
Abstract
Extracellular vesicles (EVs) are a heterogeneous group of lipid membrane-enclosed compartments that contain different biomolecules and are released by almost all living cells, including fungal genera. Fungal EVs contain multiple bioactive components that perform various biological functions, such as stimulation of the host immune system, transport of virulence factors, induction of biofilm formation, and mediation of host–pathogen interactions. In this review, we summarize the current knowledge on EVs of human pathogenic fungi, mainly focusing on their biogenesis, composition, and biological effects. We also discuss the potential markers and therapeutic applications of fungal EVs.
Similar content being viewed by others


Introduction
Extracellular vesicles are phospholipid bilayer nanostructures derived from living cells in the extracellular milieu [1]. These membrane particles are secreted by cells of all three domains: prokaryotes, eukaryotes, and archaea [2]. Their diameter ranges from 20 nm to approximately 1 μm. Based on their size, the newest grouping of EVs separates them into two major groups: small EVs or endosomal-origin EVs named “exosomes” (up to 150 nm) and medium/large EVs or microvesicles (> 150 nm) [3]. Although the characteristics and functional roles of EVs have been extensively studied in mammalian systems [4], our understanding of EVs in fungi remains relatively limited. EVs derived from fungi were first identified in Cryptococcus neoformans during a study of the trans-cell wall transport mechanisms of capsular polysaccharides [5]. Since then, EVs have been reported in various other virulent and non-virulent fungal species [6]. Similar to the mammalian, bacterial, and plant EVs, fungal EVs also contain proteins, carbohydrates, pigments, nucleic acids, toxins, and other bioactive molecules in their intercellular matrices or on their surface. These cargoes are transported to the extracellular environment for various biological functions [7,8,9,10,11,12,13,14]. Once EVs are released into the extracellular space, they migrate long distances until they encounter another cell or environment to connect with [15]. Several EV components are conserved among various fungal species and strains, while some are species-specific [8, 16]. Fungal EVs with immunogenic properties exert immunotherapeutic effects, and hence, can be used as potent tools for various therapeutic applications [17,18,19].
In this review, we summarize the biogenesis, composition, biological effects, and potential therapeutic applications of EVs of human pathogenic fungi. We also discuss the roles of EVs of non-pathogenic fungi in some sections for comparison. Finally, we outline the current challenges in fungal EV research.
Biogenesis of fungal EVs
Most studies on EV biogenesis mechanisms have focused on mammals; however the biogenesis mechanisms of fungal EVs remain unclear [6]. Extreme variations in the morphology, size, cargo, and composition of EVs suggest the presence of different EV biogenesis pathways in cells [1, 20, 21]. Several cytoplasmic proteins without any secretory signals have been discovered in fungal EVs, suggesting that EVs may be derived from cytoplasmic fractions [22]. Multivesicular bodies and membrane budding also play roles in EV biogenesis process [9, 20, 23, 24]. EVs are also produced via an invagination process (inverted macropinocytosis) and the vesicles are subsequently released into the periplasmic vacuum [22]. Moreover, EVs may be derived from vesicle-containing vacuoles that are fused with the plasma membrane subsequently [21].
Characterization of EVs formed by mutants with defects in several pathways is the key method used to study vesicle biogenesis in fungi. It’s been demonstrated that both Golgi apparatus secretory pathways and endosomal sorting complex required for transport (ESCRT) machinery are involved in fungal EV biogenesis, as well as a variety of other biogenesis regulators. Figure 1 shows the detailed biogenesis mechanisms of fungal EVs.
Golgi apparatus secretory pathways
It’s been reported that Golgi apparatus secretory regulators are associated with fungal EVs biogenesis. For instance, in Saccharomyces cerevisiae, a mutation in the SEC4 gene responsible for encoding exocytic Rab GTPase (required for post-Golgi secretory vesicle formation) alters the production of vesicles [20]. In C. neoformans, Sec6 is implicated in the exocytosis of post-Golgi secretory vesicles to the plasma membrane. Suppression of SEC6 using a small interfering RNA decreases the secretion of virulence-related molecules into EVs [25]. Sec1 is associated with the integration of Golgi-derived EVs, and the cell membrane participates in EV formation as well. However, deletion of SEC1 does not influence the secretion of EVs [20].
Golgi reassembly and stacking protein (GRASP) and autophagy-related protein 7 (Atg7) are related to EV secretion, and mutant cells lacking GRASP and Atg7 exhibit altered EV size [26]. GRASP is involved in EV-mediated mRNA transport and is a significant modulator of virulence in C. neoformans [27]. Mutant cells lacking acyl coenzyme A-binding protein (ACBP), a GRASP expression needed for conventional secretion in Dictyostelium discoideum [28] and S. cerevisiae [29] exhibit a reduction in EV components compared to the wild-type cells [20].
Although alterations in these conventional secretory genes affect EV formation, they do not completely inhibit the development of EVs, suggesting the involvement of other functional pathways in the generation and release of EVs.
ESCRT machinery
Apart from Golgi apparatus secretory pathways, ESCRT machinery is also demonstrated to be involved in the biogenesis of fungal EVs. ESCRT is associated with the development and functions of multivesicular bodies (MVBs). For example, Snf7 and Vps23 influence EV proteins in S. cerevisiae [20]. In Candida albicans, biofilm EVs contain ESCRT sub-units, Hse1 and Vps27, and mutations in different ESCRT sub-unit homologs result in reduced EV formation compared to that in wild-type strains [30]. Moreover, EVs from ESCRT-knockout yeast strains exhibit enrichment of cell wall-rebuilding enzymes, particularly chitin synthases Chs1 and Chs3, and the glucan synthase sub-unit, Fks1 [10]. Absence of Vps27 in the ESCRT complex results in the accumulation of MVBs and discharge of larger EVs in C. neoformans [31].
Other biogenesis regulators
In addition to the ESCRT apparatus, several regulators may affect EV production and cargo composition. Deletion of the APT1 gene encoding lipid flippase alters the size of C. neoformans EVs and the EV-based export of soluble glucuronoxylomannan (GXM) [32, 33]. In Cryptococcus gattii, removal of AIM25, which encodes the enzyme lipid scramblase, results in the formation of larger EVs with altered RNA content [34]. In Candida albicans, mutant species deprived of genes encoding phosphatidylserine synthase/decarboxylase (CHO1/(PSD1 and PSD2) influence the morphology, immunogenicity, and composition of EVs, which is highly suggestive of an association between the metabolism of lipids and constituents of EVs [35]. Other regulators also play a role in the release of EVs. For example, deletion of the chitin synthetase (CHS) gene in C. neoformans inhibits EVs release [36]. High EV production is observed in a C. neoformans strain lacking Cln1, a protein associated with cell cycle progression [37]. In addition, the removal of CAP10, a putative xylosyltransferase, considerably reduces the size of EVs [38]. Intracellular vesicular clusters of S. cerevisiae promote the production and selection of proteins associated with EV secretion [39]. Moreover, reduced expression of cryptococcal microvesicle marker protein 14-3-3 [40] decreases the GXM and protein content in EVs and decreases acid phosphatase and laccase activities [41].
Furthermore, the environmental stimuli, host-posed conditions and metabolic mechanisms are thought to influence the release of EVs, as well as their size and composition [7, 22, 30, 42,43,44]. The attachment of protective or non-protective monoclonal antibodies (MAbs) targeting Hsp60 to Histoplasma capsulatum fungal cells has been shown to results in altered EV cargo and release [43, 44]. Moreover, treatment with caspofungin, an antifungal drug, enhances the release of EVs from S. cerevisiae [10]. Interestingly, EVs release can also be influenced by EV size. Only multiple vesicle-leaving events (in which a group of vesicles is released simultaneously) release EVs larger than 100 nm in C. neoformans, whereas smaller vesicles may be released in multiple or single vesicle-leaving events [9]. These studies highlight the complicated mechanisms related to the formation and secretion of EVs (Fig. 1).

