
Abstract
Glyphosate (GLY), the most widely used herbicide in the world, is frequently detected in various environmental matrices, including soil, the foundation of agriculture. In practice, more than 2000 GLY-based herbicide (GBH) products are used, consisting of one or more active ingredients (AIs) and so-called “inert” co-formulants that increase the efficacy of the AIs. However, the focus of ecotoxicological assessments is mainly on AIs, while organisms are exposed to complex pesticide formulations under real-world conditions. Overall, the effects on non-target organisms indicate a broad range of biochemical and physiological modes of action, which contrasts with the general assumption that herbicides are specific and act only on target plants. Both GLY alone and GBHs have unintended side-effects on many terrestrial organisms, including non-target plants, microorganisms, insects, spiders, or earthworms, as well as vertebrates such as amphibians, reptiles, or mammals. One of the triggering mechanisms for these effects is oxidative stress with consequences on biochemical parameters and DNA damage. In addition, disruptions of various physiological, behavioral and ecological processes have been reported. Most studies have examined the short-term effects of a single application of GLY/GBH to a single species. However, the agricultural practice of applying GBHs two to three times during a cultivation season over an extended period of time, the interactions with other pesticides and agrochemicals applied to the same field, and effects on ecological interactions within the field and landscape are rarely considered. In the vast majority of cases, the toxicity of GBHs exceeds the toxicity of GLY, demonstrating that supposedly inert co-formulants are either toxic in their own right or interact and add to the toxicity of AIs. The chemical diversity of different GBHs and the non-disclosure of the co-formulants make it difficult to attribute effects to specific chemical substances within a GBH. Moreover, impurities in GBHs (e.g., heavy metals such as arsenic, chromium, cobalt) pose additional environment and food safety risks. These impacts are even more critical because GBHs are so widely distributed worldwide and interact with other pollutants and environmental stressors. Based on the available literature on terrestrial ecotoxicity, and given the drastic decline in biodiversity, we conclude that the continued high use of GBHs, resulting in increased exposure and risk, cannot be considered ecologically sustainable.
Similar content being viewed by others

Introduction
A significant part of various pesticides used in agriculture and other sectors for weed, pest and pathogen (e.g., viruses, fungi) control exert an increased chemical burden on the environment. The applied active ingredients (AIs) and also other components (co-formulants) present in commercial formulations, can reach the soil surface and waterbodies through infiltration, runoff, precipitation, leaching and as a result of outdated application technology. The ecotoxicity of chemical pollutants depends largely on various abiotic (e.g., the composition of different environmental matrices, pH, mineral content) and biotic (e.g., microbial composition and activity) factors, as well as the physico-chemical characteristics of the compounds such as water solubility, sorption capacity, and biodegradability, and the mechanisms of their biological actions. In addition, the possibility of toxic metabolite formation must also be taken into account [1, 2]. Pesticide residues are a significant environmental risk factor, and since glyphosate (GLY) is the most widely used AI in herbicides worldwide, its effects on non-target organisms has become the focus of a large and increased body of scientific research (5003 entries on PubMed and 14,557 entries on Web of Science as of June 2023).
GLY (N-(phosphonomethyl)-glycine), the phosphonomethyl derivative of the natural amino acid glycine [3] has captivated the interests of environmental and health scientists despite potentially equally or more toxic herbicides being used in large quantities [4]. Due to the large uptake of GLY-tolerant (GT) genetically modified (GM) crops, especially in North and South America, GLY-based herbicides (GBHs) have become globally the most widely used herbicides over the last two decades [5,6,7,8]. The global market of GBHs was estimated at approximately 4438.5 million USD in 2020 [9]. However, it is highly problematic to find up-to-date data on GBH use and sales, whilst details of sales for a given formulation are often hidden under the pretext of commercial sensitivity. Therefore, it is very difficult to obtain accurate information on GBH use within the European Union (EU) [10]. According to a European survey in 2017, GBH sales were about 44,250 tonnes of AI, while the average use of GLY in the agriculture was estimated as 0.24 kg AI ha–1 [11].
The mechanism of herbicidal action of GLY stems from its ability to inhibit the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), which is part of the shikimate pathway [12]. The shikimate pathway is responsible for the biosynthesis of essential aromatic amino acids (phenylalanine, tryptophan, tyrosine), and thus its inhibition by GLY in plants results in death [12]. Although the shikimate pathway is found in all plants giving GBHs a non-selective broad spectrum herbicidal activity, it is also found in most fungi and certain bacterial species and strains, but never in animals [12]. The lack of the shikimate pathway in animals, including humans, led to the claim that it is non-toxic to these classes of organisms. However, GLY and GBHs may still negatively impact soil, animal and human health by compromising the composition and function of fungal and bacteria species [13, 14]. In GBHs, the salts of GLY [e.g., GLY-isopropylammonium salt (GLY-IPA); -diammonium salt; -trimethylsulfonium salt] are used as AI to increase its solubility of AI in water while retaining its herbicidal properties [15, 16]. Commercial pesticide formulations invariably contain different types of additives, collectively called co-formulants, in addition to the declared AIs. The main purpose of pesticide co-formulation agents is to increase the effectiveness and bioavailability of the formulation by increasing the solubility, adsorption, and absorption of the AI [17]. For example, presence of surfactants such as POEA (a mixture of polyethoxylated tallow amines) promotes the penetration of GLY through plant cell walls [18].
Commonly, pesticide co-formulants are typically classified as inactive (inert) ingredients with respect to the main biological effect of the formulation. Therefore, a simpler environmental risk assessment (ERA) was considered sufficient for these substances compared to AIs [19, 20]. In addition, regulatory agencies obtain data on co-formulants in stand-alone studies rather than as part of formulations. Therefore, from a regulatory perspective the differential impacts of pesticides on ecosystems and peoples’ health are generally not due to the properties of co-formulants as stand-alone components, but to the ways in which the co-formulants modify the toxicity of AIs [21]. Recently, numerous studies have confirmed the high individual toxicity of co-formulants and the increased combined toxicity of AIs and co-formulants compared to the individual effect of AIs. This includes POEA (a mixture of polyethoxylated tallow amines) used for the manufacture of formulations of GBHs [22,23,24] and its substitutes [25, 26]. The use of POEA as a co-formulant in GBHs has been banned under current EU legislation because of its harmful effects on the environment [27]. However, an increasing body of evidence is showing that approved replacements of POEA such as Dodigen 4022, a propoxylated quaternary ammonium compound, is showing that they can also be toxic [25, 28].
The non-selective GBHs were originally used for pre-emergence weed control, but with the appearance of the GM GT crop varieties such as soybeans, maize, cotton, sugar beet and alfalfa the post-emergence use of GLY has also became possible. With the possibility of the post-emergence application of GLY, the amount of GBHs used for weed control purposes has increased dramatically [29]. So far, no GM GT varieties have been authorized for cultivation in the EU. In addition, another more recent adoption by farmers, which has led to a marked increase in GBH use, is its application as a pre-harvest desiccant on various crops particularly cereals such as oats and wheat, canola, pulses (beans, peas, lentils) and even potatoes [6, 30]. This involves spraying and intentionally killing the crop within 1–2 weeks of harvest to obtain more uniform drying of grains and an earlier time of harvest. This practice may make matters more convenient for farmers, but it also implies that GBH residues will enter the food chain in larger quantities as insufficient time is allowed for them to dissipate before harvest. The massive use of GBHs has led to environmental contamination, and GLY has become a ubiquitous contaminant [31].
During the regulatory process of pesticides, a detailed and harmonized two-tier registration system is applied in the EU [32]. According to Regulation (EC) 1107/2009, pesticide AIs are registered at the EU level by the European Commission, while the formulations containing pesticide AIs and co-formulants are authorized at the level of member states. This is a unique feature in the EU authorization of registered products [33]. Legislative steps towards Regulation (EC) 1107/2009, which is acknowledged as currently the most stringent pesticide authorization system in the world, have continued the program of substantially reducing the number of authorized pesticide AIs within the EU. As a result, between 1995 and 2009, the authorization of 700 AIs was discontinued [34], 22 AIs were deregistered between 2011 and 2013, and an additional 94 AIs were recommended for substitution between 2014 and 2021 [35]. In addition, as part of the European Green Deal [36], which aims to reduce pesticide use by 50% by 2030 through the replacement of "more hazardous" AIs with safer alternatives, harmonized risk indicators have been developed [37]. Among these, the risk indicator for authorized pesticides (HRI 1) has decreased by 22% compared to the baseline from 2011 to 2013. In contrast, the risk indicator for pesticides marketed under emergency permits (HRI 2) increased by 38% during the same period, which is an indirect but clear indication that despite stricter EU permits and bans, member states can still market outdated pesticide AIs under emergency authorization.
Herbicides are particularly affected by the tightening legislative requirements, yet a herbicide AI with a novel mode of action has not been placed on the market since 1992, which was the introduction of isoxaflutole, an inhibitor of the enzyme p-hydroxyphenylpyruvate dioxygenase (HPPD) [38, 39]. In 2020, Bayer announced that it will launch a new herbicide AI with a new mode of action on the market, but it is not expected until 2030 [39, 40].
Recently, several studies suggested the required revision or re-evaluation of the processes of pesticide authorization [41, 42]. However, despite these findings herbicides are only superficially tested for effects on non-target arthropods and generally do not include oral routes exposure ways (e.g., via food or air uptake). The indicated hazards are significantly relevant for risk assessment and suggest the need for the re-evaluation of current testing systems during the authorization process of pesticides [41]. Regulation and the ERA related to pesticides takes into account certain hazards but cannot centrally intervene in regulation and restriction. A major problem is that at the EU level, the focus is on the toxicology of the AIs, while organisms under real-world conditions are exposed to commercial formulations consisting of AIs and co-formulants. Furthermore, at the member state level, governments or government-related agencies must consider all stakeholders in their assessment, including manufacturers and patent holders [43].
It is important to point out that significant differences can be found in the global authorization of the pesticides, with nations having different requirements, guidelines, and legal limits for their application. Generally, pesticide legislation is stricter in more industrially developed countries compared to the developing nations. In several developing countries, often the lack of the required proper authorization and policies can be observed due to the lack of resources and scientific expertise to run pesticide analyses and evaluate environmental risk analysis [44, 45]. Another striking difference is that regulation of pesticides and the conditions of the risk assessment in the EU is stricter compared to the USA. Although international organizations have attempted to harmonize pesticide regulation by providing maximum residue limits (MRLs), these are not harmonized on a global scale. Thus, large differences can be found in the authorized in permitted MRL values. A globally harmonized pesticide regulation system would be required to increase global productivity and trade, and in addition to also support the protection of public and environmental health [44].
