
Abstract
Backgrounds
Populus and Salix belong to Salicaceae and are used as models to investigate woody plant physiology. The variation of karyotype and nuclear DNA content can partly reflect the evolutionary history of the whole genome, and can provide critical information for understanding, predicting, and potentially ameliorating the woody plant traits. Therefore, it is essential to study the chromosome number (CN) and genome size in detail to provide information for revealing the evolutionary process of Salicaceae.
Results
In this study, we report the somatic CNs of seventeen species from eight genera in Salicaceae. Of these, CNs for twelve species and for five genera are reported for the first time. Among the three subfamilies of Salicaceae, the available data indicate CN in Samydoideae is n = 21, 22, 42. The only two genera, Dianyuea and Scyphostegia, in Scyphostegioideae respectively have n = 9 and 18. In Salicoideae, Populus, Salix and five genera closely related to them (Bennettiodendron, Idesia, Carrierea, Poliothyrsis, Itoa) are based on relatively high CNs from n = 19, 20, 21, 22 to n = 95 in Salix. However, the other genera of Salicoideae are mainly based on relatively low CNs of n = 9, 10, 11. The genome sizes of 35 taxa belonging to 14 genera of Salicaceae were estimated. Of these, the genome sizes of 12 genera and all taxa except Populus euphratica are first reported. Except for Dianyuea, Idesia and Bennettiodendron, all examined species have relatively small genome sizes of less than 1 pg, although polyploidization exists.
Conclusions
The variation of CN and genome size across Salicaceae indicates frequent ploidy changes and a widespread sharing of the salicoid whole genome duplication (WGD) by the relatives of Populus and Salix. The shrinkage of genome size after WGD indicates massive loss of genomic components. The phylogenetic asymmetry in clade of Populus, Salix, and their close relatives suggests that there is a lag-time for the subsequent radiations after the salicoid WGD event. Our results provide useful data for studying the evolutionary events of Salicaceae.
Similar content being viewed by others

Background
Populus L. (poplars) and Salix L. (willows) include important woody trees with enormous ecological and economic value. The two genera have many valuable characteristics, such as fast growth, easy propagation, hybridization, pleasing appearance, and widespread distribution. These features make them important fiber resources, fuelwood, and candidates for ecological management projects, for example the rehabilitation of degraded land and the mitigation of climate change [1]. They also have important research value in the fields of wood formation, long-term perennial growth and seasonality. They are model systems of woody plant genetics, genomics, and biology [2].
The evolutionary history of poplars and willows is the basic biological roadmap to guide the disclosure of woody plant traits. A reliable phylogenetic relationship among poplar, willow and their relatives is essential, but it has just surfaced. The now expended Salicaceae includes three subfamilies, Samydoideae, Scyphostegioideae and Salicoideae [3]. Phylogenetic analysis based on molecular or morphological data resolved Populus and Salix as sister genera, and they are deeply nested in Salicoideae [3,4,5,6]. And the immediate sister groups to the clade containing poplars and willows are resolved as some genera that are apetalous, unisexual, and mainly dioecious, namely Poliothyrsis Oliv., Itoa Hemsl., Carrierea Franch., Idesia Maxim., Bennettiodendron Merr., Olmediella Baill., and Macrohasseltia L.O. Williams [4, 7, 8]. However, under different analysis methods and taxon sampling densities, the sister taxon with Populus and Salix remains controversial [3, 6,7,8,9].
Polyploidy or whole-genome duplication (WGD) is an important source for adaptation, speciation and evolution in plants [10]. Studies based on chromosome numbers suggested that ca. 30 % to perhaps 70 % of angiosperm are of polyploid origin [11,12,13]. Recent genome- and transcriptome-based analyses revealed that angiosperm contains at least one paleopolyploid event and lineage-specific polyploidy events are widespread [14,15,16]. Changes in gene expression and epigenetics after polyploidization can affect the morphology and physiology of polyploidies which in turn has the potential to affect the bio-environment and interspecies interactions [17,18,19,20]. Several ancient genome-doubling events have been proved to be closely related to evolution radiation and diversification in many angiosperm lineages such as Poaceae, Solanaceae, Fabaceae, and Brassicaceae [14, 21]. In Malpighiales, which Salicaceae belongs to, Cai et al. [22] identified 22 ancient WGD events which clustered around the Eocene–Paleocene transition, during which time the planet was warmer and wetter than any period in the Cenozoic. And these WGDs are usually associated with the most diverse clades in Malpighiales, for example, the clusioids, ochnoids, euphorbioids, phyllanthoids, violets, and passion flowers. The salicoid WGD event is inferred to predate the common ancestor of Populus and Salix. However, it remains unclear whether this WGD event is shared by other taxa of Salicaceae [22].
Diversification and speciation of plants are often accompanied by variations in the chromosome number and structure, together with the amount of nuclear DNA. Nuclear genome size, i.e. the DNA content of the unreplicated nucleus, 2 C [23], is an important genomic parameter that exhibits pronounced variation among angiosperm with a minimum of 1 C = 0.07 pg in Genlisea aurea [24] and a known maximum of 1 C = 152.23 pg in Paris japonica [25]. There is an increased interest in its evolutionary potential in the last decade [26,27,28,29,30,31]. For example, a study using 219 geophytes indicated a positive correlation between stomatal and genome size, and increased genome size was associated with earliness of flowering and tendency to grow in humid conditions [32]. In Veronica, life history is significantly correlated with 1 C-value, and significant genome downsizing accompanied by increased diversification rates exist in the polyploid Southern Hemisphere subgenus Pseudoveronica and two Northern Hemisphere subgenera [33]. Thus, assessments of the karyotype and nuclear DNA content are traditional and useful methods to explore genetic relationships and polyploid events [34, 35]. In Populus, the chromosome number of 24 species from five sections are known [36, 37]. They all have the basic chromosome number (BCN) x = 19, and the majority individuals are diploid, except in the north American aspen P. tremuloides Michx., triploids are widespread in unglaciated, drought-prone regions [38]. In Salix, the situation is much more complicated. Although most species are based on x = 19, BCN of x = 22 also appears in some species. In some extreme examples, different BCNs may present in the same species [39,40,41]. However, the chromosome number (CN) data of other Salicaceae genera are very insufficient (Supplementary Table S1). The pantropical Samydoideae includes 13 genera and ca. 235 species. Only one genus (7.7 %) and five species (2.1 %) have CN reports. In Salicoideae, which includes 40 genera and more than 960 species, there is few attention on cytology of the taxa except Populus and Salix. Only 9 genera (22.5 %), and 28 taxa (excluding the 201 taxa of Populus or Salix) have CN reports. The close relatives of Populus and Salix include Itoa, Poliothyrsis, Carrierea, Idesia, Bennettiodendron, Macrohasseltia and Olmediella [5]. Of these seven genera, there is only an uncertain CN report for Idesia polycarpa Maxim. [42]. Genome size, defined as the DNA mass in picograms within an un-replicated gametic nucleus, is a basic and important metric for comparing plant genomes and can provide insight into the evolutionary history of plants [23]. Kew Plant DNA C-value database [43] is a widely used resource that contains many past and current estimates of genome size. The database currently contains C-value data for 12,273 species comprising 10,770 angiosperms, 421 gymnosperms, 303 pteridophytes, 334 bryophytes, and 445 algae [43]. In Salicaceae, only three genera (Populus, Salix, and Casearia Jacq.) and 24 species have a genome size report ([43]; Supplementary Table S2). The lack of recognition of chromosome number and DNA content in Salicaceae hinders our understanding of the role of polyploidization in the evolution of Salicaceae.
