
Abstract
Purpose
Previous studies have reported the potential impact of immune cells on kidney stone disease (KSD), but definitive causal relationships have yet to be established. The purpose of this paper is to elucidate the potential causal association between immune cells and KSD by Mendelian randomization (MR) analysis.
Methods
In our study, a thorough two-sample Mendelian randomization (MR) analysis was performed by us to determine the potential causal relationship between immune cell traits and kidney stone disease. We included a total of four immune traits (median fluorescence intensity (MFI), relative cellular (RC), absolute cellular (AC), and morphological parameters (MP)), which are publicly available data. GWAS summary data related to KSD (9713 cases and 366,693 controls) were obtained from the FinnGen consortium. The primary MR analysis method was Inverse variance weighted. Cochran’s Q test, MR Egger, and MR-Pleiotropy RESidual Sum and Outlier (MR-PRESSO) were used to assess the stability of the results.
Results
After FDR correction, the CD8 on HLA DR + CD8br (OR = 0.95, 95% CI = 0.93–0.98, p-value = 7.20 × 10− 4, q-value = 0.088) was determined to be distinctly associated with KSD, and we also found other 25 suggestive associations between immune cells and KSD, of which 13 associations were suggested as protective factors and 12 associations were suggested as risk factors. There was no horizontal pleiotropy or significant heterogeneity in our MR analysis, as determined by the p-value results of our Cochrane Q-test, MR Egger’s intercept test, and MR-PRESSO, which were all > 0.05.
Conclusions
Our study has explored the potential causal connection between immune cells and KSD by Mendelian randomization analysis, thus providing some insights for future clinical studies.
Similar content being viewed by others


Introduction
Kidney stone disease (KSD) is a common and frequent disease whose incidence is steadily increasing worldwide. It has been found that the prevalence of KSD in the United States is about 10% [1]. Not only that, the prevalence and recurrence of kidney stones in children are gradually increasing worldwide [2]. KSD has a high rate of recurrence, with approximately 50% recurrence within 5–10 years [3]. KSD is prone to a variety of complications, including pain, urinary tract obstruction, infection, and even increases the likelihood of chronic kidney disease, which may ultimately develop into end-stage renal disease [4, 5]. However, the risk factors influencing the development of KSD have not been fully investigated. Treatments such as medications and minimally invasive surgery can only remove formed stones and have little effect on reducing recurrence or early prevention of kidney stones. Previous studies have found a correlation between some immune cells and the development of KSD [6]. Paul R. Dominguez-Gutierrez et al. suggest that by modulating the immune response, immunotherapy may offer a way to prevent stone recurrence in some people [7]. Immunotherapy essentially enhances cellular immunity, regulates the degree of immune activation, and improves immune cell function, thereby inhibiting the occurrence and development of various diseases [8]. Personalized neoantigen vaccine is a novel immunotherapeutic approach that uses long synthetic peptides to induce neoantigen-specific T-cell responses in patients. The peptides directly initiate the immune system and have the advantage of simple and rapid production [9]. It has been shown that peptide vaccines targeting neoantigens can effectively induce T-cell responses [10]. Then, it is worth considering and exploring whether we can use personalized neoantigen vaccines to modulate the body’s immune cell response to achieve early prevention and treatment of KSD. Therefore, it is particularly significant to explore the role played by immune cells in the formation of kidney stones. However, due to the difficulties of observational studies and the limitations of analyzing the results, evidence of a potential association between immune cells and KSD is not readily available through this method and the results are prone to reverse causation. Thus, the potential causal relationship between immune cells and KSD needs to be explained by more precise research methods. Mendelian randomization (MR) is a genetic epidemiological method that uses genetic variants as instrumental variables (IVs) to assess the causality between risk factors and target diseases [11]. MR utilizes random assignment of single nucleotide polymorphisms (SNPs) in genetic variants to simulate randomized trials in populations, thus overcoming potential confounders and interferences of reverse causality [12]. It is very critical that the causal sequence of MR be rational [13, 14]. Therefore, we obtained summary statistics from large genome-wide association studies (GWAS) and revealed the causal relationship between immune cells and KSD by a two-sample MR analysis.
Materials and methods
Study design
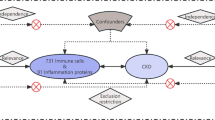
Our study followed the STROBE-MR statement used to report MR research [15]. No additional ethical approvals were required for this study, and the data we used and analyzed were published and publicly available data. An overview of the study design is shown in Fig. 1.