Generally, GLY is sprayed directly onto crop fields in both standard and no-tillage farming systems, where it is largely taken up by plants but with most being absorbed into the soil. In soil, GLY may be transported by surface water runoff, adsorbed to soil particles, enter groundwater by infiltration, or be assimilated by root transport of other non-target plants [46]. In soil, GLY is primarily metabolized to aminomethylphosphonic acid (AMPA), which is much more mobile than GLY [39, 47] and frequently detected in various environmental matrices [43]. Soil properties (e.g., mineral content, pH, phosphate concentration) greatly influence the leachability and mobility of GLY [26, 48,49,50,51,52]. Various GBHs with different co-formulants along with GLY have very different environmental fates (e.g., different half-lives and movement in soil and water). In most cases, the AI and the co-formulants almost certainly become separated relatively quickly, although very little is known about the half-lives and the environmental fate of the co-formulants. However, the presence of the GBH co-formulant POEA has been found to be widespread in mid-western US states where the cultivation of GM GT crops is most concentrated [53]. Furthermore, it has been shown that POEA can persist in soil along with GLY and AMPA and enter natural water courses [54]. Worryingly, the environmental impact of persistent POEA with GLY and AMPA is unexplored. The environmental concentration of GLY in the environment after GBH application and the occurrence of GLY in different ecological elements are strongly influenced by environmental and climatic conditions (e.g., soil composition and structure, intensity and frequency of rainfalls), as well as the timing and frequency of the pesticide treatments [50, 55]. Pesticide residue levels in or on food and feed of plant and animal origin are also regulated according to Regulation (EC) 396/2005 [56].
A high variability in detected GLY contamination levels in different environmental elements can be observed [43]. Focusing on terrestrial ecosystems GLY and/or AMPA were present in 45% of 317 topsoils collected across Europe originating from six crop systems, with a maximum concentration of 2 mg kg–1 [57] but reached as much as 8.1 mg kg–1 GLY and 38.9 mg kg–1 AMPA in no-till soils in Argentina [58]. GLY and AMPA was also ubiquitous in passive air samplers up to 3.2 µg sample–1 in Germany [59] and 116 ng sample–1 in ambient air in Austria [60]. The environmental concentration of GLY in soil highly depends on several abiotic and biotic factors environmental factors. Moreover, the circumstances and conditions of the selected analytical and sample preparation methods can highly affect the measured GLY levels [61]. Various soil characteristics such as mineral content, pH, soil redox conditions, phosphate content, and the level of organic materials as well as microbial composition and activity can affect the biodegradation and binding of GLY to different soil components. The rates of biodegradation and the adsorption of GLY determine the bioavailability and thus the effects of GLY on the non-target organisms. The contamination of soils highly depends on the intensity of agricultural use of GBHs [31, 62]. AMPA frequently appears in environmental samples via the degradation of GLY. However, the source of AMPA in the environment is not exclusively derived from GLY metabolism as it can also arise from phosphonate detergents used in washing powder [63].
Based on the high variability in detected GLY levels, the choice of environmentally relevant concentrations in ecotoxicological tests is not straightforward because habitat-specific contamination, species-specific exposure pathways, and environmentally relevant transformation under laboratory conditions are often not fully known or predictable. Due to the considerable production and use of co-formulants, these additives can easily enter the environment [54]. Therefore, the possible negative impacts of co-formulants on living organisms and ecosystem processes should be evaluated during the authorization and risk assessment of the pesticide formulations [64, 65].
Here we have summarized relevant up-to-date information and data reported since the EU Commission Directive 2010/77/EU to date on ecotoxicological concerns about GLY, its GBH formulations and co-formulants. Systematic searches were carried out in the Web of Science, Scopus, Science Direct and other scientific databases, and when necessary, references in the reports found were also followed. Results of non-public toxicity studies commissioned and funded by the industry and not included in the applications dossiers for re-approval [66] could obviously not be considered here either. In total, a reference database of over 500 scientific articles considering GLY or GBHs and ecotoxicology was compiled, and this review refers to those articles relevant to terrestrial ecosystems.
Ecotoxicity to terrestrial organisms and ecosystems
The effects of GLY and GBHs on terrestrial organisms have been reported in a large number of studies employing different species, at different developmental stages, using a variety of experimental designs (e.g., testing under laboratory/greenhouse or open field conditions) and endpoint measures (e.g., reproduction, behavior, developmental effects, enzyme activities). Most of the published studies (75%) were performed under laboratory conditions with the use of in vivo or in vitro test methods, in addition to the micro- and mesocosms. Other investigations obtained data from greenhouse (10%) and field experiments (15%). Laboratory tests are useful tools for the investigation of the different mechanisms of any toxicity observed, which is caused by the tested substances. However, studies under environmental settings such as field conditions are far more reflective of real-world scenarios compared to the laboratory-based experiments. The effects of GBHs have already been investigated in several studies on various terrestrial test organisms, such as springtails (e.g., Folsomia candida) [67], isopods (e.g., Porcellio dilatatus) [68], and earthworms (e.g., Eisenia fetida and Lumbricus terrestris) [69, 70]. Like all herbicides GLY easily enters the soil, where it is regarded as almost immobile on the basis of its sorption properties [71, 72]. However, GLY can leach out of soil due to its water solubility with other factors (e.g., phosphate concentration, pH) can contributing to increased soil mobility [72]. Thus, non-target soil-borne organisms can be harmed by GLY, its metabolites, or the co-formulants contained in GBHs via various direct or indirect exposure pathways [73, 74].
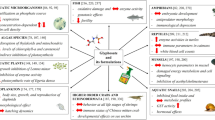
In order to investigate the potentially harmful effects of chemical compounds on non-target organisms, ecotoxicological tests always use model organisms, so-called surrogate species, that represent a specific group of organisms. In terms of regulation under Registration, Evaluation, Authorisation and Restriction of Chemicals (REACH), ecotoxicological tests must be conducted in compliance with the principles of good laboratory practice (Directive 2004/10/EC) and the international standards recognized by the EU Commission or the European Chemical Agency. During the approval of chemical compounds such as AIs in the EU, mandatory ecotoxicological tests must be performed for substances manufactured or imported in quantities of 1 tonne or more. Mandatory test organisms for the approval of either pesticide formulations or AIs in the EU consider five groups of non-target organisms: terrestrial vertebrates (birds and mammals), aquatic organisms (fish, water fleas, algae, and aquatic plants), terrestrial arthropods (including bees), soil organisms (earthworms, soil micro- and macroorganisms), and terrestrial plants [32]. The accurate and exact requirements according to the manufactured or imported quantities are summarized in Regulation 1907/2006/EC [75]. In the case of medicinal products, the prescribed ecotoxicological tests must always be carried out on the formulated products, while unfortunately during the authorization of the pesticides in certain cases they can rely solely on toxicity data of the AIs. In our review, in addition to outlining the ecotoxicological concerns associated with GBHs on terrestrial ecosystems and non-target organisms, we also summarize the effects of GLY and GBHs on specific indicator organisms such as bees and birds (Fig. 1), including the presentation of the combined effects of GLY and GBHs [76, 77] with other chemical compounds (e.g., pesticide AIs, co-formulants, heavy metals, and microplastics) or pathogens.

Main ecotoxicological effects of the herbicide active ingredient glyphosate and its commercial formulations on non-target organisms. Figure created with BioRender. (Red arrows showing up: increase; down: decrease; bi-directional: alteration)
The data on the effects of GLY on different soil organisms are complex, and sometimes the results are contradictory [78]. This is due to numerous factors that can influence the observed effects. The main factors affect the environmental fate (e.g., mobility, leaching), bioavailability and the effects on non-target organisms are the soil characteristics such as the water content, composition of different soils, for example the microbial community. In addition, mineral content, pH, soil redox conditions, phosphate content, and the level of organic materials highly influence the strength of sorption capacity and binding GLY and AMPA by soil particles. Furthermore, the variability in applied experimental design (e.g., tested concentration, exposure conditions, single or multiple exposures of the tested soils), and testing methods (e.g., testing under laboratory/greenhouse or field conditions, duration, routes of exposure) can result in contradictions in the observed effects. Additionally, the exact species tested and their developmental stage may have been significant factors [79, 80].
Effects on soil microbiota
Soil microbiota comprise a group of microscopic life forms that include bacteria, archaea, viruses, and eukaryotes like fungi. They perform many essential soil functions by regulating nutrient cycling, decomposing organic matter, defining soil structure, suppressing plant diseases, and supporting plant productivity, and they play an important role in biogeochemical cycles [81]. Several studies have shown that microbial communities are negatively affected by GBHs, while others have claimed the opposite. The results of most short-term and long-term experiments under field, under laboratory or greenhouse conditions have shown, that the composition of the soil microbial community is impacted by the use of GBHs.
In soil, GLY can act as an additional nutrient source and lead to stimulatory effects on soil biochemical parameters, such as dehydrogenase and β-glucosidase activity, and carbon and nitrogen content of microbial biomass [73, 74]. Maximum increases in enzymatic activity were observed between 18 and 37 days after treatment, while the increased β-glucosidase activity was still observed after 101 days [82]. After biodegradation of GLY, the utilization of GLY as a source of easily available carbon has been demonstrated on different bacterial strains by several studies [83,84,85]. GLY can be degraded by two metabolic pathways in bacteria. One metabolic pathway is oxidase activity that cleaves the carboxymethylene-nitrogen bond of GLY and thus converting it into AMPA and glyoxylate. In addition, C–P lyase can directly cleave the carbon–phosphorus bond to convert GLY into sarcosine. Both pathways of biodegradation use C–P lyase to cleave the C–P bond of AMPA [84]. The composition of soil microorganisms and fungi is also affected by GBHs [86, 87], with significantly lower numbers of colony-forming units (CFU) observed following treatment with the GBH Roundup PowerFlex at an application rate of 4.0 L ha–1 corresponding to 0.8 kg GLY AI ha–1 [87]. When the effects of Roundup PowerFlex (3.75 L ha–1 corresponding to 0.75 kg GLY AI ha–1—the recommended rate according to good farming practice [87]) on the individual fungal taxa was investigated, components present in GBH seemed to act as nutrients and favored growth of some fungal taxa (e.g., Colletotrichum sp., Cunninghamella sp., and Mortierella sp.), while other taxa (e.g., the fungus Mucor) were found to be completely absent [86]. Moreover, GLY can affect the nutrient composition in grapevine roots, leaves, grape juice, and xylem sap collected after 11 months of GLY application [87].
