (Fig. 9). In CK and under cold stress conditions, there were no significant changes in the expressions of dome PvHsfs, such as PvHsf01, PvHsf03, PvHsf05, PvHsf16 and PvHsf29. However, these levels were significantly altered under heat stress, indicating that these PvHsfs are highly responsive to heat stress, when compared to cold stress.

The expression of PvHsfs in radicle of common bean (P. vulgaris) at the sprout stage under cold stress (4 °C) and heat stress (45 °C). The brown, blue and red squares indicate the relative mRNA expression of PvHsfs in radicle under cold stress and heat stress
QRT-PCR was also performed to evaluate the expressions of PvHsfs under salt and heavy metal stressors (Fig. 10). Under salt stress conditions, PvHsf21 and PvHsf22 levels were significantly elevated, however, PvHsf01, PvHsf03, PvHsf09, PvHsf17 and PvHsf24 levels were markedly suppressed. Moreover, exposure to Cd2+ stress suppressed the levels of most PvHsfs, apart from PvHsf09, PvHsf21 and PvHsf22. Under salt and heavy metal stressors, levels of most PvHsfs were significantly dysregulated, apart from those of PvHsf03. PvHsf01, PvHsf17, PvHsf21, PvHsf22 and PvHsf24. Therefore, PvHsfs can be used as candidate members of the Hsf family for studies at sprouting stages under stress.

The expression of PvHsfs in radicle of common bean (P. vulgaris) at the sprout stage under salt stress (NaCl) and heavy metal stress (CdCl2 and HgCl2). Brown, white, purple and gray squares indicate the relative mRNA expression of PvHsfs in radicle under salt, CdCl2 and HgCl2 stress
Discussion
Various species have different Hsf members. There are 29 Hsf members in Tartary buckwheat (Fagopyrum tataricum) [21], 28 members in poplar (Populus) [22], 21 members in Arabidopsis [23], 19 members in grapes (Vitis vinifera) [24], 18 members in tomatoes (Solanum lycopersicum) [7] and 16 members in alfalfa (Medicago sativa) [22]. Duplication of gene family members during plant evolution is associated with genomic replication events [17]. In this study, 30 Hsf family members were identified in the genome of the common bean (PvHsf1-PvHsf30) (Fig. 1). Compared to other dicotyledons, the common bean was established to have more Hsf members. These PvHsfs were all found to be located in 11 linkage groups. Therefore, based on these results, after differentiation from early ancestors, the common bean may have had more genomic replication events.
PvHsfs in each subgroup exhibited similar motifs. Motifs-1, 2 and 3, encoding the DBD domain, were present in all PvHsfs subgroups. This domain was the most conserved and had a three helical-bundle structure [25]. Subgroup X and XI exhibited motif-9 (AHA domain) [23] while motif-8, a characteristic C-terminal motif (−LFGV-), which functions as a repressor domain, was found in PvHsf members of subgroup XI, XII, XIII and XIV [9]. Comparable findings, whereby Hsf members in each subgroup had similar motifs, were found in Populus trichocarpa [26], cotton (Gossypium hirsutum) [27] and peach (Prunus persica) [28].
Introns regulate gene expressions [29]. Therefore, it is important to elucidate on gene functions by analyzing gene structures [30]. Analyses of gene structures of PvHsf members revealed that all PvHsfs had more than one intron. However, intron lengths were different for each subgroup, and each subgroup member exhibited a similar structure. The shortest introns were in subgroup XIII, while the longest introns were in subgroup V. Comparable findings have been reported in Soybean [31], Hypericum perforatum [32] and cotton [33].
