
Abstract
Background
The re-evolution of complex characters is generally considered impossible, yet, studies of recent years have provided several examples of phenotypic reversals shown to violate Dollo’s law. Along these lines, the regain of wings in stick and leaf insects (Phasmatodea) was hypothesised to have occurred several times independently after an ancestral loss, a scenario controversially discussed among evolutionary biologists due to overestimation of the potential for trait reacquisition as well as to the lack of taxonomic data.
Results
We revisited the recovery of wings by reconstructing a phylogeny based on a comprehensive taxon sample of over 500 representative phasmatodean species to infer the evolutionary history of wings. We additionally explored the presence of ocelli, the photoreceptive organs used for flight stabilisation in winged insects, which might provide further information for interpreting flight evolution. Our findings support an ancestral loss of wings and that the ancestors of most major lineages were wingless. While the evolution of ocelli was estimated to be dependent on the presence of (fully-developed) wings, ocelli are nevertheless absent in the majority of all examined winged species and only appear in the members of few subordinate clades, albeit winged and volant taxa are found in every euphasmatodean lineage.
Conclusion
In this study, we explored the evolutionary history of wings in Phasmatodea and demonstrate that the disjunct distribution of ocelli substantiates the hypothesis on their regain and thus on trait reacquisition in general. Evidence from the fossil record as well as future studies focussing on the underlying genetic mechanisms are needed to validate our findings and to further assess the evolutionary process of phenotypic reversals.
Similar content being viewed by others


Background
Active flight is considered the key innovation to have driven lineage diversification in animals and has allowed insects to become the most species-rich group on Earth [1,2,3]. The evolution of the insect wing ~ 400 million years ago (mya) [2, 4] and the associated enhanced abilities to disperse and access unreachable resources are directly linked to the remarkable radiation and success of winged insects (Pterygota). Moreover, wings have undergone various modifications to additionally or alternatively serve other functions such as thermoregulation, mate choice and courtship, crypsis and defensive strategies [1, 5]. However, despite the numerous advantages, wings have been repeatedly reduced across all pterygote groups with several lineages being completely wingless such as heel-walkers (Mantophasmatodea), lice (Phthiraptera) or fleas (Siphonaptera) [1, 6,7,8]. Paradoxically, also the loss of flight is proposed to be in direct correlation with increased diversification rates and thus also a driver of speciation [9,10,11,12].
Flightlessness appears to occur when the selection for aerial dispersal is relaxed as in habitats with environmental stability or with unfavourable conditions for flight such as high winds or cold temperatures [6, 13, 14]. As a consequence, flightless taxa may show increased fecundity because resources previously invested in maintaining the energetically costly flight apparatus can now be allocated to reproduction [6, 15, 16]. The underlying trade-off between dispersal and reproduction has been repeatedly demonstrated for females [17,18,19,20,21] and males [22,23,24], albeit the loss of flight is generally more common in females, often resulting in wing dimorphic species with volant males [1, 6, 25]. In some species, the dilemma of dispersal capability versus fecundity is solved by wing polyphenism, where an either winged or flightless phenotype is adopted in response to specific environmental triggers [1, 16, 18, 26,27,28,29]. Yet, flightlessness is not tantamount to the complete loss of wings (aptery): A phenotype or species may exhibit shortened wing length or retain fully-sized wings but with reduced flight musculature [26, 27].
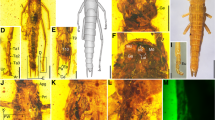
Within winged insects, the plant-mimicking lineage of stick and leaf insects (Phasmatodea) has been recognised as an expedient study system to investigate the evolution of flight due to their high diversity in wing states and sizes varying among closely related species and between sexes [30, 31] (Fig. 1). Generally, forewings are modified to abbreviated and sclerotised tegmina or wing pads, while the hindwings are membranous and folded neatly against the elongated and often slender body at rest [2]. However, most of these herbivorous insects are flightless [1, 30], being either completely wingless (apterous; Fig. 1A, B) or short-winged (micropterous/brachypterous; Fig. 1C–F), while long-winged forms (macropterous; Fig. 1G, H) may or may not be capable of ascending flight. Wings that do not sustain powered flight may serve for gliding or other derived utilities such as defensive stridulation or startle displays [32,33,34]. The latter is common in both short- and long-winged species whose wing undersides or membranes show bright warning colours in contrast to the otherwise inconspicuously coloured body maintaining crypsis (Fig. 1E–H). Sexual wing dimorphism occurs throughout the phasmatodean lineage and is exclusively female-biased [30, 31] (Fig. 1I) with the exception of two Phasmotaenia species [35]. As a result, females tend to be larger and brachypterous or apterous, whereas males are smaller and either share the female’s wing state or have more developed wings.

Photographs of various phasmatodean representatives with different wing states. Taxa may be wingless (apterous), short-winged and flightless (micropterous/brachypterous) or long-winged and presumably volant (macropterous). A Wingless couple of Oreophoetes peruana. B Wingless couple of Eurycantha insularis. C Female of Pseudodiacantha macklotti (brachypterous). D Female of Phaenopharos khaoyaiensis (micropterous/brachypterous). E and F Close-up of winglets of Phaenopharos sp. (micropterous/brachypterous). The conspicuous colouration is only visible when wings are opened presenting a startle display. G Female of Anarchodes annulipes (macropterous). The wing membranes exhibit a warning colouration to be used in startle displays. H Female of Metriophasma diocles (macropterous). The opened wings show the long hindwing and the for phasmatodeans typical shortened forewing. l Sexual size and wing dimorphism in a couple of Extatosoma tiaratum with brachypterous female on the left and macropterous male on the right. Photos by Bruno Kneubühler and Christoph Seiler
In contrast to the diverse wing states of extant stick and leaf insects, extinct species of stem group phasmatodeans have fully-developed fore- and hindwings [36,37,38] suggesting multiple and rather recent shifts from fully-winged to short-winged and wingless forms [31]. Yet, these transition events might not be unidirectional. Whiting et al. [30] proposed that wings had been completely lost in ancestral stick insects and were independently re-acquired in several descendant phasmatodean lineages. This hypothesis had been extensively debated and criticised as an overstatement in regard to the reliability of the inferred topology and the overall probability of wing regain [39, 40], and was largely considered a violation of Dollo’s law under which the loss of complex traits is irreversible (see Gould [41], but also Collin and Miglietta [42] and Goldberg and Igić [43]). Discussing the validity of Dollo’s law itself, Collin and Miglietta [42] cited Simpson [44] to have formulated its modern version, however, Simpson explicitly stated that Dollo’s law must not be confined to the “trivial case of reappearance of lost organs” [44]. Based on Dollo’s assumption, the evolution of a complex trait involves so many contingent steps that any transition in reverse (to reacquaint an ancestral state exactly) becomes effectively impossible [45]. An updated version of this principle had also been framed by Dawkins [46] who considered Dollo’s law to be a statement about the statistical improbability for a larger number of mutational steps to follow exactly the same evolutionary trajectory twice (in either direction). Wing recurrence after ancestral lost might in fact represent such a “trivial case” and was already suggested for water striders and fig wasps [47, 48] as was the re-evolution of various other traits such as eyes, digits or teeth in other animals [49,50,51,52,53]. In corroboration with Whiting et al.’s [30] results, a recent work on phasmatodean wing evolution also concluded a wingless or at least flightless ancestral state for stick and leaf insects [54]. However, both studies’ topologies do not comply with other comprehensive phylogenetic analyses based on larger datasets in terms of taxa [55] and genes [56, 57] and thus may be not reliable. Ultimately, only a robust phylogeny can provide the framework necessary for understanding the evolutionary processes underlying character evolution.
In addition to the flight apparatus itself, the specific nature of sensory systems regulating stabilisation reflexes for maintaining balance during flight such as wind-sensitive hairs [58] and ocelli [59] might provide further information for interpreting flight evolution. Pterygote insects generally possess a set of three ocelli (single-lens eyes) besides the facetted compound eyes. These photoreceptive organs process information on light levels more rapidly than compound eyes and thus significantly contribute to horizon detection and orientation in three-dimensional space during flight [59,60,61,62,63], but have also been suggested to be involved in other functions such as, for instance, the circadian rhythm (see Honkanen et al. [64], Ribi and Zeil [65] and references therein). The number and organisation of ocelli and their photoreceptors varies across all insect lineages and may be indicative of the adaptation to different selection pressures [65]. While ocelli are generally lacking in secondarily apterous pterygote insects [61], they may also be absent in lineages of winged taxa such as webspinners [66] or beetles [67]. In stick and leaf insects, ocelli appear to be associated with the capability of flight and never occur in wingless taxa [68, 69] (Fig. 2). Interestingly, numerous winged and volant species lack ocelli nevertheless (Fig. 2A). In species that possess ocelli, the degree of their development may reflect whether the species is partially- or fully-winged (Fig. 2E–G). Even within a single species they can be well-developed in the volant male, while completely absent in the flightless female (Fig. 2H–K). Given the assumed strong correlation between flight capability and presence of ocelli, the ocellar system might contribute to differentiate between primarily and secondarily winged stick insects.

Photographs showing the presence/absence of ocelli in taxa with different wing states. Arrows point to ocelli. A Macropterous female of Aschiphasma annulipes (Aschiphasmatidae). B Macropterous female of Anarchodes annulipes (Necrosciinae). C Macropterous male of Acrophylla titan (Lanceocercata). D Macropterous male of Xeroderus sp. (Lanceocercata). E Micropterous female of Peruphasma schultei (Pseudophasmatidae). F Micropterous female of Pseudophasma scabriusculum (Pseudophasmatidae). G Macropterous female of Pseudophasma fulvum (Pseudophasmatidae). H–K Sexual dimorphism in (H) micropterous female and (I) macropterous male of Phyllium philippinicum (Phylliidae), and in (J) micropterous female and (K) macropterous male of Extatosoma tiaratum (Lanceocercata). Scale bars: 1 mm. Photos by Tim Lütkemeyer and Marco Niekampf
Herein, we revisit the evolution of flight in stick and leaf insects by inferring a phylogenetic reconstruction of wings and ocelli based on a set of seven nuclear and mitochondrial genes for the largest taxon sample to date with > 500 species covering all major groups. This extremely dense taxonomic resolution appears to be necessary to satisfactorily reconstruct this highly disparate character system.