Compared to mechanical weeding, reduced root mycorrhization has been observed in grapevines treated with a GBH (Roundup PowerFlex—4.0 L ha–1 corresponding to 0.8 kg GLY AI ha–1) [87]. Effects of GLY on soil rhizosphere-associated bacterial communities were investigated in a study in which the rhizosphere of soybean and corn was sampled after 31, 37, 52, and 58 days [88]. After the application of the GBH (Roundup PowerMax), the relative abundance of Acidobacteria in the bacterial community was slightly decreased with 0.8–2.8% in the rhizosphere of the investigated crops. In contrast, the relative abundance of γ-Proteobacteria in both soybean and corn rhizosphere samples were slightly increased to 0.7 and 1.5%, respectively, by the increase of Xanthomonadaceae, suggesting that long-term application of GBH may affect the nutrient status of the rhizosphere [88]. Tolerance of the soil microbial community to GLY (in the form of AI or GBH) is not consistent with the history of the herbicide exposure to the soils at the investigated locations [89]. Functional diversity of soil microorganisms was very similar between as GLY (potassium salt) and GBH (Roundup Full II) treatments as reflected by catabolic evenness [89]. Furthermore, no difference in the overall microbial community composition and the relative number of microorganisms, such as Fusarium spp. was observed between GLY-treated plots and plots without GLY under field condition in well replicated (in time and space) studies [90]. In addition, other factors (e.g., geography, weather, and farming systems) had a much greater effects on soil microbes than the use of GLY [89]. However, GLY has been found to increase the colonization of certain Fusarium species on roots [91], which would facilitate the production of Fusarium mycotoxins [92]. This increased colonization can strongly influence crop productivity. In addition, several Fusarium spp. produce a wide range of mycotoxins which can endanger the health of animals and also humans. The most common mycotoxin groups produced by Fusarium species are zearalenones, trichothecenes, and fumonisins [93].
In a short-term greenhouse experiment, a 30% increase in soil microbial respiration was observed after the foliar application of a GBH (Roundup), which was concomitant with reduced bacterial phospholipid fatty acid content was determined for Gram+ and Gram− bacterial strains, but not for the saprotrophic fungi [94]. The GBH studied had little effect on the functional diversity of soil microbiota and only weak interactions between the GBH and soil depth, mycorrhiza, and earthworms were detected [94]. A single GLY treatment caused no changes in the microbial community in the silty clay loam soils, while more microbial biomarkers (e.g., aerobic and anaerobic bacteria markers) were reduced by the treatment in the silty loam soils [95]. In combination with the co-formulants (surfactants Activator-90, alkylphenol ethoxylate plus alcohol ethoxylate), significant differences were also found only in silt loam soils [95].
Greenhouse experiments were conducted to investigate the effects of a GBH on the diversity and composition of rhizosphere bacterial communities of transgenic GM GT Roundup Ready soybeans after two foliar herbicide applications. In addition to the decrease in abundance of Burkholderia sp., the effective resilience of the bacterial community associated with soybean roots after GBH application indicated stimulated bacterial diversity [96]. Furthermore, the increase in functional diversity of bacteria was only transient during a short-term incubation study of 21 days [97]. Most studies have examined the short-term effects of GLY on microbial biomass, rhizosphere, and terrestrial organisms at higher taxonomic levels, and mostly found only minor effects [98].
GLY can induce specific and widespread disturbance of the metabolome of Pseudomonas species in soil [99]. The GBH (Roundup R450 containing 450 g L–1 GLY) to filamentous soil fungus Aspergillus nidulans was toxic at doses 100 times below the recommended agricultural application rate with the GBH more toxic than technical GLY alone (LD50 = 90–112 mg L–1 GLY) [100]. Effects on A. nidulans included alteration of growth, defects in cellular polarity, delay in endocytosis and mitochondrial disruptions [100]. Similar effects were not found for A. section Flavi strains and A. niger [101], but adverse effects of GLY alone and in combination with N fertilization were observed on arbuscular mycorrhizal fungi (AMF) and soil enzyme activity [102]. At high doses of GBH (0.96 kg AI ha–1), an early and transitory stimulatory effect was observed, and negative effects of formulated GLY on soil fungal biomass were also detected [103]. In addition, the applied GBH affected the species richness and molecular profiles of soil fungal communities after double dosing (2 × 0.92 kg AI ha–1) or long-term application of the GBH. A temporal shift in species composition and polymorphism was detected from GLY-treated plots [103]. The addition of POEA resulted in a reduction of biomass growth of beneficial Pseudomonas species in soil by up to 60%, with a mixture of GLY and POEA also resulting in growth inhibition [104].
As mentioned, GBHs act as herbicides by inhibiting the enzyme EPSPS of the shikimate pathway, which is responsible for essential aromatic amino acid biosynthesis. The presence of the shikimate pathway in certain fungi and bacteria suggests that GLY/GBHs can also compromise the health and balance of microbiomes through the same mechanism. The finding that GLY and a GBH (Roundup MON 52276) does indeed inhibit the shikimate pathway in the gut microbiome of rats [28], has provided proof of principle that this may also take place in microorganisms in other spheres of life including soil where it could negatively impact beneficial fungal/bacterial-plant interactions within the rhizosphere. This possibility needs to be urgently explored as it has major ramifications for soil and plant health.
Further studies are needed to evaluate the detailed and potential effects of GLY and GBHs on soil microbial communities with several studies giving contradictory results, which appear to be primarily due to the diverse experimental conditions employed and testing methods. In summary, the contrasting results on the effects of GLY or GBHs on soil microorganisms highlights the difficulties in comparing or pooling data from studies that used different methods, different GBHs with unknown co-formulants, and a variety of study designs. Studies showed that meteorological and environmental parameters and soil characteristics modulate GLY/GBH-effects on microorganisms. The mechanisms by which mycorrhizal fungi respond to GLY or GBH exposure are not well understood, but it seems obvious that systemic herbicides such as GBHs immediately impact plant-associated mycorrhizal fungi. In addition, long-term effects and influences on soil parameters are hardly studied. Until a standard procedure for assessing these effects is established, a comparison of multiple assessment methods is advisable to avoid methodological bias.
Effects on terrestrial non-target plants
Non-target plants include monocots such as grasses and dicots (most others) and are defined as those that grow outside the cropping area [74]. All plants inside the cropping area other than the crop are recognized as targets and called weeds. From the soil, GLY can be taken up and transported by plant roots. In addition, GLY can have effects on non-target plants near agricultural fields due to various routes of exposure such as spray drift, rhizosphere contact or runoff [105, 106]. In a greenhouse study, application of GLY at 25% of the recommended field application rate negatively impacted 23 native plant species investigated as might be expected for such non-selective herbicide AI. Severe phytotoxicity or dieback was observed in 50% of species and growth reduction observed in 70% of the studied species. These results suggest that GLY may promote biodiversity loss of the natural environment, although some of these native species have been found to be resistant to GLY applications [107].
A review and meta-analysis of effects from GBH spray drift to non-target terrestrial plants and plant communities outside the intended area of application, showed no systematic differences between the responses of monocotyledons or dicotyledons [108]. However, the author of the study conjecture that in a certain cases wild plants may be less sensitive to GLY drift than domesticated plants [108], which agree with the result of a previous findings [109]. It is possible to assume that domestication, where crops are selected for various agriculturally important traits (e.g., faster growth, more effective resource allocation to seed and fruit), are not as resilient and thus less resistant to various stress factors [110], which may include greater susceptibility to the effects of herbicides (e.g., GLY) although more research is required to prove this hypothesis [108]. Moreover, it was assumed that restricting spray drift to a level < 5 g GLY ha–1 would protect the majority of higher plant species [108]. In Arabidopsis thaliana, an altered plant metabolome has been observed after GLY exposure due to the changes in the phyllosphere microbiome [111], which may affect plant resilience [112]. Residues of GBHs in soil can also potentially have indirect effects also on plant phytohormone homeostasis [113] with unknown consequences for ecosystems including plant–microbe and plant–insect dynamics [114].
Co-formulants used as formulating agents in GBHs such as POEA or APGs (alkyl polyglucosides) may have adverse effects on non-target plants. POEA, which facilitates the entry of GLY into the plant cuticle, was found to cause a significant reduction in the photochemical efficiency of the PS II photochemical system in treated kohlrabi, whereas this was not the case in apple leaves [115]. In contrast to POEA, the adverse effects of APGs were not detected based on the measured photochemical parameters [115]. However, GLY alone (4.05 g L–1) had only minor toxic effects on sprayed tomato plants (Solanum lycopersicum var. esculentum) compared to GBHs (Glyphogan, Roundup Grands Travaux plus, Roundup WeatherMax), which showed increased toxicity compared to GLY alone. These observations in all likelihood are due to the fact that GLY cannot penetrate plant cell walls in the absence of co-formulants and exert toxic effects. In addition, high toxicity of just POEA has also been demonstrated [18].
Another well-documented aspect of commercial GBH formulations such as Glyphogan, Roundup Classic and Medallon is the presence of toxic heavy metal impurities such as arsenic, chromium, cobalt, nickel, and lead or petroleum residues, which possibly originate from the manufacturing process. These inadvertent contaminants may also contribute to the adverse effects of GBHs as indicated by measures of endocrine disruption [18, 116, 117]. Due to the presence of these heavy metal impurities, the agricultural use of GBHs pose an additional risk to the environment and food safety by contaminating soils and crops [18]. So far, heavy metal impurities have been found in 11 GBHs, among them those registered for domestic or urban application in gardens. This could also explain findings that GLY, in contrast with the general belief, was only slightly toxic on plants at the recommended rates in agriculture [18]. The presence of arsenic in natural phosphate ores may represents a significant risk in case of the phosphate fertilizers [118] or even GBHs. During the production of these agrochemicals, the use of natural sources of phosphate can result in the presence of arsenic impurities in the manufactured formulations. Based on their potential serious health and environmental risks and consequences, the determination and evaluation of different impurities presented in commercial pesticide formulations should be undertaken with responsibility falling on manufacturers. Furthermore, a change in legislation should be enacted to make it mandatory for pesticide manufacturers to disclose the nature of any heavy metal contaminants and especially co-formulants and present in a given commercial formulation, which is currently kept hidden.
