
Abstract
Background
The dwarfing gene sdw1 has been widely used throughout the world to develop commercial barley varieties. There are at least four different alleles at the sdw1 locus.
Results
Mutations in the gibberellin 20-oxidase gene (HvGA20ox2) resulted in multiple alleles at the sdw1 locus. The sdw1.d allele from Diamant is due to a 7-bp deletion in exon 1, while the sdw1.c allele from Abed Denso has 1-bp deletion and a 4-bp insertion in the 5’ untranslated region. The sdw1.a allele from Jotun resulted from a total deletion of the HvGA20ox2 gene. The structural changes result in lower gene expression in sdw1.d and lack of expression in sdw1.a. There are three HvGA20ox genes in the barley genome. The partial or total loss of function of the HvGA20ox2 gene could be compensated by enhanced expression of its homolog HvGA20ox1and HvGA20ox3. A diagnostic molecular marker was developed to differentiate between the wild-type, sdw1.d and sdw1.a alleles and another molecular marker for differentiation of sdw1.c and sdw1.a. The markers were further tested in 197 barley varieties, out of which 28 had the sdw1.d allele and two varieties the sdw1.a allele. To date, the sdw1.d and sdw1.a alleles have only been detected in the modern barley varieties and lines.
Conclusions
The results provided further proof that the gibberellin 20-oxidase gene (HvGA20ox2) is the functional gene of the barley sdw1 mutants. Different deletions resulted in different functional alleles for different breeding purposes. Truncated protein could maintain partial function. Partial or total loss of function of the HvGA20ox2 gene could be compensated by enhanced expression of its homolog HvGA20ox1 and HvGA20ox3.
Similar content being viewed by others

Background
Semi-dwarfism is a valuable and widely used trait in intensive agriculture. The high yield potential of semi-dwarf cultivars is attributed to their improved harvest index, lodging resistance, and more efficient utilization of the environment [1]. The green revolution, led by semi-dwarf varieties in wheat, was due to the introduction of the Rht gene, which encodes a mutant form of a DELLA protein, a gibberellin signaling repressor [2]. The green revolution in rice was due to semi-dwarf varieties carrying sd1, a single locus encoding a defective gibberellin 20-oxidase-2 (GA20ox2) [3].
Semi-dwarf barley cultivars have been successfully used around the world. In China, more than 350 dwarf and semi-dwarf cultivars and entries have been developed since 1950, with an average 4.7-fold yield increase over landraces and older cultivars [4]. There are more than 30 types of dwarfs or semi-dwarfs described in barley, among which semi-brachytic 1 (uzu1), breviaristatum-e (ari-e), and semi-dwarf 1 (sdw1) are widely used in modern barley improvement [5, 6]. The ari-e mutant from Golden Promise has been used in several European cultivars and is located on chromosome 5HL [7]. The uzu gene is located on chromosome 3HL, which has been the major dwarfing gene used in East Asia barley breeding programs [8, 9]. The dwarfism controlled by uzu is caused by a missense mutation of a single nucleotide substitution in the HvBRI1 gene, which reduces the response to brassinolide [9].
The sdw1 locus has been widely used to develop modern barley varieties in Europe, North America, South America, and Australia. There are at least four alleles at the sdw1 locus, which arose from separate mutation events: sdw1.a (originally named sdw1), sdw1.c (originally named denso), sdw1.d (Diamant) and sdw1.e (mutant line ‘Ris∅ no. 9265’) [10]. The sdw1.c allele was the first reported allele at the sdw1 locus, a spontaneous mutant selected from barley cultivar Abed Denso [11]. The sdw1.c allele was successfully transferred to cultivars Deba Abed and Maris Mink, and later introduced into numerous barley crosses in Southern Swedish and Danish breeding programs [6]. The sdw1.a allele was induced by X-ray mutagenesis in a Norwegian six-rowed barley Jotun and has been used in Western USA, Canada, and Australia to breed semi-dwarf feed barley cultivars like Yerong and UC828 [12–14]. The sdw1.d allele, probably the most important for breeding, originated from a mutant selected in the M2 generation of cv. Valticky after X-ray treatment [6, 10, 11, 15]. The mutant was officially released in Czechoslovakia in 1965 as cv. Diamant, and this allele has been used for the successful release of more than 150 new malting barley cultivars in Europe [6, 15]. The sdw1.d allele has gained great acceptance in malting barley breeding programs in Europe, Canada, USA, and Australia, while the sdw1.a allele has been limited to feed barley varieties [14]. The fourth allele, sdw1.e (mutant line ‘Ris∅ no. 9265’) was found in the M2 generation of cv. Bomi after treatment with partially moderated fission neutrons in a reactor [10]. However, there are no reports of the use of this allele in variety development [6].