Schematic diagram of fungal extracellular vesicle (EV) biogenesis. Fungal EV biogenesis is regulated by multiple secretory regulators, including Golgi apparatus secretory pathways, ESCRT machinery and other biogenesis regulators. EVs may also be derived from vesicle-containing vacuoles in C. neoformans [21]. Mutant cells lacking the Golgi reassembly and stacking protein (GRASP) and autophagy-related protein 7 (Atg7) exhibit altered EV size in C. neoformans [26]. Mutation in acyl coenzyme A-binding protein (ACBP) in D. discoideum [28] and S. cerevisiae [29] indicate a reduction in components of EVs. Mutation in SEC4 gene alter the production of vesicles in S. cerevisiae [20]. SEC6 gene suppression decreases the secretion of virulence-related molecules into EVs of C. neoformans [25]. ESCRT sub-units, Snf7 and Vps23, influence EV proteins in S. cerevisiae [20]. ESCRT sub-units, Hse1 and Vps27, influence biofilm EV formation in C. albicans [30]. Deletion of APT1 gene encoding lipid flippase influences the size of EVs and EV-based export of soluble GXM in C. neoformans [32, 33]. Attachment of monoclonal antibodies targeting Hsp60 to H. capsulatum fungal cells alters the EV cargo and release [43, 44]. EE-early endosome; LE-late endosome; Aph-autophagosome; MVB-multivesicular body
Cell wall-crossing mechanisms of fungal EVs
The intricate and vibrant cell wall surrounding the fungal cells is generally considered the ultimate barrier for the secretion of fungal EVs to reach the external environment [45]. Therefore, the ability of fungal EVs to cross the cell wall is a fundamental requirement. In encapsulated fungal species, EVs must also cross the capsules of fungal cells such as those related to the Cryptococcus group [5]. However, the cellular mechanisms underlying the secretion of fungal EVs across the cell wall and capsule have not yet been clearly elucidated. The cell wall is primarily composed of a polysaccharide matrix in which various proteins and lipid groups are incorporated [45]. Three nonexclusive hypotheses have been put forth to explain the cell wall crossing mechanism of fungal EVs. First, protein channels present in the periphery of the cells can guide the fungal EVs to the extracellular space. Second, once fungal EVs are released from the cell membrane, the turgor pressure exerted by the cells forces them through the pores in the cell wall. Finally, hydrolytic enzymes loosen the cell wall by digesting specific areas of the cell wall, which allows the passage of fungal EVs through the cell wall [46,47,48].
Data supporting these non-mutually exclusive hypotheses regarding EVs passage through the fungal cell wall are dispersed in the literature. The size of the pores in the cell wall differs by strain and may be adjusted under distinct conditions depending on the cell wall remodeling enzymes, culture growth phase, extracellular pH, and deposits, such as melanin. The diameter of the pores present in the cell wall of S. cerevisiae may increase to 400 nm in response to stress [49]. This alteration represents a possible channel for larger EVs to cross cell walls. However, decrease in cell wall pore size in cryptococci can contribute to the aggregation of vesicle-like structures “trapped vesicles” seen in the region between cell wall and plasma membrane [9, 50]. Recent research has suggested that fungal cell walls have viscoelastic properties that may allow the transportation of membranous particles. Amphotericin B-containing liposomes with diameters of 60–80 nm penetrated the cell walls of C. albicans and C. neoformans from the outer environment of the cell and approached the plasma membrane in their original form, although the estimated size of the pores present in the cell wall was notably low (approximately 5.8 nm) to permit their transport [48]. Cryo-SEM analysis showed that single and multiple vesicles interacted directly with the cell wall of C. neoformans without any apparent trans-cell wall channels or pores, encouraging their passage [9]. This is in contrast to the channels in fungi that steer vesicle secretions.
Cell wall remodeling may be an effective mechanism for vesicle release. Proteomics has shown that several enzymes involved in cell wall remodeling, such as glucanases, are found in EVs, facilitating their passage through the cell wall [10, 51, 52]. Additionally, EVs are produced by polymorphological nuclear granule cells. These EVs are bound to the cell wall of Aspergillus fumigatus and are also capable of entering the hyphae of fungal cells, which results in the modification of the morphology of the fungal cell walls [53]. These studies highlight the dynamic and flexible structure of the cell wall along with its viscoelastic characteristics, which provide innovative information about the movement of vesicles through the cell wall to approach the extracellular space [6].
Composition of fungal EVs
Several studies have shown that fungal EVs carry various bioactive substances. Using EVs as a medium, these substances are involved in multiple biological activities. Fungal EVs mainly contain the following substances:
Proteins
Fungal EVs cargo is composed of a combination of different proteins that may or may not contain secretory signals. Fungal EVs contain proteins with biological roles in the biogenesis of cell wall architecture, cell membrane, response to stress, virulence/pathogenesis, cell signaling, sugar metabolism, lipid metabolism, cell growth/division, and transportation. Most of these proteins are shared among EVs from different fungal species, including C. neoformans, C. albicans, Paracoccidioides braziliensis, H. capsulatum, and S. cerevisiae, and species-specific proteins have also been widely studied [8, 20, 54,55,56].
In C. neoformans, two previous proteomic analyses reported 92 and 202 proteins in the EVs extracts [9, 21]. Enzymes involved in capsule biosynthesis, such as UDP-glucose dehydrogenase and UDP-glucuronic acid decarboxylase, are specifically found in C. neoformans EVs [21]. Recently, the analysis of EVs protein cargo from three cryptococcal species revealed many membrane-bound protein families, including Tsh proteins bearing the SUR7/PalI motif. Proteins involved in various biological processes have been identified in the EVs cargo, including Mp88 and components of the Cda, Gox, and Ril protein families [57]. In a previous study, the ability of C. gattii EVs to trigger an increased intracellular proliferation rate was eliminated when pretreated with proteinase K, indicating that C. gattii EVs contain proteins related to pathogenicity [15]. EVs of H. capsulatum revealed the presence of proteins related to REDOX, including superoxide dismutase, thiol-specific antioxidants, and catalase B, which are involved in fungal defense. Various enzymes, such as glucanase and endochitinase, are also involved in the hydrolysis of cell wall constituents that facilitate the secretion of EVs. Proteins involved in the transport and fusion of vesicles, particularly those of the Rab family, have also been identified [55]. Proteomic analysis of P. braziliensis EVs revealed a varied and complex distribution of proteins with various biological functions [8]. Protein content was compared between the EVs of P. braziliensis and those of C. neoformans, S. cerevisiae, and H. capsulatum. Twenty-six proteins were found to be common among these four fungal species. Additionally, 72 proteins are common to P. braziliensis and at least two other species [8]. Proteomic analysis of EVs derived from A. fumigatus protoplasts revealed the presence of proteins related to lipid and sugar metabolism, cell wall biosynthesis, and pathogenic processes. In EVs from A. fumigatus protoplasts, proteins not expected to be present in the extracellular environment have been identified in EVs from A. fumigatus protoplasts [23]. This finding is in agreement with various studies on the protein composition of fungal EVs [21, 51, 54, 55].
In C. albicans, the first proteomic study of EVs reported 75 proteins that fell into nine functional classes, most of which were related to the cell wall [58]. A label-free quantitative assessment identified 47 proteins that were enriched in C. albicans EVs compared to those in planktonic cells. These proteins were refined to 22 possible eV protein markers in C. albicans, including claudin-like Sur7 family proteins (Sur7 and Evp1). Additionally, azole-resistant proteins, including Cdr1 and Cdr2, have been detected [59]. Another proteomic study comparing planktonic cells and biofilm EVs in C. albicans showed that 34% of the protein cargo was specific to biofilm EVs [30]. Analysis of EVs from non-albicans Candida species (NAC) revealed the presence of 42 extracellular and surface-connected proteins from Candida glabrata, 33 from Candida parapsilosis, and 34 from Candida tropicalis [60]. Of the EVs proteins identified in Candida species, many were also found in EVs from other fungi, including S. cerevisiae and other species that are pathogenic to humans [61]. Some proteins present in fungal EVs are assumed to be moonlighting proteins [60].
A quantitative large-scale proteomic study comparing the protein content of EVs and whole-cell lysates of Malassezia sympodialis revealed the presence of many proteins that were enriched in EVs, including two allergens (Mala s1 and Mala s7) and catalytic enzymes (helicases, ligase, histone, tRNA synthetase, ribonuclease, and deacetylase) [62]. In Alternaria infectoria, a phytopathogen as well as an opportunistic pathogen in humans, proteomic analysis of EVs cargo revealed the existence of proteins related to cell metabolism and pathogenicity. Of the class of carbohydrate metabolism proteins, the beta-xylosidase, a fungi allergen, was identified in A. infectoria EVs [63]. EVs of S. cerevisiae contain proteins related to cell metabolism, cell wall formation, and stress responses [10, 20, 39, 64,65,66,67]. The EVs protein cargo of S. cerevisiae was parallel to that of C. neoformans and H. capsulatum regarding protein classification [21, 55], suggesting that EVs composition is maintained in pathogenic and non-pathogenic fungi. S. cerevisiae is an important model system for analyzing EV-mediated prion transport. A fungal prion, Sup35p, is exported via EVs in soluble and infectious forms [68], and EVs can pass prions to recipient cells where protein accumulation occurs [69].