In this article, we intend to study the chromosome number and genome size of Salicaceae especially the close relatives of Populus and Salix. We want to give a more precise process by which the chromosome number changes under the phylogenetic framework; and we intend to uncover the phylogenetic placements of the salicoid WGD.
Results
Somatic Karyotypes in Salicaceae
Chromosome number (CN) has been the most influential and ease-obtain data for detecting major genomic events, such as whole genome duplication (WGD). To reveal the evolutionary history of Populus genomes, we explored the dynamic changes of CN in Salicaceae. The phylogeny of Salicaceae and the likely sister family Lacistemataceae is presented following previous studies [3, 44,45,46,47]. We collected available karyotypes of Salicaceae and Lacistemataceae species from online database, Chromosome Counts Database (Supplementary Table S1). In addition, we detected the chromosome numbers of seventeen species from eight genera by cytological analysis (Table 1). These eight genera include Populus, Itoa, Poliothyrsis, Carrierea, Idesia, Bennettiodendron, Dianyuea, and Casearia. Among the 17 species, ten were selected from three sections (sect. Populus, sect. Tacamahaca and sect. Leucoides) of Populus. Five species from monotypic or oligotypic genera Itoa, Poliothyrsis, Carrierea, Idesia, Bennettiodendron, which are considered closely related to Populus and Salix, were sampled. The last two species were from the monotypic genus Dianyuea of Scyphostegioideae (includes two monotypic genera) and the big pantropical genus Casearia of Samydoideae (includes 13 genera and 235 species), respectively (Table 1). Of these taxa, the CN for five genera (Itoa, Poliothyrsis, Carrierea, Bennettiodendron, and Dianyuea) are reported for the first time.
Salicaceae includes three subfamilies, Salicoideae, Scyphostegioideae, and Samydoideae (Fig. 1). Samydoideae has thirteen genera and ca. 235 species [3, 48]. Five species of Casearia, a big pantropical genus in Samydoideae with ca. 180 species, were reported to have 42, 44, or 84 chromosomes in somatic cells (Supplementary Table S1). We also confirm that C. velutina Blume has CN of 2n = 44 (Table 1). The Scyphostegioideae has two monotypic genera, Scyphostegia Stapf and Dianyuea. The somatic karyotype of Scyphostegia is 2n = 18, and CN of Dianyuea is 2n = 38 or 2n = 36 +2B. The chromosomes of D. turbinata (H.J.Dong & H.Peng) C.Shang, S.Liao & Z.X.Zhang are relatively larger than the other species studied (Fig. 2). In Salicoideae, clade A includes six genera (Bembicia Oliv., Homalium Jacq., Azara Ruiz & Pav., Abatia Ruiz & Pav., Banara Aubl., and Prockia P. Browne ex L.), clade B includes eight genera (Hemiscolopia D. F. van Slooten, Scolopia Schreb., Pleuranthodendron L.O. Williams, Xylosma G. Forst., Trimeria Harv., Dovyalis E. Mey. ex Arn., Flacourtia Comm. ex L’Hér., and Oncoba Forssk.), and clade C includes seven genera (Bennettiodendron, Idesia, Poliothyrsus, Itoa, Carrierea, Populus and Salix). In clade A, species from Azara and Prockia are identified with 2n = 18, species of Homalium are based on n = 10 or 11, while a species of Abatia has n = ca. 36. In clade B, Scolopia, Xylosma, Dovyalis, Flacourtia, and Oncoba are based on n = 10 or 11, except for the CN of O. dentata Oliv. (2n = 48), which seems to be a tetraploid (4x = 48) [49]. In clade C, Bennettiodendron and Idesia share the CN of 2n = 42, which is based on n = 21. Poliothyrsus, Itoa and Carrierea share a CN of 2n = 40, which could assume to be derived from the basic chromosome number of n = 21 of Bennettiodendron and Idesia by chromosome fusions [50]. Combining our results with online data, the CN of Populus and Salix are n = 19, and n = 19, 22, 38, 44, 57, 76, 95 respectively. All the Populus species have two obviously long chromosomes as indicated by arrows in Fig. 2, which are not present in poplar relatives studied in this study (Fig. 2). Finally, as an outgroup, the Lacistema aggregatum (P.J. Bergius) Rusby from Lacistemataceae has a chromosome count of 2n = ca. 62.