Results
Phylogenetic relationships and divergence times of major phasmatodean lineages
Our comprehensive taxon sample of 513 phasmatodean taxa and two outgroup species of Embioptera provided an optimal basis for exploring the phylogenetic relationships of Phasmatodea. All Maximum Likelihood (ML) and Bayesian (BI) analyses recovered highly congruent topologies with moderate to high support across the backbone nodes and for all the major lineages (Additional file 1: Figs. S1–S4, Additional file 2: Files S1–S5). The application of different constraints had minimal effect on the topologies, and discrepancies largely pertained to the same groups such as the European Bacillinae sensu Cliquennois [70] and the African/Malagasy clade. When there were no constraints within Neophasmatodea (B3; Additional file 1: Fig. S3), the leaf insects (Phylliidae) were recovered as sister group to a clade of Occidophasmata + the remaining Oriophasmata, whereas the other inferences recovered them as closely related to the African/Malagasy group. Generally, the Occidophasmata are split in Agathemeridae + Pseudophasmatidae and Heteronemiinae + Diapheromerinae, and only in the ML tree Paraprisopus is recovered as sister taxon to Agathemera and not clustered within Pseudophasmatidae (Additional file 1: Fig. S1). The phylogenetic relationships within Oriophasmata vary slightly depending on the constraints. The most strict constraints forced the Heteropterygidae to form the sole sister group to the remaining Oriophasmata (B2; Additional file 1: Fig. S4), albeit the analyses without this constraint recovered them as sister taxon to a clade including Clitumninae sensu Cliquennois [70] (= Clitumnini, Gratidiini, Medaurini), European Bacillinae sensu Cliquennois [70] and African Gratidiidae sensu Cliquennois [70]. Yet, the B2 constraint resulted in the overall highest nodal support values. The remaining clade of Oriophasmata appears generally more congruent, especially in regard to the Lonchodidae (Lonchodinae + Necrosciinae) as sister to (Pharnaciinae + Prosentoria) + ((Palophidae + Cladomorphinae) + (Stephanacridini + Xenophasmina + Lanceocercata)).
The selection of fossils to use as calibration age priors was identical for all three BI trees (Additional file 3: Table S9) and the resulting divergence time estimates are consistent among the analyses, with slightly older estimates for the B2 constrained tree (Additional file 2: Files S3–S5). Our results estimate Phasmatodea to have originated in the Jurassic ~ 178.56 million years ago (mya) (186.6–165.14 mya) (B2), whereas the diversification of Euphasmatodea and Neophasmatodea started in the Cretaceous ~ 106.13 mya (114.16–75.11 mya) and ~ 96.29 mya (141.77–63.23 mya), respectively. However, the divergence of most of the major lineages does not predate the Cretaceous-Palaeogene (K-Pg) boundary and which thus have their origin in the Early Cenozoic (Palaeocene).
Phylogenetic signal and ancestral states reconstruction
Morphological data on wings and ocelli were collected for 513 phasmatodean specimens, albeit 18 are only known from one sex (nine females and males, respectively). The majority is found to be apterous with 272 taxa lacking wings entirely and 45 with wingless females (Fig. 3). Among winged species, brachyptery is found to be more common in females, whereas males predominantly have fully-developed wings. Ocelli never occur in wingless species and rarely in micropterous taxa (3 of 66 males, 7 of 112 females). Although mostly present in macropterous species, less than half of the examined macropterous females and males possess ocelli. In general, ocelli were found to occur in five of the major lineages, namely, Lanceocercata, Necrosciinae, Pseudophasmatidae, Palophidae and Phylliidae, and females with ocelli exist only in the former three groups.

Results of morphological examination. Observed number of wingless (apterous), partially-winged (micropterous) and fully-winged (macropterous) male and female phasmatodeans and proportion of taxa with and without ocelli
A strong phylogenetic signal was detected for all tested traits (presence of ocelli, presence of wings, capability of flight) with negative D statistics (D = −0.568, −0.410 and −0.355, respectively) and fitted lambda values of > 0.981 (Additional file 3: Table S1) indicating that these traits are more likely to be shared by closely related species and thus phylogenetically conserved. This was also supported by comparing the numbers of evolutionary transitions observed for each trait (24, 58 and 51) against a randomisation process of that trait (on average 70, 169 and 142; Additional file 3: Table S1).
Prior to reconstructing ancestral states, we fitted different models to the binary wing dataset (datawings) and compared log-likelihoods and AIC values (Additional file 3: Table S2). The ARD model was determined as the best-fit model independent of whether the root state was equally probable to be winged/wingless or forced to be wingless. By contrast, the models disallowing the regain of wings (IRR) performed the poorest, even when the root was forced to be winged. Applying the ARD model, the ancestral reconstruction of wings and ocelli estimated the ancestral state for all the major nodes to be wingless and without ocelli for males and females (Fig. 4, Additional file 1: Figs. S4, S5). When regarding the African/Malagasy stick insects as two separate clades, we recognise 21 major lineages of which 14 have winged representatives, but only for 7 the ancestral state was estimated to be winged. The probabilities of ancestral winglessness in males for the higher taxa (Phasmatodea: 48.67%, Euphasmatodea: 84%, Neophasmatodea: 95.33%, Occidophasmata: 96%, Oriophasmata: 97.67%; Fig. 4, Additional file 3: Table S3) are not found to be significantly different from the results obtained from the reconstructions based on the phylogenies with different constraints (B1, B3; Additional file 1: Figs. S2, S3) and in only one instance the macropterous state is estimated to be more likely for the phasmatodean node (B1; Additional file 3: Table S3). The reconstruction of ocelli showed a consistently high likelihood of their absence across the phylogeny and their evolution is estimated to have occurred at least once in Pseudophasmatidae, Phylliidae, Palophidae and Lanceocercata, and three times in Necrosciinae (Additional file 1: Figs. S2–S4).

Phasmatodean phylogenetic relationships and reconstruction of wing states. The phylogeny is based on the Bayesian analysis using constraints (B2) and branches are coloured based on the results of the ancestral state reconstruction of male wings (Additional file 1: Fig. S4; see also Figs. S1–S3 and S5). Branch colour for unknown states corresponds to the most likely state of the parent node. Pie charts on major nodes show the probabilities for the ancestral state. The presence of ocelli is highlighted in yellow at the tips. EUPHAS, Euphasmatodea; NEOPHAS, Neophasmatodea; Occidoph, Occidophasmata; Orioph, Oriophasmata; T, Timematodea; ASCH, Aschiphasmatidae; A, Agathemeridae; PSEU, Pseudophasmatidae; HN, Heteronemiinae; DIAPH, Diapheromerinae; HET, Heteropterygidae; GRA, Gratidiidae sensu Cliquennois [70]; BAC, Bacillinae sensu Cliquennois [70]; CLI, Clitumninae sensu Cliquennois [70]; AFR/MAD, African/Malagasy group including Achriopteridae, Anisacanthidae, Antongiliidae, Damasippoididae and Xylicinae sensu Cliquennois [70]; PHYLL, Phylliidae; PHA, Pharnaciinae + Prosentoria; P, Palophidae; CLA, Cladomorphinae; X, Xenophasmina; ST, Stephanacridini; LANCEO, Lanceocercata; LONCH, Lonchodinae; NEC, Necrosciinae
The evaluation of the number of transition events between states when wing-regain is permitted (ARD) or wing loss is irreversible (IRR) showed that when analysing the 3-state dataset under the ARD model, the number of shifts was highly increased, albeit most shifts were detected between micropterous and macropterous taxa. Comparing the binary dataset under ARD and IRR models and thus the transitions between the presence and absence of wings revealed more sensible results. Under the ARD model, the loss of wings occurred on average twice as likely as their regain with a total of ~ 64 evolutionary shifts on average between states (Fig. 5, Additional file 3: Table S4). In contrast, when revolution of wings was not permitted (IRR), the number of losses is significantly higher (~ 76; Fig. 5, Additional file 1: Fig. S6). Similar results were obtained from comparing the results of ocelli reconstruction with 8 regains and 25 losses under the ARD model compared to 55 losses under the IRR model (Additional file 3: Table S4).

Box-plot diagram of number of transitions between the winged and wingless state. Numbers were generated from performing 300 iterations of stochastic character mapping in phytools based on the binary wing dataset. The distribution of observed transitions events is displayed as box-plots of the 25–75th percentile, with the horizontal line representing the median and vertical lines representing the range (excluding potential outliers). For the ARD model, transition events from wingless to winged and vice versa, and the combined amount are shown. Under the IRR model disallowing wing regain, all transitions are unidirectional. The number of all transitions observed under ARD and IRR was compared using a paired t-test. ***p-value < 2.2e-16
Trait correlation
We tested for correlation of wings and ocelli and recovered strong evidence for their correlated evolution in males and females (Additional file 3: Table S5). Specifically, models performed best when ocelli were set as the dependent variable implying that the evolution of ocelli was depending on the evolution of wings. However, applying different root states influenced the estimation resulting in varying AIC values and Akaike model weights (AICw). In one instance, when the root state for females was set to wingless + no ocelli, the evolution of ocelli and wings was recovered as intradependent. The wingless root state also resulted in the lowest AICw values for males (AICw = 0.5136), whereas the unconstrained, winged + no ocelli and winged + ocelli root states resulted in higher values (AICw = 0.6462, 0.9434 and 0.7895, respectively). The correlation of wings and ocelli was further examined by fitting models of correlated evolution between ocelli and the three individual wing states. While we did not find any evidence that short wings and ocelli are correlated, there was statistically significant correlation recovered between ocelli and long wings as well as winglessness.
Diversification rates
The BAMM analysis converged well and ESS values are well above 200. A total of nine diversification rate shifts was detected across the branches of the phasmatodean phylogeny (Additional file 1: Fig. S7) and the best shift configuration (posterior probability = 0.71) features two shifts that are localised in the clade Euphasmatodea and in the clade of European Bacillinae. The second-best shift configuration additionally shows a slight rate shift for the lineage including among others the Cladomorphinae and Lanceocercata. Overall, the net diversification shows a gradual and constant increase in rate over time and remains comparably low only for Timematodea (Additional file 1: Fig. S7). The results of our trait-dependent diversification rate analysis to assess the influence of wings on the diversification process show that character-independent HiSSE (CID-4) models were inferred to best fit our data (Additional file 3: Table S6). The best-fit model (AICw = 0.9412), the CID-4 model with four hidden states, does not include the focal character (wings) and thus assumes that changes in diversification rate are independent from the presence or absence of wings.