The sdw1 locus is located on chromosome 3HL, but more distal from the centromere than uzu1 [16]. Comparative genomic analysis revealed that the sdw1 gene in barley is located in the syntenic region of the rice green revolution semi-dwarf gene sd1, encoding a gibberellin 20-oxidase enzyme [13]. However, it is not clear what the gene structure changes resulted in different functional alleles. The objectives of this study were to (i) confirm gibberellin 20-oxidase as the functional gene, (ii) provide a detailed molecular characterization of different alleles at the sdw1 locus, (iii) understand how gene expression at the locus is regulated, and (iv) develop an allele-specific diagnostic marker for barley breeding programs.
Results
Cloning the HvGA20ox2 gene from barley genomic DNA
A fragment of 4831 bp was isolated from the tall barley varieties AC Metcalfe, Hamelin, and Valticky following PCR amplification of genomic DNA (Additional file 1: Figure S1). Based on FGENESH gene annotation, the barley HvGA20ox2 gene (3486 bp) contains three exons and two introns, with 1030 bp for exon 1, 325 bp for exon 2, 490 bp for exon 3, 173 bp for intron 1, and 1468 bp for intron 2. The coding sequence is 1242 bp in length, with a 371 bp 5’ untranslated region in exon 1 and a 232 bp 3’ untranslated region in exon 3 (Additional file 1: Figure S1). In addition, the isolated 4831 bp barley DNA fragment contains a 974-bp 5' upstream sequence and a 371-bp 3' downstream sequence of the HvGA20x2 gene.
The putative protein of the HvGA20ox2 gene has 414 amino acids. The predicted protein contains a conserved domain of the 2OG-Fe(II) oxygenase superfamily, non-haem dioxygenase in morphine synthesis, and gibberellin 20-oxidase (Fig. 1a, b).

Allelic variations of HvGA20ox2 gene in barley. a: structure of HvGA20ox2 gene; b: conserved domain of HvGA20ox2 protein; c: sdw1.d allele; d: verification of deletion in sdw1.d allele in a DH pupation of Baudin/AC Metcalfe; e: sdw1.c allele mutation
The barley HvGA20ox2 orthologous genes were identified by BLASTP in rice (sd1 OsGA20ox2, AAL87949), wheat (CDM85079.1), Aegilops (EMT17460), Brachypodium (XP003567337), maize (XP008654721), sorghum (XP002456751), Setaria italica (XP004970813) and Arabidopsis (GA20ox1 gene, NP194272). The amino acid sequence identity of the predicted HvGA20ox2 proteins in other grass species and Arabidopsis is listed in Additional file 2: Table S1. The predicted protein of the barley HvGA20ox2 gene was more similar to wheat and Aegilops (94.0 and 95.4% identity, respectively) than maize and Brachypodium (74.4 and 74.7% identity, respectively). As expected, the lowest level of identity was found for Arabidopsis (46.9%).
The barley HvGA20ox1 (AAT49058) and HvGA20ox3 (AAT49059) genes, previously isolated, are also involved in GA (gibberellic acid) biosynthesis [17]. The predicted protein of HvGA20ox2 only shares 50.6 and 48.5% of sequence identity with HvGA20ox1 (AAT49058) and HvGA20ox3 (AAT49059), respectively. Phylogenetic trees of the predicted proteins of barley HvGA20ox2 and the orthologous proteins HvGA20ox1 and HvGA20ox3 were constructed (Fig. 2).

Phylogenetic trees of the predicted proteins of HvGA20ox2 gene including the ortholog proteins
Allelic variation of HvGA20ox2 in semi-dwarf barley
The nucleotide sequences of the HvGA20ox2 gene from the three tall barley varieties (AC Metcalfe, Hamelin and Valticky) were identical. DNA sequences of the HvGA20ox2 gene were isolated from Baudin and Diamant, two semi-dwarf barley varieties known to have the sdw1.d allele. No nucleotide differences were detected between Baudin and Diamant. A comparison between the three tall barley varieties and sdw1.d allele semi-dwarf barley (Baudin and Diamant) identified a 7-bp (GACTCCC) deletion in the coding region of exon 1, from position 473 to 479, in the sdw1.d allele (Fig. 1c). In addition, the previously detected A/G substitution was also confirmed in this study [13]. The deletion in the sdw1.d allele was predicted to cause coding frame shifts and premature translation termination. Sequence analysis showed that there are ten internal ‘ATG’ start sites in the sdw1.d coding sequence. Among them, three ‘ATG’ sites located in position 1026–1028 (exon 1),1232–1234 (exon 2) and 1334–1336 (exon 2) could translate to a truncated protein with a conserved domain of the 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily (Fig 1).