In conclusion, controlling or eradicating weeds is, of course, the aim of using GBHs. However, the contribution of GBHs among all herbicides used to the decline in the weed flora in Europe is unclear [119]. Also, to what extent GLY or GBHs also contributed to a decline in the arable weed seedbanks, with only a few species dominating and being able to adapt to herbicide use in agriculture is unknown [120]. Furthermore, it is remarkable that only higher plants are subjected to an ERA from GLY and GBH exposure with the sensitivity of terrestrial algae, mosses, ferns, and lichens is lacking [108]. There is an urgent need for studies investigating long-term effects of GBH use on native plant species and overall plant biodiversity. Because plants are important components of the food web in agricultural landscapes by providing pollen, nectar, or biomass, pollinators and herbivores are affected indirectly and therefore biodiversity as a whole is impacted. For example, the destruction of milkweed in US agricultural fields where GM GT crops are grown, has resulted in the almost extinction of the migrating monarch butterfly, whose larvae use these plants as their sole food source [121, 122].
Effects on terrestrial invertebrates
Nematodes
Nematodes are important components of the soil food web that cover all trophic stages from bacterial-feeding, fungal-feeding, root-feeding, omnivorous and predatory genera [123]. Studies of the effects of GLY on nematode populations have shown mixed results with some observing an alteration in community diversity [98, 124, 125] whilst another investigation found no change in the total number of nematodes or nematode trophic groups [126]. The soil-dwelling nematode Caenorhabditis elegans has emerged as an important experimental model organism in various scientific fields such as ecotoxicology, neurobiology, and developmental biology due to the ease of its maintenance, fully described development, short life cycle, and well-characterized genome. C. elegans as a test organism has a high sensitivity to different pollutants and represent important functional levels in the soil [127, 128]. Therefore, it is not surprising, that in the last decade, ecotoxicological studies have mainly used C. elegans as a test organism in the assessment of the adverse effects caused by GLY and GBHs.
Potential deleterious effects of a GBH (Touchdown) on the nervous system of C. elegans have been demonstrated [129]. The observed effects during the development, may cause individuals to be more susceptible to neurodegenerative diseases in adulthood [129]. The production of reactive oxygen species (ROS; reactive molecules and free radicals derived from molecular oxygen), and an enhanced oxidative stress response were detected in C. elegans exposed to the GBH Glifosato Atanor II (4.8 mM GLY) [130]. The induced oxidative imbalance in the treated nematodes resulted in DAF-16 activation leading to the increased expression of genes involved in antioxidant defense as indicated by biomarkers such as catalase activity [130]. In addition, ROS production and the enhancement of an oxidative stress response were observed in the GBH-treated nematodes [130]. Furthermore, inhibition of locomotion (10 µM GLY resulted in 87% inhibition) and fertility (23% and 93% inhibition caused by the levels of 0.01 µM and 10 μM GLY) were observed after exposure a Roundup GBH, although, the growth of the nematodes was not affected by the treatments [131]. Both GLY and its formulation Roundup triggered increased convulsive behavior in C. elegans by targeting receptors in the central nervous system [132].
Fields in northern latitudes present a special case as the period over which agents that can decompose GLY is shorter than in more southerly regions and thus lead to less degradation of this compound. A study comparing the effects of a Roundup GBH and hoeing on the abundance of nematodes found that killing plants by hoeing had drastic effects on soil fauna and functioning, and apparently, distinguishing these effects from direct GLY effects is profoundly important when evaluating GLY risks in soils [126]. In contrast, the effects of Roundup on nematodes were minor and transient and no GLY residues were not found in the soil at the end of the experiment [126]. A meta-analysis of nematode responses to different herbicides suggests that the abundance of bacterial-feeding, plant-parasitic and omnivorous nematodes increase in herbicide treated soils, while those of fungivorous and predatory nematodes decrease [124]. However, only one of the studies evaluated the effects of GLY, which showed that the total nematode abundance and the proportions of trophic groups in conventionally tilled and no-till soybean and corn fields in the USA, did not differ under GLY application (1.12 kg ha−1 applied once or twice during the growth period) from those observed under application of other herbicides [126]. A short-term laboratory experiment studying outcomes from a single GLY application on the nematodes of Australian banana plantations, showed no significant effects of GLY application on the total number of nematodes or nematode trophic groups [98].
A field experiment investigating the response of plant-parasitic nematode communities upon application of GLY to GM GT soybean and maize found no effect on Meloidogyne incognita, M. javanica and Pratylenchus spp. numbers [133]. Furthermore, no significant differences were observed between the GLY-treated and non-treated plots for Meloidogyne population densities [133]. In contrast, glasshouse and field studies conducted in South Africa found higher root-knot and lesion nematodes under GM GT soybean cultivars compared to previous surveys conducted 19 years previously indicating effects from GBH treatment [134].
Despite an increasing concern of consequences of using vast amounts of GBHs in agroecosystems and elsewhere, their effects on nematodes have been little studied. Given the diverse lifecycles of nematodes from predatory to parasitic to crop plants and agricultural pests non-target effects of GBHs on nematodes can have broad consequences for agroecosystem functions and deserve a more detailed investigation. Currently, it is not clear whether GLY/GBHs have direct effects on nematodes or rather indirect effects via reduced root carbon flow and increased dead plant material. Also, it would be important to examine the whole decomposer food web, and it is possible that soil microorganisms or soil invertebrates are more sensitive to GLY or GBH and alter food web interactions.
Collembolans, terrestrial insects and spiders
Collembolans or springtails are a diverse group of small, omnivorous arthropods that interact with soil microorganisms and affect the decomposition of organic matter in soil [135]. Terrestrial insects make up the bulk of terrestrial diversity, and reports of insect declines suggest that 40% of insect species in temperate countries may face extinction over the next few decades due to habitat losses, invasive species, climate change, and pollution with agrochemicals [136,137,138]. Spiders are a very diverse order that have a very important role in agroecosystems as they prey on insects of which some are pests [139].
GLY and GBH residues in soils may have adverse effects on non-target terrestrial organisms [98]. The EC50 value for reproduction of the Collembola Folsomia candida exposed to GLY in a natural soil over 4 weeks was 4.63 mg GLY kg soil–1. Lower EC50 values (0.87–1.49 mg kg of soil–1) were reported for exposure to the GBH Montana (containing POEA as co-formulant) indicating higher toxicity in the presence of POEA [67]. Moreover, the investigated GBH caused abnormal cellular respiration and lipid metabolism, leading to oxidative stress and damage to biological life cycle stages such as molting [67]. At the recommended field dose, GBH Montana did not affect the avoidance behavior and reproduction of the F. candida and the earthworm Eisenia andrei. However, collembolans were more sensitive to the GBH [140].
In a greenhouse experiment, GBHs (Roundup LB Plus, Touchdown Quattro, Roundup PowerFlex) and their AIs (isopropylammonium, diammonium or potassium salts of GLY, respectively) increased the surface activity of the springtail Sminthurinus niger at the recommended dosages [51]. The detected activity of springtails was higher for the GBHs than for the AIs. It was also found that soil organic matter (SOM) can have an impact on the effects of GLY AI and GBHs with the stimulating effect of GLY treatments on springtails generally greater in soil with a higher SOM content (4.1%) compared to low (3.0%) SOM content indicating significant interaction between the treatments and levels of organic matter. A significant interactive effect between SOM levels and GBH treatment on soil moisture was also observed, but no such interactive effect was observed between SOM levels and AIs [51]. Such interactions between effects of GLYs/GBHs and soil organic matter is important as great efforts are taken to increase organic matter levels [141].
An investigation of the preference of F. candida between GLY-treated and uncontaminated areas, the results showed a preference for the uncontaminated areas [142]. Investigations in vineyards in Romania showed that collembolan activity density in untreated soil was positively correlated with tillage and herbicide application in neighboring rows treated with a combination of a GBH and flazasulfuron (GLY acid at 360 g L–1 and 5% flazasulfuron at 0.07 kg L–1) applied at 1.6 L ha–1 [143]. Collembolans was interpreted as the result of a GBH/flazasulfuron-induced decrease in competitors and predators of collembolans, and a stimulation of microorganisms by nutrients contained in the herbicide.
Investigations of possible adverse effects of GLY and GBHs on spiders have shown no effects of GLY alone (GLY-IPA 0.18–2.16 kg ha–1) on Lepthyphantes tenuis spiders [144], but increased mortality and altered behavior were observed in Pardosa milvina and Neoscona theisi spiders exposed to GBHs [145, 146]. Synergistic effects of GBH (Roundup Klasik Pro) and tank-mixing adjuvants (Wetcit and Agrovital surfactants), which are commonly added to formulations prior to application, were observed in Pardosa wolf spiders. Significantly lower predator activity was observed in spiders exposed to the pure surfactants and the mixture of GBH and tank mix adjuvants. However, no effects from just Roundup Klasik Pro treatment on predator activity were observed [147].
Increased levels of antioxidant capacity and gene expression of the antioxidant defense system were observed in the fruit fly Drosophila melanogaster exposed to a GBH (Roundup) with the results indicating an early activation of the antioxidant defense system to enhance prevention of damage associated with the presence of ROS [148]. The effects of GBH exposure were sex-, age-, and temperature-dependent in-field crickets (Gryllus lineaticeps) [149]. Moreover, the additive effects of temperature and GBH exposure on adult growth and feeding rates were demonstrated although no adverse effects from GLY alone were observed [149]. It has also been found that GLY and AMPA can inhibit phenoloxidase activity in Galleria mellonella moths, resulting in inhibition of melanization and a suppressed melanin-based immune response in the exposed moths [150].
The effects of GLY and GBHs on mosquitos have also been investigated. GLY exposure resulted in dose-dependent deleterious effects on the learning ability of the Aedes aegypti mosquito larvae at field-realistic doses [151]. It has also been reported that GLY treatment resulted in altered composition but not in the density of midgut microbiota (Asaia and Enterobacteriaceae populations) in Anopheles gambiae [150]. Furthermore, the survival of adult A. gambiae females was increased at the tested low GLY doses compared to the control [150].
GLY and GBHs have only cursorily tested for effects on non-target arthropods. In laboratory experiments, lacewings (Chrysoperla carnea) larvae were exposed to a GBH (Roundup WeatherMax, 540 g L–1 GLY) via food at concentrations below the recommended field application rate [41]. This resulted in developmental arrest, impaired cocoon formation and high mortality of this species with a clear dose–response relationship.