Discussion
Independent of the alternative backbone phylogenies (for a more detailed discussion, see Additional file 4), the reconstruction of the two key characters involved in insect flight produced almost identical results in all analyses (Fig. 4, Additional file 1: Figs. S2–S4). Applying the ARD model and thus allowing transitions rates to be different for gains and losses, the ancestral reconstruction of wings and ocelli estimated the ancestral state to be wingless and anocellate for the major nodes of male and female phasmatodeans (Fig. 4, Additional file 1: Figs. S4, S5). Although Timema and their extinct relatives are completely wingless, the node at the split between Timematodea and Euphasmatodea is however not estimated to be unequivocally in favour of a wingless ancestral form. Under consideration of the significant time gap of approximately more than 50 million years between their divergence and the start of euphasmatodean diversification that has repeatedly been inferred [4, 54,55,56,57, 71, 72], it cannot be ruled out that stem group euphasmatodeans were winged. The scarce fossil record [69, 73, 74] shows that extinct Phasmatodea from the Cretaceous were predominantly winged [36,37,38, 75,76,77], but are thought to belong to the stem group of Phasmatodea rather than Euphasmatodea [69, 74, 77]. Hence, the fossil evidence is not contradicting a wingless ancestor of crown Phasmatodea or Euphasmatodea. Moreover, it is noteworthy that the wing morphology of these Mesozoic winged stick insects is different from that of recent phasmatodeans [38, 78]: All fossil specimens exhibit two long pairs of wings [36,37,38], whereas the sclerotised forewings (tegmina) of extant macropterous forms are shortened with very few exceptions (see below). Long tegmina are in fact considered the plesiomorphic condition of Polyneoptera, which are suggested to have been independently reduced in various lineages such as in stoneflies or webspinners as well as stick and leaf insects [79], albeit it remains unclear when and how often forewing shortening occurred in Phasmatodea. Also the fossils’ tegmina venation is different and its modification in modern phasmatodeans might be due to a convergent evolutionary process of wing shortening and not because of common ancestry, especially when considering the few extant, unrelated and strongly subordinated species that exhibit long tegmina with venation differing from other recent forms (i.e., Heteropteryx, Prisopus, Phylliidae) [78, 80, 81]. While the large tegmina of the flightless leaf insect females (Phylliidae) contribute to their leaf mimicry and therefore might have been secondarily elongated [82, 83] (the volant males possess shorter tegmina), the wing morphology of male Heteropteryx is most similar to that of stem phasmatodeans [36, 78, 81]. As subordinated lineage within the ground-dwelling and predominantly wingless Heteropterygidae, it was suggested that Heteropteryx secondarily became arboreal and the male regained the capability of active flight. Due to the most plesiomorphic wing morphology among extant stick and leaf insects, its wings were hypothesised to be the product of an atavistic reversion [69, 80]. Our results support this hypothesis and not only for this clade, where morphology is corroborative, but also for the more diverse winged lineages of Euphasmatodea.
Regardless of whether wings were present or absent in the common ancestor of Euphasmatodea, our results show an increase in diversification rate with the start of their radiation (Additional file 1: Fig. S7). This rapid radiation that largely follows the K-Pg mass extinction (~ 66 mya) was previously suggested to be linked to the diversification of flowering plants [56]—a co-evolutionary pattern also observed in other plant-associated insect groups [84,85,86]. Our results substantiate this hypothesis, particularly, when considering the results of the trait-dependent diversification analysis (HiSSE) from which we concluded that flight or flightlessness might not have been the main driver of euphasmatodean diversification. Alternatively, the evolution of hard-shelled eggs [55, 81, 87] and the acquisition of endogenous pectinase genes [88] further shaping the co-evolution with their food plants are most likely to have contributed to the success of the early euphasmatodean lineages. Also the recurrent opportunities for colonising new land masses (i.a., the Indo-Pacific region) appears to have promoted speciation [80, 89] and might explain the increased diversification rate recovered for Lonchodidae, Lanceocercata and relatives within Oriophasmata (square symbol; Additional file 1: Fig. S7). By contrast, the slight decrease in the rate for the European Bacillinae (circle symbol) might be an artefact resulting from oversampling closely related taxa, most of which are only represented by one single gene, but could also correspond to the ecological shift to a temperate region. Given that both the presence of wings and the lack thereof were shown to potentially increase diversification in insects [1, 2, 6, 9,10,11,12], it cannot be excluded that either condition had an equal impact on the diversification of the individual euphasmatodean lineages (e.g., for the predominantly macropterous and most species-rich lineage Necrosciinae versus the speciose and mostly wingless Lonchodinae [69, 90, 91]). Ultimately, that no significant change in rate among subgroups was detected and character-independent diversification was favoured is indicative of the wing state not being subject to Dollo’s law of irreversible evolution [43].
Within Euphasmatodea, only few lineages were recovered as ancestrally winged (e.g., Pseudophasmatidae, Phylliidae), whereas in others, the winged taxa are found rather subordinated among otherwise wingless forms (e.g., Lonchodinae, Diapheromerinae). In contrast to the seemingly random distribution of apterous species in ancestrally winged lineages, the winged species in predominantly wingless groups are closely related, indicating single origins of wing re-evolution. Discrepancies may be explained by incomplete taxon sampling or wrongly recovered phylogenetic position, which might diminish in a phylogenomic context or when applying an even denser taxon sampling. Conversely, the multiple instances of secondarily apterous taxa found among winged lineages are the result of the higher probability of wing loss in comparison to that of regain [30, 39, 40, 48]. Generally, evolutionary transitions were proposed to occur rapidly and often in Phasmatodea—not only between apterous and macropterous taxa but also between wing states [31]. Previous authors argued that the probability or cost ratio of wing gain was considered too high compared with that of its loss [40]. In our reconstruction, the estimated number of potential regains ranges between 9 and 36 events (~ 22) with ~ 42 losses (27–59) versus ~ 76 losses (71–82) if wing loss is considered irreversible (Fig. 5), hence, permitting re-evolution appears to be more parsimonious. For instance, in predominantly wingless Diapheromerinae, we find 1–3 potential wing regain events versus an alternative of a total of ten losses if wing recovery was impossible. Although our results clearly support the hypothesis of re-evolution, we nevertheless acknowledge that the reacquisition of a complex trait such as wings must be assumed less likely than its loss, and that it is possible that ultimately, independent wing loss might have occurred much more often in phasmatodeans than in other insect groups. However, how can we then explain the distant disjunct distribution of ocelli in Phasmatodea?
Insect ocelli have not been intensively studied in a phylogenetic context before, but it is well-known that there are winged lineages within Pterygota that lack ocelli such as earwigs (Dermaptera) and webspinners (Embioptera). Although both groups have undergone a unique modification of wings, their specific lifestyle led to a reduced need for aerial dispersal over time (ground-dwelling lifestyle of Dermaptera; silk galleries of Embioptera) [79, 92, 93], which generally resulted in 20–40% of flightlessness and the complete loss of wings in all embiopteran females [1] and potentially promoting the loss of ocelli. In contrast, the majority of phasmatodean taxa are flightless or wingless [1, 30, 69], and some winged species do possess ocelli (Fig. 3). While wings are certainly homologous, we were not able to identify any lineage-specific differences except for those mentioned above (e.g., female leaf insects). In regard to ocelli, only those of male Lanceocercata are found to be particularly prominent (Fig. 2C, D, K). However, the differences of size and spacing are generally more dependent on the shape of the head than on specific lineages, with larger differences found between sexes of a single species than present among lineages. Considering the significant number of macropterous anocellate species and the seemingly arbitrary distribution of ocelli-bearing lineages raises the question whether ocelli might have re-evolved; otherwise, if ocelli had been in the ground plan of winged (Eu)Phasmatodea, why would most lineages that retained the capability of flight have reduced this flight-stabilising sensory system? For instance, the Aschiphasmatidae, sister group to the remaining Euphasmatodea (= Neophasmatodea), comprise besides wingless forms several fully-winged species that are capable of active flight, yet, none possess ocelli. Our results show that the presence of ocelli is restricted to five subordinated and completely unrelated lineages, albeit winged and volant taxa are found more frequently across Phasmatodea (Fig. 4). While it is plausible to assume that ocelli might have been lost in winged lineages that contemporarily lost the capability of flight (i.e., micropterous lineages), the high number of macropterous and volant species lacking ocelli is perplexing. The alternative scenario involving the possibility of wing regain implies that secondarily winged lineages re-evolved the ocelli subsequent to or simultaneously with the recovery wings, which is highly supported by our analyses. Specifically, the ancestral state of Pseudophasmatidae, Palophidae and Phylliidae was recovered as possessing ocelli along with wings, whereas ocelli are estimated to have occurred only in subordinated clades in Lanceocercata and Necrosciinae in spite of wings and flight being recovered to have a more ancestral origin. The necrosciine taxa represent an optimal example, since ocelli occur only in one single subclade within a highly diverse macropterous lineage and are otherwise only present in two unrelated species, where they must have re-appeared independently (Hemisosibia incerta, Gargantuoidea triumphalis). Conversely, it might be a common trend for brachypterous species within the ocelli-bearing clades to reduce ocelli (as observed in other insect groups [94,95,96,97]), albeit there are also few macropterous species lacking ocelli (e.g., Creoxylus, Eurynecroscia, Paraprisopus).
In comparison to the other ocelli-bearing clades, the distribution of ocelli within Phylliidae appears more ambiguous. While females are sedentary and flightless, all males are volant and depend on flight for mate localisation [69, 89, 98], thus, it is questionable whether there have been multiple independent secondary losses of ocelli as estimated by our analysis. Particularly the phylogenetically incoherent degree of ocelli development in the species of Phyllium, where ocelli may be absent or weakly, moderately or well developed (not coded in our analysis), suggests that the ocellar system is a disparate character, which might have gradually and independently re-evolved in several phylliid lineages. The possibility of ocelli reacquisition is further supported by photographs [99] and specimens that we examined of the Giant Malaysian Leaf Insect (Pulchriphyllium giganteum) showing the presence of ocelli not only in males, but also in females. This detail is lacking from the original species description [100] and from subsequent morphological studies including this species [101, 102]. Contradicting the previous assumption that female leaf insects are entirely devoid of ocelli [69, 103], this discovery may be interpreted as further evidence for secondary ocelli gain—even in flightless species or sexes. Generally, however, it cannot be excluded that a potentially functional ocellar system may be present internally in the absence of an external (visible) lens as it has been observed in other insects [104,105,106,107,108] and that this condition preceded the development of fully-formed ocelli at least in some lineages such as Phyllium. The alternative scenario involving (gradual) reduction of ocelli in volant stick and leaf insects does not appear plausible especially under consideration of the different functions of ocelli besides those related to flight [60, 61, 64, 65, 109]. Also the predominantly nocturnal life style would rather promote the enlargement of ocelli and not their reduction [61, 110]. Similarly, it is also argued that in dense vegetation, where there is no clearly visible horizon line, flying insects such as Necrosciinae would undergo specific modifications to improve the performance of the ocellar system to compensate for the obscured horizon [65, 109], yet, there are no habitat differences between ocellate and anocellate Necrosciinae. Hence, we favour the hypothesis that ocelli are not plesiomorphic with copious losses but instead were re-evolved as was also proposed for lineages of the predominantly anocellate beetles [67, 111]. To further corroborate this assumption, future studies are needed to elucidate the organisation and development of ocelli across phasmatodean taxa as well as to examine the internal morphology of the head capsule to clarify the potential presence of internal structures in anocellate species that facilitate ocelli emergence.