Another important semi-dwarf allele of the HvGA20ox2 gene is sdw1.c (originally named denso). The DNA sequence of HvGA20ox2 was determined from a semi-dwarf barley Deba Abed. This allele did not have the sdw1.d (Diamant, also called as denso in literature) allele deletion. Five different sequence variations were identified by comparing the HvGA20ox2 gene sequence of Deba Abed with the tall barley cultivars (AC Metcalfe, Hamelin and Valticky). The deletion of a single “A” and a “GTTA” insertion were located in the untranslated region of exon 1 in positions 42 and 64, respectively. The 4-bp insertion in the sdw1.c allele was further confirmed by using barley varieties with known genotype (Fig. 3). In addition, two synonymous mutations were also detected at positions 659 (coding sequence of exon 1, G/A transition) and 3161 (coding sequence of exon 3, C/G transversion). An A/C transversion was also detected at position 3321 in the 3’ UTR region (Fig. 1e). However, none of the synonymous mutations in coding region and the transversion in 3’ UTR is expected to explain the dwarf phenotype.

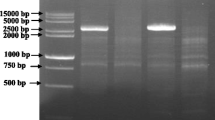
The 4-bp insertion in the sdw1.c allele amplified by the marker MC40861P in HvGA20ox2 gene. Lanes 4, 7 and 9 represent the sdw1.a allele. DNA templates (from left to right): 1. AC Metcalfe, 2. Baudin, 3. Deba Abed, 4. Jotun, 5. Hamelin, 6. Triumph, 7. Yerong, 8. Diamont, 9. Jotun, 10. Maris Mink
In contrast to sdw1.c and sdw1.d alleles, all primer combinations of the whole gene in Additional file 2: Table S2 failed to amplify any fragment from the sdw1.a mutants. PCR amplification analyses spanning the HvGA20ox2 gene locus and the neighboring genes identified a possible deletion of the whole HvGA20ox2 gene in sdw1.a varieties (data not shown).
Mapping the HvGA20ox2 gene in the Baudin/AC Metcalfe population
Two molecular linkage maps have been constructed for the Baudin/AC Metcalfe DH (double haploid) population. The first map was constructed with 178 DH lines and 234 SSR and AFLP markers [18]. The second map has 12,998 SNP tags anchored to seven chromosomes, spanning a cumulative 967.6 cM genetic distance [19]. In both maps the 7-bp indel polymorphism mapped to the expected location on chromosome 3H (data not shown).
Plant heights from three different field trials were used for QTL analysis. The average height of sdw1.d allelic plants was 16 to 19 cm shorter than the wild type plants in all trials (Additional file 1: Figure S2). However, large variation in plant height was observed within an allelic class (Additional file 1: Figure S2). A major QTL was identified for plant height and explained 37.2–44.5% of the plant height variation (Additional file 2: Table S3). The QTL peak co-located with the HvGA20ox2 gene-specific marker (Additional file 1: Figure S3).
Association analysis of the gene-specific marker in a natural population
One hundred and ninety-seven barley varieties, breeding lines and landraces were collected from Australia, Africa, China, European, North and South America and their plant heights varied from 50 to 105 cm. Of those, 28 accessions had the 7-bp deletion, three accessions had the 4-bp insertion while two did not yield an amplification product (Table 1). The 7-bp deletion points to the sdw1.d allele, the 4-bp insertion points to the sdw1.c allele and the lack of amplification points to the sdw1.a allele. Twenty-one barley accessions with the sdw1.d allele belong to the obvious dwarf types, with heights varying from 50 to 70 cm. Seven lines with the sdw1.d allele have a medium stature, from 75 to 80 cm. One sdw1.c allelic barley variety Tx9425 is the dwarf type. The two sdw1.a allelic barley varieties Yerong and Yan90260 are of the dwarf type. The sdw1.a and sdw1.d alleles explained 29% of plant height variation in the 197 barley varieties (P < 0.0001). We only detected the sdw1.a and sdw1.d alleles in modern barley varieties. The results provide further support for GA20 oxidase 2 (HvGA20ox2) as the functional gene for the sdw1 locus. We also observed that 52 barley varieties/lines displayed the short stature without the sdw1.a, sdw1.c and sdw1.d alleles in this population.