In summary, there are comparatively few studies considering the effects of GLY and GBHs on terrestrial non-target organisms. Given that collembolans are considered bioindicators of soil health, comparatively few studies are available testing their response to GLY or GBHs. Generally, effects on collembolans appeared to be due to a combination of direct effects and associated changes in plant cover and soil parameters. Most investigations of GLY/GBH effects on invertebrates have focused on insect pollinators such as honeybees or wild bees. An exception is a recent study looking at the effects of chronic oral exposure of lacewings [41]. This study is highly significant as lacewings are important predators in many agricultural crop fields where they feed on aphids, chinch bugs, mealybugs, scales, whiteflies, leafhoppers, lepidopterous eggs and larvae, and mites [152]. Clearly, more studies are needed to investigate to what extent GLY/GBHs affect food webs and ecological interactions in agroecosystems.
Honeybees and wild bees
Insect pollinators underpin global food production, but they are suffering significant declines across the world with pesticides such GLY/GBHs are thought to be important drivers in this process [153, 154]. Generally, bees may come in contact with GLY/GBH by being sprayed directly, by touching a sprayed plant part, or orally via foraging for nectar and pollen [155, 156]. GLY and GBH effects have already been covered in excellent reviews recently including effects on their behaviors, growth and development, metabolic processes, and immune defense, providing a detailed reference for studying the mechanism of action of pesticides [157, 158]. Most of a sets of 16 publications show that the treatments with GLY caused higher mortality of bees and therefore, GLY and GBHs can be considered toxic to honeybees as well as wild and solitary bees [157]. GLY concentrations detected a few days after application ranged from 2.8–31.3 mg kg–1 in nectar and 87.2–629.0 mg kg–1 in pollen. The detected GLY levels decreased with time being strongly affected by environmental conditions and plant species [159, 160]. The most influential parameters include various abiotic (e.g., the composition of different environmental matrices, pH, mineral content, hydrogeological conditions) and biotic (e.g., microbial composition and activity, vegetation) environmental factors, the climatic conditions (e.g., weather, rainfalls), and the circumstances of farming systems (e.g., timing and frequency of the GBH applications). In addition, the environmental fate of GLY also depends on the presence of the various co-formulants (e.g., anti-formulating agents, surfactants) as well. The application of GBH as a pre-harvest crop desiccant, can result in the contamination of non-target wild plant pollen and nectar. The presence of GLY was proven in the samples collected two to seven days after the GBH application, and thus the flowering of the contaminated plants serves as a significant exposure route for pollinators. However, after two or more months, GLY was no longer detected in the collected samples [161]. Foraging bees exposed to contaminated pollen and nectar can transport the herbicide into hives, thus residues of GLY can be appear also in honey [162]. A field study evaluated GLY contamination of nectar and pollen of oilseed rape and blackberries grown in hedgerows [161]. Pollen loads were taken from honeybees and bumblebees foraging on the crop at the same time and GLY was detected in blackberry nectar and pollen samples that were taken within 7 days after GBH application on oilseed rape as a desiccant. AMPA was not detected in any samples [161].
Studies of GLY/GBH effects on honeybee sensory and cognitive abilities report negative impacts on associative learning processes of foragers, cognitive and sensory abilities of young hive bees and promotes delays in brood development [159]. Due to the widespread use of GBHs, exposure to GLY is inevitable for several bee species. The consequences of GLY exposure on honeybees including effects on their behavior, growth and development, metabolic processes, and immune defenses, have been demonstrated in several studies [158]. Direct toxicity of a GBH was demonstrated in bee species (Apis mellifera and Hypotrigona ruspolii) under laboratory conditions [163]. The dose-dependent mortality of bees was observed after the contact with plants freshly sprayed with the GBH (Sunphosate 360 SL) at the recommended concentration (10 mL L–1) [163]. GLY is known to interact with various bacteria, including gut bacteria. Disruption of gut bacteria makes bees more vulnerable, leading to various negative health outcomes including decreased resistance to pathogens, which in turn results in the weakening or complete loss of bee colonies [164,165,166]. When tested alone, GLY (1.5 mM in sugar syrup) itself, but not AMPA, significantly altered the gut microbiota of honeybees [167]. Exposure for 5 days to GLY AI and a GBH at field relevant concentrations (0.01–1 mM), resulted in reduced relative abundance of a beneficial bee gut bacterium (Snodgrassella) by 50% compared to controls. However, this difference was not significant at days 3 and 7 post-exposure indicating the resilience of the microbiota population. During the 7-day post-exposure period, reduced survival rates were detected in bees exposed to GBH at the concentration of 1 mM GLY equivalent concentration [168]. The chronic ingestion of GBHs (exposure via food uptake for 10 days) did not cause any effects on the food consumption in exposed Melipona quadrifasciata bees indicating that they do not avoid the contaminated food [169]. In addition, after exposure the height of midgut digestive cells was not affected. However, in bees fed on food contaminated with GBHs, differences were detected in the ultrastructure of digestive cells and impaired midgut digestive cell cohesion was observed due to the disorganization of the smooth septate junctions between cells, which was probably caused by the presence of the POEA co-formulant [169]. Similar to many other animals, the honey bee A. mellifera relies on a beneficial gut microbiota for regulation of immune homeostasis. Honey bees exposed to GBHs or antibiotics, usually exhibit dysbiosis and increased susceptibility to bacterial infection [170]. Considering the relevance of the microbiota–immunity axis for host health, the available data suggest that exposure to GLY/GBHs could potentially negatively affect other components of the bee physiology, such as the immune system.
A study has also investigated three GBHs (the consumer products Roundup Ready-To-Use and the agricultural products Roundup ProActive and Weedol), and a herbicide without GLY (Roundup No Glyphosate) for their effects on bumble bees Bombus terrestris audax [171]. After spraying the herbicides at label recommended rates, the bees suffered between 30% (Roundup ProActive) and 94% mortality (Roundup Ready-To-Use) over 24 h. Weedol did not cause significant mortality, suggesting that GLY alone is not the cause of the heightened dearth rates. The observed 96% mortality caused by Roundup No Glyphosate supports this conclusion [171]. Roundup products were found to cause comprehensive matting of bee body hair, suggesting that co-formulants in the products, may cause death by incapacitating the animal’s gas exchange system. These results demonstrate that Roundup products pose a significant hazard to bees, in both agricultural and urban systems. Moreover, the toxicity of GBHs was dependent on the different additives in the commercial herbicide formulations [171].
In a choice test where bumblebees could forage on plants treated with GLY AI, it was observed that GLY and AMPA residues were present in collected pollen with these insects indiscriminately foraging on both GLY-treated and untreated plants [172]. However, the time spent on individual flowers was slightly lower on GLY-treated plants but this did not affect the bumblebees' choice overall. Floral resources remained present in plants for at least 5 days after lethal treatment with GLY and that GLY residues were present in pollen for at least 70 h posttreatment. These results show that bees could be exposed to herbicide in the environment, both topically and orally, by foraging on plants in the period between herbicide treatment and death.
With regard to solitary wild bees (Megachile spp.), significant negative effects of the GBH Roundup ControlMax were detected on the reproductive success, with exposure at the recommended field application of 8 g AI L–1 or twice this level, halving the probability of finding brood cells [173]. In addition, a decrease in the rate of full larval development of 25% was also detected. Survival at immature stages of development was significantly reduced compared to the unexposed controls, whilst survival in treated nests was lowered by 14% and 4%, respectively, depending on the tested concentration [173]. In another study investigating developmental effects, delayed molting and lower larval weight were observed in honeybee (A. mellifera) broods that consumed GLY-containing feed (1.25–5.0 mg L–1 food) [174]. Furthermore, metabolomic analysis of the effects of GLY (7.12 mg L–1) on A. mellifera showed significant downregulation of several essential amino acids (leucine, lysine, valine, and isoleucine) was observed two days after oral exposure, which indicates a major metabolic perturbation [175].
Sublethal doses of GLY (2.5, 5, and 10 mg L–1 corresponding to 0.125, 0.250, and 0.500 μg bee–1) can disrupt the honeybee navigation, while exposed bees spend more time flying home compared to controls [176]. Similarly, significantly reduced bee navigation performance, decreased motor skills, and abnormal movement were detected following exposure of A. mellifera to a non-lethal range of a GBH (Roundup, 5–10 mg L–1 GLY equivalent concentration) in a dose-dependent manner [177]. At concentrations of GBH below recommended use for weed control (0–3.7 mg GLY acid equivalent L–1) and as measured in environmental elements (1.4–7.6 mg GLY acid equivalent L–1), reduced sensitivity to sucrose and nectar reward was observed with a concomitant decrease of elemental learning performance and short-term memory retention [178]. Acute field-realistic exposure to a GBH (0.1 µL Roundup Gold containing 0.045 mg AI) resulted in impaired fine-color discrimination and long-term memory in the exposed bumblebees (B. terrestris), which may reduce individual and colony fitness, although the olfaction or general vision of the exposed bees were not affected due to the exposure [179]. After oral exposure by ingestion to sublethal GLY concentrations (2.5 and 10 mg L–1), delayed brood development and deterioration of associative learning processes of foragers, as well as altered cognitive and sensory abilities of young bees were observed [159]. After an 11-day exposure to a GBH Roundup formulation at recommended use concentrations, a significant decrease in water and sucrose sensitivity of honeybees was observed. The tested GBH also negatively affected olfactory, memory, and climbing abilities in honeybees was observed [180]. Ingestion of 50 ng or more of GLY reduced both the frequency of sleep periods and the antennal activity of honeybees, which is probably related to induced metabolic stress [181]. After long-term oral GLY exposure (via feed consumption: 5 mg L–1 GLY in sugar solution), both individual and collective thermoregulation of bumblebees was altered [182]. Although the effect was minimal at the level of the individual, a significant difference was observed at the collective level. Moreover, a decrease of more than 25% was observed in required high brood temperatures during periods of resource limitation [182].
Based on the results of ecotoxicological testing on bees, the use of GBHs would appear to pose a serious health risk to these pollinators. GLY can be directly and also indirectly toxic to bees even at concentrations that can be frequently detected in the environmental samples. Exposure of bees to very low and environmentally relevant concentrations can affect, among other parameters, the composition of gut microbiota, reproduction success, thermoregulation, and foraging abilities, which collectively can weaken the individual’s and colony’s potential to survive. Studies of GLY/GBH effects on wild bees and other flower-visiting insects are scarce. GBHs and other herbicides are the most widely applied type of pesticides and are broadly considered ‘bee safe’ by regulatory bodies who explicitly allow their application directly onto foraging bees. As pollen samples contain numerous pesticide residues [183], more focus should be applied on interactions between GLY/GBH and other agrochemicals. Generally, results of the studies conducted to date reveal that the manner of GLY/GBH contact (ingestion or contact), the phase of the biological life cycle (adults or larvae), and the dose (ecologically relevant dose and recommended by the manufacturer) are crucial elements to consider. Overall, publications reporting the toxicity of GLY/GBHs on bees are still scarce, for both lethal and sublethal effects, and mainly for stingless and solitary bee species.