Overview and assumptions of the Mendelian randomization study design. Significant instrumental variables were selected for immune cells and KSD, and the causalities were then explored. Three basic assumptions of MR analysis were illustrated in this graph, namely, relevance (assumption 1), independence (assumption 2), and exclusion restrictions (assumption 3)
Genetic instrument selection
A summary of GWAS statistics for each immune trait is publicly available from the GWAS Catalog (accession numbers from GCST0001391 to GCST0002121) [16]. We included immunophenotypes that belonged to four different immune traits: absolute cell (AC) count, median fluorescence intensity (MFI), morphologic parameter (MP), and relative cell (RC) count. When categorized by cell panels, they belong to seven different panels, the Treg, TBNK (T cells, B cells, natural killer cells), B cells, monocytes, CDCs, myeloid cells, and mature stages of T cells. Initial immune traits GWAS was obtained from data on 3,757 Europeans and there were no overlapping cohorts. Approximately 22 million SNPs genotyped with high-density arrays were estimated using a reference panel based on Sardinian sequences and tested for correlation after adjusting for covariates [17].
According to recent research [16, 18], we set the significance level of IVs for each immune trait at 1 × 10− 5. Meanwhile, all SNPs were clumped according to linkage disequilibrium (LD), defined by r 2 < 0.1 with a clumping window > 500kb [19]. In this way, we could include more SNPs to ensure the stability of the results. We removed palindromic SNPs as well as SNPs that could not be found in the KSD data.
Data source for kidney stone disease
We obtained research data on kidney stone disease (KSD) from the FinnGen consortium, derived from the ninth release of the FinnGen consortium with 9,713 cases and 366,693 controls of kidney stone disease.
Statistical analysis
We first harmonized single nucleotide polymorphisms (SNPs) with the same allele, and then we performed two-sample MR analyses using inverse variance weighting (IVW), MR-Egger, and weighted median. The results primarily relied on the IVW method, with support from the other two methods. The MR-Egger method has the ability to tolerate potential pleiotropy and estimate results conservatively [20]. The weighted median method is a method that provides reliable estimates of causal effects and has the great advantage of allowing for 50% invalid IVs [21]. Also, we utilized the Wald ratio method to infer the effect of a single IV on KSD. We grouped the results of the IVW analysis according to the seven different cell panels, performed the false discovery rate (FDR) correction according to the groups, and compared the corrected p-value, also known as q-value, with the threshold we set at 0.1 [22]. We considered the association between immune cells and KSD to be suggestive if the p-value was < 0.05 but the q-value was ≥ 0.1, and significant if the q-value was < 0.1. We used Cochrane’s Q-test to assess heterogeneity, along with the MR-Egger intercept to assess pleiotropy. To assess whether the results were affected by outlier SNPs, we used leave-one-out analysis, MR-Pleiotropy Residual Sum and Outlier method (MR-PRESSO) to validate. We excluded results with heterogeneity and horizontal pleiotropy and retained results with the same direction under the three analyses of IVW, MR-Egger, and weighted median. All analyses were performed with the software packages TwoSampleMR (version 0.5.7) and MR-PRESSO (version 1.0) in R (version 4.3.0), and the correlation heat map was drawn using ChiPlot (https://www.chiplot.online/).
Results
Overview
Table 1 lists all the results that reached a suggestive significance (p-value < 0.05) for a total of 26 pairs between immune traits and KSD. When categorizing results based on the cell panels,12 traits belonged to the Treg, 4 from TBNK, 3 classified as monocyte, 1 from Maturation stages of T cell, 2 belonged to cDC, and 4 within B cell were found to be suggestively associated with KSD. We corrected the p-values in the different panels using the FDR method to reduce possible false positive results (Supplementary Table S1). After sensitivity analysis and multiple correction analysis (Supplementary Table S2), the CD8 on HLA DR + CD8br remained statistically significant (q-value < 0.1). Meanwhile, we categorized immune cells into seven sections according to the panel, grouped complex findings according to the panel, and found multiple suggestive associations between immune cells and KSD.
Treg and KSD
12 pairs between Treg/KSD reached suggestive association (p-value < 0.05) by using IVW MR analysis. RC had the greatest number of suggestive associations than other trait types. CD39 and CD28 were the most commonly expressed molecules in different types of immune traits. And, CD28 on CD39 + CD4 + was the most essential trait compared with other traits in this section (p-value = 3.70 × 10− 3). Furthermore, as shown in the scatter plot (Fig. 2), the traits (e.g., CD3 on CD39 + CD4+, CD25 on CD39 + CD4+) were positively associated with KSD (Fig. 2a–f), while the traits (e.g., CD39 + secreting Treg %CD4 Treg, CD39 + secreting Treg %secreting Treg) were negatively associated with KSD (Fig. 2g-l).