A summary phylogeny based on published trees of Salicaceae [3, 46]. The inferred chromosome numbers superimposed onto the phylogenetic tree are predominant counts for each genus based on available data. The chromosome numbers and mean 1 C DNA amounts assigned based on cited publications or database are in grey. Hypothesized placement of the salicoid WGD event is indicated with star. Scy, Scyphostegioideae. Sam, Samydoideae. Lac, Lacistemataceae

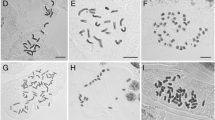
Somatic chromosomes of Populus and its relatives. A Casearia velutina (2n = 44). B-C Dianyuea turbinata (2n = 38). D Bennettiodendron leprosipes (2n = 42). E-G Idesia polycarpa (2n = 42). H Itoa orientalis (2n = 40). I Carrierea calycina (2n = 40). J Poliothyrsis sinensis (2n = 40). K Populus adenopoda (2n = 38). L P. cathayana (2n = 38). M P. ciliata (2n = 38). N P. davidiana (2n = 38). O P. glauca (2n = 38). P P. pamirica (2n = 38). Q P. szechuanica (2n = 38). R P. wilsonii (2n = 38). S P. yatungensis (2n = 38). T P. yunnanensis (2n = 38). Arrows in B-C indicate the possible extra-chromosomes, and in K-T indicate the two large chromosomes of poplars
Salicaceae DNA C-values
Besides chromosome number, we observed representative signatures of chromosome size in different Salicaceae species (Fig. 1; Table 2). By searching the Plant DNA C-values database of Kew, we found only species from three genera of Salicaceae have DNA C-values estimates [43]. The 1 C DNA amount of Casearia bourdillonii Mukh. (Samydoideae) is 0.63 pg. The 1 C DNA amount varies from 0.45 pg in P. tremula L. to 0.55 pg in P. tremuloides in Populus, and it varies from 0.36 pg in S. amygdaloides Andersson (2n = 38) to 0.86 pg in S. ×fragilis L. (2n = 38, 76, 114) in Salix ([43]; Supplementary Table S2). In this study, we estimate the DNA C-values of 35 taxa from 14 genera of Salicaceae by flow cytometric analysis, as illustrated in Fig. 3. In Samydoideae, the 1 C DNA amount of Casearia graveolens Dalz. is 0.696 pg, which is similar to that of C. bourdillonii. In Scyphostegioideae, the 1 C DNA amount of Dianyuea turbinata is 4.315 pg, which is the biggest in Salicaceae. In Salicoideae clade A, two resources of Homalium ceylanicum (Gardner) Benth. have 1 C DNA amount of 0.416 pg in resource C17057 and 0.404 pg in 00GN0039, respectively. However, there is no statistically significance difference between the two resources (p = 0.136, t-test). In Salicoideae clade B, 1 C DNA contents of five genera range from 0.315 pg in Scolopia chinensis (Lour.) Clos to 0.568 pg in Flacourtia indica (Burm. f.) Merr. And they are different from each other (p < 0.002, t-test) except for Xylosma and Oncoba (p = 0.498, t-test). In Salicoideae clade C, Bennettiodendron (n = 21, 1 C DNA amount = 3.296 pg) and Idesia (n = 21, 1 C DNA amount = 1.138-1.211 pg) have relatively large 1 C DNA amounts with almost six and two times of the average (about 0.530 pg) of the other five genera, respectively. The rest five genera have similar 1 C DNA amounts, varying from 0.552 pg in Poliothyrsis to 0.685 pg in Itoa. The 1 C DNA amount of Populus varies from 0.45 pg in P. tremula to 0.577 pg in P. euphratica Olivier, except 0.705 pg in an individual of P. suaveolens Fisch. which probably represent a triploid. In Salix, it varies from 0.36 to 0.86 pg, which reflect the frequent polyploidy in the genus.

Estimation of nuclear DNA amount (genome size). The histograms of PI fluorescence were obtained by flow cytometric analysis. Glycine max Williams 82 served as internal standard for Populus szechuanica (A) and Carrierea calycina (B), while Zea mays L. B73 served as internal standard for Idesia polycarpa (C) and Dianyuea turbinate (D). Small peaks at about twice the DNA content of the marker G1 peak represent the cells in G2 phase
Discussion
The taxonomic implications of chromosome number and 1 C DNA amount
The monotypic Dianyuea includes D. turbinate, which is an enigmatic species endemic to the western Yunnan Province, China. It was first described and placed in the genus Flacourtia Comm. ex L’Hér. of Salicoideae as F. turbinata H.J. Dong & H. Peng in 2013 [51]. Using plastid trnL-F, matK, and rbcL sequence, Shang et al. found F. turbinata and the genus Scyphostegia of Scyphostegioideae form a strongly supported clade which is sister to the crown group of Salicoideae [46]. These indicate that F. turbinata does not belong to Salicoideae, and should be separated from Flacourtia. In addition, F. turbinata has many characteristics which are different from that of Flacourtia. For example, male flowers with six stamens connate to form a column (polystemonous and free in Flacourtia), basal placentation (parietal in Flacourtia), capsule (berry in Flacourtia), and fleshy, pubescent, and lobed appendages around seeds (absent in Flacourtia) [46]. Previous studies show the chromosome number (CN) of Flacourtia is n = 10 or 11. Our results show that the CN of F. turbinata is 2n = 38 or 36 +2B. Among all the 38 chromosomes, there are also two small chromosomes without any visible constriction stably appeared in almost all metaphase cells of F. turbinata examined. If consider them as two extra-chromosomes, the CN of F. turbinate is alternatively 2n = 36 + 2B. In order to provide conclusive evidence, further research is needed. The genome size of F. turbinata (4.315 pg) is much bigger than that of Flacourtia (0.433-0.568 pg). The chromosome number and genome size give additional evidence for excluding F. turbinata from Flacourtia.
The monotypic Scyphostegia includes S. borneensis Stapf, which is endemic to the northern part of Borneo [52]. It has been placed in or considered to be closely related to several different and distantly-related families, including Monimiaceae, Moraceae, Tamaricaceae, and Flacourtiaceae due to its unusual combination of external morphology (dioecy, basal placentation, 3-merous flowers, and telescoping inflorescence bracts) and anatomical features (stem, leaf, flower, and fruit) [52,53,54,55]. Shang et al. found a strongly supported sister relationship of Dianyuea and Scyphostegia, and they are sister to all taxa of Salicoideae [46]. The CN of Scyphostegia and Dianyuea are 2n = 18 and 38 (perhaps 36 + 2B), respectively. They are possibly based on the same basic chromosome number, and a polyploid event probably happened in Dianyuea, which is fairly common in Malpighiales [56]. Therefore, our results provide additional evidence for the sister relationship between Dianyuea and Scyphostegia, which indicate by molecular phylogenetic study [46].