There is strong evidence that pesticide co-formulants, particularly those in GBHs, may contribute to global bee declines. Thus, we recommend that government regulators make it mandatory for manufacturers to publicly fully disclose co-formulant identities to enable a scientific assessment of their role to insect declines. Another key knowledge gap is the routes of exposure of bees to GLY/GBHs in the environment, including whether they may forage on treated plants before they die. After spraying, residues have been found in nectar and pollen collected by bees foraging on treated plants. This dietary exposure to GLY and GBH with co-formulants could pose a hazard for flower-visiting animals including bees, and severely compromise to conduct pollination of crops and other plants. In order to gain further insight into the potential extent of translocation within both plants and soil when a crop is desiccated shortly before harvest with GBHs, and the potential impact of this practice on bees, systematic monitoring needs to be undertaken.
Snails and slugs
Snails and slugs serve a variety of functions within an ecosystem, they are important herbivores, are decomposers of organic material, are a food source for arthropods and vertebrate, and can even be pollinators [184, 185]. Under laboratory conditions, long-term exposure to the GBH Bypass at the recommended field doses as well as a tenfold higher dose had no significant effects on the survival and growth of Helix aspersa, although, the GBH treatment resulted in reduced development of the albumen gland during maturation of the snail genital tract [186]. Importantly, the accumulation of GLY and its metabolite AMPA in snail tissues is well documented [186, 187]. In snails (Archachatina marginata), a concentration-dependent weight loss was observed after exposure to GBH Grassate [188]. The genotoxic potential of GBH Roundup Flash (30 mg L–1 of GLY equivalent concentration) was demonstrated in land snail embryos (Cantareus aspersus) exposed to throughout embryonic development [189]. Generally, responses of snails to GLY/GBHs are poorly investigated whilst effects on slugs have not been studied at all. As some slug species, such as Arion spp. are important invasive species in agricultural crops [190, 191], it would be interesting to see if they are more resistant to direct and indirect GLY/GBH exposure than native slug species.
Earthworms and enchytraeids
In many soils, earthworms represent the largest fraction of soil biomass. Earthworm well-being is a key factor in soil health, as they are the major decomposers of organic matter, and have a significant role in soil aeration, infiltration, structure, nutrient cycling, and water movement [192]. Enchytraeids resemble very small earthworms that can be found in various habitats, including agricultural land and, like earthworms, they influence soil structure and organic matter dynamics by affecting microbial communities [193].
Earthworms have also been used as bioindicators for the effects of agrochemicals [194]. Several independent studies indicate that both GLY and GBHs pose direct and indirect effects to earthworms [73, 74]. The earthworms most commonly used for ecotoxicological testing are the Eisenia species, so the discussion of the effects starts with these organisms. In soil treated with GLY, the weight of E. fetida was reduced by 50%. Although, the toxicity of GBHs is highly dependent on the co-formulants present [195]. The difference in toxicity of different GBHs was demonstrated in E. andrei under laboratory conditions, with a 4.5-fold difference in LC50 values of GBHs Roundup FG and MON 8750 [196]. DNA and lysosomal damage were detected at low concentrations of Roundup FG (14.4 mg AI acid equivalent m–2). At sublethal concentrations, a concentration-dependent weight loss was observed, consistent with the effects of GLY as an uncoupler of oxidative phosphorylation [196]. These observations agree with the results of a previous investigation performed on E. fetida exposed to GLY-treated soil [197]. The exposure to 26.3 mg kg–1 GLY isopropylammonium salt (IPA) in soil compost caused a 15–26% reduction in body mass of E. fetida, and resulted in 22–33% faster death of earthworms in a stress test compared to the control [70]. GLY-IPA caused a 15–26% reduction in biomass and survival of exposed earthworms compared to the control. In contrast, significant changes were not detected in earthworms exposed to GBHs (Roundup-Ready-To-Use III and Roundup Super Concentrate) according to the measured endpoints [70]. Results indicated that nitrates and phosphates in the GBHs offset the toxic effects of GLY by the stimulating microbial growth and its consequent biodegradation [70].
Another study using the earthworm Pheretima eguana, found that GBH exposure caused aneugenic effects (spindle fiber disfunction, increased micronuclei in coelomocytes), while the risk of gene mutation and its genotoxicity via a clastogenic (DNA damage) mechanism was not observed [198]. Sensitivity of E. fetida to the GBH Roundup-Ready-To-Use III (26.3 mg GLY kg–1 dry soil, 29 days) depended on soil temperature, whilst exposure to the GBH resulted a temperature-dependent survival [199]. E. fetida exposed to GBH Rodeo XL (1080 g AI ha–1) incorporated slightly less straw into the soil than earthworms in soil without GBH treatment [69]. One week after exposure to GBH Roundup (60.7 mg GLY kg–1 soil), E. fetida showed the greatest decrease in survival in a stress test, while after 2 weeks the greatest decline was observed in soil microbial biomass, although both investigated test communities recovered by the third week [200]. In earthworms sampled from vineyards treated with GBH (Roundup PowerFlex—4.0 L ha–1 corresponding to 0.8 kg GLY AI ha–1), density, reproduction, biomass, and surface casting activity were not different compared to the control samples and furthermore, the GBH treatment did not alter litter decomposition in soil [87]. E. fetida tolerated GLY exposure and its presence improved GLY and AMPA removal in biobed [201].
In E. fetida and E. andrei earthworms exposed to two GBHs (Touchdown Quattro, Roundup PowerFlex), their AIs (GLY ammonium and potassium salt) and the co-formulant APG presented in the GBHs, biomass and reproduction were significantly affected by the tested substances and temperature (15 °C and 20 °C) also with significant interaction of the two factors [26]. Generally in this study, more pronounced effects of GLY AIs was observed compared to the corresponding GBHs. Moreover, cocoon production was increased and stimulated by APG alone [26], which may be explained by the hormesis effect [202]. However, no effects of GLY/GBHs were detected on the avoidance behavior of the exposed earthworms. It should be borne in mind that most studies were conducted on Eisenia species, which do not actually live in agricultural soils, thus these species do not normally come into contact with GBHs.
Adverse effects of a GBH (Roundup 360) on survival rates and the number of cocoons were observed in earthworm species as a consequence of the exposure to GBH Roundup 360 in different concentrations (0, 0.5, 29, 57.9, 116 kg ha–1 of GLY equivalent concentration) in vineyards in the northeastern Italy [203]. The rates recommended to farmers were between 7.2 to 43.2 kg ha–1 of GLY [203]. A decrease in the survival of the earthworms was observed. However, differences were detected in the sensitivity of the tested earthworm species. The significant decline of the survival of the most sensitive Octodrilus complanatus was detected even at the lowest concentration (0.59 g m–2 of GLY) tested. Furthermore, a drastic decrease in the number of cocoons was also observed at the lowest test concentration. In the tested Lumbricus terrestris collected from a conventionally managed GLY-treated vineyard for at least three decades, the number of the cocoons was not affected by the treatments suggesting buildup of some resistance mechanisms to GBH exposure among anecic earthworms [203].
An improved performance in growth, fruit yield and quality were detected in tomatoes planted with earthworm casts produced by different species (Alma millsoni, Eudrilus eugeniae, and Libyodrilus violaceus), while increased vitamin C and β-carotene contents were measured in the fruits of tomatoes planted with the casts of earthworms not exposed to GBH Roundup Alphée (8.3 kg ha–1) compared to the normal soil control units [204]. Moreover, the tomatoes planted with the use of casts produced by the exposed earthworms were not able to set fruit [204]. Consequently, casts of unexposed earthworms promoted tomato performance more than NPK fertilizer.
In a model mesocosm greenhouse experiment, GBH (Roundup Speed) had adverse effects on belowground interactions between earthworms (L. terrestris) and symbiotic AMF. Moreover, significant reductions in root mycorrhization, soil AMF spore biomass, and earthworm activity were also observed [205]. In addition, soil hyphal biomass was increased and soil water infiltration was reduced [205]. GBHs (Roundup LB Plus, Roundup PowerFlex, Touchdown Quattro) and their corresponding AIs (GLY-IPA, -potassium, and -diammonium salts) decreased earthworm (L. terrestris) casting and movement activity at the recommended application rates [52]. In contrast, no significant differences were found in the effects of the GBHs and their associated GLY AIs on earthworm activity [52]. According to the results obtained, toxicity had substance-specific characteristics, and the amount of leachate in the experimental pots after simulating heavy rainfall was higher in GBH treatments than in AI treatments [52]. Oxidative stress, induction of acetylcholinesterase (AChE) activity, and increased concentration of malondialdehyde were observed after short-term exposure to a GBH (0.3, 3, and 30 µg kg dw soil–1 GLY) in earthworms (Dendrobaena veneta), but reproduction was not affected [206]. In L. terrestris, vertical burrowing activity almost ceased 3 weeks after GBH application, and reproduction was also reduced by 56% within 3 months after herbicide treatment. In contrast, no significant differences were observed in the activity of the soil-dwelling earthworm Aporrectodea caliginosa [207]. A concentration-dependent growth inhibition (reduced size and length) was detected in Aporrectodea longa exposed to the GBH Grassate [188]. Furthermore, in a pilot study, disruptions were observed in the gut microbiome of earthworms (Alma millsoni, Eudrilus eugeniae, and Libyodrilus violaceus) exposed to GBH Roundup Alphée (8.3 kg ha–1) [208]. Moreover, a significant shift in bacterial community composition was observed, especially by the increased abundance of Enterobacter, Pantoea, and Pseudomonas genera compared to the control [208]. GBHs (Roundup Original, Trop, Zapp, Crucial) at the recommended application rate affected earthworms, collembolans, and isopods, with non-avoidance behavior observed in earthworms and impaired feeding activity in collembolans [68]. To the best of our knowledge, no study has assessed the effects of GLY/GBH on enchytraeids [209].
Earthworms are commonly distinguished in three ecological groups according to their feeding habits: endogeics or horizontal burrowers, anecics or vertical burrowers, and epigeics or litter dwellers. Most studies on GLY/GBH effects have been conducted on epigeics. This is due to the fact that they are also an approved surrogate species in official regulatory environmental risk assessments such as those required in the EU. Findings that earthworm species vary in their sensitivity to GLY/GBH [207, 210, 211] call for more diversity in the earthworm species used to study the effects of this and other herbicides. Overall, the studies carried out to date make it clear that worms show differing sensitivities to GLY/GBHs with respect to distinctive response parameters (survival, growth, activity, reproduction). As with all other non-target species, shifts in soil faunal communities are not addressed, although changes in community structure are known to be the most significant effect of some pesticides [212].