Scatter plot for the relationship between the SNP effect size of causal immune traits (x-axis) and the corresponding effect size estimates of kidney stone disease (KSD) (y-axis). The a ~ l plots all belong to the Treg group panel. (a) CD3 on CD39 + CD4 + on KSD, (b) CD25 on CD39 + CD4 + on KSD, (c) CD28- CD25 + + CD8br AC on KSD, (d) CD28- CD25 + + CD8br %T cell on KSD, (e) CD28 + CD45RA + CD8br %T cell on KSD, (f) CD28 + CD45RA + CD8dim %T cell on KSD, (g) CD39 + secreting Treg %CD4 Treg on KSD, (h) CD39 + secreting Treg %secreting Treg on KSD, (i) CD39 on CD39 + secreting Treg on KSD, (j) CD28 on CD39 + CD4 + on KSD, (k) CD39 + CD4 + AC on KSD, (l) CD25 + + CD8br %CD8br on KSD.
TBNK and KSD
Table 1 shows the suggestive significance of the 4 pairs between the TBNK panel and the KSD (p-value < 0.05). RC and MFI had the largest number of suggestive associations. After FDR correction, the CD8 on HLA DR + CD8br (OR = 0.95, 95% CI = 0.93–0.98, p-value = 7.20 × 10− 4, q-value = 0.088) had significant causal effects on KSD estimated from IVW (q-value < 0.1) (Supplementary Table S1). Moreover, for the most significant pair, no significant heterogeneity or horizontal pleiotropy was detected by sensitivity analysis (Supplementary Table S2), and the leave-one-out plot showed robust results for the relationship between CD8 on HLA DR + CD8br and KSD (Fig. 3g). We observed consistent correlation signals using other methods (Fig. 4) (Supplementary Table S3). As shown in Fig. 3, negative associations were found between CD8 on HLA DR + CD8br, HLA DR on HLA DR + CD8br, and KSD. On the other hand, the positive associations between CD3- lymphocyte %leukocyte, CD8br NKT %lymphocyte, and KSD were also detected. (Fig. 3a-d)

Scatter plot for the relationship between the SNP effect size of causal immune traits (x-axis) and the corresponding effect size estimates of kidney stone disease (KSD) (y-axis) and leave-one-out plot for the causal relationships between the TBNK group panel and KSD. The a ~ d scatter plots and e ~ h leave-one-out plots all belong to the TBNK group panel. (a) and (e) CD3- lymphocyte %leukocyte on KSD, (b) and (f) CD8br NKT %lymphocyte on KSD, (c) and (g) CD8 on HLA DR + CD8br on KSD, (d) and (h) HLA DR on HLA DR + CD8br on KSD.

Circle heatmap of the results of the MR analysis between the 26 immune traits that reached suggestive significance and the risk of KSD. The circle from the outer to the inner represented the weighted median, MR-Egger, and IVW, respectively
B cell, Monocyte, cDC, and KSD
A total of 4 B cell /KSD pairs, 3 Monocyte/KSD, and 2 cDC/KSD were detected at the suggestive level of p-value < 0.05(Table 1). Among them, the traits of MFI were particularly prominent, as a high percentage of suggestive significant pairs was detected up to 77.78% (7 MFI pairs vs. 9 total pairs). Meanwhile, 13 of the 26 pairs of results obtained were classified as belonging to MFI based on immune traits, which further suggested that immune cells belonging to MFI may play an important role in the development of KSD. It should be noted that BAFF-R on sw mem, which belongs to the B cell panel, was significantly associated with KSD after correction FDR correction (q-value = 0.099) (Supplementary Table S1). However, after the Cochrane Q-test and MR Egger’s intercept test, the results were of heterogeneity and pleiotropy, so we did not include it in the discussion.
Maturation stages of T cell, and KSD
Compared to the 12 pairs of results from Treg and KSD, only 1 pair between the Maturation stages of T cell /KSD reached suggestive association (p-value < 0.05) (Table 1). This suggests that Treg may have a more complex role in the progression of KSD compared to the Maturation stages of T cell.
Myeloid cell
Although five immune cell trait types in the Myeloid cell panel showed suggestive correlation with KSD, and one of them, CD33 on CD33dim HLA DR-, remained significantly correlated after FDR correction (Supplementary Table S1). Nevertheless, after the Cochrane Q-test and MR Egger’s intercept test, the results were of heterogeneity and pleiotropy, so we excluded the results in the Myeloid cell panel from the discussion.