Under the assumption of Dollo’s law of irreversible evolution, even in regard to single traits, the developmental genetic foundation of a complex structure is lost along with the trait itself and therefore cannot be reacquired because evolving the same complex structures de novo is not considered possible. However, if the underlying genetic architecture for the lost trait remains conserved, reinitiation of suppressed genes might result in its recovery. Moreover, evolutionary changes in gene regulatory networks are proposed to have a major influence on morphological evolution [112, 113]. Given that the genetic components and developmental processes of a complex character were conserved, evolutionary shifts in developmental gene expression patterns might be responsible for the loss as well as for the regain of a trait [112]. Also wing polyphenism in caste systems is underlying a similar concept, where wingless morphs are produced from the same genetic basis as winged morphs [29]. For instance in the wingless castes of ants it was shown that wing loss is caused by the interruption of the wing formation pathway at different points in different species, which thus might easily be reversed in the absence of such interruption [114]. The key elements underlying insect flight have been conserved over hundreds of million years of evolution and may remain available for reactivation also in secondarily wingless insects for a long period of time [115], albeit it was estimated that silenced genetic pathways cannot retain function for more than 10 million years [116]. Yet, regulatory elements may be reinitiatable over longer periods of time [53], especially when the genetic developmental programme is underlying pleiotropic maintenance and is accommodated in similar structures or tissues still present [30, 50, 51, 53, 112]. Since the genetic basis for insect wing development is retained in leg formation [117, 118], genes may have been co-opted from these existing expression patterns that then elicited wing re-evolution in stick and leaf insects [30]. Although a gradual process of the reversal of wing loss involving the necessary deployment of multiple genes can generally be assumed, it has been demonstrated that musculature and innervation associated with flight can be maintained in flightless phasmatodeans [119], while in other insects flight loss may be explained by degradation of these structures [27]. Therefore, we suggest that in phasmatodeans apparently a latent and hitherto not understood capacity to produce wings and flight-associated traits exists (or existed) that facilitated the reappearance of wings, flight and ocelli to occur multiple times independently. Our hypothesis thus incorporates that the disjunct distribution of these traits did not result from plesiomorphy and numerous losses but from a radiation of wingless ancestors whose descendants independently regained wings (and ocelli). This is further supported by the effects attributed to the loss of flight in the wingless ancestor such as increase in body size and reduction in eye size [1, 60, 120] of which the latter is still observable in modern volant species indicating a rather recent regain of flight. Since the advantages of both flight and flightlessness are equally abundant, the trade-off between dispersal and fecundity is reflected in phasmatodean capability of the dynamic transition between wing states as well as by the high occurrence of sexual dimorphism in terms of size and wings [31].
Conclusion
The currently expanding body of research reporting re-evolution and reversals clearly challenges our assumptions about the irreversibility of complex traits. By demonstrating that wings as well as the flight-associated ocelli have re-evolved in Phasmatodea based on an extensive taxon sampling, our study contributes to this development and establishes a basis for future work to further investigate the regain of wings and flight in stick and leaf insects including the comparison of wing lengths as a continuous character and the assessment of flight capability. Ideally, morphological, developmental, phylogenomic and palaeontological work can eventually be united into a comprehensive picture to understand phasmatodean wing and flight evolution and the underlying genetic developmental processes that lead to the manifestation of atavistic reversals.
Material and methods
Taxon and gene sampling
Our comprehensive dataset consisted of 513 species and specimens with representatives of all major clades of Phasmatodea and covering almost 50% of its generic diversity (Additional file 3: Table S7). For the outgroup, we further included two species of Embioptera, which were repeatedly recovered as the sister group of Phasmatodea [4, 30, 56, 121]. Our gene sampling comprised three nuclear (18S, 28S, H3) and four mitochondrial loci (12S, 16S, COI, COII) of which data for numerous specimens were already available on GenBank [30, 54, 55, 71, 80, 89, 90, 101, 122,123,124,125,126,127,128,129,130,131]. Additionally, we generated new data for 194 specimens following the protocol given by Bank et al. [80] with primers adopted from Buckley et al. [124] and Robertson et al. [55] (see Bank et al. [80]), and deposited the sequences in GenBank under accession numbers OK314533–OK314857, OK324156–OK324324, OK333379–OK334012 (Additional file 3: Table S7).
Phylogenetic analysis and divergence time estimation
Phylogenetic comparative analyses are highly dependent on a robust phylogeny. Hence, in order to optimise our topology, we modified our tree inferences to mimic the phylogenetic relationships obtained from transcriptomic studies [56, 57] by constraining the higher taxonomical groups (i.e., Phasmatodea, Euphasmatodea, Neophasmatodea, Oriophasmata and Occidophasmata).
The final supermatrix of 5636 bp (Additional file 2: File S6) was obtained by aligning, trimming and concatenating the sequences as described by Bank et al. [80]. We partitioned the dataset by gene and by codon position for ribosomal (12S, 16S, 18S, 28S) and for protein-coding (COI, COII, H3) genes, respectively, prior to determining the best-fit partitioning scheme and evolutionary model using ModelFinder [132] (options -m MF –merge greedy) integrated in IQ-TREE v. 2.1.2 [133]. The third codon position of the COI and COII genes as well as the second and third position of H3 was merged, resulting in 11 partitions (see Additional file 3: Table S8).
We applied two approaches to infer the phylogenetic relationships: Maximum Likelihood (ML) and Bayesian Inference (BI). For the ML tree inference, we used IQ-TREE [133, 134] to conduct 100 independent tree searches using the aforementioned constraints and the best-fit partitioning scheme and substitution models. For each search, we used a random starting tree, increased the number of unsuccessful iterations (-nstop 500) and set the perturbation strength to 0.2. We computed the log-likelihoods and at the same time tested the tree topologies to identify the best-scoring tree. Nodal support was subsequently calculated using the Ultrafast Bootstrap approximation (UFBoot) and the SH approximate likelihood ratio test (aLRT) [135] by generating 10,000 replicates. UFBoot support values were plotted on the best tree, whereas for the SH-aLRT test, a new phylogeny was generated by default in IQ-TREE.
For the BI tree, we simultaneously conducted the phylogenetic analysis with the divergence time estimation in BEAST2 v. 2.6.1 [136]. We used the same partitioning scheme as for the ML analysis, but substitution models were selected by bModelTest v. 1.2.1 [137] implemented in BEAST. We linked trees and clocks across all partitions and applied the Yule tree prior and a relaxed clock with an uncorrelated lognormal distribution (UCLD) and a clock-rate of 1e−7. We time-calibrated the tree using five fossils unambiguously assigned to Phasmatodea (Additional file 3: Table S9) and chose a lognormal prior distribution with the minimum age selected as the offset and log-mean and log-sd set to 1.0. In the case of the two fossils from Dominican amber (Clonistria sp. and Malacomorpha sp.), the calibrated node was selected to include the sister group (option “use originate”). For these fossils as well as for the Phasmatodea calibration, the log-mean was set to 2.0 to allow a softer maximum bound. To account for potential phylogenetic incongruences that might have an impact on the reconstruction of ancestral states, we performed three independent analyses based on different constraints: For the first, we used the same constraints as for the ML analysis (B1), for the second we additionally constrained the Heteropterygidae as the sister group to all the remaining Oriophasmata (following the results of transcriptomic studies [56, 57]) (B2), and for the third, we did not set any constraints for Oriophasmata and Occidophasmata and thus only for Phasmatodea, Euphasmatodea and Neophasmatodea (B3). All analyses were run for 300 million generations with parameter and tree sampling every 10,000 generations. Convergence was assessed in Tracer v. 1.7.1 [138] and a Maximum Clade Credibility tree was generated after removing the first 3000 trees as burn-in.
Morphological data
To compile our morphological data matrix, we gathered information on the wing states and the possession of ocelli for both sexes for each taxon included (Additional file 3: Table S10). For specimens not available for examination, we reviewed the literature (e.g., Redtenbacher [139, 140]) and searched for suitable photographs of wild, captive-bred or type species online (www.phasmida.speciesfile.org, www.phasmatodea.com). While we assembled the ocelli data based on their presence or absence and did not distinguish between different types of ocelli (e.g., number, relative size), wing states were coded as wingless (apterous, 0), partially-winged (micropterous, 1) or fully-winged (macropterous, 2). The differentiation between micropterous and macropterous was made arbitrarily as we defined a morph as fully-winged when the hind wings exceeded the fourth abdominal segment [81]. Since some analyses require binary data, we created two additional datasets by combining the three wing states based on wingless (0) versus winged (1) taxa (datawings) and based on flightless (0) versus fully-winged and presumably volant (1) taxa (dataflight). Missing data were generally coded as absent (0). For all analyses, the fossil-calibrated BEAST tree with B2 constraints was used.
Phylogenetic signal
Using the binary datasets for males, we estimated the phylogenetic signal of the presence of ocelli, presence of wings (datawings) and capability of flight (dataflight) by calculating the D statistic [141] in the R package caper v. 1.0.1 [142] using the phylo.d function with default parameters and 10,000 permutations. Furthermore, we assessed the phylogenetic signal using the function fitDiscrete in the R package geiger v. 2.0.7 [143] to fit Pagel’s lambda [144] under the ARD transition model. We expected D estimates to be ≤ 0 and lambda values closer to 1, both serving as an indicator for traits having evolved under a Brownian motion model and not to be recovered as randomly distributed across the phylogeny. To further evaluate a potential random distribution, we created a randomised character matrix to compare the number of evolutionary transitions with the results from our true dataset as outlined elsewhere [145, 146].
Ancestral state reconstruction
Ancestral states of wings and ocelli were estimated in the R package phytools v. 0.7-70 [147]. To determine the best-fit model, we used the functions fitMk and fitDiscrete to fit different models to the binary dataset of male wing traits (datawings). Using fitDiscrete, we tested models that permitted evolutionary rates between wingless and winged to be equal in both directions (equal-rates, ER), different in all directions (all-rates-different, ARD) and unidirectional from winged to wingless (customised irreversible model, IRR). We repeated the tests using the function fitMk and additionally tested the IRR model with the root state fixed to “winged” and the ARD model with the root state fixed to “wingless”. Finally, for the ancestral state reconstruction, we used the 3-states datasets of wings and the binary datasets of ocelli for both males and females, and coded missing data with equal probabilities for each of the possible states. We performed stochastic character mapping [148] with the function make.simmap implemented in phytools based on the best-fit model (ARD) and ran 300 iterations of Markov chain Monte Carlo (MCMC) sampling. Using the option Q = “mcmc”, the rate matrix Q is sampled 300 times from its posterior probability distribution using MCMC and stochastic maps and node states are generated conditioned on each sampled value of Q. To assess whether the topology of the tree had an impact on the estimated ancestral states, we also performed the analysis using the alternative trees (B1 and B3 constraints).
Given the 300 inferred possible evolutionary histories resulting from stochastic character mapping, we calculated the average number of transition events between wing states and the presence/absence of ocelli. For comparison with an evolutionary hypothesis where wing/ocelli regain is not permitted, we repeated simulating stochastic maps as described above but using the IRR model. To be even more conservative, we also reconstructed ancestral states using both models for the binary wing dataset (datawings).