Proteomic approaches have identified proteins enriched in EVs from many fungal species involved in host–pathogen interactions [8, 20, 21, 54, 55, 60]. In addition, proteins responsible for transport across membranes, such as ATP and ADP carriers and ATPase, have also been demonstrated in EVs of P. braziliensis, S. cerevisiae, C. neoformans, and H. capsulatum [8, 20, 21, 55]. Cell wall remodeling enzymes, including Scw4 and Exg1, or proteins associated with the cell wall, such as Ecm33, have also been detected in the EVs of S. cerevisiae [20]. This indicated a common phenomenon involving the presence of these proteins in EVs among different fungal organisms, which may be correlated with their secretion. The proteins enriched in fungal EVs and their functional classifications are listed in Table 1.
Lipids
Fungal lipids play significant roles in biofilm formation, drug resistance, microdomain production, and EV production. Any alteration in the composition of lipids in the fungal membrane and EVs alters fungal pathogenicity [78]. Lipid composition of fungal EVs is suggested to be similar to that of cells of the same origin. However, advanced investigations have revealed that only a few lipids are designated as a particular type of EVs [79]. The lipid composition of EVs from pathogenic fungal species mainly includes sterol derivatives, phospholipids, and glycosphingolipid GlcCer, which are essential constituents of the plasma membrane [5]. Phospholipids, such as phosphatidylcholine, phosphatidylethanolamine, and phosphatidylserine, are the major components of EVs membranes across a range of fungal species, including C. neoformans, H. capsulatum, and P. braziliensis [5, 16, 55, 80].
EVs derived from C. neoformans are composed of phospholipids, GlcCer, and sterols, such as ergosterols and obtusifoliol [5, 80], similar to the lipids reported for C. albicans EVs [35, 54]. GlcCer and sterols are the main neutral lipids present in C. albicans EVs. GlcCer is critical for dimorphism and disease progression in C. albicans [81]. Lipidomic analysis of EVs from C. albicans and Candida auris demonstrated higher ergosterol concentrations in the EVs of C. albicans than in those of C. auris [54]. Furthermore, the concentrations of diacylglycerols and triacylglycerols are increased in C. albicans [35]. In contrast, most glycerophospholipids were dominant in C. auris [70]. Sphingolipids, such as hexosylceramides, were also found in both species. EVs derived from biofilm-forming C. albicans possess phospholipids, such as phosphatidylethanolamine, phosphatidylinositol, and phosphatidylcholine, together with sphingolipids [30]. In EVs of other Candida species, including C. glabrata, C. parapsilosis, and C. tropicalis, lipid analysis demonstrated the presence of lecithin phospholipids in all three species, with C. parapsilosis exhibiting the highest quantity of lecithin compared to the other two species [82].
In H. capsulatum EVs, different lipids, particularly phosphatidylcholine, phosphatidylserine, and phosphatidylethanolamine, have been identified at specific concentrations in H. capsulatum EVs. All these lipids are considered to serve multiple purposes and are the main components of the structure, along with the biological functioning of the double-layered lipids in both mammals and fungi [55]. H. capsulatum EVs contain ergosterols, diacylglycerides, triacylglycerides, sphingomyelins, phosphoinositol ceramides and lysophospholipids [76]. Lipidomics comparison of fatty acids and sterols between whole-cell lipid extracts and EVs extracts from two separate isolates of P. braziliensis showed compositional similarities and proportional differences. The sterol composition of EVs was brassicasterol, followed by ergosterol and lanosterol, which were prevalent in both isolates. In addition, P. braziliensis EVs are rich in phospholipids, including phosphatidylethanolamine, phosphatidylserine, phosphatidylglycerol, phosphatidylcholine, phosphatidic acid, and phosphatidylinositol [16]. Table 2 lists the types of lipids enriched in the EVs from different fungi.
Carbohydrates
Complex carbohydrates, such as GXM have been shown to be exported by fungal EVs [56] across the cell wall [5]. GXM is a unique feature of C. neoformans and is an essential virulence factor [83]. GXM has been found to be enriched in EVs of Cryptococcus species, both in C. neoformans and C. gattii [5, 34].
In A. fumigatus, carbohydrate analysis showed the presence of mannosyl (Man) and galactosyl (Gal) and a significant amount of glucosyl (Glc) and N-acetyl-galactosaminyl (GalNAc) residues in EVs derived from regenerating protoplasts [23]. GalNAc is an epitope of galactosaminogalactan (GAG), which is a part of the extracellular matrix of A. fumigatus and is associated with virulence [84, 85]. In P. braziliensis, EVs carbohydrates are comprised of glucose, mannose, and galactose residues containing a heavy molecular mass α-4,6-glucan and galactofuranosylmannan, which is probably an oligomer having a 2-α-Manp backbone chain linked to β-Galf (1,3) and α-Manp (1,6) residues [86]. EVs from P. braziliensis carry highly immunogenic α-linked galactopyranosyl epitopes present both on the surface and inside the EVs [87].
In addition, lectin microarrays have shown the presence of terminal Man and GlcNAc residues exposed on the surface of P. braziliensis and Paracoccidioides lutzii EVs identified by DC-SIGN receptors [86]. EVs derived from C. albicans biofilms displayed a high degree of consistency in composition with the matrix material, including proteins and polysaccharides, primarily glucan and mannan [30]. These findings reveal that EVs may be a significant source of matrix material and contribute to resistance against antifungal agents. A description of the carbohydrates enriched in the fungal EVs is presented in Table 3.
Nucleic acids
EVs cargo is generally composed of diverse genetic materials. EVs allow the flow of genetic information owing to the data contained in their RNA and DNA cargoes. However, mitochondrial and genomic DNA are rarely found in EVs [89,90,91]. However, most EVs are rich in RNA, including small non-coding RNAs (such as microRNAs [miRNAs], small nuclear RNA [snRNAs], small nucleolar RNAs [snoRNAs], and tRNA fragments [tRFs]), long non-coding RNAs, and mRNAs that have been identified in EVs from both eukaryotic and prokaryotic organisms [92]. RNA composition in fungal EVs has been described in a range of species including C. neoformans, P. braziliensis, C. albicans, S. cerevisiae, C. gattii, Pichia fermentans, M. sympodialis, and H. capsulatum [13,14,15, 93, 94].
The earliest detailed study on the RNA content in fungal vesicles, known as evRNAs, characterized C.albicans, P. braziliensis, S. cerevisiae, and C. neoformans [14]. In these organisms, EVs transport small RNA molecules secured by EVs membranes through the degradation of exogenous rNase. A previous fungal EVs study showed that most EVs small RNA molecules are less than 250 nucleotides in length on average. Overall, 1246 conserved miRNA-like sequences (milRNAs) were identified, of which 20 sequences were common among the four tested species (P. braziliensis, C. neoformans, C. albicans, and S. cerevisiae). Interestingly, among the ncRNA species, snoRNAs and tRFs were the most frequently observed EVs RNAs in all tested species [14]. Comparative analysis of EVs RNA content derived from the two strains of H. capsulatum revealed a total of 124 mRNAs with significant differences in their composition. Strain-specific short reads ranging from 25 to 40 nucleotides were also identified. Half of these fragments were associated with the reverse transcript strand, indicating that milRNA was present in the fungal EVs. Because these two highly virulent H. capsulatum strains produce EVs that are abundant in RNA classes related to stress responses and translation, an association between EVs RNA and virulence has also been proposed [12]. Comparison of sub-populations of RNA (less than or more than 200 nucleotides in length) in EVs from P. braziliensis (Pb18 and Pb3) and P. lutzii (Pb01) demonstrated that 15 ncRNAs were shared among all samples. snoRNAs were enriched in P. braziliensis EVs, whereas the proportions of snoRNAs, tRNA, and rRNA in P. lutzii EVs were identical. Putative exonic small RNAs are abundant in EVs of Pb18 strain [95]. sRNA classes are involved in modulating transcription and translation. These studies showed that variations in virulence among fungal isolates could be attributed to their distinct EVs RNA contents [95].