Sensitive analysis
Cochran’s Q test, MR-Egger intercept test, and MR-PRESSO showed no heterogeneity or horizontal pleiotropy in the 26 pairs of results we retained in the analyses of immune cells and risk of KSD (all p-value > 0.05) (Supplementary Table S2). The leave-one-out plot and scatter plot showed that the results were robust.
Discussion
Despite the important role of the immune system in the formation of KSD, current research remains unclear as to its specific complex role in the progression of KSD. As far as we know, it’s the first time that the potential causal relationship between multiple types of immune cells and KSD has been explored by MR analytical methods. Our study found 12, 4, 4, 3, 2, 1, pairs between 6 panels of immune traits (i.e., Treg, TBNK, B cell, Monocyte, cDC, Maturation stages of T cell) and KSD were detected at the suggestive level of p-value < 0.05, respectively. Among them, the CD8 on HLA DR + CD8br/KSD reached the level of significant correlation (q-value < 0.1).
We found that the risk of KSD decreased as the proportion of the CD8 on HLA DR + CD8br increased. In the TBNK panel, T lymphocytes were split into six subsets based on the expression of the CD4 and CD8 markers: CD4-CD8- (DN), CD4-CD8dim (CD8dim), CD4-CD8bright (CD8br), CD4 + CD8br (DP), CD4 + CD8dim, CD4 + CD8- (CD4+) [16]. The HLA DR positivity of CD4 + and CD8br T cells and NK cells was considered as an activation marker [16].
CD8 on HLA DR + CD8br, which can be interpreted as a CD4-CD8bright (CD8br) subset of T lymphocytes in the HLA-DR activated state. CD8 is a heterodimer composed of α and β peptide chains, and its main function is to assist in the process of antigen recognition by immune cells and to participate in the activation of T cells. signals, which can enhance the interaction between T-cells and APCs or target cells [23]. CD8br cells are limited by their own MHC class I molecules and differentiate into cytotoxic T cells (CTLs) with cytotoxicity after activation, which can specifically kill target cells. Antigens or cytokines activate these cells, and activated cells induce apoptosis through two primary pathways: lytic (cytotoxicity) and non-lytic (cytokine production) mechanisms respectively [24]. The lysis process mainly utilizes secreted particles containing effector substances, such as granzyme, perforin, etc. These particles are released to act directly on the target cells, thereby killing them. Non-lytic mechanisms mainly refer to the secretion of cytokines such as IFN-γ and TNF-α, which are essential for inflammatory responses [25]. Meanwhile, CD8 + T cells in infectious diseases have been reported to produce cytokines to act on themselves, thereby enhancing their cytotoxic activity upon activation [26, 27].
An association between kidney stone formation and urinary tract infections has been reported in certain patient groups [28, 29]. Localized infection in the urinary tract is one of the factors that trigger stone formation, and the stones themselves are foreign bodies that can cause obstruction in the urinary tract, which can exacerbate the degree of infection, and then enter a vicious circle [30]. Urease-producing bacteria such as Proteus mirabilis, Klebsiella pneumoniae, Staphylococcus aureus, and Pseudomonas aeruginosa are invariably associated with the formation and recurrence of struvite calculi [31]. These bacteria split urea and promote the formation of ammonia and carbon dioxide, leading to renal tubular damage and alkalinization of the urine with subsequent formation of phosphate and struvite calculi [31]. Also, de Cógáin et al. described secondary infection stones caused by non-urease-producing bacteria, including Escherichia coli and Enterococcus spp [32, 33]. During the formation of kidney stones, cytotoxic T cells are able to directly kill the bacteria that cause urinary tract infections. They do this by releasing cytotoxins, such as granzymes and perforins, which disrupt the cell membranes of the pathogens, thereby triggering the death of the bacteria. This direct killing effect can effectively control and clear the infection, preventing it from spreading further and leading to the deterioration of kidney stones. Using immunoinformatics and structural vaccinology, Suar et al. designed multi-epitope peptide vaccines against Klebsiella pneumoniae [34], Enterococcus faecalis [35], Staphylococcus aureus [36, 37], and Acinetobacter baumannii [38, 39] with highly immunogenic cytotoxic T-lymphocyte epitopes and high binding affinities for most human leukocyte antigen (HLA). This offers feasibility and credibility for our future utilization of vaccines to modulate immune responses, such as T-cell activation, to target bacteria during kidney stone formation and thereby prevent KSD.