Rice and maize are monocotyledons, while common bean (P. vulgaris) and Arabidopsis are dicotyledons. Phylogenetic tree analysis revealed that Hsf members in the above four species were in the same subgroups (14) while subgroup III and XI had no monocot members, which could be attributed to evolution of plants into monocots and dicots. Hsfs of the same subgroup in these four species exhibited similar gene structures. However, the order of motifs in the four species exhibited some differences. Same subgroup Hsfs had similar motifs, such as motifs-1, 2, 3 (DBD domain) as well as motif-8 (C-terminal domain) and they also exhibited similar motif compositions, implying that Hsf members play similar functions in proteins. Comparable outcomes have been found among Hsf members in maize [34], moso bamboo (Phyllostachys edulis) [35], pumpkin (Cucurbita moschata) [36] and tea plant (Camellia sinensis) [37]. Moreover, Hsf members appear before plants differentiate into monocotyledons or dicotyledons, and same subgroup members exhibit similar motifs and evolutionary relationships [21].
The cis-elements in promoter regions of gene family members regulate the expressions of metabolic pathway-related genes [38]. In this study, seven cis-elements related to hormones (such as ARBE, TATC-box, P-box, AuxRR-core, TCA and TGA elements) were identified, indicating that PvHsfs may be involved in the roles of hormones in plant growth and development. Three stress-related cis-elements (LTR, ARE and MBS elements) were found in different PvHsfs, including PvHsf09, PvHsf10, PvHsf15, PvHsf17 and PvHsf24. Under abiotic stress conditions, the expression levels of these PvHsfs exhibited significant changes (Figs. 9 and 10), confirming that stress-related cis-acting elements are responsive to abiotic stress. Also, Hormone-related cis-elements, including ABRE, AuxRR-core, P-box and TGA-elements, have been found in carnation (Dianthus caryophyllus) [39] and Hypericum perforatum [32]. Hsf members while three stress-related cis-elements (LTR, ARE and MBS elements) have also been found in Hsf members from Brassica juncea [40] and Hypericum perforatum [32]. Moreover, Hsf members in carnation (Dianthus caryophyllus) [39], Brassica juncea [40], Hypericum perforatum [32] and tea plants (Camellia sinensis) [37] had a CAT-box element. Under abiotic stress, the expressions of Hsf members exhibited some variations.
Collinearity analysis revealed that PvHsfs had more pairs of homologous genes in soybean (37) than in Arabidopsis (1) and rice (1), indicating that PvHsfs are closely associated with legumes. The collinear gene of PvHsfs in Arabidopsis, AT2G41690, has been reported to exert some effects during abiotic stress [41], indicating that PvHsfs influence abiotic stress responses in plants.
Gene expression patterns can be used to investigate the biological functions of various genes [42]. In crops, Hsf members exhibit tissue-specific expressions. For instance, DcaHsfs exhibit different expression patterns in carnation, as well as in members of the same class [39]. StHsf genes are highly expressed in various potato (Solanum tuberosum) tissues [43]. The expressions of PvHsfs in the phytozome database exhibit tissue-specificity (flowers, flower buds, young pods, leaves, green mature pods, stems and roots), indicating that PvHsfs are involved in plant growth and development.
During plant growth, the sprouting stage is the first and most important stage. It directly affects plant development and yield. Moreover, during abiotic stress, it is the most sensitive stage [44, 45]. As a result, studies have evaluated the effects of stress on the sprouting stage [46]. Herein, PvHsfs levels at the sprout stage were specific in the cotyledon and radicle, indicating that these tissues can be used as target tissues to assess PvHsfs at the sprout stage.
Stress, especially abiotic stress, alters the expressions of Hsf members [7]. For instance, heat stress alters the expressions of CarHsfs in chickpea (Cicer arietinum) [47]. Moreover, various stresses dysregulate the expression levels of CsHsf genes in tea plants [37]. In pumpkins (Cucurbita moschata), cold and heat stress have been shown to significantly alter the expressions of some CmHsfs [36]. Abiotic stressors, such as heat, cold, salt, drought and cadmium have been shown to alter the expressions of most TaHsfs in bread wheat (Triticum aestivum) [48]. Collectively, these findings imply that Hsf members can respond and resist abiotic stress. qRT-PCR assays were performed to assess the expression profiles of PvHsfs under heat, cold, salt and heavy metal stresses. PvHsf01, PvHsf17, PvHsf21 and PvHsf24 had a significant different expression under heat, cold, salt and heavy metal stress compared with CK treatment, which was similar expression-patterns in DcaHsfs (D. caryophyllus) [39]. These abiotic stressors altered the expressions of PvHsfs, similar to the patterns observed in other plants.