Transcription levels of genes encoding the final steps of GA biosynthesis
Our previous result demonstrated that the mutations in sdw1.d and sdw1.a reduced the gene expression of HvGA20ox2 [20]. In this study, we also measured the expression of the other two homologous genes HvGA20ox1 and HvGA20ox3 (Fig. 4a,c). It is surprised that the expression level of HvGA20ox1 was 1.7 times higher in Baudin (sdw1.d) and 4.7 times higher in Jotun (sdw1.a) while HvGA20ox3 showed three times higher in Baudin and 1.4 times higher in Jotun. The result suggests that partial or total loss of function of HvGA20ox2 can be compensated by other GA20 oxidases, especially HvGA20ox1.

Relative gene expression levels of HvGA20ox1and HvGA20ox3. a: transcription level of HvGA20ox1 at stem elongation stage in AC Metcalfe (wild type), Baudin (sdw1.d allele) and Joutn (sdw1.a allele); b: bulk-segregating analysis of HvGA20ox1 gene expression at tillering stage in Baudin/AC Metcalfe DH population, each bulk contained 20 DH lines with different alleles of the HvGA20ox2 gene; c: transcription level of HvGA20ox3 at stem elongation stage in AC Metcalfe (wild type), Baudin (sdw1.d allele) and Joutn (sdw1.a allele)
To further confirm if the increased expression of HvGA20ox1 was due to partial loss of function of HvGA20ox2, we conducted a bulked segregant analysis of gene expression in the Baudin (sdw1.d)/AC Metcalfe (tall) DH population. The expression level of the sdw1.d bulk matched with the sdw1.d parent Baudin, with higher expression and reversed trend observed in the tall bulk and AC Metcalfe (tall parent) (Fig 4b). From those results we conclude that partial loss (sdw1.d) or total loss (sdw1.a) of HvGA20ox2 may be compensated by increased expression of HvGA20ox1.
Discussion
Modification of the gibberellin biosynthetic and signal transduction pathways was a crucial step in crop breeding, as it conferred the agronomically important semi-dwarf phenotype [21]. The rice green revolution gene sd1 was the result of reduced function of GA 20-oxidase-2 [3]. The GA 20-oxidases are involved in the later steps of GA biosynthesis, in which GA53 is converted into GA44 [17]. It is now clear that reduced function of the GA 20-oxidase gene leads to reduction in plant height in rice. A previous study has demonstrated that the sdw1 gene may be orthologous to the rice sd1 gene [13]. However, it is not clear how the gene structure changes resulted in dfiierent functional alleles. In this study, we characterized a full-length copy and alleles of the barley HvGA20ox2 gene, which has a conserved gene structure when compared to the rice sd1 gene. Sequence similarity analysis showed that the predicted protein of the barley HvGA20ox2 gene shared 83.1% of identity to its rice ortholog.
Four alleles have been reported at the sdw1 locus. In this study, we characterized the HvGA20ox2 gene from three independent mutants. The sdw1.a allele might be the result of a total deletion of the HvGA20ox2 gene. Nearly no expression of HvGA20ox2 was detected for the sdw1.a mutant (Jotun) previously [20], which was consistent with a total deletion of the HvGA20ox2 gene, as our study suggests. A recent study demonstrated that sdw1.e (mutant line ‘Ris∅ no. 9265’) also resulted from a total deletion of the HvGA20ox2 [22]. The sdw1.c allele has a 1-bp deletion and a 4-bp “GTTA” insertion in the untranslated region of exon1, respectively. The sdw1.d (Diamant) allele is caused by a 7-bp deletion in exon1, which resulted in coding frame shifts and premature translation termination. As there is an internal ATG, the sdw1.d (Diamant) allele may lead to a truncated protein with a conserved domain of the 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily. Thus, the sdw1.d (Diamant) allele still maintains partial function of GA 20-oxidase. Sequencing of different alleles at the sdw1 locus points to HvGA20ox2 as the functional gene responsible for the phenotype.