In the early stages of kidney stone formation, the primary role of CD8 + T cells is to activate and direct the involvement of other immune cells in the immune response. They are able to secrete a range of cytokines, such as IFN-γ, and TNF-α, to promote an inflammatory response. Cytokines are a class of proteins that regulate the function of immune cells and participate in inflammatory responses and cell proliferation [40], and they play an important role in kidney stone formation [6]. TNF-α, upon stone stimulation of renal tubular epithelial cells, leads to aggregation of immune cells and induces an inflammatory response [6], thereby further damaging renal tissue. CD8 + T cells are also able to interact with other types of immune cells, such as macrophages and B-cells, to regulate the extent and direction of the immune response through the mediation of cytokines [41]. This immunomodulatory effect can limit excessive inflammatory responses, reduce kidney stone damage to kidney tissue, and aid in stone expulsion and removal. These findings are consistent with those of Noble et al. They found that CD8 + T cells can secrete anti-inflammatory cytokines with inhibitory activity, such as IL-10 [42]. Anti-inflammatory cytokines (e.g., IL-10) inhibit the inflammatory response, while it has been suggested that exogenous IL-10 treatment can reverse oxalate-mediated outcomes in vitro [43, 44]. Meanwhile, IL-10 was identified as a urinary tract inflammation-associated factor that can accurately differentiate between control individuals and patients with urinary tract stones [45]. Equally important, IL-10 has been reported to be essential for the induction of M2 macrophage polarization [46]. M2 anti-inflammatory macrophages have been found to phagocytose and degrade CaOx kidney stone fragments [47]. Because the M2 phenotype is more able to phagocytose crystals than the M1 phenotype [48], regulating the direction of their polarization may have therapeutic value [49]. Taguchi, K et al. found that M1Mϕ infusion and LPS and IFN-γ induction promoted renal crystal formation, whereas M2Mϕ infusion and IL-4 and IL-13 induction inhibited renal crystal formation [48]. Zhu et al. also modulated macrophage CSF-1 expression by inhibiting androgen receptor, thereby reducing crystal deposition in the kidney by altering macrophage recruitment/M2 polarization [50]. These experimental pieces of evidence provide us with the possibility of utilizing the cytokines secreted by CD8 + T cells to regulate the polarization of macrophages for the treatment of kidney stones. The combination of effective molecular adjuvants can cause massive expansion of antigen-specific CD8 T cells and show previously unseen efficacy in vaccine and tumor models [51], so whether we can utilize them for the treatment of KSD deserves to be explored further in the future.
Our findings may suggest that immune cells have a complex role in the pathogenesis and progression of KSD. These new findings may contribute to a more comprehensive understanding of immune cells in KSD and provide new potential biomarkers for early diagnosis and monitoring of KSD. Meanwhile, these findings also raise the question of what role these immune cells could play in KSD prevention and treatment. Currently, the recurrence rate of KSD after surgery remains high due to individual differences and environmental factors [52], and the damage to the kidney from repeated surgeries can easily lead to life-threatening renal failure. In addition, there is a lack of effective drugs for the treatment of kidney stones. Then immunotherapy based on the results of our findings may provide new ideas for the prevention and treatment of kidney stones. The application of immunotherapy has revolutionized cancer treatment by harnessing the body’s immune system to work against cancer and significantly improve patient prognosis [53]. Based on our findings and what has been discussed above, we propose the following hypothesis for the application of immunotherapy against KSD: firstly, we can design a multiple-epitope peptide vaccine that induces a CD8 + T-cell immune response to directly kill bacteria in infected kidney stones. Secondly, the infusion of exogenous cytokines can be utilized to promote the polarization of M2 macrophages and reduce renal crystal deposition. Finally, combined vaccine molecular adjuvants can be designed to induce the expansion of CD8 + T cells. Then exploring the specific relationship between the interactions of these immune cells so as to accurately regulate the immune microenvironment of the renal mesenchyme and thus intervene in kidney stone formation by utilizing immunotherapeutic methods will be the focus of future research.
This study was the result of a two-sample MR analysis based on published results from a large genomic cohort study and was therefore statistically valid. The findings of this study were derived from causal inference utilizing various MR analysis methods with genetic instruments as variables. These results are reliable, robust, and not confounded by horizontal pleiotropy and other factors. However, there are still some limiting parts of our study. First, even though we performed multiple sensitivity analyses, we were unable to fully assess the horizontal pleiotropy effect. Second, the data in this study comes from the European population, so it is worth considering whether the results can be generalized to other races, which limits the generalizability of the application of the results. Third, the lack of individual-specific information prevented us from further stratifying the population. Finally, in order to be able to more fully explore the relationship between immune cells and kidney stones, we used a more relaxed threshold to assess the results, but this may have increased some false positives.