Conclusions
In summary, this study identified 30 members of PvHsf from the reference genome and comprehensively analyzed the location, motifs, gene structure, cis-elements and collinearity among PvHsf members. The PvHsfs’ expression from the phytozome database and the analysis at the sprout stage in different tissues all revealed that PvHsfs had the tissue-specific expression. In addition, the expression of PvHsfs under heat, cold, salt and heavy metal stress showed PvHsfs might regulate responses to abiotic stresses in common bean. This study lays the foundation for further identification of PvHsfs and adds to our understanding on the role of PvHsfs in the regulation of abiotic stress resistance in common bean.
Materials and methods
Identification of Hsf members in P. vulgaris
Genomic data (genes, cDNAs and proteins) of P. vulgaris (PhaVulg1_0) was derived from the ensembl plants database [49] while data of Hsf protein domain (PF00447) was obtained from the PFAM database [50]. These data were screened using the HMMER software [51] to identify Hsf members. In addition, ExPASy Proteomics Server [52] and Plant Protein Phosphorylation DataBase (P3DB) [53] were screened to identify the Hsf members in common bean (PvHsf). The location of PvHsf members was mapped based on the reference genome and named depending on their chromosomal location using TBtools [54].
Analysis of Hsf members in P. vulgaris
MEGA X [55] was used to align protein sequences of PvHsf and Hsf members in three species (Arabidopsis, maize, rice) reported previously [11, 12, 14]. Maximum Likelihoodphy analysis was performed using 1000 replicates as bootstrap values and the jtt + g + i model predicted by MEGA. The MEME tool [56] was used to identify motifs with E-value of less than 1e− 20 and 10–50 amino acids numbered based on their corresponding E-values. The gene structure of PvHsfs was analyzed using Gene Structure Display Server (GSDS) [57]. Gene-wise [58] was used to determine the coordinates corresponding to DNA (containing exon and intron) and protein sequences. The cis-acting elements of PvHsfs were uncovered using the plantCARE software [38]. Circos software [59] was used to analyze gene duplication events in PvHsfs via the MCScanX function [60]. The expression of PvHsfs was visualized using a heatmap constructed using TBtools (phytozome data) [61]. All databases and software links are shown in Table S4.
Preparation of plant materials and qRT-PCR analysis
A locally-grown common bean variety “Longjiang Ziyun” was obtained from the National Coarse Cereals Engineering Research Center (Daqing, Heilongjiang, China). The seeds were placed in an incubator away from light at 26 °C to allow sprouting. The plants were separately exposed to different stress treatments on the fifth day. Cold stress was induced by exposing plants to a temperature of 4 °C and heat stress was induced by exposing plants to a temperature of 45 °C [62, 63]. Salt stress was triggered by treatment with 70 mmol/L (NaCl), while heavy metal stress was simulated by exposing plants to 0.5 mg/L (CdCl2) and 60 mg/L (HgCl2) [64,65,66]. For control (CK) treatment, hypocotyl, radicle and cotyledon were collected as for samples in the analysis of tissue-specific expression. The radicles under abiotic stress treatments were collected as samples respectively while the CK was served as the control tissue sample. RNA Easy Fast Kit (DP452, Tiangen, Beijing) was used to extract RNA and cDNA was obtained by total RNA reverse transcription using HiScript SuperMix was used to extract for qPCR (+gDNA wiper) (R223–01, Vazyme, Nanjing). The Primer premier 5.0 software (PREMIER Biosoft, San Francisco, USA) was used to design primers of PvHsf members (Table S5). In this experiment, Pvactin11 gene served as the internal reference gene [67]. The expression of each PvHsf member was determined through qRT-PCR using the Light Cycler system (Roche 480II, Roche, Switzerland) and TransStart® Top Green qPCR SuperMix (AQ131–04, TransGen Biotech, Beijing). For each treatment, three biological replicates were prepared, and for each sample, three technical replicates were prepared. The relative mRNA expression was calculated as previously described [68].