Based on our sequencing results, we designed an allele-specific marker. As expected, the allele-specific marker co-segregated with a major QTL controlling plant height in the DH population of Baudin/AC Metcalfe. The gene-specific marker was further tested in a natural population. We found the sdw1.a and sdw1.d alleles only in modern barley varieties and associated with plant height. These results provide further support for HvGA20ox2 as the functional gene of the sdw1 locus. However, the molecular marker for the 4 bp insertion in the sdw1.c allele seems not associated with plant height in the natural population. We speculate that the 1 bp deletion may be more important for the gene function in the sdw1.c allele as the sdw1.d allele.
Until now, no malting barley variety has been developed from the sdw1.a allele. Bioactive gibberellins are not only essential regulators for barley growth and development, but are also essential for malting and brewing [23]. It is expected that the deletion of the HvGA20ox2 gene in sdw1.a allele would result in reduced GA biosynthesis during the malting process. This would explain why the sdw1.a allele has been used exclusively in feed barley.
A recent study in Arabidopsis thaliana reported 21 independent loss-of-function alleles at GA locus 5 (GA5), which encodes gibberellin 20-oxidase 1 (GA20ox1), causing semi-dwarfness [24]. These results suggest that GA 20-oxidase might be a hot spot for phenotypic variation in crop and other plant species. Further research is required to establish whether there is further allelic variation in HvGA20ox2 in barley.
The predicted protein of the barley HvGA20ox2 gene shared high identity with the Aegilops and wheat orthologs (Fig. 2), which raises the question why no such semi-dwarf mutants have been identified in these species thus far. Such mutants have already demonstrated great potential to increase yield in rice and barley, and thus it seems worthwhile creating similar mutants in wheat as an alternative source of dwarfing genes. Our results further demonstrate that GA20 oxidase homologs can functionally compensate for each other (Fig. 4b). This means that to achieve a similar feat in wheat, GA20 oxidase expression in all three genomes would have to be modified simultaneously. Advances in sequencing and gene editing technologies may provide an efficient approach to identifying or producing such mutants in wheat.
Previously, a SNP in intron 2 was detected between semi-dwarf barley variety Baudin and tall variety AC Metcalfe [13]. The SNP marker was mapped to chromosome 3H in the double haploid population of Baudin/AC Metcalfe, while co-segregating with plant height [13]. However, this SNP is not unique for the sdw1.d allele. In contrast, the allele-specific marker in this study can be used as a diagnostic test for the sdw1.a, sdw1.d and wild-type alleles.
The sdw1 alleles explained part of the height variation in both the DH population and the test barley varieties. Some barley varieties without the sdw1.a and sdw1.d alleles also displayed short stature. These results indicated that some novel dwarfing genes have already used to breed barley varieties [6, 9, 25–29]. We also observed the plant height variation within allele classes was much greater than the variation between sdw1.d allele class and wild type class. This indicated that some novel dwarfing genes also responsible for the height variation between Baudin and AC Metcalfe [6, 9, 25–29].
Methods
Genetic materials and agronomic traits
The medium tall barley varieties used in this study included AC Metcalfe, Valticky (parent of Diamant), and Hamelin. The semi-dwarf barley varieties Diamant and Baudin represent the sdw1.d allele. The sdw1.d allele in Baudin was from Triumph, which derived its sdw1.d gene from Diamant. The barley variety Deba Abed represents the sdw1.c (denso) allele. Jotun is the sdw1.a mutant. Yerong is a semi-dwarfing dual-purpose (feed and graze) barley variety carrying sdw1.a gene [30].
A doubled haploid population comprising 178 lines was generated via anther culture from the F1 progeny of a Baudin/AC Metcalfe cross. The 197 barley varieties and lines used in this study were collected from Australia, Africa, Europe, North and South America, and are listed in Table 1.
The mapping population (178 DH lines) with its parents and the 197 barley accessions were planted at three sites in Western Australia. The field trial sites were located in the high rainfall agricultural zone, in order to achieve the maximum growing potential for the semi-dwarf genotypes. The DH lines and parents were planted in 1 × 5 m plots and the same randomized design was used at each site for convenience. Parental and local barley varieties were used as grid controls for spatial analysis.
Cloning of HvGA20ox2 gene from barley varieties
Polymerase chain reaction (PCR) primers were designed from the cloned fragments of the HvGA20ox2 gene [13] and barley genome sequencing information (Additional file 2: Table S2). The relative positions of each primer to the HvGA20ox2 gene are shown in Additional file 1: Figure S1. All primers were synthesized by Gene Works Pty. Ltd. (Australia). The PCR reactions consisted of 50 ng genomic DNA as template, 0.1 μM of each primer, in a final volume of 10 μl containing 1 × PCR buffer, 1.5 mM MgCl2, 0.2 mM dNTP, and 0.5 U Taq polymerase (Bioline, Australia). The PCR reactions were performed using the following program: denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, annealing for 45 s and extension at 72 °C for 1 min, and a final extension at 72 °C for 5 min. The optimal annealing temperature of each pair of primer combination was determined by gradient PCR. The PCR products were cloned into pGEM-T Easy Vector (Promega), and at least two independent clones from each PCR product were sequenced using an automated sequencing system (ABI 377, Applied Biosystems).