Terrestrial vertebrates
Terrestrial vertebrates are important herbivores and predators in agroecosystems. Amphibians and reptiles are among the most threatened vertebrate species worldwide. Problems faced by amphibians is particularly acute as they have a life cycle that encompasses aquatic and terrestrial phases as well as migration to and from spawning waters and can thus be exposed to pesticides in the water and on land [213, 214]. To estimate the risk of GLY/GBHs for terrestrial vertebrate species, only birds and rodent mammals are obligatory in ERA. Amphibians, reptiles, and bats are not, at least in the EU [215]. As the focus of this review is on terrestrial ecosystems, we will only consider studies focusing on GLY/GBH effects on the terrestrial life stages of amphibians.
Ground dwelling vertebrates
Effects of GLY and its GBHs on rodents in the wild, as well as reptiles continue to be neglected. However, the extensive use of GBHs raises great concern about its impacts on wildlife. In a study of dermal exposure to two GBHs (0.144 g AI L–1 of Agpro Glyphosate 360 and Yates Roundup Weedkiller) with spraying aimed to simulate field conditions, no significant effects on the mass of exposed Oligosoma polychrome skinks were observed [216]. However, significantly higher skin temperatures were observed after treatment with Yates Roundup Weedkiller, suggesting increased metabolism and physiological stress responses [216]. This kind of heat-seeking behavior could reflect an effort to trigger a fever response to environmental stress. The increased body temperature may allow reptiles to better deal with harmful effects caused by environmental stressors through more effectively functioning physiological and biochemical systems (e.g., higher biotransformation capacity) [217,218,219]. However, the heat-seeking behavior can also result in also side-effects such as increased water loss and predation risk. In addition, higher biotransformation capacity can cause not only a reduce level of the contaminant, but also can generate more toxic metabolites. The temperature-dependent toxicity of insecticides (e.g., chlorpyrifos) resulting in altered thermoregulation in exposed lizards has also been demonstrated [219]. In newborn lizards (Salvator merianae), a significant increase in DNA damage was observed as a result of embryonic exposure to GBH Roundup Full II (66.2% GLY) at 50–1600 μg GBH egg–1 (corresponding to 33.1–1059.2 μg AI egg–1), which translates into an application rate of 1.3–2.0 kg ha–1, although levels of micronucleus, nuclear abnormalities and size of the newborns at birth and 6 months after exposure were not significantly altered [220]. After low dose GLY exposure at low doses (0.05 and 0.5 μg kg bw–1), toxic effects on the liver of Podarcis siculus lizard were observed regardless of the sex [221]. In addition, the appearance of fibrotic lesions, reduced physiological liver function and increased enzyme activities (e.g., superoxide dismutase and glutathione peroxidase) and the upregulation of estrogen receptor α and vitellogenin gene expression also noted [221]. In male lizards, decreased spermatogenesis, altered testicular morphology, and modified localization of estrogen receptors in germ cells were observed following 3weeks of GLY exposure (at 0.05 and 0.5 μg/kg body weight), but steroidogenesis was not affected [222]. Toxic effects from a GBH has not been demonstrated in the Sceloporus occidentalis lizard [223].
Several studies have demonstrated adverse effects of GBHs on amphibians even at the environmentally relevant concentrations. The effects on amphibians were found to be strongly correlated with the composition of the GBH, as well as the sensitivity and life stages of the tested species. Toxic effects of GLY were often associated with the presence of the co-formulants. Generally, most studies undertake ecotoxicological testing aims to investigate the effects of GLY and GBHs on the early developmental stages (e.g., tadpoles) as this is a window of greater sensitivity.
The toxicity of a GBH and arsenite as individual toxicants and in a 50:50 mixture was determined in Rhinella arenarum tadpoles during acute (48 h) and chronic assays (22 days). [224]. In both types of assays, the levels of enzymatic activity (AChE, carboxylesterase, and glutathione S-transferase (GST)), and the levels of thyroid hormones were examined. Additionally, the mitotic index of red blood cells and DNA damage index were calculated for the chronic exposure phase of the investigation. LC50 values at 48 h were 46.0 mg L–1 GLY equivalent concentration for GBH, 37.3 mg L–1 for arsenite, and 30.3 mg L–1 for the GBH-arsenite mixture. The study demonstrated strong synergistic toxicity of the GBH-arsenite mixture, negatively altering antioxidant systems and thyroid hormone levels, with consequences on red blood cell production and DNA damage in treated tadpoles. In addition, skin damages, developmental effects including morphological changes and genotoxicity were also observed in the tadpoles [225,226,227,228]. Morphological alterations in liver and other hepatotoxic effects (e.g., increased melanin area) were observed in Leptodactylus latinasus exposed to GLY (100, 1000, 10,000 µg g–1 equivalent concentration) [229]. In the common toad Rhinella arenarum exposed to GLY (20 mg L–1), increased oxidative stress, immunological depression and neurotoxic effects were detected [230]. In a study on North American anuran species investigating GBH co-formulant toxicity, the determined 96-h LC50 values were in the range 0.68–1.32 mg L–1 following exposure to surfactant MON 0818 (POEA), which demonstrates the highly toxic nature of this substance [231].
Laboratory animal studies are mainly conducted with rats and mice and occasionally with rabbits. Rats are the preferred model system as these animals are a regulatory accepted surrogate for human health risk assessment. However, such investigations could also provide information on the effects of wild rodents. In one study, Sprague–Dawley rats were exposed via drinking water to either GLY or GBH Roundup Bioflow at the US GLY acceptable daily intake (ADI) of 1.75 mg kg–1 body weight (bw) day–1 [232]. One cohort was continuously dosed until sexual maturity (6-week cohort) and another cohort was continuously dosed until adulthood (13-week cohort). Survival, body weight, food and water consumption of the rats were not affected by the treatment with either GLY or the Roundup Bioflow [232]. Also, the concentrations of both GLY and AMPA in urine treated with GLY were similar to that observed in animals treated with GBH. The majority of GLY was excreted unchanged. Up to 100-fold higher urinary GLY levels (0.48–2.28 mg kg–1) were detected in the treated animals compared to the detected AMPA levels (0.011–0.027 mg kg–1). Co-formulants present in the GBH did not seem to exert a major effect on the absorption and excretion of GLY [232]. Another investigation showed that the gut microbiota of mice was also disturbed, with altered relative abundance and phylogenic diversity of key microbes were observed following exposure of 250 or 500 mg kg–1 bw day–1 GLY equivalent of a GBH (Roundup) [233]. In addition, subchronic and chronic exposure increased anxiety- and depression-like behaviors. This study reinforces the essential link between gut microbiota and GBH toxicity in mice and suggest that intestinal dysbiosis can increase the prevalence of neurobehavioral alterations [233]. Fungal and bacterial diversity of the caecum microbiome of rats exposed to GLY (0.5, 5, 50 mg kg body weight–1 day–1), or to its GBH formulations (Roundup Bioflow and RangerPro) at the same GLY equivalent dose starting at a prenatal stage of development up until to adulthood was markedly altered by the GBHs in a dose-dependent manner with exposure to GLY alone significantly altered only bacterial diversity [234]. An assessment of the gut metabolome of adult rats exposed to GLY and Roundup MON52276 (0.5, 50, 175 mg kg−1 bw day−1) showed an oxidative stress response and inhibition of the shikimate pathway in the gut microbiota, a metabolic effect that was previously thought to be restricted to plants as part of this compounds mechanism of acting as weed killer [28]. This observation of shikimate pathway inhibition in gut microbiota can have major implications for the integrity of microbiome composition and function in the environment (soil, water courses, animals). In addition, analysis of the liver from the same animals showed marked oxidative stress and genotoxicity in GLY and especially MON52276 treatments suggesting a carcinogenic capability [25]. Exposure to GLY and a GBH Roundup formulation resulted in an increased in homocysteine levels in treated male rats at the US ADI valued for GLY (1.75 mg kg bw day–1) [235]. In addition, significant toxicity of GLY on rabbit biochemical (e.g., alanine aminotransferase, urea, and creatinine) and hematological (e.g., white blood cells, hemoglobin, hematocrit) parameters was found in a concentration- and time-dependent manner [236]. GLY and its formulation (Roundup 3 Plus) (EU ADI 0.5 mg kg−1 bw day−1 plus 5 and 50 mg kg–1 bw day–1 groups) negatively affected the male reproductive system in mice following perinatal exposure to GLY via drinking water (tested concentrations: the current EFSA ADI 0.5 mg kg–1 day–1, in addition to 5 and 50 mg kg–1 day–1) resulting in altered testis morphology and decreased serum testosterone concentrations [237]. Reduced sperm count was detected at 0.5 and 5 mg kg–1 day–1 of GLY and Roundup 3 Plus with decreased numbers of spermatozoa also observed [237]. The female reproductive system has also been observed to be severely impacted by GLY exposure with as evidenced by increased ovarian weight and follicle number, as well as an altered ovarian proteome following chronic GLY exposure at low-levels [238]. Furthermore, GLY has been shown to induce the transgenerational inheritance of disease (e.g., obesity, kidney, and ovarian diseases) and germline (e.g., sperm) epimutations [239, 240]. In Chaetophractus villosus armadillo exposure to GBH Roundup Full resulted in a significant increase in the chromosome aberration (e.g., chromatid breaks), altered cellular and genetic biomarkers (e.g., cell proliferation kinetics and mitotic index), as well as increased frequency of sister chromatid exchanges, confirming the possible genotoxicity of the GBHs [241, 242].
As exposure of vertebrates living in agroecosystems to GLY/GBHs seems inevitable, a high risk to these populations can be anticipated. However, most studies focus on single-species scenarios and do not investigate effects on community composition. Moreover, studies on long-term effects of GLY/GBH in reproduction of vertebrates are missing. Reptiles that lay eggs in soil, and amphibians living in soil caves contaminated with GLY/GBH residues can have long-term contact with these herbicides, but to the best of our knowledge, this has not been studied thus far.
Birds
Birds are among the best-studied wild animals regarding GLY and GBH effects. This is mainly because birds are an integral part of ERA during the regulatory GLY registration process. Among the estimated 10,000 bird species present worldwide, avian toxicity data for risk assessments are produced in feeding studies using only a few model bird species: e.g., Japanese quail (Coturnix japonica), bobwhite quail (Colinus virginianus), or Mallard duck (Anas platyrhynchos).