Trait correlation
We used Pagel’s binary character correlation test [149] as implemented in phytools to assess potential coevolutionary dynamics between wings and ocelli in males and females. By applying the ARD model and using different parameters for the argument “dep.var”, we estimated whether wings and ocelli evolved independently, intradependently or whether the evolution of wings was dependent on the evolution of ocelli or vice versa, and determined the best model to explain the correlated evolution with the Akaike information criterion (AIC) and Akaike weights. As tests for correlation of traits may be sensitive to the root state, we explored the impact of applying different root states by fixing the root to represent either wingless + no ocelli, winged + no ocelli, or wings + ocelli. We further tested for the correlation of ocelli and either of the three wing states (apterous, micropterous, macropterous). We estimated and compared the values of log-likelihoods and Akaike weights, and corrected p-values for multiple comparisons using the Bonferroni correction.
Diversification analysis
We explored evolutionary rate dynamics with the Bayesian analysis of macroevolutionary mixtures (BAMM) and the R package BAMMtools [150, 151]. Priors were estimated using the BAMMtools function “setBAMMpriors” and modified accordingly (expectedNumberOfShifts = 1.0; lambdaInitPrior = 4.80107993368211; lambdaShiftPrior = 0.00644760145800591; muInitPrior = 4.80107993368211). We ran four chains of 10 million generations being sampled every 1000 generations using the “speciationextinction” model and setting “globalSamplingFraction” to 0.15 (~ 500 taxa of ~ 3400 described species). Convergence was assessed and subsequent data analysis performed in BAMMtools.
Whether the evolution of wings had an impact on diversification rates was tested using the Hidden State-dependent Speciation and Extinction model (HiSSE) in the R package hisse [152]. We used the binary dataset of male wings and removed the outgroup (Embioptera). Following Song et al. [153], we then fitted 24 different models [152] including original BiSSE models [154], character-independent diversification models (CID-2 and CID-4) and HiSSE models, which assume a hidden state. All models were compared using log-likelihood, AIC and Akaike weights.
Availability of data and materials
The data supporting the conclusions of this article are included within the article and its additional files. Newly obtained sequences have been deposited in GenBank under accession number OK314533–OK314857, OK324156–OK324324, OK333379–OK334012 (see also Additional file 3: Table S7). Additional data and code for this paper are available at GitHub (https://github.com/sb4nk/Phasmatodea_flightevol).
References
Wagner DL, Liebherr JK. Flightlessness in insects. Trends Ecol Evol. 1992;7:216–20. https://doi.org/10.1016/0169-5347(92)90047-F.
Grimaldi D, Engel MS. Evolution of insects. Cambridge: Cambridge University Press; 2005.
May RM. How many species are there on Earth? Science. 1988;241:1441–9. https://doi.org/10.1126/science.241.4872.1441.
Misof B, Liu S, Meusemann K, Peters RS, Donath A, Mayer C, et al. Phylogenomics resolves the timing and pattern of insect evolution. Science. 2014;346:763–7. https://doi.org/10.1126/science.1257570.
Douglas MM. Thermoregulatory significance of thoracic lobes in the evolution of insect wings. Science. 1981;211:84–6. https://doi.org/10.1126/science.211.4477.84.
Roff DA. The evolution of flightlessness in insects. Ecol Monogr. 1990;60:389–421.
Roff DA. The evolution of flightlessness: is history important? Evol Ecol. 1994;8:639–57. https://doi.org/10.1007/BF01237847.
Wipfler B, Klug R, Ge S-Q, Bai M, Göbbels J, Yang X-K, et al. The thorax of Mantophasmatodea, the morphology of flightlessness, and the evolution of the neopteran insects. Cladistics. 2015;31:50–70. https://doi.org/10.1111/cla.12068.
Emerson BC, Wallis GP. Phylogenetic relationships of the Prodontria (Coleoptera; Scarabaeidae; subfamily Melolonthinae), derived from sequence variation in the mitochondrial cytochrome oxidase II gene. Mol Phylogenet Evol. 1995;4:433–47. https://doi.org/10.1006/mpev.1995.1040.
Bell KL, Yeates DK, Moritz C, Monteith GB. Molecular phylogeny and biogeography of the dung beetle genus Temnoplectron Westwood (Scarabaeidae: Scarabaeinae) from Australia’s wet tropics. Mol Phylogenet Evol. 2004;31:741–53. https://doi.org/10.1016/j.ympev.2003.09.010.
Ikeda H, Nishikawa M, Sota T. Loss of flight promotes beetle diversification. Nat Commun. 2012;3:648. https://doi.org/10.1038/ncomms1659.
Mitterboeck TF, Liu S, Adamowicz SJ, Fu J, Zhang R, Song W, et al. Positive and relaxed selection associated with flight evolution and loss in insect transcriptomes. Gigascience. 2017;6:1–14. https://doi.org/10.1093/gigascience/gix073.
Denno RF, Roderick GK, Olmstead KL, Döbel HG. Density-related migration in planthoppers (Homoptera: Delphacidae): the role of habitat persistence. Am Nat. 1991;138:1513–41.
Roff DA. Habitat persistence and the evolution of wing dimorphism in insects. Am Nat. 1994;144:772–98. https://doi.org/10.1086/285706.
Roff DA. Life history consequences of bioenergetic and biomechanical constraints on migration. Am Zool. 1991;31:205–16. https://doi.org/10.1093/icb/31.1.205.
Zera AJ, Denno RF. Physiology and ecology of dispersal polymorphism in insects. Annu Rev Entomol. 1997;42:207–30. https://doi.org/10.1146/annurev.ento.42.1.207.
Zera AJ. Differences in survivorship, development rate and fertility between the longwinged and wingless morphs of the waterstrider, Limnoporus canaliculatus. Evolution. 1984;38:1023–32. https://doi.org/10.2307/2408436.
Roff DA. The evolution of wing dimorphism in insects. Evolution. 1986;40:1009–20.
Mole S, Zera AJ. Differential allocation of resources underlies the dispersal-reproduction trade-off in the wing-dimorphic cricket, Gryllus rubens. Oecologia. 1993;93:121–7.
Braendle C, Davis GK, Brisson JA, Stern DL. Wing dimorphism in aphids. Heredity (Edinb). 2006;97:192–9. https://doi.org/10.1038/sj.hdy.6800863.
Lorenz MW. Oogenesis-flight syndrome in crickets: Age-dependent egg production, flight performance, and biochemical composition of the flight muscles in adult female Gryllus bimaculatus. J Insect Physiol. 2007;53:819–32. https://doi.org/10.1016/j.jinsphys.2007.03.011.
Langellotto GA, Denno RF, Ott JR. A trade-off between flight capability and reproduction in males of a wing-dimorphic insect. Ecology. 2000;81:865–75. https://doi.org/10.1890/0012-9658(2000)081[0865:ATOBFC]2.0.CO;2.
Guerra PA. Evaluating the life-history trade-off between dispersal capability and reproduction in wing dimorphic insects: a meta-analysis. Biol Rev. 2011;86:813–35. https://doi.org/10.1111/j.1469-185X.2010.00172.x.
Kotyk M, Varadínová Z. Wing reduction influences male mating success but not female fitness in cockroaches. Sci Rep. 2017;7:2367. https://doi.org/10.1038/s41598-017-02647-7.
Fong DW, Kane TC, Culver DC. Vestigialization and loss of nonfunctional characters. Annu Rev Ecol Syst. 1995;26:249–68. https://doi.org/10.1146/annurev.es.26.110195.001341.
Harrison RG. Dispersal polymorphisms in insects. Annu Rev Ecol Syst. 1980;11:95–118.
McCulloch GA, Wallis GP, Waters JM. Do insects lose flight before they lose their wings? Population genetic structure in subalpine stoneflies. Mol Ecol. 2009;18:4073–87. https://doi.org/10.1111/j.1365-294X.2009.04337.x.
Zera AJ. Wing polymorphism in Gryllus (Orthoptera: Gryllidae): proximate endocrine, energetic and biochemical mechanisms underlying morph specialization for flight vs. reproduction. In: Whitman DW, Ananthakrishnan TN, editors. Phenotypic plasticity of insects: mechanisms and consequences. Enfield: Science Publishers Inc; 2009. p. 609–53.
Simpson SJ, Sword GA, Lo N. Polyphenism in insects. Curr Biol. 2011;21:R738–49. https://doi.org/10.1016/j.cub.2011.06.006.
Whiting MF, Bradler S, Maxwell T. Loss and recovery of wings in stick insects. Nature. 2003;421:264–7. https://doi.org/10.1038/nature01274.1.
Zeng Y, O’Malley C, Singhal S, Rahim F, Park S, Chen X, et al. A tale of winglets: evolution of flight morphology in stick insects. Front Ecol Evol. 2020;8:121. https://doi.org/10.3389/fevo.2020.00121.
Robinson MH. The defensive behavior of Pterinoxylus spinulosus Redtenbacher, a winged stick insect from Panama (Phasmatodea). Psyche (Stuttg). 1968;75:195–207. https://doi.org/10.1155/1968/19150.
Bedford GO. Biology and ecology of the Phasmatodea. Annu Rev Entomol. 1978;23:125–49. https://doi.org/10.1146/annurev.en.23.010178.001013.
Carlberg U. Defensive stridulation in Heteropteryx dilatata Parkinson (Insecta: Phasmida). Zool Anz. 1989;223:165–73.
Hennemann FH, Conle OV. Studies on the genus Phasmotaenia Navás, 1907, with the descriptions of five new species from the Solomon Islands, a revised key to the species and notes on its geographic distribution (Phasmatodea: ‘Anareolatae’: Phasmatidae s. l: Stephanacridini). Zootaxa. 2009;2011:1–46. https://doi.org/10.11646/zootaxa.2011.1.1.
Shang L, Béthoux O, Ren D. New stem-Phasmatodea from the Middle Jurassic of China. Eur J Entomol. 2011;108:677–85. https://doi.org/10.14411/eje.2011.086.
Wang M, Béthoux O, Bradler S, Jacques FMB, Cui Y, Ren D. Under cover at pre-angiosperm times: a cloaked phasmatodean insect from the early Cretaceous Jehol biota. PLoS ONE. 2014;9: e91290. https://doi.org/10.1371/journal.pone.0091290.
Yang H, Yin X, Lin X, Wang C, Shih C, Zhang W, et al. Cretaceous winged stick insects clarify the early evolution of Phasmatodea. Proc R Soc B Biol Sci. 2019;286:20191085. https://doi.org/10.1098/rspb.2019.1085.
Stone G, French V. Evolution: have wings come, gone and come again? Curr Biol. 2003;13:R436–8. https://doi.org/10.1016/S0960-9822(03)00364-6.
Trueman JWH, Pfeil BE, Kelchner SA, Yeates DK. Did stick insects really regain their wings? Syst Entomol. 2004;29:138–9. https://doi.org/10.1111/j.0307-6970.2004.00251.x.
Gould SJ. Dollo on Dollo’s law: irreversibility and the status of evolutionary laws. J Hist Biol. 1970;3:189–212. https://doi.org/10.1007/BF00137351.
Collin R, Miglietta MP. Reversing opinions on Dollo’s Law. Trends Ecol Evol. 2008;23:602–9. https://doi.org/10.1016/j.tree.2008.06.013.
Goldberg EE, Igić B. On phylogenetic tests of irreversible evolution. Evolution. 2008;62:2727–41. https://doi.org/10.1111/j.1558-5646.2008.00505.x.