Availability of data and materials
All data generated or analysed during this study are included in this published article and its supplementary information files.
Abbreviations
- Hsf:
-
Heat shock transcription factor.
- qRT-PCR:
-
Quantitative Real-time PCR
- Ka/Ks:
-
The ratio of the number of nonsynonymous substitutions per nonsynonymous site (Ka) to the number of synonymous substitutions per synonymous site (Ks)
References
Nichols NN, Sutivisedsak N, Dien BS, Biswas A, Lesch WC, Cotta MA. Conversion of starch from dry common beans (Phaseolus vulgaris L.) to ethanol. Ind Crop Prod. 2011. https://doi.org/10.1016/j.indcrop.2010.12.029.
Lee SC, Gepts PL, Whitaker JR. Protein structures of common bean (Phaseolus vulgaris) alpha-amylase inhibitors. J Agric Food Chem. 2002. https://doi.org/10.1021/jf020189t.
Singh RS, Walia AK. Microbial lectins and their prospective mitogenic potential. Crit Rev Microbiol. 2014. https://doi.org/10.3109/1040841x.2012.733680.
Delgado-Salinas A, Turley T, Richman A, Lavin M. Phylogenetic analysis of the cultivated and wild species of Phaseolus (Fabaceae). Syst Bot. 1999. https://doi.org/10.2307/2419699.
Ohama N, Sato H, Shinozaki K, Yamaguchi-Shinozaki K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017. https://doi.org/10.1016/j.tplants.2016.08.015.
Scharf KD, Rose S, Zott W, Schöffl F, Nover L. Three tomato genes code for heat stress transcription factors with a region of remarkable homology to the DNA-binding domain of the yeast HSF. EMBO J. 1990. https://doi.org/10.1002/j.1460-2075.1990.tb07900.x.
Scharf KD, Berberich T, Ebersberger I, Nover L. The plant heat stress transcription factor (Hsf) family: structure, function and evolution. Biochim Biophys Acta. 2012. https://doi.org/10.1016/j.bbagrm.2011.10.002.
Wu C. Heat shock transcription factors: structure and regulation. Annu Rev Cell Dev Biol. 1995. https://doi.org/10.1146/annurev.cb.11.110195.002301.
Fragkostefanakis S, Röth S, Schleiff E, Scharf KD. Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks. Plant Cell Environ. 2015. https://doi.org/10.1111/pce.12396.
Bharti K, Von Koskull-Döring P, Bharti S, Kumar P, Tintschl-Körbitzer A, Treuter E, et al. Tomato heat stress transcription factor HsfB1 represents a novel type of general transcription coactivator with a histone-like motif interacting with the plant CREB binding protein ortholog HAC1. Plant Cell. 2004. https://doi.org/10.1105/tpc.019927.
Busch W, Wunderlich M, Schöffl F. Identification of novel heat shock factor-dependent genes and biochemical pathways in Arabidopsis thaliana. Plant J. 2005. https://doi.org/10.1111/j.1365-313X.2004.02272.x.
Guo J, Wu J, Ji Q, Wang C, Luo L, Yuan Y, et al. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J Genet Genomics. 2008. https://doi.org/10.1016/s1673-8527(08)60016-8.
Chung E, Kim KM, Lee JH. Genome-wide analysis and molecular characterization of heat shock transcription factor family in Glycine max. J Genet Genomics. 2013. https://doi.org/10.1016/j.jgg.2012.12.002.