Analysis of EVs produced by yeast and pseudohyphal forms of Pichia fermentans showed higher RNA content in EVs from the yeast form. In pseudohyphal EVs, stress-induced spliceosomes and miRNAs associated with hypoxia and cell differentiation are highly expressed. Nevertheless, miRNAs or snRNAs involved in splicing regulation or RNA degradation in both growth forms were similarly expressed in EVs [93]. EVs derived from the human fungal pathogen Rhizopus delemar contain many RNA species, such as mRNAs, lncRNAs, tRNAs, and miRNAs [96]. Next-generation sequencing was performed to investigate the RNA profiles of EVs derived from C. auris. The results showed that EVs contain 104 sequences of non-coding RNAs (tRNAs and tRNAs-half) and 563 sequences of messenger RNAs, snoRNAs, and rRNAs [70]. M. sympodialis also possesses small RNAs which are a collection of 16–22 nucleotides with well-defined start and stop loci. Although no genes encoding the components of the RNAi machinery are found in this organism, they possess an RNAi-independent mechanism for the biogenesis of these small RNA molecules [97].
RNA molecules are considered as crucial players in intercellular communication. Their involvement in communication derives from their ability to control gene expression in recipient cells by sending specific RNAs [98]. EVs have been used for RNA transfer in C. gattii. Macrophages infected with a non-outbreak C. gattii strain take up EVs originating from a highly virulent C. gattii strain and lead to increased survival of C. gattii inside macrophages [15]. This indicates that EV RNA is necessary for virulence transmission.
Effects of EVs on fungal pathogenicity
Fungal EVs contain various bioactive substances. This suggests that they potentially participate in intercellular communication and influence fungal pathogenicity.
Biofilm generation
Biofilms are fixed communities of microorganisms that allow cells to firmly adhere to each other and other surfaces and are guarded by a polymeric extracellular matrix composed of polysaccharides [99]. Cells in biofilms exhibit enhanced resistance along with distinctive phenotypes compared to planktonic or unadhered cells, and are also associated with the perseverance of infections [100]. Production of biofilms is a major phenomenon in the pathogenesis of C. albicans [101], and the biofilm matrix forms a defense mechanism against antifungal drugs [102]. The cargo of biofilm EVs consists of proteins and carbohydrates, mainly mannan and glucan, similar to those in the biofilm matrix, making EVs a notable factor in establishing the biofilm matrix [30]. As determined by 1 H and 2D 1 H-13 C NMR, the major mannan constituents in the vesicle cargo exhibited structural resemblance to the biofilm matrix mannan–glucan complex, which is a biofilm-related drug resistance-determining factor [103]. The activity of biofilm EVs was analyzed using C. albicans ESCRT mutants, which led to decreased biofilm EVs production, decreased matrix polysaccharide levels, and higher fluconazole vulnerability than wild-type C. albicans. The “add-back” of EVs from wildtype biofilms reestablishes matrix accumulation and biofilm drug resistance [30]. This finding suggests that the biofilm EVs of C. albicans play critical roles in the development of matrices and antimicrobial resistance. Furthermore, biofilms may mediate drug resistance through the overexpression of drug efflux pump proteins [102, 104]. The presence of drug efflux pump proteins in the EVs of C. albicans is indicative of the biological function of EVs in azole resistance by transporting efflux pump proteins to non-resistant strains [59]. Additionally, EVs of biofilms have large amounts of oxidative, heat stress, and virulence-inducing proteins, indicating a possible role for EVs in pathogenesis [59].
Mediation of fungal virulence
The importance of fungal EVs in mediating virulence has been previously proposed. The fatal human pathogen C. gattii secretes EVs to mediate the virulence of neighboring fungal cells [15]. The outbreak lineage of C. gattii has the ability proliferates rapidly within host phagocytes [105]. Previously, reports has demonstrated that this rapid proliferation is driven by a ‘division of labor’ mechanism [106]. Division of labor is mediated by the secretion of fungal EVs. Virulent-strain-derived EVs are taken up by infected host macrophages and trigger rapid intracellular proliferation of the non-outbreak lineage [15].
Effects of fungal EVs on host–pathogen interactions
The presence of different bioactive components in fungal EVs increases the probability of their association with prospective hosts. The interaction between fungal EVs and the host mainly occurs through direct biological effects and immunomodulation. Figure 2 provides an overview of fungal EVs in host–pathogen interactions.
Direct biological effects on the host
Several studies have demonstrated that virulence-related molecules are exported directly or via fungal EVs, which could have direct biological effects on the host. The virulence factors enriched in fungal EVs and their biological effects on the host are summarized in Table 4.
In C. neoformans, GXM is a vital element of the fungal polysaccharide capsule that exerts both immunosuppressive and cytotoxic effects on immune cells [107,108,109,110], in order to avoid the killing of fungi by macrophages through phagocytosis [111]. Melanin production enhances the resistance to macrophage phagocytosis and reactive oxygen species derived from host cells [7, 112]. EVs released by C. neoformans are also capable of enhancing brain infection, as they boost the transmigration of yeast across the blood-brain barrier [40]. Small RNAs, including microRNAs (miRNAs), are significant constituents of EVs in C. albicans as well as C. neoformans. The RNAi machinery present in both fungal species suggests the role played by EV-derived small RNA molecules during fungal infections by mimicking endogenous miRNAs to modulate gene expression in host cells [113]. Recently, proteomic analysis demonstrated that EVs derived from Talaromyces marneffei contain heat shock proteins and mannoprotein 1, which maintain homeostasis and pathogenicity [77].
Fungal EVs contain components with both pathogenic and immunogenic properties that allow fungal EVs to trigger the host immune response [58]. Phospholipid phosphatidylserine is an essential element of C. albicans EVs to activate immune cells, as phosphatidylserine synthase CHO1 deficient EVs are unable to stimulate NF-κB in bone marrow-derived macrophages (BMDMs) [35]. Glucosylceramide is another vital antigenic molecule that triggers immune responses and is found in the EVs of C. albicans and C. neoformans [114, 115]. EVs of P. braziliensis carry epitopes of α-galactopyranosyl, an extremely immunogenic particle, which were recognized efficiently by anti-α-gal antibodies within patients with paracoccidioidomycosis [87]. Recently, proteomic analysis of EVs derived from NAC, such as C. tropicalis, C. parapsilosis, and C. glabrata, demonstrated abundant moonlighting proteins or their orthologs with the ability to induce immune responses, adhesive roles, and pathogenic potential [60].
Modulation of the host immune response
Recent studies have shown that fungal EVs strongly influence host immunity [17, 116]. In C. neoformans, EVs are involved in modulating the phagocytosis index and antimicrobial potential of macrophages, as they induce the secretion of high concentrations of cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-10, transforming growth factor-beta (TGF-β), and nitric oxide (NO). Interestingly, both TNF and NO production, together with an improved capacity to phagocytose and destroy fungal cells, indicate the dual role of EVs in triggering positive and negative activation of macrophages [117]. As observed with C. neoformans EVs, immunomodulation has also been reported for C. albicans in host immune cells. EVs internalization by dendritic cells (DCs) and macrophages induces the production of IL-12, TNF-α, IL-10, TGF-α, and nitric oxide as well as increased levels of MHC-II and CD86 expression in DCs [54]. This cytokine signature is balanced because anti-inflammatory cytokines (IL-10 and TGF-β) serve as host response breaks that can inhibit or decrease tissue damage induced by an intensified inflammatory reaction.
EVs derived from M. sympodialis were found to interact with human keratinocytes and induce increased expression of intercellular adhesion molecule-1, which is indicative of a significant step in the skin defense against this fungal pathogen [118]. EVs from the saprophytic fungus Aspergillus flavus stimulate macrophages to produce inflammatory mediators, including TNF-α, IL-1, IL-6, and NO, and induce M1 polarization [17]. Similar results were delineated by EVs derived from P. braziliensis, while secondary stimulation promoted the secretion of pro-inflammatory cytokines, such as TNF-α, IL-6, and IL-12, and favored M1 polarization [119].
Macrophages expressing the M1 phenotype play an important role in eliminating fungal pathogens. The development of M1 and inhibition of M2 phenotypic expression have also been demonstrated, as they provide protection against infections caused by C. neoformans, H. capsulatum, and A. fumigatus [120,121,122]. EVs from H. capsulatum lower the phagocytic activity and intracellular killing by BMDMs [44]. These results indicate the immunomodulatory activity of EVs. Depending on the pathogen, fungal EVs can intensify or attenuate the progression of the infection [110].