Simpson GG. The major features of evolution. New York: Columbia University Press; 1953.
Gould SJ, Robinson BA. The promotion and prevention of recoiling in a maximally snaillike vermetid gastropod: a case study for the centenary of Dollo’s Law. Paleobiology. 1994;20:368–90.
Dawkins R. The blind watchmaker. New York: W. W. Norton & Company; 1994.
Andersen NM. Phylogenetic tests of evolutionary scenarios: the evolution of flightlessness and wing polymorphism in insects. In: Grandcolas P, editor. The origin of biodiversity in insects: phylogenetic tests of evolutionary scenarios. Paris: Mémoires du muséum national d’histoire naturelle; 1997. p. 91–108.
Whiting MF, Whiting AS. Is wing recurrence really impossible?: A reply to Trueman et al. Syst Entomol. 2004;29:140–1. https://doi.org/10.1111/j.1365-3113.2004.0255.x.
Oakley TH. On homology of arthropod compound eyes. Integr Comp Biol. 2003;43:522–30. https://doi.org/10.1093/icb/43.4.522.
West-Eberhard MJ. Developmental plasticity and evolution. Oxford: Oxford University Press; 2003.
Kohlsdorf T. Reversibility of digit loss revisited: limb diversification in Bachia lizards (Gymnophthalmidae). J Exp Zool Part B Mol Dev Evol. 2021. https://doi.org/10.1002/jez.b.23024.
Wiens JJ. Re-evolution of lost mandibular teeth in frogs after more than 200 million years, and re-evaluating Dollo’s law. Evolution. 2011;65:1283–96. https://doi.org/10.1111/j.1558-5646.2011.01221.x.
Lynch VJ. Return of a lost structure in the evolution of felid dentition revisited: a DevoEvo perspective on the irreversibility of evolution. bioRxiv. 2021. https://doi.org/10.1101/2021.02.04.429820.
Forni G, Martelossi J, Valero P, Hennemann FH, Conle OV, Luchetti A, et al. Macroevolutionary analyses provide new evidences of phasmids wings evolution as a reversible process. bioRxiv. 2020. https://doi.org/10.1101/2020.10.14.336354.
Robertson JA, Bradler S, Whiting MF. Evolution of oviposition techniques in stick and leaf insects (Phasmatodea). Front Ecol Evol. 2018;6:216. https://doi.org/10.3389/fevo.2018.00216.
Simon S, Letsch H, Bank S, Buckley TR, Donath A, Liu S, et al. Old World and New World Phasmatodea: phylogenomics resolve the evolutionary history of stick and leaf insects. Front Ecol Evol. 2019;7:345. https://doi.org/10.3389/fevo.2019.00345.
Tihelka E, Cai C, Giacomelli M, Pisani D, Donoghue PCJ. Integrated phylogenomic and fossil evidence of stick and leaf insects (Phasmatodea) reveal a Permian-Triassic co-origination with insectivores. R Soc Open Sci. 2020;7: 201689. https://doi.org/10.1098/rsos.201689.
Hustert R, Klug R. Evolution of a new sense for wind in flying phasmids? Afferents and interneurons. Naturwissenschaften. 2009;96:1411–9. https://doi.org/10.1007/s00114-009-0597-7.
Krapp HG. Ocelli. Curr Biol. 2009;19:R435–7. https://doi.org/10.1016/j.cub.2009.03.034.
Taylor CP. Contribution of compound eyes and ocelli to steering of locusts in flight: I. Behavioural analysis. J Exp Biol. 1981;93:1–18.
Mizunami M. Information processing in the insect ocellar system: comparative approaches to the evolution of visual processing and neural circuits. Adv In Insect Phys. 1995;25:151–265. https://doi.org/10.1016/S0065-2806(08)60065-X.
Stange G, Stowe S, Chahl JS, Massaro A. Anisotropic imaging in the dragonfly median ocellus: a matched filter for horizon detection. J Comp Physiol A Sensory, Neural, Behav Physiol. 2002;188:455–67. https://doi.org/10.1007/s00359-002-0317-7.
Wilson M. The functional organisation of locust ocelli. J Comp Physiol A. 1978;124:297–316.
Honkanen A, Saari P, Takalo J, Heimonen K, Weckström M. The role of ocelli in cockroach optomotor performance. J Comp Physiol A Neuroethol Sensory, Neural, Behav Physiol. 2018;204:231–43. https://doi.org/10.1007/s00359-017-1235-z.
Ribi W, Zeil J. Diversity and common themes in the organization of ocelli in Hymenoptera, Odonata and Diptera. J Comp Physiol A Neuroethol Sensory, Neural, Behav Physiol. 2018;204:505–17. https://doi.org/10.1007/s00359-018-1258-0.
Szumik CA. The higher classification of the order Embioptera: a cladistic analysis. Cladistics. 1996;12:41–64. https://doi.org/10.1111/j.1096-0031.1996.tb00192.x.
Leschen RAB, Beutel RG. Ocellar atavism in Coleoptera: plesiomorphy or apomorphy? J Zool Syst Evol Res. 2004;42:63–9. https://doi.org/10.1046/j.0947-5745.2003.00241.x.
Beier M. Phasmida (Stab- oder Gespenstschrecken). Handbuch der Zoologie 4 (2) 2/10. Walter de Gruyter & Company, Berlin, Germany; 1968. 1–56 p.
Bradler S, Buckley TR. Biodiversity of Phasmatodea. In: Foottit RG, Adler PH, editors. Insect biodiversity: science and society, vol. II. 1st ed. Wiley-Blackwell: Hoboken; 2018. p. 281–313. https://doi.org/10.1002/9781118945582.ch11.
Cliquennois N. Ordre des Phasmatodea (Phasmes). In: Aberlenc H-P, editor. Les insectes du Monde. 2020. https://doi.org/10.5962/bhl.title.122844.
Bradler S, Cliquennois N, Buckley TR. Single origin of the Mascarene stick insects: ancient radiation on sunken islands? BMC Evol Biol. 2015;15:196. https://doi.org/10.1186/s12862-015-0478-y.
Forni G, Plazzi F, Cussigh A, Conle O, Hennemann FH, Luchetti A, et al. Phylomitogenomics provides new perspectives on the Euphasmatodea radiation (Insecta: Phasmatodea). Mol Phylogenet Evol. 2021;155: 106983. https://doi.org/10.1016/j.ympev.2020.106983.
Wedmann S, Bradler S, Rust J. The first fossil leaf insect: 47 million years of specialized cryptic morphology and behavior. Proc Natl Acad Sci. 2007;104:565–9.
Bradler S, Buckley TR. Stick insect on unsafe ground: does a fossil from the early Eocene of France really link Mesozoic taxa with the extant crown group of Phasmatodea? Syst Entomol. 2011;36:218–22. https://doi.org/10.1111/j.1365-3113.2010.00564.x.
Nel A, Delfosse E. A new Chinese Mesozoic stick insect. Acta Palaeontol Pol. 2011;56:429–32. https://doi.org/10.4202/app.2009.1108.
Nel A, Delfosse E, Robillard T, Petrulevičius JF. An early winged crown group stick insect from the Early Eocene amber of France (Insecta, Phasmatodea). Syst Entomol. 2010;35:340–6. https://doi.org/10.1111/j.1365-3113.2009.00515.x.
Yang H, Shi C, Engel MS, Zhao Z, Ren D, Gao T. Early specializations for mimicry and defense in a Jurassic stick insect. Natl Sci Rev. 2021;8:nwaa056. https://doi.org/10.1093/nsr/nwaa056.
Willmann R. Die phylogenetischen Beziehungen der Insecta: Offene Fragen und Probleme. Verhandlungen des Westdtsch Entomol. 2001;2003:1–64.
Wipfler B, Letsch H, Frandsen PB, Kapli P, Mayer C, Bartel D, et al. Evolutionary history of Polyneoptera and its implications for our understanding of early winged insects. Proc Natl Acad Sci. 2019;116:3024–9. https://doi.org/10.1073/pnas.1817794116.
Bank S, Buckley TR, Büscher TH, Bresseel J, Constant J, de Haan M, et al. Reconstructing the nonadaptive radiation of an ancient lineage of ground-dwelling stick insects (Phasmatodea: Heteropterygidae). Syst Entomol. 2021;46:487–507. https://doi.org/10.1111/syen.12472.
Bradler S. Die Phylogenie der Stab- und Gespenstschrecken (Insecta: Phasmatodea). Species, Phylogeny Evol. 2009;2:3–139.
Klante H. Die Wandelnden Blätter—Eine taxonomische Revision der Gattung Phyllium Ill. (Insecta Orthoptera, Phasmatoptera). Zool Beiträge. 1976;22:49–79.
Zompro O, Größer D. A generic revision of the insect order Phasmatodea: the genera of the areolate stick insect family Phylliidae (Walking Leaves). Spixiana. 2003;26:129–41.
Fagua G, Condamine FL, Horak M, Zwick A, Sperling FAH. Diversification shifts in leafroller moths linked to continental colonization and the rise of angiosperms. Cladistics. 2017;33:449–66. https://doi.org/10.1111/cla.12185.
McKenna DD, Sequeira AS, Marvaldi AE, Farrell BD. Temporal lags and overlap in the diversification of weevils and flowering plants. Proc Natl Acad Sci. 2009;106:7083–8. https://doi.org/10.1073/pnas.0810618106.
Zhang Q, Wang B. Evolution of lower brachyceran flies (Diptera) and their adaptive radiation with angiosperms. Front Plant Sci. 2017;8:631. https://doi.org/10.3389/fpls.2017.00631.
Sellick J. The range of egg capsule morphology within the phasmatodea and its relevance to the taxonomy of the order. Ital J Zool. 1997;64:97–104. https://doi.org/10.1080/11250009709356178.
Shelomi M, Danchin EGJ, Heckel D, Wipfler B, Bradler S, Zhou X, et al. Horizontal gene transfer of pectinases from bacteria preceded the diversification of stick and leaf insects. Sci Rep. 2016;6:26388. https://doi.org/10.1038/srep26388.
Bank S, Cumming RT, Li Y, Henze K, Le Tirant S, Bradler S. A tree of leaves: phylogeny and historical biogeography of the leaf insects (Phasmatodea: Phylliidae). Commun Biol. 2021;4:932. https://doi.org/10.1038/s42003-021-02436-z.
Bradler S, Robertson JA, Whiting MF. A molecular phylogeny of Phasmatodea with emphasis on Necrosciinae, the most species-rich subfamily of stick insects. Syst Entomol. 2014;39:205–22. https://doi.org/10.1111/syen.12055.
Brock PD, Büscher TH, Baker E. Phasmida Species File Online. Version 5.0/5.0. 2021. Available from: http://phasmida.speciesfile.org.
Haas F. The evolution of wing folding and flight in the Dermaptera (Insecta). Acta Zool cracoviensia. 2003;46:67–72.