Lin YX, Jiang HY, Chu ZX, Tang XL, Zhu SW, Cheng BJ. Genome-wide identification, classification and analysis of heat shock transcription factor family in maize. BMC Genomics. 2011. https://doi.org/10.1186/1471-2164-12-76.
Prieto-Dapena P, Almoguera C, Personat JM, Merchan F, Jordano J. Seed-specific transcription factor HSFA9 links late embryogenesis and early photomorphogenesis. J Exp Bot. 2017. https://doi.org/10.1093/jxb/erx020.
Lin Q, Jiang Q, Lin J, Wang D, Li S, Liu C, et al. Heat shock transcription factors expression during fruit development and under hot air stress in Ponkan (Citrus reticulata Blanco cv. Ponkan) fruit. Gene. 2015. https://doi.org/10.1016/j.gene.2015.01.024.
Liu M, Ma Z, Zheng T, Wang J, Huang L, Sun W, et al. The potential role of Auxin and Abscisic acid balance and FtARF2 in the final size determination of Tartary buckwheat fruit. Int J Mol Sci. 2018. https://doi.org/10.3390/ijms19092755.
Huang Y, Li MY, Wang F, Xu ZS, Huang W, Wang GL, et al. Heat shock factors in carrot: genome-wide identification, classification, and expression profiles response to abiotic stress. Mol Biol Rep. 2015. https://doi.org/10.1007/s11033-014-3826-x.
Huang YC, Niu CY, Yang CR, Jinn TL. The heat stress factor HSFA6b connects ABA signaling and ABA-mediated heat responses. Plant Physiol. 2016. https://doi.org/10.1104/pp.16.00860.
Zhang J, Jia H, Li J, Li Y, Lu M, Hu J. Molecular evolution and expression divergence of the Populus euphratica Hsf genes provide insight into the stress acclimation of desert poplar. Sci Rep. 2016. https://doi.org/10.1038/srep30050.
Liu M, Huang Q, Sun W, Ma Z, Huang L, Wu Q, et al. Genome-wide investigation of the heat shock transcription factor (Hsf) gene family in Tartary buckwheat (Fagopyrum tataricum). BMC Genomics. 2019. https://doi.org/10.1186/s12864-019-6205-0.
Wang F, Dong Q, Jiang H, Zhu S, Chen B, Xiang Y. Genome-wide analysis of the heat shock transcription factors in Populus trichocarpa and Medicago truncatula. Mol Biol Rep. 2012. https://doi.org/10.1007/s11033-011-0933-9.
Nover L, Bharti K, Döring P, Mishra SK, Ganguli A, Scharf KD. Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need? Cell Stress Chaperones. 2001. https://doi.org/10.1379/1466-1268(2001)006<0177:aathst>2.0.co;2.
Liu G, Chai F, Wang Y, Jiang J, Duan W, Wang Y, et al. Genome-wide identification and classification of HSF family in grape, and their transcriptional analysis under heat acclimation and heat stress. Horticult Plant J. 2018. https://doi.org/10.1016/j.hpj.2018.06.001.
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ, Coutu J, et al. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell. 2005. https://doi.org/10.1105/tpc.104.026971.
Zhang J, Liu B, Li J, Zhang L, Wang Y, Zheng H, et al. Hsf and Hsp gene families in Populus: genome-wide identification, organization and correlated expression during development and in stress responses. BMC Genomics. 2015. https://doi.org/10.1186/s12864-015-1398-3.
Wang J, Sun N, Deng T, Zhang L, Zuo K. Genome-wide cloning, identification, classification and functional analysis of cotton heat shock transcription factors in cotton (Gossypium hirsutum). BMC Genomics. 2014. https://doi.org/10.1186/1471-2164-15-961.
Tan B, Yan L, Li H, Lian X, Cheng J, Wang W, et al. Genome-wide identification of HSF family in peach and functional analysis of PpHSF5 involvement in root and aerial organ development. PeerJ. 2021. https://doi.org/10.7717/peerj.10961.