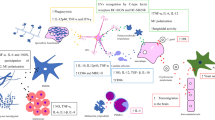
Biological effects of fungal EVs on fungal pathogenicity and host–pathogen interactions. As an important medium of intercellular communication, fungal EVs can mediate both fungal cell–cell communication and host–pathogen interactions. Biofilm EVs carrying biological molecules are involved in biofilm generation and resistance in C. albicans [30]. C. gattii EVs mediate the virulence of neighboring fungal cells [15]. C. neoformans EVs contain glucuronoxylomannan (GXM) and exert both immunosuppressive and cytotoxic effects on immune cells [107,108,109,110]. MicroRNAs are significant constituents of EVs in C. albicans and C. neoformans and modulate the gene expression in host cells [113]. EVs modulate the phagocytosis index and induce cytokine production by macrophages in C. neoformans [117]. In A. flavus, EVs induce M1 polarization of macrophages [17]. C. albicans EVs activate dendritic cells (DCs) for antigen presentation and cytokine secretion [54]
Potential fungal EV markers
Suitable fungal EV markers are required to improve EVs isolation methods and conduct detailed research on the composition and biological functions of EVs. Exosomes from mammals have been purified and analyzed using marker proteins, including CD63, Alix, and TSG101 [123, 124], whereas for all EVs, the markers CD9 and CD81 have been used [125]. However, most of the markers for mammalian EVs do not possess orthologs in fungal EVs [126]. Vps27 and Hse1, proteins of the ESCRT complex, have been found in EVs of C. albicans but at very low concentrations [30]. Comparative proteomic analysis of EVs derived from H. capsulatum, S. cerevisiae, P. braziliensis, and C. neoformans has revealed 26 protein orthologs in EVs from all four species that may serve as marker candidates [8]. Nonetheless, these results are less convincing when analyzed for individual fungal species [126]. Hsp70, a marker protein found in mammalian exosomes [127], has also been detected in C. albicans EVs. An Hsp71-like protein found in C. neoformans, which shares more than 80% sequence similarity with the Hsp70 protein found in the proteomic dataset of C. albicans EVs, suggests that Hsp70 could be a possible marker for fungal EVs [126].
A recent analysis revealed 22 possible protein markers, including the claudin-like Sur7 family proteins Sur7 and Evp1, in the EVs of C. albicans. Based on their possible topological resemblance to tetraspanin markers used for mammalian EVs, Sur7 and Evp1 have been proposed as putative positive markers for C. albicans EVs [59]. Sur7 proteins have also been recently described in EVs from Cryptococcus [57]. In another study, a monoclonal antibody (mAb476) that specifically recognized Galactofuranose(Galf)-bearing glycoconjugates of Aspergillus was described. EVs fractions of both Aspergillus culture supernatant and clinical samples of patients with invasive aspergillosis (IA) were recognized by mAb476. This finding suggesting that Galf is considered a potential marker of Aspergillus EVs and Galf-bearing Aspergillus EVs may be used for clinical diagnostic applications of IA [128]. Moreover, the major lipid component GlcCer is an intriguing fungal EV marker, although certain species, including C. glabrata and S. cerevisiae, do not generate or synthesize this GSL [129]. No other fungus-specific markers have been identified to date. Unstandardized methods for isolating fungal EVs have significantly limited the development of fungal EV markers.
Potential therapeutic applications of fungal EVs
There is an increasing need for the development of innovative methodologies for preventing and treating infections caused by pathogenic fungi, as demonstrated by the elevated number of fatalities caused by invasive mycoses. Several attempts have been made to develop vaccines for the prevention of fungal infections, but none have been licensed for public use [110, 130]. In mammalian studies, exosomes are used as biomarkers, cell-free therapeutic agents and cancer vaccine [131, 132]. Fungal EVs have nano-sized structures, contain several immunogenic molecules, and interact with the host in multiple ways, making them suitable candidates for therapeutic applications. Fig. 3 summarizes the potential clinical applications of fungal EVs.
Vaccine development
EVs from fungal species possess molecules capable of stimulating immunogenic reactions, endowing fungal EVs with potential for vaccine development. Bgl2 or 1,3-glucosyltransferase is a protein component of EVs in C. albicans, which is responsible for the biosynthesis of fungal cell walls and virulence, and can elicit humoral immunity by reacting with the sera of patients with candidiasis [133]. A mouse model showed increased survival following treatment with BgI2 in the vaccine before infection with C. albicans [58]. Moreover, the ortholog of the enolase protein (Eno1) present in the EVs of C. albicans was found to be abundant as an immunodominant antigenic protein in candidiasis patients [134]. Furthermore, glycoprotein 43 (gp43) is a protein conjugate present in the fungal species P. braziliensis that is also present in EVs of P. braziliensis [8]. Immunization with P10 lowered the fungal load and improved the survival of healthy or immunosuppressed mice through a process based on the Th1 immune response after lethal challenge with P. braziliensis [135,136,137].
Cryo-EM analysis revealed a novel antigenic structure present on the surface of Cryptococcus EVs [138], resembling the spike complexes present on the envelope of viruses [139, 140], suggesting their role as potential vaccines. Indeed, mice immunized with EVs acquired from a mutated capsular strain of C. neoformans showed a robust antibody response and substantially extended survival following infection with C. neoformans [138]. Cryptococcal infection can be prevented through prophylactic immunization, and several antigens of cryptococcus have been examined for their potential as vaccines [141, 142]. A mutant strain of C. neoformans lacking sterylglucosidase 1 (Sgl1), an enzyme required for sterylglucoside degradation, is nonpathogenic in cryptococcosis mouse models. Interestingly, this mutant strain acted as a vaccine that triggered a protective immune response in immunocompromised (CD4 cell-depleted) mice against cryptococcosis after challenge with WT C. neoformans or C. gattii. The deposition of SGs in Δsgl1 mutant may be responsible for this protective immunity. These results are especially relevant in the sense of HIV/AIDS immune deficiency, indicating that a new vaccine strategy against cryptococcosis could be given by the Δsgl1 strain [143].
Immunotherapy application
Numerous studies have demonstrated the immunotherapeutic potential of fungal EVs [17,18,19]. Prophylaxis of Galleria mellonella with C. albicans EVs prior to infection with live C. albicans reduces fungal stress and enhances larval survival [54]. Pretreatment of G. mellonella with A. flavus EVs also decreases CFU levels and enhances the larval survival [17]. Similarly, GXM-containing EVs of C. neoformans induce immunity in G. mellonella against lethal challenge with C. neoformans [144]. EVs from C. albicans may exert a protective effect against murine candidiasis [19].
Collectively, these studies demonstrate the potential of fungal EVs to modulate the natural immune response and intercept pathogenesis in vivo to control fungal infection, suggesting native or engineered EVs as promising candidates for therapeutic applications.

Potential clinical applications of fungal EVs. The membranous structure of fungal EVs allows them to carry various bioactive components, providing fungal EVs great potential for clinical applications. Suitable fungal EVs markers may be useful for fungal infection diagnosis [145] and new EVs isolation strategy [59]. Antigens and immunogenic proteins presented in fungal EVs are able to activate immune response in the host, providing the possibility of fungal EVs in vaccine development and immunotherapy application [54]
Conclusion and future challenges
In recent years, fungal EVs have gained increasing attention in infectious disease and fungal pathogen research. More extensive and in-depth research will further enhance our understanding of fungal EVs. Despite the recent advancements in fungal EV research, several challenges must be addressed in future studies.
-
1)
Unlike mammalian EVs, the absence of specific biomarkers in fungal EVs has hampered their in-depth study. Detection of new EV markers is a useful resource for studying the biogenesis, cargo packing, isolation, and characterization of fungal EVs and determining their role in pathogenesis.
-
2)
Molecular processes governing the biogenesis, cell wall-crossing mechanisms, and compositional diversity of EVs must be investigated to better understand their biological roles.
-
3)
In-depth studies of the host–pathogen interactions mediated by fungal EVs will provide insights on the pathogenic mechanism of fungi and aid in the development of new treatment strategies for fungal infections.
-
4)
Influence of fungal EVs on recipient fungi and other microorganisms also requires further investigation.
-
5)
Lastly, the potential of fungal EVs as biomarkers for the early diagnosis and therapeutic monitoring of fungal infections needs to be assessed in future studies.
Data Availability
Not applicable.
References
Van Niel G, d’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–28.
Gill S, Catchpole R, Forterre P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol Rev. 2019;43(3):273–303.
Bielska E, May RC. Extracellular vesicles of human pathogenic fungi. Curr Opin Microbiol. 2019;52:90–9.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science, 2020. 367(6478).