Ross ES. Biosystematics of the Embioptera. Annu Rev Entomol. 1970;15:157–72.
Schuh RT, Slater JA. The true bugs of the world (Hemiptera: Heteroptera): classification and natural history. Ithaca: Cornell University Press, Ithaca, New York, USA; 1995.
Thomas DB. The anocellate, flightless genus Lojus McDonald (Heteroptera: Pentatomidae). Ann Entomol Soc Am. 1997;90:569–74.
Brothers DJ. Phylogeny and classification of the aculeate Hymenoptera, with special reference to Mutillidae. Univ Kansas Sci Bull. 1975;50:483–648.
Narendra A, Ramirez-Esquivel F, Ribi WA. Compound eye and ocellar structure for walking and flying modes of locomotion in the Australian ant, Camponotus consobrinus. Sci Rep. 2016;6:22331. https://doi.org/10.1038/srep22331.
Boisseau RP, Büscher TH, Klawitter LJ, Gorb SN, Emlen DJ, Tobalske BW. Multi-modal locomotor costs favor smaller males in a sexually dimorphic leaf-mimicking insect. BMC Ecol Evol. 2022;22:39. https://doi.org/10.1186/s12862-022-01993-z.
Photographs of Pulchriphyllium pulchrifolium. Available from: http://www.phasmatodea.com/phyllium-pulchriphyllium-giganteum-tapah-hills.
Hausleithner B. Eine neue Phyllium-Art aus Malaysia (Phasmatodea: Phylliidae). Entomol Zeitschrift. 1984;94:39–42.
Cumming RT, Bank S, Le Tirant S, Bradler S. Notes on the leaf insects of the genus Phyllium of Sumatra and Java, Indonesia, including the description of two new species with purple coxae (Phasmatodea, Phylliidae). Zookeys. 2020;913:89–126. https://doi.org/10.3897/zookeys.913.49044.
Cumming RT, Le Tirant S, Teemsma SN. On the Phyllium of Peninsular Malaysia and Sumatra, Indonesia, with range expansions for currently known species, description of the previously unknown Phyllium (Pu.) abdulfatahi Seow-Choen female, and description of the new species Phyllium (Ph.) rubrum n. sp. from Peninsular Malaysia (Phasmida: Phylliidae). Faunitaxys. 2018;6:1–21.
Shi C, Shih C, Chen S, Ren D. Phasmatodea—stick insects and leaf insects. In: Ren D, Shih C, Gao T, Wang Y, Yao Y, editors. Rhythms of insect evolution: evidence from the Jurassic and Cretaceous in Northern China. 1st ed. Wiley-Blackwell: Hoboken; 2019. p. 165–73. https://doi.org/10.1002/9781119427957.ch13.
Lohrmann V, Waldren GC, Reiß M, Engel MS, Ohl M. An anocellar polistine wasp (Hymenoptera, Vespidae, Polistinae) from Texas. Zoosystematics Evol. 2016;92:251–5. https://doi.org/10.3897/zse.92.10548.
Eaton JL. Insect photoreceptor: an internal ocellus is present in sphinx moths. Science. 1971;173:822–3.
Seifert P, Smola U, Schinko I. Internal extraocular photoreceptors in a dipteran insect. Tissue Cell. 1987;19:111–8. https://doi.org/10.1016/0040-8166(87)90062-0.
Miller FE. A histological study of the eye and brain of a one-eyed bee. Ann Entomol Soc Am. 1936;29:66–9. https://doi.org/10.1093/aesa/29.1.66.
Dickens JC, Eaton JL. External ocelli in Lepidoptera previously considered to be anocellate. Nature. 1973;242:205–6.
Warrant EJ, Kelber A, Wallén R, Wcislo WT. Ocellar optics in nocturnal and diurnal bees and wasps. Arthropod Struct Dev. 2006;35:293–305. https://doi.org/10.1016/j.asd.2006.08.012.
Kerfoot WB. Correlation between ocellar size and the foraging activities of bees (Hymenoptera: Apoidea). Am Nat. 1967;101:65–70.
Newton AF, Thayer MK. Protopselaphinae new subfamily for Protopselaphus new genus from Malaysia, with a phylogenetic analysis and review of the Omaliine group of Staphylinidae including Pselaphidae. In: Pakaluk J, Ślipiński SA, editors. Biology, phylogeny, and classification of Coleoptera: papers celebrating the 80th birthday of Roy A Crowson. Muzeum i Instytut Zoologii PAN: Warszawa; 1995. p. 219–320.
Prud’homme B, Gompel N, Carroll SB. Emerging principles of regulatory evolution. Proc Natl Acad Sci. 2007;104:8605–12.
Engel MS, Davis SR, Prokop J. Insect wings: the evolutionary development of nature’s first flyers. In: Minelli A, Boxshall G, Fusco G, editors. Arthropod biology and evolution. Springer: Berlin Heidelberg; 2013. p. 269–98.
Abouheif E, Wray GA. Evolution of the gene network underlying wing polyphenism in ants. Science. 2002;297:249–52. https://doi.org/10.1126/science.1071468.
McCulloch GA, Oliphant A, Dearden PK, Veale AJ, Ellen CW, Waters JM. Comparative transcriptomic analysis of a wing-dimorphic stonefly reveals candidate wing loss genes. EvoDevo. 2019;10:21. https://doi.org/10.1186/s13227-019-0135-4.
Marshall CR, Raff EC, Raff RA. Dollo’s law and the death and resurrection of genes. Proc Natl Acad Sci. 1994;91:12283–7. https://doi.org/10.1073/pnas.91.25.12283.
Kim J, Sebring A, Esch JJ, Kraus ME, Vorwerk K, Magee J, et al. Integration of positional signals and regulation of wing formation and identity by Drosophila vestigial gene. Nature. 1996;382:133–8.
Cohen B, Simcox AA, Cohen SM. Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development. 1993;117:597–608.
Kutsch W, Kittmann R. Flight motor pattern in flying and non-flying Phasmida. J Comp Physiol A. 1991;168:483–90.
Liebherr JK. Brachyptery and phyletic size increase in Carabidae (Coleoptera). Ann Entomol Soc Am. 1988;81:157–63.
Evangelista DA, Wipfler B, Béthoux O, Donath A, Fujita M, Kohli MK, et al. An integrative phylogenomic approach illuminates the evolutionary history of cockroaches and termites (Blattodea). Proc R Soc B Biol Sci. 2019;286:20182076. https://doi.org/10.1098/rspb.2018.2076.
Mantovani B, Passamonti M, Scali V. The mitochondrial cytochrome oxidase II gene in Bacillus stick insects: ancestry of hybrids, androgenesis, and phylogenetic relationships. Mol Phylogenet Evol. 2001;19:157–63. https://doi.org/10.1006/mpev.2000.0850.
Passamonti M, Mantovani B, Scali V. Phylogeny and karyotype evolution of the Iberian Leptynia attenuata species complex (Insecta Phasmatodea). Mol Phylogenet Evol. 2004;30:87–96. https://doi.org/10.1016/S1055-7903(03)00156-8.
Buckley TR, Attanayake D, Bradler S. Extreme convergence in stick insect evolution: phylogenetic placement of the Lord Howe Island tree lobster. Proc R Soc B Biol Sci. 2009;276:1055–62. https://doi.org/10.1098/rspb.2008.1552.
Buckley TR, Attanayake D, Nylander JAA, Bradler S. The phylogenetic placement and biogeographical origins of the New Zealand stick insects (Phasmatodea). Syst Entomol. 2010;35:207–25. https://doi.org/10.1111/j.1365-3113.2009.00505.x.
Buckley TR, Attanayake D, Park D, Ravindran S, Jewell TR, Normark BB. Investigating hybridization in the parthenogenetic New Zealand stick insect Acanthoxyla (Phasmatodea) using single-copy nuclear loci. Mol Phylogenet Evol. 2008;48:335–49. https://doi.org/10.1016/j.ympev.2008.02.016.
Milani L, Ghiselli F, Pellecchia M, Scali V, Passamonti M. Reticulate evolution in stick insects: the case of Clonopsis (Insecta Phasmida). BMC Evol Biol. 2010;10:258. https://doi.org/10.1186/1471-2148-10-258.
Goldberg J, Bresseel J, Constant J, Kneubühler B, Leubner F, Michalik P, et al. Extreme convergence in egg-laying strategy across insect orders. Sci Rep. 2015;5:7825. https://doi.org/10.1038/srep07825.
Valero P, Ortiz AS. Description and DNA barcoding of a new Iberian species of Pijnackeria (Scali, 2009) from Sierra Nevada, Spain (Phasmida: Diapheromeridae). Zootaxa. 2015;4058:535–50. https://doi.org/10.11646/zootaxa.4058.4.5.
Büscher TH, Buckley TR, Grohmann C, Gorb SN, Bradler S. The evolution of tarsal adhesive microstructures in stick and leaf insects (Phasmatodea). Front Ecol Evol. 2018;6:69. https://doi.org/10.3389/fevo.2018.00069.
Cumming RT, Bank S, Bresseel J, Constant J, Le Tirant S, Dong Z, et al. Cryptophyllium, the hidden leaf insects—descriptions of a new leaf insect genus and thirteen species from the former celebicum species group (Phasmatodea, Phylliidae). Zookeys. 2021;1018:1–179. https://doi.org/10.3897/zookeys.1018.61033.
Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods. 2017;14:587–9. https://doi.org/10.1038/nmeth.4285.
Minh BQ, Schmidt HA, Chernomor O, Schrempf D, Woodhams MD, von Haeseler A, et al. IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol Biol Evol. 2020;37:1530–4. https://doi.org/10.1093/molbev/msaa015.
Chernomor O, von Haeseler A, Minh BQ. Terrace aware data structure for phylogenomic inference from supermatrices. Syst Biol. 2016;65:997–1008. https://doi.org/10.1093/sysbio/syw037.
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol. 2010;59:307–21.
Bouckaert RR, Vaughan TG, Barido-Sottani J, Duchêne S, Fourment M, Gavryushkina A, et al. BEAST 2.5: an advanced software platform for Bayesian evolutionary analysis. PLoS Comput Biol. 2019;15:e1006650. https://doi.org/10.1371/journal.pcbi.1006650.
Bouckaert RR, Drummond AJ. bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol Biol. 2017;17:42. https://doi.org/10.1186/s12862-017-0890-6.
Rambaut A, Drummond AJ, Xie D, Baele G, Suchard MA. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst Biol. 2018;67:901–4. https://doi.org/10.1093/sysbio/syy032.
Redtenbacher J. Phasmidae: Areolatae. In: Die Insektenfamilie der Phasmiden, vol. 1. Leipzig: Wilhelm Engelmann; 1906. p. 1–180.
Redtenbacher J. Phasmidae: Anareolatae (Phibalosomini, Acrophyllini, Necrosciini). In: Die Insektenfamilie der Phasmiden, vol. 3. Leipzig: Wilhelm Engelmann; 1908. p. 341–589.