The identity of the sister taxon of Populus-Salix and the relationship of Populus, Salix, and their relatives have been long-term discussed and remained controversial. There are two proposed phylogenetic relationships. In the first situation, the two Asia genera Idesia and Bennettiodendron have closer relationships with Populus-Salix than Poliothyrsis, Itoa, and Carrierea. This is supported by a phylogenetic study of Malpighiales using 13 gene regions, including 10 plasmid genes and 3 nuclear genes [7]. A similar relationship has been revealed by Xi et al. [8] and Zhang et al. [6] using plastome sequence phylogeny. Besides, the close relationship of Idesia with Populus and Salix was supported by the occurrence of the rust fungus, Melampsora, in Idesia, Populus and Salix [9]. In the second situation, the relationships of Poliothyrsis, Itoa, Carrierea and Populus-Salix are closer than that between Idesia and Populus-Salix as illustrated in Fig. 1. This relationship is supported by the landmark phylogenetic research of Salicaceae which used plastid rbcL DNA sequence, and included a comprehensive sampling of Salicaceae [3]. Both evolutionary relationships are mainly based on plasmid sequences, and may be affected by chloroplast capture. Of the 3 subfamilies and 55 genera in Salicaceae, the three works support the first relationship covered 6 genera (2 subfamilies), 11 genera (3 subfamilies), and 11 genera (3 subfamilies), respectively [6,7,8]. In the second proposed phylogeny, they sampled 22 genera (3 subfamilies). More and more studies show that even if the same gene marker set is used, the difference of taxon sampling density will lead to the contradiction of phylogenetic trees [56]. And the increasing importance of taxon sampling for phylogenetic inference has been proposed and high-lightened [56,57,58]. Our results in the chromosome number and genome size also provide some hints. The history of genome duplication events in Populus provided evidence that the progenitor of Populus had a base chromosome number of 10. The salicoid whole-genome duplication (WGD) led to the doubling of chromosome number and the subsequently genome-wide reorganization and joining of chromosomes result in the n = 19 chromosome karyotype of Populus [59]. Itoa, Poliothyrsis, and Carrierea all share the CN of n = 20. Their DNA contents are similar with that of Populus and Salix. All the DNA C-values of these five genera are ranged from 0.36 to 0.685 pg, and even the polyploidy genomes of Populus and Salix are less than 1 pg. Idesia and Bennettiodendron both share the CN of n = 21. They also have much bigger DNA C-values up to 1.211 pg and 3.296 pg, respectively. In addition, Itoa, Poliothyrsis, and Carrierea have more morphological similarities with Populus and Salix than Idesia and Bennettiodendron, for example, the capsule and winged seeds. Therefore, we adopt the second phylogeny in this study. However, we can’t exclude other possibilities until in-depth analysis with more data and more comprehensive sampling are performed.
The phylogenetic placement of the salicoid whole-genome duplication
The salicoid WGD event is present in all sequenced poplars and willows [22, 59,60,61,62,63,64]. The time of salicoid WGD was deduced as 8 to 13 Ma when naively calibrated the molecular clock using synonymous rates observed in the Brassicaceae [59]. However, the WGD event is probably shared by poplars and willows [22, 59], and fossil record shows that the Populus and Salix lineages diverged 60 to 65 Ma [65, 66]. Thus, the salicoid WGD is placed at or near the lineages diverged time of 60 to 65 Ma [59]. This time point is coincident with the previous hypothesis that multiple WGD events in independent lineages of land plants appear to cluster around the Cretaceous - Paleogene (K-Pg) boundary, around 66 Ma [16]. In addition, the Salicoideae was supposed to split from Scyphostegioideae at 68.9 (78.7-59.8) Ma [8]. In this study, we found species in clade A and B of Salicoideae mainly have n = 9, 10, or 11 and all species in clade C have twice or more CNs (Fig. 1). Thus, the occurrence of WGD event in the crown group of Salicoideae clade C is the most parsimony evolutionary scenario. Otherwise, if assume the WGD event in the crown group of the whole Salicoideae, we have to suppose a reduction of ploidy level in clade A and B which is improbability due to failure of homologous pairing in meiosis and the fact that, polyploid abundance is only expected to increase over time, since polyploidization is an irreversible process [67]. If assume several WGD events independently occurred in genera of clade C, genome-wide reconstruction of gene families and molecular clock analysis across these genera are required to confirm that. However, we must take this conclusion carefully before dense sampling of genomic sequence data are investigated.
The success of poplars and willows
Polyploidy is thought to be a major evolutionary driving force in angiosperm diversification [14]. However, there is often a lag-time or delay between the WGD event and subsequent radiations [68, 69]. In clade C, Poliothyrsis, Itoa, Carrierea, Idesia, Bennettiodendron, Olmediella, Macrohasseltia are considered to be sister genera close to Populus and Salix [4, 7, 8]. It is noteworthy that the position of Olmediella and Macrohasseltia is uncertain, so they are excluded from the tree in Fig. 1. Altogether, Poliothyrsis, Idesia, Olmediella and Macrohasseltia are monotypic, while Itoa, Carrierea, and Bennettiodendron all have two species respectively [70]. For comparison, Populus and Salix including approximately 22-45 and 330-500 species, respectively [1, 71]. Our results indicate that the WGD may occur in the crown group of clade C. The species richness in Populus-Salix group and species poverty in their relatives suggest that there is a lag-time for the subsequent radiations after the salicoid WGD event in clade C.
The WGD radiation lag-time model proposed by Schranz et al. suggested that major radiation events are likely not directly driven by the WGDs, but rather by secondary dispersal events triggered by later changing environmental conditions (climate, geological, etc.), evolutionary arms races (e.g. herbivore and plant host), co-radiations (e.g. specialized pollinator and plant host), and migration events into new environments [68]. According to our own observation and literature records, all the seven sister genera of Populus and Salix tend to have a narrow and fragmentized habitat, and they are common but not key elements of their habitat [70, 72]. The first five genera (Poliothyrsis, Itoa, Carrierea, Idesia, and Bennettiodendron) are restricted to East Asia and Southeast Asia, and the latter two genera (Olmediella and Macrohasseltia) are endemic to Central America [4]. They survive in tropical and subtropical forest except for Idesia which can reach the southern edge of temperate zone [70]. Populus and Salix have their maximum species richness in temperate regions of the northern hemisphere and are diversified extensively in high latitude [1]. Many poplars and willows are keystone species in northern hemisphere especially in riparian forest [1].
In conclusion, the radiation and adaptation of Populus and Salix might be driven by both the WGD and environmental changes. As our prediction, the seven genera in Clade C shared a WGD, which provided sources for adaptation, speciation and evolution. After the WGD, five genera were retained in narrow and fragmentized habitats, while Populus and Salix migrated to colder environments. A suit of adaptive innovations, including cold tolerance (in almost all poplars and willows), drought tolerance (Populus euphratica, P. alba, etc.), and plateau adaptability (in Salix sect. Lindleyanae) enable Populus-Salix to occupy northern hemisphere temperate area.