Rodrigues ML, et al. Vesicular polysaccharide export in Cryptococcus neoformans is a eukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot Cell. 2007;6(1):48–59.
Rizzo J, Rodrigues ML, Janbon G. Extracellular vesicles in Fungi: past, Present, and future perspectives. Front Cell Infect Microbiol, 2020. 10.
Eisenman HC, et al. Vesicle-associated melanization in Cryptococcus neoformans. Microbiology. 2009;155(Pt 12):3860.
Vallejo MC, et al. Vesicle and vesicle-free extracellular proteome of Paracoccidioides brasiliensis: comparative analysis with other pathogenic fungi. J Proteome Res. 2012;11(3):1676–85.
Wolf JM, et al. Interaction of Cryptococcus neoformans extracellular vesicles with the cell wall. Eukaryot Cell. 2014;13(12):1484–93.
Zhao K, et al. Extracellular vesicles secreted by Saccharomyces cerevisiae are involved in cell wall remodelling. Commun biology. 2019;2(1):1–13.
Herkert PF, et al. Extracellular vesicles as vehicles for the delivery of biologically active fungal molecules. Curr Protein Pept Sci. 2019;20(10):1027–36.
Alves LR et al. Extracellular vesicle-mediated RNA release in Histoplasma capsulatum. Msphere, 2019. 4(2).
Rayner S, et al. Identification of small RNAs in extracellular vesicles from the commensal yeast Malassezia sympodialis. Sci Rep. 2017;7(1):1–9.
Da Silva RP, et al. Extracellular vesicle-mediated export of fungal RNA. Sci Rep. 2015;5:7763.
Bielska E, et al. Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus gattii. Nat Commun. 2018;9(1):1–9.
Vallejo MC, et al. Lipidomic analysis of extracellular vesicles from the pathogenic phase of Paracoccidioides brasiliensis. PLoS ONE. 2012;7(6):e39463.
Brauer VS et al. Extracellular vesicles from Aspergillus flavus Induce M1 polarization in Vitro. mSphere, 2020. 5(3).
Marina CL, et al. Nutritional conditions modulate C. neoformans extracellular vesicles’ capacity to elicit host immune response. Microorganisms. 2020;8(11):1815.
Vargas G, et al. Protective effect of fungal extracellular vesicles against murine candidiasis. Cell Microbiol. 2020;22(10):e13238.
Oliveira DL, et al. Characterization of yeast extracellular vesicles: evidence for the participation of different pathways of cellular traffic in vesicle biogenesis. PLoS ONE. 2010;5(6):e11113.
Rodrigues ML, et al. Extracellular vesicles produced by Cryptococcus neoformans contain protein components associated with virulence. Eukaryot Cell. 2008;7(1):58–67.
Rodrigues ML, et al. Vesicular mechanisms of traffic of fungal molecules to the extracellular space. Curr Opin Microbiol. 2013;16(4):414–20.
Rizzo J et al. Characterization of extracellular vesicles produced by aspergillus fumigatus protoplasts. bioRxiv, 2020.
Rodrigues ML, et al. Human antibodies against a purified glucosylceramide from Cryptococcus neoformans inhibit cell budding and fungal growth. Infect Immun. 2000;68(12):7049–60.
Panepinto J, et al. Section 6-dependent sorting of fungal extracellular exosomes and laccase of Cryptococcus neoformans. Mol Microbiol. 2009;71(5):1165–76.
da Peres R, et al. Golgi reassembly and stacking protein (GRASP) participates in vesicle-mediated RNA export in Cryptococcus neoformans. Genes. 2018;9(8):400.
Kmetzsch L, et al. Role for golgi reassembly and stacking protein (GRASP) in polysaccharide secretion and fungal virulence. Mol Microbiol. 2011;81(1):206–18.
Kinseth MA, et al. The golgi-associated protein GRASP is required for unconventional protein secretion during development. Cell. 2007;130(3):524–34.
Duran JM, et al. Unconventional secretion of Acb1 is mediated by autophagosomes. J Cell Biol. 2010;188(4):527–36.
Zarnowski R, et al. Candida albicans biofilm–induced vesicles confer drug resistance through matrix biogenesis. PLoS Biol. 2018;16(10):e2006872.
Park Y-D, et al. Role of the ESCRT pathway in laccase trafficking and virulence of Cryptococcus neoformans. Infection and Immunity; 2020.
Rizzo J, et al. Role of the Apt1 protein in polysaccharide secretion by Cryptococcus neoformans. Eukaryot Cell. 2014;13(6):715–26.
Rizzo J et al. The putative flippase Apt1 is required for intracellular membrane architecture and biosynthesis of polysaccharide and lipids in Cryptococcus neoformans Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2018. 1865(3): p. 532–41.
Reis FC, et al. A novel protocol for the isolation of fungal extracellular vesicles reveals the participation of a putative scramblase in polysaccharide export and capsule construction in Cryptococcus gattii. MSphere. 2019;4(2):e00080–19.
Wolf JM, et al. Lipid biosynthetic genes affect Candida albicans extracellular vesicle morphology, cargo, and immunostimulatory properties. Eukaryot Cell. 2015;14(8):745–54.
Rodrigues J, et al. Lack of chitin synthase genes impacts capsular architecture and cellular physiology in Cryptococcus neoformans. Cell Surf. 2018;2:14–23.
García-Rodas R et al. Capsule growth in Cryptococcus neoformans is coordinated with cell cycle progression. MBio, 2014. 5(3).
Tefsen B, et al. Deletion of the CAP10 gene of Cryptococcus neoformans results in a pleiotropic phenotype with changes in expression of virulence factors. Res Microbiol. 2014;165(6):399–410.
Winters CM, Hong-Brown LQ, Chiang HL. Intracellular vesicle clusters are organelles that synthesize extracellular vesicle-associated cargo proteins in yeast. J Biol Chem. 2020;295(9):2650–63.
Huang S-H, et al. Cryptococcus neoformans-derived microvesicles enhance the pathogenesis of fungal brain infection. PLoS ONE. 2012;7(11):e48570.
Li J, et al. The 14-3-3 gene function of Cryptococcus neoformans is required for its growth and virulence. J Microbiol Biotechnol. 2016;26(5):918–27.
Winters M, C. and, Chiang H-L. Yeast as a model system to study trafficking of small vesicles carrying signal-less proteins in and out of the cell. Curr Protein Pept Sci. 2016;17(8):808–20.
Baltazar LM et al. Antibody binding alters the characteristics and contents of extracellular vesicles released by Histoplasma capsulatum. Msphere, 2016. 1(2).
Baltazar LM, et al. Concentration-dependent protein loading of extracellular vesicles released by Histoplasma capsulatum after antibody treatment and its modulatory action upon macrophages. Sci Rep. 2018;8(1):1–10.
Nimrichter L, et al. The multitude of targets for the immune system and drug therapy in the fungal cell wall. Microbes Infect. 2005;7(4):789–98.
Brown L, et al. Through the wall: extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat Rev Microbiol. 2015;13(10):620–30.
Wolf JM, Casadevall A. Challenges posed by extracellular vesicles from eukaryotic microbes. Curr Opin Microbiol. 2014;22:73–8.
Walker L et al. The viscoelastic properties of the fungal cell wall allow traffic of AmBisome as intact liposome vesicles. MBio, 2018. 9(1).
de Souza Pereira R, Geibel J. Direct observation of oxidative stress on the cell wall of Saccharomyces cerevisiae strains with atomic force microscopy. Mol Cell Biochem. 1999;201(1–2):17–24.
Jacobson ES, Ikeda R. Effect of melanization upon porosity of the cryptococcal cell wall. Med Mycol. 2005;43(4):327–33.
Nimrichter L, et al. Extracellular vesicle-associated transitory cell wall components and their impact on the interaction of fungi with host cells. Front Microbiol. 2016;7:1034.
Rodrigues ML, et al. The impact of proteomics on the understanding of functions and biogenesis of fungal extracellular vesicles. J Proteom. 2014;97:177–86.
Shopova IA et al. Human neutrophils produce antifungal extracellular vesicles against aspergillus fumigatus. Mbio, 2020. 11(2).
Vargas G, et al. Compositional and immunobiological analyses of extracellular vesicles released by C andida albicans. Cell Microbiol. 2015;17(3):389–407.
Albuquerque PC, et al. Vesicular transport in Histoplasma capsulatum: an effective mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell Microbiol. 2008;10(8):1695–710.