Fritz SA, Purvis A. Selectivity in mammalian extinction risk and threat types: a new measure of phylogenetic signal strength in binary traits. Conserv Biol. 2010;24:1042–51. https://doi.org/10.1111/j.1523-1739.2010.01455.x.
Orme D. The caper package: comparative analysis of phylogenetics and evolution in R. 2013. Available from: https://cran.r-project.org/web/packages/caper/index.htm.
Harmon LJ, Weir JT, Brock CD, Glor RE, Challenger W. GEIGER: investigating evolutionary radiations. Bioinformatics. 2008;24:129–31. https://doi.org/10.1093/bioinformatics/btm538.
Pagel M. The Maximum Likelihood approach to reconstructing ancestral character states of discrete characters on phylogenies. Syst Biol. 1999;48:612–22. https://doi.org/10.1080/106351599260184.
Bush SE, Weckstein JD, Gustafsson DR, Allen J, DiBlasi E, Shreve SM, et al. Unlocking the black box of feather louse diversity: a molecular phylogeny of the hyper-diverse genus Brueelia. Mol Phylogenet Evol. 2016;94:737–51. https://doi.org/10.1016/j.ympev.2015.09.015.
Maddison WP, Slatkin M. Null models for the number of evolutionary steps in a character on a phylogenetic tree. Evolution. 1991;45:1184–97.
Revell LJ. phytools: an R package for phylogenetic comparative biology (and other things). Methods Ecol Evol. 2012;3:217–23. https://doi.org/10.1111/j.2041-210X.2011.00169.x.
Bollback JP. SIMMAP: stochastic character mapping of discrete traits on phylogenies. BMC Bioinformatics. 2006;7:88. https://doi.org/10.1186/1471-2105-7-88.
Pagel M. Detecting correlated evolution on phylogenies: a general method for the comparative analysis of discrete characters. Proc R Soc B Biol Sci. 1994;255:37–45. https://doi.org/10.1098/rspb.1994.0006.
Rabosky DL, Grundler M, Anderson C, Title P, Shi JJ, Brown JW, et al. BAMMtools: an R package for the analysis of evolutionary dynamics on phylogenetic trees. Methods Ecol Evol. 2014;5:701–7. https://doi.org/10.1111/2041-210X.12199.
Rabosky DL. Automatic detection of key innovations, rate shifts, and diversity-dependence on phylogenetic trees. PLoS ONE. 2014;9: e89543. https://doi.org/10.1371/journal.pone.0089543.
Beaulieu JM, O’Meara BC. Detecting hidden diversification shifts in models of trait-dependent speciation and extinction. Syst Biol. 2016;65:583–601. https://doi.org/10.1093/sysbio/syw022.
Song H, Béthoux O, Shin S, Donath A, Letsch H, Liu S, et al. Phylogenomic analysis sheds light on the evolutionary pathways towards acoustic communication in Orthoptera. Nat Commun. 2020;11:4939. https://doi.org/10.1038/s41467-020-18739-4.
Maddison WP, Midford PE, Otto SP. Estimating a binary character’s effect on speciation and extinction. Syst Biol. 2007;56:701–10. https://doi.org/10.1080/10635150701607033.
Acknowledgements
We are indebted to the countless people who provided numerous specimens over the last years, with special thanks going to Thomas R. Buckley (New Zealand), Nicolas Cliquennois (Madagascar), Sylvain Hugel (France), Gavin Svenson (USA), Michael F. Whiting (USA), Albert Kang (Malaysia), Bruno Kneubühler (Switzerland), Kristien Rabaey (Belgium), Valerio Scali (Italy), Katja Kramp (Germany), Savel R. Daniels (South Africa) and Donald Windsor (Panama). For information concerning the ocelli presence in certain taxa, we thank Royce T. Cumming (USA), Albert Kang (Malaysia), Nicolas Cliquennois (Madagascar) and Francis Seow-Choen (Singapore). We thank Katharina Henze for helping in the molecular lab. Photos were kindly provided by Bruno Kneubühler, Tim Lütkemeyer, Marco Niekampf and Christoph Seiler.
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was supported by the German Research Foundation (DFG Grants BR 2930/4-2, BR 2930/5-1 and BR 2930/7-1).
Author information
Authors and Affiliations
Contributions
SBa and SBr designed the project. SBr provided materials. SBa generated and analysed the data. SBa prepared the figures and wrote the manuscript with contributions from SBr. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Maximum Likelihood phylogeny based on the best-scoring tree with nodal support (UFBoot and SH-aLRT) at each node (Files S1 and S2). Lock symbols represent constrained clades (B1 constraints). Character states for wings and ocelli are depicted at tips for females and males. Figure S2. Ancestral state reconstruction for males. Analysis based on the BI tree with B1 constraints (see lock symbols at nodes; File S3). Nodal support values (< 1 posterior probability) depicted at each node. Stars represent the fossils used for calibration and numbering corresponds to Table S9. T, Timematodea; ASCH, Aschiphasmatidae; A, Agathemeridae; PSEU, Pseudophasmatidae; HN, Heteronemiinae; DIAPH, Diapheromerinae; HET, Heteropterygidae; GRA, Gratidiidae sensu Cliquennois [70]; CLI, Clitumninae sensu Cliquennois [70]; BAC, Bacillinae sensu Cliquennois [70]; PHA, Pharnaciinae + Prosentoria; LANCEO, Lanceocercata; X, Xenophasmina; ST, Stephanacridini; P, Palophidae; CLA, Cladomorphinae; LONCH, Lonchodinae, NEC, Necrosciinae; AFR/MAD, African/Malagasy group including Achriopteridae, Anisacanthidae, Antongiliidae sensu Cliquennois [70], Damasippoididae and Xylicinae sensu Cliquennois [70]; PHYLL, Phylliidae. Figure S3. Ancestral state reconstruction for males. Analysis based on the BI tree with B3 constraints (see lock symbols at nodes; File S5). Nodal support values (< 1 posterior probability) depicted at each node. Stars represent the fossils used for calibration and numbering corresponds to Table S9. Abbreviations as in caption of Fig. S2. Figure S4. Ancestral state reconstruction for males. Analysis based on the BI tree with B2 constraints (see lock symbols at nodes; File S4). Nodal support values (< 1 posterior probability) depicted at each node. Stars represent the fossils used for calibration and numbering corresponds to Table S9. Abbreviations as in caption of Fig. S2. Figure S5. Ancestral state reconstruction for females. Analysis based on the BI tree with B2 constraints (see lock symbols at nodes; File S4). Nodal support values and divergence times are identical to those in Fig. S4. Stars represent the fossils used for calibration and numbering corresponds to Table S9. Abbreviations as in caption of Fig. S2. Figure S6. Ancestral state reconstruction for the binary dataset of males using the IRR model. Analysis based on the BI tree with B2 constraints (see lock symbols at nodes; File S4). Nodal support values and divergence times are identical to those in Fig. S4. Stars represent the fossils used for calibration, and numbering corresponds to Table S9. Abbreviations as in caption of Fig. S2. Figure S7. Phylorate plots resulting from diversification rate estimation in BAMM. (A) Credible set of shift configurations with posterior probabilities (pp). Mean rate parameters are model-averaged across all samples assignable to a given configuration. (B) Phylorate plot of net diversification. Model shifts are depicted as symbols on branches, with star (= Euphasmatodea) and circle (= European Bacillinae) according to the best shift configuration. The red colouration indicates rate acceleration. The grey square represents an additional potential rate shift added from the second best configuration (clade includes Pharnaciinae + Prosentoria, Palophidae, Cladomorphinae, Xenophasmina, Stephanacridini and Lanceocercata).
Additional file 2: File S1.
Best scoring ML tree with UFBoot support values in newick format. File S2. Alternative ML tree with SH-aLRT support values in newick format. File S3. BI tree with B1 constraints in nexus format. File S4. BI tree with B2 constraints in nexus format. File S5. BI tree with B3 constraints in nexus format. File S6. Final concatenated supermatrix in fasta format.
Additional file 3: Table S1.
Phylogenetic signal. Results of phylogenetic signal estimations using D statistics and Pagel's lambda, and the comparison of the observed number of evolutionary transitions for each trait against a randomised character matrix. Wings = presence of wings (macropterous or micropterous), Flight = potential capability of flight (macropterous taxa), Ocelli = presence of ocelli. Table S2. Model testing. Results of testing the fit of different character evolution models to the binary wing dataset (datawings). ER, equal rates; ARD, all-rates-different; IRR, irreversible model disallowing the regain of wings. Table S3. Probabilites [%] for the three wing states for the major nodes. Comparison of the results for the B1, B2 and B3 constrained phylogenies. Note that Oriophasmata were not recovered as monophyletic in the B3 phylogeny and thus both Oriophasmata and Occidophasmata were excluded here. Table S4. Average number of transitions between states resulting from 300 iterations of stochastic character mapping. Based on phylogeny with B2 constraints and both 3-states and binary wing and ocelli datasets using the ARD and the IRR model. The asterisk indicates that depicted ranges show the confidence interval of 95% highest posterior density. apt, apterous; micro, micropterous; macro, macropterous. Table S5. Correlation test. Results of fitting Pagel's model to test for correlation between the evolution of wings and ocelli. Correlation was tested on the binary datasets of males and females with different settings for the root state. Variable x represents the wing state and y the ocelli state. For males, the correlation of ocelli was further tested for the three wing states. Significance in bold. AIC, Akaike Information criterion, AICw, Akaike weights. Table S6. Diversification analysis. Results of fitting 24 models of diversification including HiSSE, BiSSE and character-independent (CID) models. Analysis was based on the B2 constraint phylogeny and the binary wing dataset with outgroup taxa (Embioptera) removed. Best-fit model is depicted in bold. AIC, Akaike information criterion. Table S7. Information on taxon and gene sampling. GenBank accession numbers are provided for each specimen used in this study. For taxa retrieved from GenBank, an ID with consecutive numbering is used (GBXXX). In alphabetical order. Table S8. Best partitioning scheme (subsets) and best-fit substitution models determined with ModelFinder in IQ-TREE. Table S9. Fossil calibrations used for the divergence time estimation in BEAST2. The numbering (1–5) corresponds to the depiction of calibrated nodes in Figs. S2–S5. Table S10. Morphological data. Character matrix for females and males for ocelli and the 3-state and binary datasets for wings. Ocelli are coded as either absent or present and wings are coded in three states; apterous (0), micropterous (1) and macropterous (2). Missing data are shown as question mark. The binary datasets represent male wing states only and code presence of wings (datawings: wingless (0) vs. winged (1)) and flight capability (dataflight: flightless (0) vs. flighted (1)). Missing data for binary datasets are coded as absent (0). For taxa retrieved from GenBank, an ID with consecutive numbering is used (GBXXX).
Additional file 4.
Supplementary discussion of phasmatodean phylogenetic relationships.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Bank, S., Bradler, S. A second view on the evolution of flight in stick and leaf insects (Phasmatodea). BMC Ecol Evo 22, 62 (2022). https://doi.org/10.1186/s12862-022-02018-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-022-02018-5