Conclusions
In this study, we report the somatic CN of seventeen species from eight genera in Salicaceae. Of these, CNs for twelve species (Itoa orientalis, Poliothyrsis sinensis, Carrierea calycina, Bennettiodendron leprosipes, Dianyuea turbinata, Casearia velutina, Populus glauca, P. pamirica, P. szechuanica, P. wilsonii, P. yatungensis, and P. yunnanensis) and for five genera (Itoa, Poliothyrsis, Carrierea, Bennettiodendron, and Dianyuea) are reported for the first time. The CN of Idesia polycarpa was confirmed to be 2n = 42. The genome sizes of 35 taxa belonging to 14 genera of Populus and Populus-relatives were estimated. Of these, the genome size of 12 genera (Dianyuea, Homalium, Scolopia, Xylosma, Dovyalis, Flacourtia, Oncoba, Bennettiodendron, Idesia, Carrierea, Poliothyrsis, and Itoa) and of all taxa except Populus euphratica are first reported. Our research greatly enriched the basic cytological characteristics of Salicaceae. The variation of CN and genome size across Salicaceae indicate frequent ploidy changes and a widespread sharing of the salicoid whole genome duplication (WGD) by the relatives of Populus and Salix. The phylogenetic asymmetry in clade of Populus, Salix, and their close relatives suggests that there is a lag-time for the subsequent radiations after the salicoid WGD event. Our results provide useful data for studying the evolutionary events of Salicaceae.
Methods
Taxon sampling and identification
In this study, we included 17 taxa from eight genera for cytological analysis (Table 1) and 35 taxa from fourteen genera for genome size estimation (Table 2). Individuals were collected from field work by Zhong-Shuai Zhang in the vast area of China conducted in 2020 and 2021, as well as Xishuangbanna Tropical Botanical Garden and Kunming Botanical Garden in China (Tables 1 and 2). Where possible, more than one and up to six individuals were included per taxon. Sampled individuals were identified by all the authors according to appropriate literatures, and type materials from different herbariums. Due to the taxonomy of Populus is still in heated debate and there is no recent taxonomic revisions of many genera studied in this article. We give all the sampling site and relevant pictures to aid identification (Figs. 4, 5, 6 and 7). The voucher specimens of the studied materials are all preserved in the herbarium of Chinese Academy of Forestry (CAF) [73]. The source numbers of voucher specimens are listed in Tables 1 and 2.

Morphological features of Salicaceae (I). (A) Bennettiodendron leprosipes, (B-C) Carrierea calycina, (D) Casearia velutina, (E) C. graveolens, (F-H) Dianyuea turbinate, (I-J) Dovyalis caffra, (K-L) Flacourtia indica, (M) F. inermis, (N) F. rukam, (O) Homalium ceylanicum, and (P) Idesia polycarpa

Morphological features of Salicaceae (II). (A) Itoa orientalis, (B) Oncoba echinate, (C) Poliothyrsis sinensis, (D-E) Scolopia chinensis, (F-G) Xylosma congesta, (H-I) X. longifolia, (J-K) Populus adenopoda, (L-M) P. cathayana, (N-O) P. ciliate, (P-Q) P. davidiana, and (R-T) P. glauca

Morphological features of Salicaceae (III). (A-C) Populus haoana var. megaphylla, (D-F) P. kangdingensis, (G-I) P. koreana, (J-L) P. lasiocarpa, (M-O) P. pamirica, (P) P. qiongdaoensis, (Q-R) P. rotundifolia, (S-T) P. simonii, (U-V) P. suaveolens, and (W-Y) P. szechuanica

Morphological features of Salicaceae (IV). (A-C) Populus wilsonii, (D-F) P. yatungensis, (G-I) P. yuana, (J-L) P. yunnanensis, and (M-O) P. yunnanensis var. pedicellata
Cytological analysis
Branch cuttings or seedlings of samples were collected and planted in flowerpots in the greenhouse at 25℃. Vigorous root tips were pre-treated with ice-water mixture in dark room for 24 h. After incubation, the tips were fixed in Carnoy I solution (3:1 ethanol: glacial acetic acid) at 4 °C for at least 3 h. They were then digested at 37 °C in a combination (1:1) of 2 % cellulase and 2 % pectinase for 30 to 60 min before staining with an improved carbol-fuchsin solution and squashed for cytological observation [74]. Standard liquid nitrogen method was used to make permanent slides that were preserved at Chinese Academy of Forestry. The photo micrographs were taken using an Axio Imager A1 microscope (Zeiss, Germany). Only complete cells with clear outline and scattered chromosomes were selected for observation. The chromosome number of each taxon was determined by checking multiple random selected mitosis metaphase cells of individuals. We detected more than 5, up to 42 (in Dianyuea turbinata) cells of root tips of each taxon, and determined the chromosome number only when all cells showed the same count.