Rodrigues ML, et al. Vesicular trans-cell wall transport in fungi: a mechanism for the delivery of virulence-associated macromolecules? Lipid insights. 2008;2:LPI.
Rizzo J et al. New insights into Cryptococcus extracellular vesicles suggest a new structural model and an antifungal vaccine strategy BioRxiv, 2020: p. 2020.08. 17.253716.
Gil-Bona A, et al. Proteomics unravels extracellular vesicles as carriers of classical cytoplasmic proteins in Candida albicans. J Proteome Res. 2015;14(1):142–53.
Dawson CS, et al. Protein markers for Candida albicans EVs include claudin-like Sur7 family proteins. J Extracell vesicles. 2020;9(1):1750810.
Karkowska-Kuleta J, et al. Characteristics of extracellular vesicles released by the pathogenic yeast-like fungi Candida glabrata, Candida parapsilosis and Candida tropicalis. Cells. 2020;9(7):1722.
de Toledo Martins S et al. Extracellular vesicles in fungi: composition and functions. Fungal Physiol Immunopathogenesis, 2018: p. 45–59.
Johansson HJ, et al. Extracellular nanovesicles released from the commensal yeast Malassezia sympodialis are enriched in allergens and interact with cells in human skin. Sci Rep. 2018;8(1):1–11.
Silva BM, et al. Characterization of Alternaria infectoria extracellular vesicles. Med Mycol. 2014;52(2):202–10.
Giardina BJ, Stein K, Chiang H-L. The endocytosis gene END3 is essential for the glucose-induced rapid decline of small vesicles in the extracellular fraction in Saccharomyces cerevisiae. J Extracell vesicles. 2014;3(1):23497.
Kabani M, Pilard M, Melki R. Glucose availability dictates the export of the soluble and prion forms of Sup35p via periplasmic or extracellular vesicles. Mol Microbiol. 2020;114(2):322–32.
Stein K, Winters C, Chiang HL. Vps15p regulates the distribution of cup-shaped organelles containing the major eisosome protein Pil1p to the extracellular fraction required for endocytosis of extracellular vesicles carrying metabolic enzymes. Biol Cell. 2017;109(5):190–209.
Kabani M, Melki R. Sup35p in its soluble and prion states is packaged inside extracellular vesicles. MBio. 2015;6(4):e01017–15.
Kabani M, Melki R. Sup35p in its soluble and prion states is packaged inside extracellular vesicles. MBio, 2015. 6(4).
Liu S et al. Horizontal transmission of cytosolic Sup35 prions by extracellular vesicles. MBio, 2016. 7(4).
Zamith-Miranda D et al. Comparative molecular and immunoregulatory analysis of extracellular vesicles from Candida albicans and Candida auris bioRxiv, 2020.
Konečná K, et al. A comparative analysis of protein virulence factors released via extracellular vesicles in two Candida albicans strains cultivated in a nutrient-limited medium. Microb Pathog. 2019;136:103666.
Martínez-López R et al. Small extracellular vesicles secreted by Candida albicans hyphae have highly diverse protein cargoes that include virulence factors and stimulate macrophages bioRxiv, 2020: p. 2020.10.02.323774.
Rizzo J, et al. Characterization of extracellular vesicles produced by aspergillus fumigatus protoplasts. MSphere. 2020;5(4):e00476–20.
Souza JAM, et al. Characterization of aspergillus fumigatus extracellular vesicles and their effects on macrophages and neutrophils functions. Front Microbiol. 2019;10:2008.
Matos Baltazar L, et al. Antibody binding alters the characteristics and contents of extracellular vesicles released by Histoplasma capsulatum. MSphere. 2016;1(2):e00085–15.
Cleare LG, et al. Media matters! Alterations in the loading and release of Histoplasma capsulatum extracellular vesicles in response to different nutritional milieus. Cell Microbiol. 2020;22(9):e13217.
Yang B, et al. Extracellular vesicles derived from Talaromyces marneffei yeasts mediate inflammatory response in macrophage cells by bioactive protein components. Front Microbiol. 2021;11:603183.
Abels ER, Breakefield XO. Introduction to Extracellular vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol Neurobiol. 2016;36(3):301–12.
Llorente A et al. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids, 2013. 1831(7): p. 1302–9.
Oliveira DL, et al. Cryptococcus neoformans cryoultramicrotomy and vesicle fractionation reveals an intimate association between membrane lipids and glucuronoxylomannan. Fungal Genet Biol. 2009;46(12):956–63.
Gow NA, et al. Candida albicans morphogenesis and host defence: discriminating invasion from colonization. Nat Rev Microbiol. 2012;10(2):112–22.
Karkowska-Kuleta J et al. Characteristics of Extracellular vesicles released by the pathogenic yeast-like Fungi Candida glabrata, Candida parapsilosis and Candida tropicalis. Cells, 2020. 9(7).
O’Meara TR, Alspaugh JA. The Cryptococcus neoformans capsule: a sword and a shield. Clin Microbiol Rev. 2012;25(3):387–408.
Fontaine T, et al. Galactosaminogalactan, a new immunosuppressive polysaccharide of aspergillus fumigatus. PLoS Pathog. 2011;7(11):e1002372.
Gresnigt MS, et al. A polysaccharide virulence factor from Aspergillus fumigatus elicits anti-inflammatory effects through induction of Interleukin-1 receptor antagonist. PLoS Pathog. 2014;10(3):e1003936.
Peres da Silva R, et al. Extracellular vesicles from Paracoccidioides pathogenic species transport polysaccharide and expose ligands for DC-SIGN receptors. Sci Rep. 2015;5:14213.
Vallejo MC, et al. The pathogenic fungus paracoccidioides brasiliensis exports extracellular vesicles containing highly immunogenic α-Galactosyl epitopes. Eukaryot Cell. 2011;10(3):343–51.
da Silva RP, et al. Extracellular vesicles from Paracoccidioides pathogenic species transport polysaccharide and expose ligands for DC-SIGN receptors. Sci Rep. 2015;5(1):1–12.
Guescini M, et al. Astrocytes and glioblastoma cells release exosomes carrying mtDNA. J Neural Transm. 2009;117(1):1.
Balaj L, et al. Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat Commun. 2011;2(1):180.
Waldenström A, et al. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS ONE. 2012;7(4):e34653.
Yáñez-Mó M, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell vesicles. 2015;4(1):27066.
Leone F, et al. Analysis of extracellular vesicles produced in the biofilm by the dimorphic yeast Pichia fermentans. J Cell Physiol. 2018;233(4):2759–67.
Alves LR, et al. Extracellular vesicle-mediated RNA release in Histoplasma capsulatum. Msphere. 2019;4(2):e00176–19.
Peres da Silva R et al. Comparison of the RNA content of Extracellular vesicles derived from Paracoccidioides brasiliensis and paracoccidioides lutzii. Cells, 2019. 8(7).
Liu M, et al. Comparative genome-wide analysis of extracellular small RNAs from the mucormycosis pathogen Rhizopus delemar. Sci Rep. 2018;8(1):5243.
Rayner S, et al. Identification of small RNAs in extracellular vesicles from the commensal yeast Malassezia sympodialis. Sci Rep. 2017;7:39742.
Iguchi H, Kosaka N, Ochiya T. Secretory microRNAs as a versatile communication tool. Commun Integr Biol. 2010;3(5):478–81.
Chatterjee S, Das S. Developmental stages of biofilm and characterization of extracellular matrix of manglicolous fungus aspergillus niger BSC-1. J Basic Microbiol. 2020;60(3):231–42.
Kowalski CH, et al. Fungal biofilm architecture produces hypoxic microenvironments that drive antifungal resistance. Proc Natl Acad Sci. 2020;117(36):22473–83.
Nobile CJ, Johnson AD. Candida albicans Biofilms and Human Disease. Annu Rev Microbiol. 2015;69:71–92.
Taff HT, et al. A Candida biofilm-induced pathway for matrix glucan delivery: implications for drug resistance. PLoS Pathog. 2012;8(8):e1002848.
Mitchell KF, et al. Community participation in biofilm matrix assembly and function. Proc Natl Acad Sci. 2015;112(13):4092–7.
Holmes AR, et al. ABC transporter Cdr1p contributes more than Cdr2p does to fluconazole efflux in fluconazole-resistant Candida albicans clinical isolates. Antimicrob Agents Chemother. 2008;52(11):3851–62.