Shaul O. How introns enhance gene expression. Int J Biochem Cell Biol. 2017. https://doi.org/10.1016/j.biocel.2017.06.016.
Li Y, Chen D, Luo S, Zhu Y, Jia X, Duan Y, et al. Intron-mediated regulation of β-tubulin genes expression affects the sensitivity to carbendazim in Fusarium graminearum. Curr Genet. 2019. https://doi.org/10.1007/s00294-019-00960-4.
Li PS, Yu TF, He GH, Chen M, Zhou YB, Chai SC, et al. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genomics. 2014. https://doi.org/10.1186/1471-2164-15-1009.
Zhou L, Yu X, Wang D, Li L, Zhou W, Zhang Q, et al. Genome-wide identification, classification and expression profile analysis of the HSF gene family in Hypericum perforatum. PeerJ. 2021. https://doi.org/10.7717/peerj.11345.
Rehman A, Atif RM, Azhar MT, Peng Z, Li H, Qin G, et al. X., genome wide identification, classification and functional characterization of heat shock transcription factors in cultivated and ancestral cottons (Gossypium spp.). Int J Biol Macromol. 2021. https://doi.org/10.1016/j.ijbiomac.2021.05.016.
Zhang H, Li G, Fu C, Duan S, Hu D, Guo X. Genome-wide identification, transcriptome analysis and alternative splicing events of Hsf family genes in maize. Sci Rep. 2020. https://doi.org/10.1038/s41598-020-65068-z.
Huang B, Huang Z, Ma R, Chen J, Zhang Z, Yrjälä K. Genome-wide identification and analysis of the heat shock transcription factor family in moso bamboo (Phyllostachys edulis). Sci Rep. 2021. https://doi.org/10.1038/s41598-021-95899-3.
Shen C, Yuan J. Genome-wide characterization and expression analysis of the heat shock transcription factor family in pumpkin (Cucurbita moschata). BMC Plant Biol. 2020. https://doi.org/10.1186/s12870-020-02683-y.
Zhang X, Xu W, Ni D, Wang M, Guo G. Genome-wide characterization of tea plant (Camellia sinensis) Hsf transcription factor family and role of CsHsfA2 in heat tolerance. BMC Plant Biol. 2020. https://doi.org/10.1186/s12870-020-02462-9.
Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, et al. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002. https://doi.org/10.1093/nar/30.1.325.
Li W, Wan XL, Yu JY, Wang KL, Zhang J. Genome-wide identification, classification, and expression analysis of the Hsf gene family in carnation (Dianthus caryophyllus). Int J Mol Sci. 2019. https://doi.org/10.3390/ijms20205233.
Li M, Xie F, Li Y, Gong L, Luo Y, Zhang Y, et al. Genome-wide analysis of the heat shock transcription factor gene family in Brassica juncea: structure, evolution, and expression profiles. DNA Cell Biol. 2020. https://doi.org/10.1089/dna.2020.5922.
Sarry JE, Kuhn L, Ducruix C, Lafaye A, Junot C, Hugouvieux V, et al. The early responses of Arabidopsis thaliana cells to cadmium exposure explored by protein and metabolite profiling analyses. Proteomics. 2006. https://doi.org/10.1002/pmic.200500543.
Maheswari U, Jabbari K, Petit JL, Porcel BM, Allen AE, Cadoret JP, et al. Digital expression profiling of novel diatom transcripts provides insight into their biological functions. Genome Biol. 2010. https://doi.org/10.1186/gb-2010-11-8-r85.
Tang R, Zhu W, Song X, Lin X, Cai J, Wang M, et al. Genome-wide identification and function analyses of heat shock transcription factors in potato. Front Plant Sci. 2016. https://doi.org/10.3389/fpls.2016.00490.
Fahad S, Bajwa AA, Nazir U, Anjum SA, Farooq A, Zohaib A, et al. Crop production under drought and heat stress: plant responses and management options. Front Plant Sci. 2017. https://doi.org/10.3389/fpls.2017.01147.