Flow cytometry
The fresh leaves of the majority individuals were collected from the transplanted plants cultivated in greenhouse. These leaves were kept on ice and used for flow cytometer analysis within 12 h. The materials from Xishuangbanna Tropical Botanical Garden and Kunming Botanical Garden were collected directly from the garden trees. For botanical garden materials, populations from different origin are labeled by resource numbers. The materials were stored at 0 ℃ immediately and measured within three days. One to three technical repetitions were tested according to the availability of materials. We used internal standards for all measurements and the internal standards were selected based on appropriate non-overlapping genome size. Fresh leaves of Zea mays L. B73 (2.425 pg/1 C) were used as internal standards for Dianyuea turbinata and Idesia polycarpa [75]. Fresh leaves of Glycine max (L.) Merr. Williams 82 (1.155 pg/1 C) were used as internal standards for the rest samples [76]. Approximately 0.5 cm2 leaf of the standard and samples were co-chopped with a sharp razor blade for ca. 10 to 20 s in a Petri dish containing 0.25 mL ice-cold nuclei extracting buffer (30 mmol/L Na3C6H5O7·2H2O, 45 mmol/L MgCl2, 20 mmol/L MOPS, 20 mmol/L NaCl, 20 mmol/L EDTA-Na2, 0.1 % volume percentage Triton X-100, 0.5 % volume percentage Tween-20, 1 % volume percentage PVP, pH=7.0). The nuclei extracting buffer is slightly modified from Galbraith’s buffer and was preserved at 4℃ until use [77]. The homogenate was gently sucked up by pipette and passed through 48 μm nylon mesh filters into 5 mL plastic round-bottom Falcon tubes (Corning, New York, N.Y., USA). A volume of 0.5 ml staining buffer (CyStain PI Absolute P, Sysmex Partec GmbH Görlitz, Germany), 3 µl propidium iodide (CyStain PI Absolute P, Sysmex Partec GmbH Görlitz, Germany) and 1.5 µl RNaseA were added and mixed by gentle shaking. Samples were incubated with the staining solution on ice for 15 min in darkness prior to flow cytometry analysis. The homogenates were analyzed based on light scatter and fluorescence signals produced from 20 mW laser illumination at 488 nm using a BD LSRFortessaTM cell analyzer (BD Biosciences, Franklin Lakes, NJ). At least 3 × 103 nuclei were collected in each measurement. Data were collected and analyzed by BD FACSDiva 7.0 (BD Biosciences, Franklin Lakes, NJ). The coefficient of variation among nuclei (CVn) was calculated as follow: CVn = SD/M, where SD was the standard deviation of the nuclei distribution, and M was the mean channel number [78]. We performed a pre-analyze on some samples of Populus and all samples of the other genera. We collected the PI fluorescence intensity of each sample without internal standard and checked whether there were several peaks arranged in an endoreplication fashion first. If endoreplication exists, there will be additional peaks with 8 C, 16 C, 32 C and even higher DNA levels besides the 2 C (G1) and 4 C (G2) peaks (Response Fig. 1 A). As a result, we did not find any polyploidization peaks of DNA. Then, we preformed analysis on samples with internal standard, and used the two large peaks representing G1 nuclei of the reference and the sample to estimate the DNA contents. The formula 1 C DNA content (pg) of the standard × average of sample G1 peak / average of standard G1 peak was used to estimate the 1 C DNA content of sample cells at G1 phase.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files. The voucher specimens of the materials analyzed during the current study are preserved in the herbarium of Chinese Academy of Forestry (CAF) and available from the corresponding author on reasonable request.
References
Dickmann DI, Kuzovkina J. Poplars and willows of the world, with emphasis on silviculturally important species. In: Isebrands JG, Richardson J. editors. Poplars and Willows: Trees for Society and the Environment. Rome: CAB International & FAO; 2014. p. 8–91.
Jansson S, Douglas CJ. Populus: A model system for plant biology. Annu. Rev. Plant Biol. 2007;58:435–58.
Chase MW, Zmarzty S, Lledó MD, Wurdack KJ, Swensen SM, Fay MF. When in doubt, put it in Flacourtiaceae: a molecular phylogenetic analysis based on plastid rbcL DNA sequences. Kew Bull. 2002;57:141–81.
Alford MA. Systematics studies in Flacourtiaceae. PhD thesis. Cornell University; 2005.
The Angiosperm Phylogeny Group, Chase MW, Christenhusz MJM, Fay MF, Byng JW, Judd WS, et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016;181(1):1–20.
Zhang L, Xi ZX, Wang MC, Guo XY, Ma T. Plastome phylogeny and lineage diversification of Salicaceae with focus on poplars and willows. Ecol. Evol. 2018;8(16):7817–23.
Wurdack KJ, Davis CC. Malpighiales phylogenetics: gaining ground on one of the most recalcitrant clades in the angiosperm tree of life. Am. J. Bot. 2009;96(8):1551–70.
Xi Z, Ruhfel BR, Schaefer H, Amorim AM, Sugumaran M, Wurdack KJ, et al. Phylogenomics and a posteriori data partitioning resolve the Cretaceous angiosperm radiation in Malpighiales. Proc Natl Acad Sci USA. 2012;109(43):17519–24.
Cronquist A. An integrated system of classification of flowering plants. New York: Columbia University Press; 1981.
Alix K, Gérard PR, Schwarzacher T, Heslop-Harrison JS. Polyploidy and interspecific hybridization: partners for adaptation, speciation and evolution in plants. Ann. Bot. 2017;120(2):183–94.
Landis JB, Soltis DE, Li Z, Marx HE, Barker MS, Tank DC, et al. Impact of whole-genome duplication events on diversification rates in angiosperms. Am. J. Bot. 2018;105(3):348–63.
Darlington CD. 1936. Recent advances in cytology. Philadelphia: Blakiston; 1936.
Stebbins GL. Variation and evolution in plants. New York: Columbia University Press; 1950.
Soltis DE, Albert VA, Leebens-Mack J, Bell CD, Paterson A, Zheng C, et al. Polyploidy and angiosperm diversification. Am. J. Bot. 2009;96(1):336–48.
Jiao YN, Wickett NJ, Ayyampalayam S, Chanderbali AS, Landherr L, Ralph PE, et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011;473(7345):97–100.
Clark JW, Donoghue PC. Whole-genome duplication and plant macroevolution. Trends Plant Sci. 2018;23(10):933–45.
McCarthy E, Chase M, Knapp S, Litt M, Leitch AR, Le Comber S. Transgressive phenotypes and generalist pollination in the floral evolution of Nicotiana polyploids. Nat. Plants 2016;16119.
Segraves KA, Anneberg TJ. Species interactions and plant polyploidy. Am. J. Bot. 2016;103(10):1–10.
Gross K, Schiestl FP. Are tetraploids more successful? Floral signals, reproductive success and floral isolation in mixed-ploidy populations of a terrestrial orchid. Ann. Bot. 2015;115:263–73.
McCarthy EW, Arnold SEJ, Chittka L, Le Comber SC, Verity R, Dodsworth S, et al. The effect of polyploidy and hybridization on the evolution of floral colour in Nicotiana (Solanaceae). Ann. Bot. 2015;115(7):1117–31.
Cannon SB, McKain MR, Harkess A, Nelson MN, Dash S, Deyholos MK et al. Multiple polyploidy events in the early radiation of nodulating and nonnodulating legumes. Mol. Biol. Evol. 2015;32(1):193–210.
Cai L, Xi Z, Amorim AM, Sugumaran M, Rest JS, Liu L, et al. Widespread ancient whole-genome duplications in Malpighiales coincide with Eocene global climatic upheaval. New Phytol. 2019;221:565–76.