Ma H, et al. The fatal fungal outbreak on Vancouver Island is characterized by enhanced intracellular parasitism driven by mitochondrial regulation. Proc Natl Acad Sci. 2009;106(31):12980–5.
Voelz K, et al. Division of labour’in response to host oxidative burst drives a fatal Cryptococcus gattii outbreak. Nat Commun. 2014;5(1):1–12.
Monari C, et al. Cryptococcus neoformans capsular glucuronoxylomannan induces expression of fas ligand in macrophages. J Immunol. 2005;174(6):3461–8.
Monari C, Bistoni F, Vecchiarelli A. Glucuronoxylomannan exhibits potent immunosuppressive properties. FEMS Yeast Res. 2006;6(4):537–42.
Villena SN, et al. Capsular polysaccharides galactoxylomannan and glucuronoxylomannan from Cryptococcus neoformans induce macrophage apoptosis mediated by Fas ligand. Cell Microbiol. 2008;10(6):1274–85.
Freitas MS, et al. Fungal extracellular vesicles as potential targets for immune interventions. Msphere. 2019;4(6):e00747–19.
Zaragoza O, et al. The capsule of the fungal pathogen Cryptococcus neoformans. Adv Appl Microbiol. 2009;68:133–216.
Coelho C, Bocca AL, Casadevall A. The tools for virulence of Cryptococcus neoformans. Adv Appl Microbiol. 2014;87:1–41.
da Peres R, et al. Extracellular vesicle-mediated export of fungal RNA. Sci Rep. 2015;5(1):1–12.
Rittershaus PC, et al. Glucosylceramide synthase is an essential regulator of pathogenicity of Cryptococcus neoformans. J Clin Investig. 2006;116(6):1651–9.
Noble SM, et al. Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat Genet. 2010;42(7):590–8.
Esher SK, Zaragoza O, Alspaugh JA. Cryptococcal pathogenic mechanisms: a dangerous trip from the environment to the brain. Mem Inst Oswaldo Cruz, 2018. 113(7).
Oliveira DL, et al. Extracellular vesicles from Cryptococcus neoformans modulate macrophage functions. Infect Immun. 2010;78(4):1601–9.
Vallhov H, et al. Extracellular vesicles released from the skin commensal yeast Malassezia sympodialis activate human primary keratinocytes. Front Cell Infect Microbiol. 2020;10:6.
Da Silva TA, et al. Extracellular vesicles from Paracoccidioides brasiliensis induced M1 polarization in vitro. Sci Rep. 2016;6:35867.
Wager CL, Wormley F. Classical versus alternative macrophage activation: the Ying and the Yang in host defense against pulmonary fungal infections. Mucosal Immunol. 2014;7(5):1023–35.
Moreira AP, et al. Serum amyloid P attenuates M2 macrophage activation and protects against fungal spore–induced allergic airway disease. J Allergy Clin Immunol. 2010;126(4):712–21. e7.
Müller U, et al. IL-13 induces disease-promoting type 2 cytokines, alternatively activated macrophages and allergic inflammation during pulmonary infection of mice with Cryptococcus neoformans. J Immunol. 2007;179(8):5367–77.
Mathivanan S, Simpson RJ. ExoCarta: a compendium of exosomal proteins and RNA. Proteomics. 2009;9(21):4997–5000.
Willms E, et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci Rep. 2016;6(1):1–12.
Yoshioka Y et al. Comp marker Anal Extracell vesicles different Hum cancer types J Extracell Vesicles 2. 2013.
Bleackley MR, Dawson CS, Anderson MA. Fungal extracellular vesicles with a focus on proteomic analysis. Proteomics. 2019;19(8):1800232.
Lancaster GI, Febbraio MA. Exosome-dependent trafficking of HSP70 a novel secretory pathway for cellular stress proteins. J Biol Chem. 2005;280(24):23349–55.
Kausik Datta KW, Rock M, Lowary JesseFrancisT. Susmita Sarkar, Kieren Marr. Characterisation of mAb476-reactive galactofuranose-bearing Aspergillus antigens excreted in urine. in European Congress of Clinical Microbiology & Infectious Diseases 2019 (ECCMID 2019). 2019. Amsterdam, Netherlands.
Tavares PM, et al. In vitro activity of the antifungal plant defensin RsAFP2 against Candida isolates and its in vivo efficacy in prophylactic murine models of candidiasis. Antimicrob Agents Chemother. 2008;52(12):4522–5.
Boniche C, et al. Immunotherapy against systemic fungal infections based on monoclonal antibodies. J Fungi. 2020;6(1):31.
Rezaie J, Etemadi T, Feghhi M. The distinct roles of exosomes in innate immune responses and therapeutic applications in cancer. Eur J Pharmacol. 2022;933:175292.
Rezaie J, Feghhi M, Etemadi T. A review on exosomes application in clinical trials: perspective, questions, and challenges. Cell Commun Signal. 2022;20(1):145.
Pitarch A, et al. Decoding serological response to Candida cell wall immunome into novel diagnostic, prognostic, and therapeutic candidates for systemic candidiasis by proteomic and bioinformatic analyses. Mol Cell Proteom. 2006;5(1):79–96.
Pitarch A, Nombela C, Gil C. Serum antibody signature directed against Candida albicans Hsp90 and enolase detects invasive candidiasis in non-neutropenic patients. J Proteome Res. 2014;13(11):5165–84.
Taborda CP, et al. Mapping of the T-Cell epitope in the major 43-Kilodalton glycoprotein of Paracoccidioides brasiliensisWhich induces a Th-1 response protective against fungal infection in BALB/c mice. Infect Immun. 1998;66(2):786–93.
Marques AF, et al. Additive effect of P10 immunization and chemotherapy in anergic mice challenged intratracheally with virulent yeasts of Paracoccidioides brasiliensis. Microbes Infect. 2008;10(12–13):1251–8.
Munoz JE, et al. Immunization with P10 peptide increases specific immunity and protects immunosuppressed BALB/c mice infected with virulent yeasts of Paracoccidioides brasiliensis. Mycopathologia. 2014;178(3–4):177–88.
Wong SSW et al. New insights into Cryptococcus extracellular vesicles suggest a new structural model and an antifungal vaccine strategy. bioRxiv, 2020.
Neuman BW, et al. Supramolecular architecture of severe acute respiratory syndrome coronavirus revealed by electron cryomicroscopy. J Virol. 2006;80(16):7918–28.
Zanetti G, et al. Cryo-electron tomographic structure of an immunodeficiency virus envelope complex in situ. PLoS Pathog. 2006;2(8):e83.
Van Dyke MCC, Wormley FL Jr. A call to arms: quest for a cryptococcal vaccine. Trends Microbiol. 2018;26(5):436–46.
Ueno K, et al. Vaccines and protective immune memory against cryptococcosis. Biol Pharm Bull. 2020;43(2):230–9.
Rella A, et al. Role of sterylglucosidase 1 (Sgl1) on the pathogenicity of Cryptococcus neoformans: potential applications for vaccine development. Front Microbiol. 2015;6:836.
Colombo AC et al. Cryptococcus neoformans glucuronoxylomannan and sterylglucoside are required for host protection in an animal vaccination model. MBio, 2019. 10(2).
Dufresne SF, et al. Detection of urinary excreted fungal galactomannan-like antigens for diagnosis of invasive aspergillosis. PLoS ONE. 2012;7(8):e42736.
Acknowledgements
Not applicable.
Funding
The efforts in preparing this review were supported by grants from the National Science Fund for Distinguished Young Scholars (82025024) and National Natural Science Foundation of China (82102459 and 82272384), China Postdoctoral Science Foundation (2019M662990), the President Foundation of Nanfang Hospital Southern Medical University (2021B023), Guangdong Basic and Applied Basic Research Foundation (2021A1515110578), Guangzhou Science and Technology Plan Project-Basic and Applied Basic Research Projects (202201011480), Clinical research subject of Guangdong Medical Association (yueyihui2020[342]).
Author information
Authors and Affiliations
Contributions
AU, YYH, QW, XMH and LZ, conceived the manuscript and supervised the whole project. AU and YYH drafted the manuscript and drew the figures. KNZ and YNH drafted and organized the tables. SU, MUR and JYW participated in manuscript revision and references organization.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of Interest
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ullah, A., Huang, Y., Zhao, K. et al. Characteristics and potential clinical applications of the extracellular vesicles of human pathogenic Fungi. BMC Microbiol 23, 227 (2023). https://doi.org/10.1186/s12866-023-02945-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-023-02945-3