Conclusion
In conclusion, we fully utilized the bidirectional MR analysis to investigate the potential relationship between immune cells and KSD, giving a little insight to researchers exploring the complex interactions between the immune system and KSD. And based on the results of the study, we propose hypotheses for future applications and future priorities that still need to be explored further. Moreover, our study reduces the potential for reverse causality as well as multiple unavoidable confounding factors in research, which may offer a novel approach for scientific researchers to continue exploring the Immunological mechanisms of KSD.
Data availability
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Abbreviations
- KSD:
-
kidney stone disease
- MFI:
-
median fluorescence intensity
- RC:
-
relative cellular
- AC:
-
absolute cellular
- MP:
-
morphological parameters
- GWAS:
-
genome-wide association studies
- IV:
-
instrumental variable
- IVW:
-
inverse variance weighted
- MR:
-
Mendelian randomization
- MR-PRESSO:
-
MR-Pleiotropy Residual Sum and Outlier method
- OR:
-
odds ratio
- SNP:
-
single nucleotide polymorphism
- CTLs:
-
cytotoxic T cells
- IL-10:
-
Interleukin-10
- IL-4:
-
Interleukin-4
- IL-13:
-
Interleukin-13
- IFN-γ:
-
Interferon-gamma
- LPS:
-
Lipopolysaccharides
References
Chewcharat A, Curhan G. Trends in the prevalence of kidney stones in the United States from 2007 to 2016. Urolithiasis. 2021;49:27–39.
Tasian GE, Copelovitch L. Evaluation and medical management of kidney stones in children. J Urol. 2014;192:1329–36.
Rule AD, Lieske JC, Li X, Melton LJ, Krambeck AE, Bergstralh EJ. The ROKS nomogram for predicting a second symptomatic stone episode. J Am Soc Nephrol JASN. 2014;25:2878–86.
Zhe M, Hang Z. Nephrolithiasis as a risk factor of chronic kidney disease: a meta-analysis of cohort studies with 4,770,691 participants. Urolithiasis. 2017;45:441–8.
Rule AD, Lieske JC, Pais VM. Management of kidney stones in 2020. JAMA. 2020;323:1961–2.
Taguchi K, Okada A, Unno R, Hamamoto S, Yasui T. Macrophage function in calcium oxalate kidney stone formation: a systematic review of literature. Front Immunol. 2021;12:673690.
Dominguez-Gutierrez PR, Kwenda EP, Khan SR, Canales BK. Immunotherapy for stone disease. Curr Opin Urol. 2020;30:183–9.
Shen K-Y, Zhu Y, Xie S-Z, Qin L-X. Immunosuppressive tumor microenvironment and immunotherapy of hepatocellular carcinoma: current status and prospectives. J Hematol OncolJ Hematol Oncol. 2024;17:25.
Mørk SK, Skadborg SK, Albieri B, Draghi A, Bol K, Kadivar M, et al. Dose escalation study of a personalized peptide-based neoantigen vaccine (EVX-01) in patients with metastatic melanoma. J Immunother Cancer. 2024;12:e008817.
Mørk SK, Kadivar M, Bol KF, Draghi A, Westergaard MCW, Skadborg SK, et al. Personalized therapy with peptide-based neoantigen vaccine (EVX-01) including a novel adjuvant, CAF®09b, in patients with metastatic melanoma. Oncoimmunology. 2022;11:2023255.
Lawlor DA, Harbord RM, Sterne JAC, Timpson N, Davey Smith G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008;27:1133–63.
Birney E. Mendelian randomization. Cold Spring Harb Perspect Med. 2022;12:a041302.
Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014;23:R89–98.
Timpson NJ, Wade KH, Smith GD. Mendelian randomization: application to cardiovascular disease. Curr Hypertens Rep. 2012;14:29–37.
Skrivankova VW, Richmond RC, Woolf BAR, Yarmolinsky J, Davies NM, Swanson SA, et al. Strengthening the reporting of Observational studies in Epidemiology using mendelian randomization: the STROBE-MR Statement. JAMA. 2021;326:1614–21.
Orrù V, Steri M, Sidore C, Marongiu M, Serra V, Olla S, et al. Complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nat Genet. 2020;52:1036–45.