Greilhuber J, Doležel J, Lysák MA, Bennett MD. The origin, evolution and proposed stabilization of the terms ‘genome size’ and ‘C-value’ to describe nuclear DNA contents. Ann. Bot. 2005;95(1):255–60.
Fleischmann A, Michael TP, Rivadavia F, Sousa A, Wang WQ, Temsch EM et al. Evolution of genome size and chromosome number in the carnivorous plant genus Genlisea (Lentibulariaceae), with a new estimate of the minimum genome size in angiosperms. Ann. Bot. 2014;114(8):1651–63.
Pellicer J, Michael FF, Ilia JL. The largest eukaryotic genome of them all?. Bot. J. Linn. Soc. 2010;164(1):10–5.
Leitch IJ, Kahandawala I, Suda J, Hanson L, Ingrouille MJ, Chase MW et al. Genome size diversity in orchids: consequences and evolution. Ann. Bot. 2019;104(3):469–81.
Bai CK, Alverson WS, Follansbee A, Waller DM. New reports of nuclear DNA content for 407 vascular plant taxa from the United States. Ann. Bot. 2012;110(8):1623–9.
Jersáková J, Trávníček P, Kubátová B, Krejčíková J, Urfus T, Liu ZJ et al. Genome size variation in Orchidaceae subfamily Apostasioideae: filling the phylogenetic gap. Bot. J. Linn. Soc. 2013;172(1):95–105.
Pellicer J, Hidalgo O, Dodsworth S, Leitch IJ. Genome size diversity and its impact on the evolution of land plants. Genes 2018;9(88):1–14.
Trávníček P, Čertner M, Ponert J, Chumová Z, Jersáková J, Suda J. Diversity in genome size and GC content shows adaptive potential in orchids and is closely linked to partial endoreplication, plant life-history traits and climatic conditions. New Phytol. 2019;224:1642–56.
Roddy AB, Théroux-Rancourt G, Abbo T, Benedetti JW, Brodersen CR, Castro M, et al. The scaling of genome size and cell size limits maximum rates of photosynthesis with implications for ecological strategies. Int. J. Plant Sci. 2020;181(1):75–87.
Veselý P, Bureš P, Šmarda P, Pavlíček T. Genome size and DNA base composition of geophytes: the mirror of phenology and ecology? Ann. Bot. 2012;109(1):65–75.
Meudt HM, Rojas-Andrés BM, Prebble JM, Low E, Garnock-Jones PJ, Albach DC. Is genome downsizing associated with diversification in polyploid lineages of Veronica? Bot. J. Linn. Soc. 2015;178(2):243–66.
Carović-Stanko K, Liber Z, Besendorfer V, Javornik B, Bohanec B, Kolak I, et al. Genetic relations among basil taxa (Ocimum L.) based on molecular markers, nuclear DNA content, and chromosome number. Plant Syst. Evol. 2009;285(1):13–22.
Murray BG, De Lange PJ, Ferguson AR. Nuclear DNA variation, chromosome numbers and polyploidy in the endemic and indigenous grass flora of New Zealand. Ann. Bot. 2005;96(7):1293–305.
Eckenwalder JE. Systematics and evolution of Populus. In: Stettler RF, Bradshaw HD, Heilman PE, Hinckley TM, editors. Biology of Populus, and its Implications for Management and Conservatio. Ottawa: NRC Research Press; 1996. p. 7–32.
Rice A, Glick L, Abadi S, Einhorn M, Kopelman NM, Salman-Minkov A, et al. The Chromosome Counts Database (CCDB) - a community resource of plant chromosome numbers. New Phytol. 2015;206(1):19–26.
Mock KE, Callahan CM, Islam-Faridi MN, Shaw JD, Rai HS, Sanderson SC, et al. Widespread triploidy in western North American aspen (Populus tremuloides). PLoS ONE 2012;7(10):e48406.
Wilkinson J. The cytology of Salix in relation to its taxonomy. Ann. Bot. 1944;8(30/31):269–84.
Blackburn KB, Harrison JWH. A preliminary account of the chromosomes and chromosome behavior in the Salicaceae. Ann. Bot. 1924;38(105):361–78.
Suda Y, Argus GW. Chromosome numbers of some North American Salix. Brittonia 1968;20(3):191-7.
Darlington CD, Wylie AP. Chromosome atlas of flowering plants. London: George Allen & Unwin; 1955.
Pellicer J, Leitch IJ. The Plant DNA C-values database (release 7.1): an updated online repository of plant genome size data for comparative studies. New Phytol. 2020;226:301–5.
Baker WJ, Bailey P, Barber V, Barker A, Bellot S, Bishop D, et al. A comprehensive phylogenomic platform for exploring the angiosperm Tree of Life. BioRxiv. 2021;doi: https://doi.org/10.1101/2021.02.22.431589.
Boucher LD, Manchester SR, Judd WS. An extinct genus of Salicaceae based on twigs with attached flowers, fruits, and foliage from the Eocene Green River Formation of Utah and Colorado, USA. Am. J. Bot. 2003;90(9):1389–99.
Shang C, Liao S, Guo YJ, Zhang ZX. Dianyuea gen. nov. (Salicaceae: Scyphostegioideae) from southwestern China. Nord. J. Bot. 2017;35(4):499-505.
Tokuoka T, Tobe H. Phylogenetic analyses of Malpighiales using plastid and nuclear DNA sequences, with particular reference to the embryology of Euphorbiaceae sens. str. J. Plant Res. 2006;119(6):599–616.
Stevens PF. Angiosperm Phylogeny Website. Version 14. 2001 (onwards). http://www.mobot.org/MOBOT/research/APweb/. Accessed 30 Aug 2021.
Gadella TWJ. Cytological studies on some flowering plants collected in Africa. Bull. Jard. Bot. Natl. Belg. (Impr.) 1972;42(4):393–402.
Schubert I. Chromosome evolution. Curr. Opin. Plant Biol. 2007;10(2):109–15.
Dong HJ, Peng H. Flacourtia turbinata (Salicaceae: Flacourtieae), a new species from Yunnan, China. Phytotaxa 2013;94(2):56–60.
Van Steenis CGGJ. Scyphostegiaceae. In: Van Steenis CGGJ, editor. Flora Malesiana. Ser. I. Vol. 5. Djakarta: Noordhoff-Kolff N.V.; 1957. p. 297–99.