Ibrahim EA. Seed priming to alleviate salinity stress in germinating seeds. J Plant Physiol. 2016. https://doi.org/10.1016/j.jplph.2015.12.011.
Yadav PV, Kumari M, Ahmed Z. Seed priming mediated germination improvement and tolerance to subsequent exposure to cold and salt stress in Capsicum. Res J Seed Sci. 2011. https://doi.org/10.3923/rjss.2011.125.136.
Chidambaranathan P, Jagannadham PTK, Satheesh V, Kohli D, Basavarajappa SH, Chellapilla B, et al. Genome-wide analysis identifies chickpea (Cicer arietinum) heat stress transcription factors (Hsfs) responsive to heat stress at the pod development stage. J Plant Res. 2018. https://doi.org/10.1007/s10265-017-0948-y.
Zhou M, Zheng S, Liu R, Lu J, Lu L, Zhang C, et al. Genome-wide identification, phylogenetic and expression analysis of the heat shock transcription factor family in bread wheat (Triticum aestivum L.). BMC Genomics. 2019. https://doi.org/10.1186/s12864-019-5876-x.
Zhang Q, Zhang WJ, Yin ZG, Li WJ, Zhao HH, Zhang S, et al. Genome- and Transcriptome-wide identification of C3Hs in common bean (Phaseolus vulgaris L.) and structural and expression-based analyses of their functions during the sprout stage under salt-stress conditions. Front Genet. 2020. https://doi.org/10.3389/fgene.2020.564607.
Finn RD, Bateman A, Clements J, Coggill P, Eberhardt RY, Eddy SR, et al. Pfam: the protein families database. Nucleic Acids Res. 2014. https://doi.org/10.1093/nar/gkt1223.
Finn RD, Clements J, Arndt W, Miller BL, Wheeler TJ, Schreiber F, et al. HMMER web server: 2015 update. Nucleic Acids Res. 2015. https://doi.org/10.1093/nar/gkv397.
Hoogland C, Mostaguir K, Appel RD, Lisacek F. The world-2DPAGE constellation to promote and publish gel-based proteomics data through the ExPASy server. J Proteome. 2008. https://doi.org/10.1016/j.jprot.2008.02.005.
Yao Q, Xu D. Bioinformatics analysis of protein phosphorylation in plant systems biology using P3DB. Methods Mol Biol. 2017. https://doi.org/10.1007/978-1-4939-6783-4_6.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020. https://doi.org/10.1016/j.molp.2020.06.009.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018. https://doi.org/10.1093/molbev/msy096.
Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, et al. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009. https://doi.org/10.1093/nar/gkp335.
Hu B, Jin J, Guo AY, Zhang H, Luo J, Gao G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics. 2015. https://doi.org/10.1093/bioinformatics/btu817.
Simmons MP, Sloan DB, Springer MS, Gatesy J. Gene-wise resampling outperforms site-wise resampling in phylogenetic coalescence analyses. Mol Phylogenet Evol. 2019. https://doi.org/10.1016/j.ympev.2018.10.001.
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, et al. Circos: an information aesthetic for comparative genomics. Genome Res. 2009. https://doi.org/10.1101/gr.092759.109.
Wang Y, Li J, Paterson AH. MCScanX-transposed: detecting transposed gene duplications based on multiple colinearity scans. Bioinformatics. 2013. https://doi.org/10.1093/bioinformatics/btt150.
Goodstein DM, Shu S, Howson R, Neupane R, Hayes RD, Fazo J, et al. Phytozome: a comparative platform for green plant genomics. Nucleic Acids Res. 2012. https://doi.org/10.1093/nar/gkr944.
Alsamir M, Mahmood T, Trethowan R, Ahmad N. An overview of heat stress in tomato (Solanum lycopersicum L.). Saudi. J Biol Sci. 2021. https://doi.org/10.1016/j.sjbs.2020.11.088.