Sidore C, Busonero F, Maschio A, Porcu E, Naitza S, Zoledziewska M, et al. Genome sequencing elucidates sardinian genetic architecture and augments association analyses for lipid and blood inflammatory markers. Nat Genet. 2015;47:1272–81.
Yu X-H, Yang Y-Q, Cao R-R, Bo L, Lei S-F. The causal role of gut microbiota in development of osteoarthritis. Osteoarthritis Cartilage. 2021;29:1741–50.
1000 Genomes Project Consortium, Auton A, Brooks LD, Durbin RM, Garrison EP, Kang HM, et al. A global reference for human genetic variation. Nature. 2015;526:68–74.
Burgess S, Thompson SG. Interpreting findings from mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32:377–89.
Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some Invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40:304–14.
Storey JD, Tibshirani R. Statistical significance for genomewide studies. Proc Natl Acad Sci U S A. 2003;100:9440–5.
Cole DK, Gao GF. CD8: adhesion molecule, co-receptor and immuno-modulator. Cell Mol Immunol. 2004;1:81–8.
Gonzalez SM, Taborda NA, Rugeles MT. Role of different subpopulations of CD8 + T cells during HIV exposure and infection. Front Immunol. 2017;8:936.
Perdomo-Celis F, Velilla PA, Taborda NA, Rugeles MT. An altered cytotoxic program of CD8 + T-cells in HIV-infected patients despite HAART-induced viral suppression. PLoS ONE. 2019;14:e0210540.
Xing L, Liu C, Fu R, Wang H, Wang J, Liu X, et al. CD8 + HLA-DR + T cells are increased in patients with severe aplastic anemia. Mol Med Rep. 2014;10:1252–8.
Schmidt ME, Knudson CJ, Hartwig SM, Pewe LL, Meyerholz DK, Langlois RA, et al. Memory CD8 T cells mediate severe immunopathology following respiratory syncytial virus infection. PLoS Pathog. 2018;14:e1006810.
Tavichakorntrakool R, Prasongwattana V, Sungkeeree S, Saisud P, Sribenjalux P, Pimratana C, et al. Extensive characterizations of bacteria isolated from catheterized urine and stone matrices in patients with nephrolithiasis. Nephrol Dial Transplant off Publ Eur Dial Transpl Assoc -. Eur Ren Assoc. 2012;27:4125–30.
Borghi L, Nouvenne A, Meschi T. Nephrolithiasis and urinary tract infections: the chicken or the egg dilemma? Nephrol Dial Transplant off Publ Eur Dial Transpl Assoc -. Eur Ren Assoc. 2012;27:3982–4.
Ripa F, Pietropaolo A, Montanari E, Hameed BMZ, Gauhar V, Somani BK. Association of Kidney Stones and recurrent UTIs: the Chicken and Egg Situation. A systematic review of literature. Curr Urol Rep. 2022;23:165–74.
Espinosa-Ortiz EJ, Eisner BH, Lange D, Gerlach R. Current insights into the mechanisms and management of infection stones. Nat Rev Urol. 2019;16:35–53.
de Cógáin MR, Lieske JC, Vrtiska TJ, Tosh PK, Krambeck AE. Secondarily infected nonstruvite urolithiasis: a prospective evaluation. Urology. 2014;84:1295–300.
Flannigan R, Choy WH, Chew B, Lange D. Renal struvite stones–pathogenesis, microbiology, and management strategies. Nat Rev Urol. 2014;11:333–41.
Dey J, Mahapatra SR, Lata S, Patro S, Misra N, Suar M. Exploring Klebsiella pneumoniae capsule polysaccharide proteins to design multiepitope subunit vaccine to fight against pneumonia. Expert Rev Vaccines. 2022;21:569–87.
Dey J, Mahapatra SR, Raj TK, Kaur T, Jain P, Tiwari A, et al. Designing a novel multi-epitope vaccine to evoke a robust immune response against pathogenic multidrug-resistant Enterococcus faecium bacterium. Gut Pathog. 2022;14:21.
Chatterjee R, Mahapatra SR, Dey J, Raj Takur K, Raina V, Misra N, et al. An immunoinformatics and structural vaccinology study to design a multi-epitope vaccine against Staphylococcus aureus infection. J Mol Recognit JMR. 2023;36:e3007.
Mahapatra SR, Dey J, Raj TK, Misra N, Suar M. Designing a Next-Generation Multiepitope-based vaccine against Staphylococcus aureus using reverse vaccinology approaches. Pathog Basel Switz. 2023;12:376.