Sequence assembly and alignment
The target sequences of each variety were assembled by the SeqMan program (DNAStar). Clustal X2 was used for multiple sequence alignment. The exon and intron, and protein sequences of the HvGA20ox2 gene from each variety were identified by using BLASTN, TBLASTN, and online gene prediction software FGENESH (http://linux1.softberry.com/berry.phtml?topic=fgenesh&group=programs&subgroup=gfind). The orthologs of the barley HvGA20x2 gene from other grass species and Arabidopsis were confirmed by BLASTP. The identity of the deduced amino acid of the HvGA20x2 gene among the orthologs was analyzed by DNAStar. Phylogenetic trees of the predicted proteins of the barley HvGA20ox2 gene, including the orthologous proteins HvGA20ox1 and HvGA20ox3 was constructed using MEGA 6.0 by maximum likelihood approach, and the confidence of the nodes was evaluated using 1000 bootstrap replications.
Real-time quantitative RT-PCR
RNA was extracted from the stems at tillering or stem elongation stage using a Spin Column Plant total RNA Purification Kit(Sanggon Biotech (Shanghai) Co., Ltd. cDNA was prepared from 1 μg RNA using AMV First Strand cDNA Synthesis Kit(Sanggon Biotech (Shanghai) Co., Ltd). qPCR reactions were performed using SYBR Green (SG Fast qPCR Master Mix(High Rox), BBI) and the Applied Biosystems Stepone plus Real-time PCR System. The Real-time PCR assays were performed in triplicate for each cDNA sample. To determine transcription levels of barley HvGA20ox2 and genes encoding the final steps of GA biosynthesis, HvACTIN and HvGAPDH were employed as reference genes for barley. The oligonuleotide sequences used for quantitative RT-PCR are listed in Additional file 2: Table S4.
To determine if other genes are regulated by HvGA20ox2, 20 doubled haploid lines from the Baudin/AC Metcalfe population were selected based on the genotype of the HvGA20ox2 gene to construct two pools (sdw1.d and wild type) for measurement of the expression of other genes in the GA biosynthesis pathway. Three biological repeats were used for RNA extraction.
Verification of the denso allele in a DH population
Presence of the sdw1.d allele was verified in the DH population of Baudin/AC Metcalfe and barley cultivars. Genomic DNA was extracted from young leaves using the standard CTAB protocol. DNA samples were quantified using the Nanodrop equipment and adjusted to a final concentration of 50 ng/μL for PCR. Primers used are listed in Additional file 2: Table S1. PCR amplification conditions were as described above. The PCR products were separated in 6% PAGE gels.
QTL analysis for plant height
The software package MapQTL 5.0 was used to conduct QTL analysis for plant height after import of the files for genotypes, phenotypes and genetic maps. Interval analysis was first performed to estimate the closest markers associated with plant height, followed by multiple QTL model (MQM) analysis. LOD threshold values applied to declare the presence of a QTL were estimated by performing whole-genome wide permutation tests using 10,000 permutations. The QTL map was then generated using Mapchart 2.2.
Conclusions
Our research provided further evidence that the gibberellin 20-oxidase gene (HvGA20ox2) is the functional gene for the barley sdw1 mutants. The sdw1.d allele from Diamant is due to a 7-bp deletion in exon 1, while the sdw1.c allele from Abed Denso has 1-bp deletion and a 4-bp insertion in the 5’ untranslated region. The sdw1.a allele from Jotun resulted from a total deletion of the HvGA20ox2 gene. Partial or total loss of function of the HvGA20ox2 gene could be compensated by enhanced expression of its homolog HvGA20ox1 and HvGA20ox3. A diagnostic molecular marker was developed to differentiate between the wild-type, sdw1.d and sdw1.a alleles and another molecular marker for differentiation of sdw1.c and sdw1.a. Further research is required to establish whether the truncated protein could maintain partial function and whether there is further allelic variation in HvGA20ox2 in barley.