Wang F, Chen X, Dong S, Jiang X, Wang L, Yu J, et al. Crosstalk of PIF4 and DELLA modulates CBF transcript and hormone homeostasis in cold response in tomato. Plant Biotechnol J. 2020. https://doi.org/10.1111/pbi.13272.
Mohammadi S, Pourakbar L, Siavash Moghaddam S, Popović-Djordjević J. The effect of EDTA and citric acid on biochemical processes and changes in phenolic compounds profile of okra (Abelmoschus esculentus L.) under mercury stress. Ecotoxicol Environ Saf. 2021. https://doi.org/10.1016/j.ecoenv.2020.111607.
Zhang Q, Zhang W-j, Yin Z-g, Li W-j, Xia C-Y, Sun H-Y, et al. Genome-wide identification reveals the potential functions of the bZIP gene family in common bean (Phaseolus vulgaris) in response to salt stress during the sprouting stage. J Plant Growth Regul. 2021. https://doi.org/10.1007/s00344-021-10497-x.
Zhao Q, Wang H, Du Y, Rogers HJ, Wu Z, Jia S, et al. MSH2 and MSH6 in mismatch repair system account for soybean (Glycine max (L.) Merr.) tolerance to cadmium toxicity by determining DNA damage response. J Agric Food Chem. 2020. https://doi.org/10.1021/acs.jafc.9b06599.
Zhang Q, Li M, Xia CY, Zhang WJ, Yin ZG, Zhang YL, et al. Transcriptome-based analysis of salt-related genes during the sprout stage of common bean (Phaseolus vulgaris) under salt stress conditions. Biotechnol Biotechnol Equip. 2021. https://doi.org/10.1080/13102818.2021.1954091.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−ΔΔC(T)) method. Methods. 2001. https://doi.org/10.1006/meth.2001.1262.
Acknowledgements
We thank all of the members of Agricultural College, Heilongjiang Bayi Agricultural University and National Coarse Cereals Technology Engineering Research Center for their support throughout the study.
Funding
This study was financially supported by National Key Research and Development Program of China (2020YFD1001402); and the Research Project of Heilongjiang Bayi Agricultural University (XDB2011–02). The funders had no role in the experimental design, data collection, analysis and interpretation of the data or writing the manuscript.
Author information
Authors and Affiliations
Contributions
Qi Zhang: Methodology, Data curation and Writing original draft; Jing Geng: Data curation; Yanli Du and Qiang Zhao: Conceptualization and Methodology; Wenjing Zhang and Qingxi Fang: Software; Jianghui Li: Formal analysis, Xiankai Yuan: Preparation of materials; Yaru Fan: Methodology; Jidao Du: Conceptualization, Data curation and Funding acquisition. All authors reviewed the manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
None.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
The Motif structure (Motif1-Motif10) of PvHsf members.
Additional file 2: Figure S2.
The Motif structure (Motif1-Motif20) of Hsf members in Arabidopsis, rice (Oryza sativa), maize (Zea mays) and common bean (Phaseolus vulgaris).
Additional file 3: Table S1.
Identification of PvHsf members in P. vulgaris.
Additional file 4: Table S2.
The cis-acting elements of PvHsfs.
Additional file 5: Table S3.
KA/KS of PvHsfs.
Additional file 6: Table S4.
The database and software websites.
Additional file 7: Table S5.
QRT-PCR primer of PvHsfs designed by Primer premier 5.0 software.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

{kind=link}
{kind=link}
Cite this article
Zhang, Q., Geng, J., Du, Y. et al. Heat shock transcription factor (Hsf) gene family in common bean (Phaseolus vulgaris): genome-wide identification, phylogeny, evolutionary expansion and expression analyses at the sprout stage under abiotic stress. BMC Plant Biol 22, 33 (2022). https://doi.org/10.1186/s12870-021-03417-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-021-03417-4