Dey J, Mahapatra SR, Singh PK, Prabhuswamimath SC, Misra N, Suar M. Designing of multi-epitope peptide vaccine against Acinetobacter baumannii through combined immunoinformatics and protein interaction–based approaches. Immunol Res. 2023;71:639–62.
Mahapatra SR, Dey J, Jaiswal A, Roy R, Misra N, Suar M. Immunoinformatics-guided designing of epitope-based subunit vaccine from Pilus assembly protein of Acinetobacter baumannii bacteria. J Immunol Methods. 2022;508:113325.
Liu C, Chu D, Kalantar-Zadeh K, George J, Young HA, Liu G. Cytokines: from clinical significance to quantification. Adv Sci Weinh Baden-Wurtt Ger. 2021;8:e2004433.
Klein Geltink RI, Kyle RL, Pearce EL. Unraveling the Complex Interplay between T Cell metabolism and function. Annu Rev Immunol. 2018;36:461–88.
Noble A, Giorgini A, Leggat JA. Cytokine-induced IL-10-secreting CD8 T cells represent a phenotypically distinct suppressor T-cell lineage. Blood. 2006;107:4475–83.
Atri C, Guerfali FZ, Laouini D. Role of human macrophage polarization in inflammation during infectious diseases. Int J Mol Sci. 2018;19:1801.
Kumar P, Laurence E, Crossman DK, Assimos DG, Murphy MP, Mitchell T. Oxalate disrupts monocyte and macrophage cellular function via Interleukin-10 and mitochondrial reactive oxygen species (ROS) signaling. Redox Biol. 2023;67:102919.
Okada A, Ando R, Taguchi K, Hamamoto S, Unno R, Sugino T, et al. Identification of new urinary risk markers for urinary stones using a logistic model and multinomial logit model. Clin Exp Nephrol. 2019;23:710–6.
Lopes RL, Borges TJ, Zanin RF, Bonorino C. IL-10 is required for polarization of macrophages to M2-like phenotype by mycobacterial DnaK (heat shock protein 70). Cytokine. 2016;85:123–9.
Xu Z, Yao X, Duan C, Liu H, Xu H. Metabolic changes in kidney stone disease. Front Immunol. 2023;14:1142207.
Taguchi K, Okada A, Hamamoto S, Unno R, Moritoki Y, Ando R, et al. M1/M2-macrophage phenotypes regulate renal calcium oxalate crystal development. Sci Rep. 2016;6:35167.
Chen T, Cao Q, Wang Y, Harris DCH. M2 macrophages in kidney disease: biology, therapies, and perspectives. Kidney Int. 2019;95:760–73.
Zhu W, Zhao Z, Chou F, Zuo L, Liu T, Yeh S, et al. Loss of the androgen receptor suppresses intrarenal calcium oxalate crystals deposition via altering macrophage recruitment/M2 polarization with change of the miR-185-5p/CSF-1 signals. Cell Death Dis. 2019;10:1–19.
Kornbluth RS, Stone GW. Immunostimulatory combinations: designing the next generation of vaccine adjuvants. J Leukoc Biol. 2006;80:1084–102.
Daudon M, Jungers P, Bazin D, Williams JC. Recurrence rates of urinary calculi according to stone composition and morphology. Urolithiasis. 2018;46:459–70.
Mitra A, Kumar A, Amdare NP, Pathak R. Current Landscape of Cancer Immunotherapy: harnessing the Immune Arsenal to Overcome Immune Evasion. Biology. 2024;13:307.
Acknowledgements
we are grateful to all the investigators and participants in a longitudinal study (SardiNIA project) for SardiNIA dataset and their contribution to those studies. Genetic association estimates for kidney stone disease were obtained from the FinnGen consortium (www.finngen.fi/en). We thank all investigators for sharing these data.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 81974092).
Author information
Authors and Affiliations
Contributions
D.F. Yuan and J.Y. Yang prepared and drafted the manuscript. X. Yu obtained funding for the study and provided critical revision of the manuscript for important intellectual content. W.S. Wu, Y. Amier, X.M. Li, and W.L. Wan assisted in obtaining data for the review article and revised the manuscript. Y.S. Huang and J.B. Li, confirmed the authenticity of all the raw data. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Our analysis used publicly available GWAS summary statistics. No new data were collected, and no new ethical approval was required.
Consent for publication
Not applicable’ for that section.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yuan, D., Yang, J., Wu, W. et al. The immune factors have complex causal regulation effects on kidney stone disease: a mendelian randomization study. BMC Immunol 25, 34 (2024). https://doi.org/10.1186/s12865-024-00627-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12865-024-00627-x