Abbreviations
- AFLP:
-
Amplified restriction fragment polymorphism
- cM:
-
Centimorgan
- DH:
-
Double haploid
- GA:
-
Gibberellic acid
- PCR:
-
Polymerase chain reaction
- QTL:
-
Quantitative trait loci
- Rht :
-
Reduced height
- sd1 :
-
Semidwarf-1
- sdw1 :
-
Semi-dwarf 1
- SNP:
-
Single nucleotide polymorphism
- SSR:
-
Simple sequence repeats
References
Milach S, Federizzi L. Dwarfing genes in plant improvement. Adv Agron. 2001;73(1):35–63.
Peng J, Richards DE, Hartley NM, Murphy GP, Devos KM, Flintham JE, Beales J, Fish LJ, Worland AJ, Pelica F. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature. 1999;400(6741):256–61.
Monna L, Kitazawa N, Yoshino R, Suzuki J, Masuda H, Maehara Y, Tanji M, Sato M, Nasu S, Minobe Y. Positional cloning of rice semidwarfing gene, sd-1: rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002;9(1):11–7.
Zhang J, Zhang W. Tracing sources of dwarfing genes in barley breeding in China. Euphytica. 2003;131(3):285–93.
Kuczynska A, Surma M, Adamski T, Mikolajczak K, Krystkowiak K, Ogrodowicz P. Effects of the semi-dwarfing sdw1/denso gene in barley. J Appl Genet. 2013;54:381–90.
Dockter C, Hansson M. Improving barley culm robustness for secured crop yield in a changing climate. J Exp Bot. 2015;66(12):3499–509.
Forster BP. Mutation genetics of salt tolerance in barley: an assessment of Golden Promise and other semi-dwarf mutants. Euphytica. 2001;120(3):317–28.
Zhang J. Inheritance of agronomic traits from the Chinese barley dwarfing gene donors ‘Xiaoshan Lixiahuang’and ‘Cangzhou Luodama’. Plant Breed. 2000;119(6):523–4.
Chono M, Honda I, Zeniya H, Yoneyama K, Saisho D, Takeda K, Takatsuto S, Hoshino T, Watanabe Y. A semidwarf phenotype of barley uzu results from a nucleotide substitution in the gene encoding a putative brassinosteroid receptor. Plant Physiol. 2003;133(3):1209–19.
Franckowiak JD, Lundqvist U. Descriptions of barley genetic stocks for 2012. Barley Genet Newsl. 2012;42:36–792.
Haahr V, Von Wettstein D. Studies of an induced, high-yielding dwarf-mutant of spring barley, Proceedings of the 3rd International Barley Genetics Symposium: 1975; Garching. München: Verlag Karl Thiemig; 1976. p. 215–8.
Mickelson HR, Rasmusson DC. Genes for short stature in barley. Crop Sci. 1994;34(5):1180–3.
Jia Q, Zhang J, Westcott S, Zhang XQ, Bellgard M, Lance R, Li C. GA-20 oxidase as a candidate for the semidwarf gene sdw1/denso in barley. Funct Integr Genomic. 2009;9(2):255–62.
Hellewell KB, Rasmusson DC, Gallo-Meagher M. Enhancing yield of semidwarf barley. Crop Sci. 2000;40(2):352–8.
Grausgruber H, Bointner H, Tumpold R, Ruckenbauer P, Fischbeck G. Genetic improvement of agronomic and qualitative traits of spring barley. Plant Breed. 2002;121(5):411–6.
Barua U, Chalmers K, Thomas W, Hackett C, Lea V, Jack P, Forster B, Waugh R, Powell W. Molecular mapping of genes determining height, time to heading, and growth habit in barley (Hordeum vulgare). Genome. 1993;36(6):1080–7.
Spielmeyer W, Ellis M, Robertson M, Ali S, Lenton JR, Chandler PM. Isolation of gibberellin metabolic pathway genes from barley and comparative mapping in barley, wheat and rice. Theor Appl Genet. 2004;109(4):847–55.
Cakir M, Gupta S, Li C, Hayden M, Mather DE, Ablett GA, Platz GJ, Broughton S, Chalmers KJ, Loughman R. Genetic mapping and QTL analysis of disease resistance traits in the barley population Baudin × AC Metcalfe. Crop Pasture Sci. 2011;62(2):152–61.
Zhou G, Zhang Q, Zhang X-q, Tan C, Li C. Construction of high-density genetic map in barley through restriction-site associated DNA sequencing. PLoS One. 2015;10(7):e0133161.