Swamy BGL. On the floral structure of Scyphostegia. Proc. Natl. Acad. Sci. India Sect B Bio.l Sci. 1953;19:127-42.
Van Heel WA. Anatomical and ontogenetic investigations on the morphology of the flowers and the fruit of Scyphostegia borneensis Stapf (Scyphostegiaceae). Blumea 1967;15(1):107–25.
Alford MA. Scyphostegia borneensis. In: The Tree of Life Web Project. 2007. http://tolweb.org/Scyphostegia_borneensis/68360/2007.02.06. Accessed 30 Aug 2021.
Natsidis P, Tsakogiannis A, Pavlidis P, Tsigenopoulos CS, Manousaki T. Phylogenomics investigation of sparids (Teleostei: Spariformes) using high-quality proteomes highlights the importance of taxon sampling. Commun. Biol. 2019; 2(400):1–10.
Nabhan AR, Sarkar IN. The impact of taxon sampling on phylogenetic inference: a review of two decades of controversy, Brief. Bioinformatics. 2012;13(1):122–34.
Majure LC, Soltis DE, Soltis PS. Effects of taxon sampling and tree reconstruction methods on phylodiversity metrics. Ecol. Evol. 2019;9:9479–99.
Tuskan GA, DiFazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006;313(5793):1596–604.
Ma T, Wang JY, Zhou GK, Yue Z, Hu QJ, Chen Y, et al. Genomic insights into salt adaptation in a desert poplar. Nat. Commun. 2013;4(2797):1–7.
Dai XG, Hu QJ, Cai QL, Feng K, Ye N, Tuskan GA, et al. The willow genome and divergent evolution from poplar after the common genome duplication. Cell Res. 2014;24(10):1274–7.
Yang WL, Wang K, Zhang J, Ma JC, Liu JQ, Ma, T. The draft genome sequence of a desert tree Populus pruinosa. Gigascience 2017;6(9):1–7.
Lin YC, Wang J, Delhomme N, Schithaler B, Sundström G, Zuccolo A, et al. Functional and evolutionary genomic inferences in Populus through genome and population sequencing of American and European aspen. Proc. Natl. Acad. Sci. U.S.A. 2018;115:E10970-E10978.
Liu YJ, Wang XR, Zeng QY. De novo assembly of white poplar genome and genetic diversity of white poplar population in Irtysh River basin in China. Sci. China Life Sci. 2019;62(5):609–18.
Manchester SR, Dilcher DL, Tidwell WD. Interconnected reproductive and vegetative remains of Populus (Salicaceae) from the middle Eocene Green River Formation, northeastern Utah. Am. J. Bot. 1986;73(1):156–60.
Collinson ME. The early fossil history of Salicaceae: a brief review. Proc. R. Soc. Edinb. 1992;98B:155-67.
Van de Peer Y, Mizrachi E, Marchal K. The evolutionary significance of polyploidy. Nat. Rev. Genet. 2017;18(7):411–24.
Schranz ME, Mohammadin S, Edger, PP. Ancient whole genome duplications, novelty and diversification: the WGD Radiation Lag-Time Model. Curr. Opin. Plant Biol. 2012;15(2):147–53.
Tank DC, Eastman JM, Pennell MW, Soltis PS, Soltis DE, Hinchliff CE, et al. Nested radiations and the pulse of angiosperm diversification: Increased diversification rates often follow whole genome duplications. New Phytol. 2015;207(2):454–67.
Yang QE, Zmarzty S. 2007. Flacourtiaceae. In: Wu ZY, Raven PH, editors. Flora of China, vol. 13. Beijing: Science Press & St. Louis.: Missouri Botanical Garden Press; p. 112–37.
Cronk QCB. Plant eco-devo: the potential of poplar as a model organism. New Phytol. 2005;166(1):39–48.
Alford MH, Belyaeva IV. Neotropical Salicaceae. In: Milliken W, Klitgård B, Baracat A, editors. Neotropikey-Interactive key and information resources for flowering plants of the Neotropics. 2009. http://www.kew.org/science/tropamerica/neotropikey/families/Salicaceae.htm. Accessed 30 Aug, 2021.
Thiers B. Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. 2016. http://sweetgum.nybg.org/science/ih/. Accessed 1 July 2021.
Zhu C. Plant chromosome and chromosome technology. Beijing: Science Press; 1982.
Rayburn AL, Biradar DP, Bullock DG, Mcmurphy LM. Nuclear-DNA content in F1 hybrids of maize. Heredity 1993;70(3):294–300.
Arumuganathan K, Earle ED. Nuclear DNA content of some important plant species. Plant Mol. Biol. Rep. 1991;9(3):208–18.
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP, Firoozabady E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983;220(4601):1049–51.
Ormerod MG. Flow cytometry: a basic introduction. 2008. https://flowbook.denovosoftware.com/. Accessed 3 Aug 2021.
Acknowledgements
We thank Dr. Zhang Jindan from Plant Science Facility at the Institute of Botany, Chinese Academy of Sciences for her excellent technical assistance on flow cytometry. We thank Xishuangbanna Tropical Botanical Garden (XTBG) and Ding Hongbo, Sheng Caiyu, Mo Haibo for their kind help in sample collection in XTBG. We also thank Kunming Botanical Garden and Jiang Liqiong from Yunnan University for kindly providing us with fresh materials of Xylosma congesta and Poliothyrsis sinensis for flow cytometry analysis. We acknowledge Zhu Xinxin from Xinyang Normal University, Jiang Rihong from Guangxi Academy of Forestry and Xiao Bo from Malipo County, Yunnan province for helping us collect tree seedlings of Populus glauca, Itoa orientalis, and Bennettiodendron leprosipes respectively.
Funding
This study was supported by the National Natural Science Foundation of China (31822011 and 32001329) and the fund of the State Key Laboratory of Tree Genetics and Breeding, Chinese Academy of Forestry (TGB2020001 and TGB2021001).
Author information
Authors and Affiliations
Contributions
Y.-J.L. and Q.-Y.Z. designed research; Z.-S.Z. collected materials, performed the experiments, and analyzed the data; Y.-J.L. and Z.-S.Z. wrote the article. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, ZS., Zeng, QY. & Liu, YJ. Frequent ploidy changes in Salicaceae indicates widespread sharing of the salicoid whole genome duplication by the relatives of Populus L. and Salix L.. BMC Plant Biol 21, 535 (2021). https://doi.org/10.1186/s12870-021-03313-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-021-03313-x