Jia Q, Zhang X-Q, Westcott S, Broughton S, Cakir M, Yang J, Lance R, Li C. Expression level of a gibberellin 20-oxidase gene is associated with multiple agronomic and quality traits in barley. Theor Appl Genet. 2011;122(8):1451–60.
Peter H, Stephen GT. Gibberellin biosynthesis and its regulation. Biochem J. 2012;444(1):11–25.
Jia QJ, Li CD, Shang Y, Zhu JH, Hua W, Wang J, Yang JM, Zhang GP. Molecular characterization and functional analysis of barley semi-dwarf mutant Riso no. 9265. BMC Genomics. 2015;16:927.
Yadav S, Luthra Y, Sood D, Aggarwal N. Gibberellic acid (GA3) induced changes in proanthocyanidins and malt quality of two and six-row husked barleys. Plant Food Hum Nutr. 2000;55(2):87–96.
Barboza L, Effgen S, Alonso-Blanco C, Kooke R, Keurentjes JJ, Koornneef M, Alcazar R. Arabidopsis semidwarfs evolved from independent mutations in GA20ox1, ortholog to green revolution dwarf alleles in rice and barley. Proc Natl Acad Sci U S A. 2013;110(39):15818–23.
Yu GT, Horsley RD, Zhang B, Franckowiak JD. A new semi-dwarfing gene identified by molecular mapping of quantitative trait loci in barley. Theor Appl Genet. 2010;120(4):853–61.
Ren X, Sun D, Guan W, Sun G, Li C. Inheritance and identification of molecular markers associated with a novel dwarfing gene in barley. BMC Genet. 2010;11(4):1–7.
Vu GT, Wicker T, Buchmann JP, Chandler PM, Matsumoto T, Graner A, Stein N. Fine mapping and syntenic integration of the semi-dwarfing gene sdw3 of barley. Funct Inter Genomics. 2010;10(4):509–21.
Wang J, Yang J, Jia Q, Zhu J, Shang Y, Hua W, Zhou M. A new QTL for plant height in barley (Hordeum vulgare L.) showing no negative effects on grain yield. PLoS One. 2014;9(2):e90144.
Li H, Chen G, Yan W. Molecular characterization of barley 3H semi-dwarf genes. PLoS One. 2015;10(3), e0120558.
Read BJ, Macdonald AF. Hordeum vulgare (barley) cv. Yerong. Anim Prod Sci. 1991;31(6):866.
Funding
This work was carried out with the financial support from the Australian Grain Research and Development Corporation (to CL) and the National Natural Science Foundation of China (No. 31201212) (to YX), National Key Research and Development Program (2016YFD0102101) and the Talent Youth Foundation of Hubei Province (to YX).
Availability of data and materials
The data supporting the results of this article are included within the article and its additional files. Genetic materials are available by contacting with the corresponding authors.
Authors contribution
YX: conduct gene sequencing, developing molecular marker, analyze data and write the manuscript; QJ: identify the candidate gene and quantitative PCR; GZ: QTL analysis and gene mapping; XQZ: molecular marker and field phenotype; TA: genetic material collection and population development; SB: population development; ZGY: field phenotype; WZ: design the experiment; CL: develop project concept, design the experiments, write and finalize the paper. All the authors have read through the manuscript and agree to the submission of the final version.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Author information
Authors and Affiliations
Corresponding authors
Additional files
Additional file 1: Figure S1.
Structure of barley HvGA20ox2 gene and the relative position of the primers used in this study. Figure S2. Plant height (cm) variation in Baudin/AC Metcalfe DH population from three independent field trials (SP-Ht: South Perth plant height; KD Ht: Plant height in Kendup trials. Figure S3. A major QTL for plant height co-segregated with HvGA20ox2 on chromosome 3H. The genetic map is based on Zhou et al. (2015). (ZIP 118 kb)
Additional file 2: Table S1.
Identity of the deduced amino acid sequence of the HvGA20ox2 gene with selected orthologs. Table S2. Primers used to amplify the HvGA20ox2 gene and inspect sdw1 allelic variations. Table S3. Barley varieties used in this study and their genotype at the sdw1 gene locus. Table S4. The oligonuleotide sequences used for quantitative RT-PCR for different genes. (DOCX 28 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Xu, Y., Jia, Q., Zhou, G. et al. Characterization of the sdw1 semi-dwarf gene in barley. BMC Plant Biol 17, 11 (2017). https://doi.org/10.1186/s12870-016-0964-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-016-0964-4