Dietary exposure to GBH RoundUp Flex (200 mg kg–1) resulted in 0.76 mg kg–1 GLY residues in Japanese quail (Coturnix japonica) eggs and caused a decreased embryonic development, and in addition a 20% higher lipid damage was detected in the embryonic brain tissue of exposed parental quails compared to controls was also observed [243, 244]. In another study involving Japanese quails, a preference for the GLY-containing feed (12–20 mg GLY kg−1 bw day–1) was observed compared to that without GLY [245]. In addition, delayed plumage development was observed in the exposed females with the appearance of GLY in the eggs and liver [245]. Furthermore, after dietary exposure to a GBH (160 mg kg–1 RoundUp Flex), decreased activity of antioxidant biomarkers in the liver and lower testosterone levels in male quails was observed in exposed C. japonica [245]. In contrast, increased plasma testosterone levels and growth performance, as well as the fattening of offspring and reduced sperm motility were demonstrated in roosters after chronic dietary exposure to GBH Roundup (46.8 mg kg–1 bw day–1 GLY) [246]. GBHs have also been reported to negatively impact the gut microbiome in quails especially at early stages of development [245] and in poultry where a reduction in the beneficial bacterial community in the gastrointestinal tract was observed after exposure to GBH Roundup UltraMax at a sublethal concentrations (0.075–5 mg mL–1) with the observed adverse effects possibly leading to disruption of the normal functions in the gut of exposed birds [247].
Despite evidence of GLY/GBH effects on birds through ERA procedures, many questions remain as to how well these tests can describe the toxicological situation for the majority of bird species. Both standard test species of quail and the Mallard duck, belong to species whose nestlings are very mobile, feed themselves very early in life, and do not rely on their parents feeding them, while many other bird species depend on parental care (like all passerine species). Moreover, in these standard tests only the oral uptake of contaminated food is addressed. However, dermal uptake by the feet and indirect effects via changes in the availability and quality of insect and plant food following GLY/GBH treatment are also important and need to be considered [73, 74].
Combined effects between GLY and other environmental pollutants
Most environmental pollutants (e.g., pesticide AIs, drug residues, formulating agents, and other additives) are present in the environment as a mixture of different contaminants where they may come into contact with each other in various matrices. Thus, GLY and its metabolites, or the other components of GBHs presented in the environment in all likelihood will interact with other pollutants. The determination and evaluation of the combined effects of the various pollutants and chemical compounds is necessary to conduct an appropriate and more accurate ERA.
Interactions with other AIs, co-formulants, and pollutants
Serious concerns have been raised about the ecotoxicological consequences of pesticide residues related to the combined effects of compounds of agricultural (or other) origin. It is well established that the toxicity of certain pesticide residues depends not only on the applied concentration and environmental conditions, but also on the co-occurrence of various other compounds. The effects of seed dressings on winter wheat (Triticum aestivum L. var. Capo) with neonicotinoid insecticides and/or fungicides (strobilurin and triazolinthione) on earthworm activity were enhanced by the presence of GLY due to the one-time application of a GBH, although soil basal respiration, litter decomposition, and microbial biomass were not affected by either seed dressing or GBH application [248]. The combined application of a GBH and the insecticides lambda-cyhalothrin or fipronil reduced the efficiency of weed control on certain plants compared to GBH alone, while the simultaneous application of GBH and the organophosphate insecticide dicrotophos enhanced cotton aphid control four days after treatment compared to insecticide with control of thrips also improved in the presence of GBH and imidacloprid together [249]. Additive effects of the GBH (Roundup) and atrazine-based (Atrazine 50 SC) formulations were found on lethality, fertility, and locomotion in C. elegans, which were associated with several toxic responses related to oxidative stress [131].
Chronic toxicity of pesticide AIs was evaluated on A. mellifera in studies targeted on assessing the effects on mortality of a fungicide AI (difeconazole) applied individually or in combination with a herbicide AI (GLY) and or an insecticide AI (imidacloprid) [250]. Fungicide toxicity after a single treatment via spraying at the registered application rate (75 g ha–1) on the seventh day of the treatment setup was increased after previous chronic oral exposition to GLY (0.01–0.1 μg L–1 in food for 30 days). Astonishingly, fungicide toxicity was reduced by a similar previous exposure to the neonicotinoid type insecticide (the same chronic exposure condition as for GLY), and this mitigating affect could counteract the toxicity enhancement effect of GLY [250]. Similar effects were seen when all three AIs were applied orally, alone, or in binary or ternary mixtures at a broader dose range (0.01–10 μg L–1 in food for 20 days). However, individual toxicities of the three AIs appeared to be more similar to each other, the mitigating effect of imidacloprid decreased, and the pesticide combinations altering certain physiological markers of honey bees (e.g., increased GST activity in the head, midgut and abdomen) [251].
GLY and 2,4-D are two herbicides commonly mixed together in many products [77]. A lab experiment tested single and joint lethal toxicity of the GBH Atanor (43.8% GLY-IPA) and the 2,4-D-based herbicide Así Max 50 (602 g L–1 of 2,4-D) on amphibian Rhinella arenarum larvae. The GBH (504 h-LC50 = 0.039 g AI L–1) was significantly more toxic than the 2,4-D-based formulation (504 h-LC50 = 0.250 g AI L–1) and their toxicity was time-dependent. At LC50 and LC90 effect levels, the mixture interaction resulted synergistic outcomes of toxicity for all exposure times [77]. This is the first study to report the synergistic interactions between GBH and a 2,4-D-based formulation on amphibians, highlighting its negative impact on aquatic ecosystems.
Synergistic effects were also detected of a tank-mixed adjuvant (KAO A-134) on the post-emergence activity of GBH Roundup against crabgrass (Digitaria sanguinalis) [252]. As expected, the field study showed that the adjuvant can increase the herbicidal effect of the GBH. The main purpose of co-formulants is to enhance the effectiveness and bioavailability of the formulation by increasing the adsorption, solubility, distribution, adhesion, and penetration properties, as well as the environmental stability of the AIs [17, 253]. The exact composition of pesticide formulations with respect to co-formulants is generally withheld by manufacturers invoking claiming commercial confidentiality and thus not listed on product packaging labels. However, although listed as “inert”, an increasing number of demonstrate that co-formulants can significantly affect the toxicity, including cytotoxicity, genotoxicity, and endocrine-disrupting effects of GBHs on non-target organisms [24, 95, 254,255,256].
Other substances such as heavy metals, which can be present naturally or as human generated pollutants, are present in the environment and may also interact with GLY residues. GLY was originally marketed as a potent metal chelating agent for heavy metals [257], and can therefore it can influence the environmental fate of toxic metals in the soil–water system and pose ecosystem toxicity issues [258]. The functional groups of GLY such as carboxylate, amine, and carboxylate can bind cations and form metal complexes resulting in reduced bioavailability of heavy metals. Exposure of the earthworm E. fetida to GLY, Cu or a combination of the two, found that GLY significantly decreased the acute toxicity of Cu, while the mortality rate and Cu uptake were decreased after simultaneous exposure to GLY and Cu [259]. In a follow-up study, a relative weight loss and decreased catalase, malondialdehyde, and superoxide dismutase activities were also observed from GLY and Cu exposure [260]. Additive negative effects of heavy metals (Cu, Cd) and a GBH (Glialka Star) on the chicken embryos has been demonstrated with decreased body weight and increased embryonic mortality, as well as developmental abnormalities were detected after injection of the GBH-heavy metal mixture compared to the individual toxicity effects from the individual tested compounds [261].
The chelation properties of GLY can also result in reducing bioavailability of metal micronutrients resulting in deficiencies in plants, and thus altered processes and pathogen resistance of plants can be caused. In addition, the various plant–microorganism interactions (e.g., nitrogen fixation) can be negatively impacted by GBHs. In addition, GLY may indirectly contribute to higher toxicity by concentrating toxic heavy metal residues. The exact role and contribution of the chelating property of GLY in the development of adverse effects in the environment and non-target organisms are far from being fully known [258]. However, the metal chelating properties of GLY are well known, and this additional potential environmental effect and risk are never considered adequately during the regulatory risk assessment of this herbicide AI [258, 262, 263]. It has also been found that GLY residues or drift may reduce the uptake and translocation of micronutrients (e.g., Mn or Fe), in non-target plants and suggested GLY–metal complex formation in plant tissues and/or plant rhizospheres that hinder their root uptake and translocation by the crops [264]. Several other studies link the ability of GLY to inhibit the acquisition of micronutrients, such as Mn, Fe, Zn and B, in plants exposed to GLY [265]. Such interactions of GLY with plant nutrition may potentially pose consequences on crop performance and pose a health risk to consumers [46]. The toxic heavy metal chelation properties of GLY has been suggested as a possible mechanism for the unexplained high incidence of chronic kidney disease amongst agricultural workers [266].
Plastic debris in our environment can be degraded into micro- and nanoplastics by natural processes. In a 30-day soil incubation experiment, the combination of GLY and a low content of microplastics (homopolymer polypropylene powder) negatively affected the dynamics of dissolved organic matter and carbon in soil [267]. These adverse effects resulted in a loss of bioavailable carbon and phosphorus content in soil, while the interaction of GLY and high microplastics content reduced dissolved organic nitrogen in soil [267]. The co-occurrence of GLY and micrometer-sized plastic debris as microplastics resulted in altered soil microbial respiration with altered dynamics of soil urease, β-glucosidase, and phosphatase although no effect on GLY degradation [268]. In a 14-day mesocosm experiment, simultaneous exposure to GLY and light-density polyethylene microplastics negatively affected the weight of the exposed earthworms [269]. Additionally, GLY and AMPA were concentrated in the first centimeter of top soil layer and the downward transport of the tested compounds was only detected in the earthworm burrows (0.04–4.25 μg g–1). The rate of GLY transport (including AMPA) by earthworms was highly depended on the synergetic effects of combined application of GLY and microplastics [269].
Interactions with pathogens and parasites
GLY disrupts the shikimate pathway in different plants and important groups of microorganisms with the prevention of the production of essential aromatic amino acids. GLY treatment results in prolonged lethal effects on plants and reduced resistance to root pathogens [270]. GLY can alter the susceptibility of the moth G. mellonella and the mosquito A. gambiae to infections. A reduced survival was observed in GLY-exposed moths infected with the fungus Cryptococcus neoformans, possibly due to the weakened melanin-based immune response of G. mellonella. Additionally, increased susceptibility of A. gambiae was detected to infection with the Plasmodium falciparum, the major vector of human malaria in Africa [150].
GLY is known to affect the functioning of various bacteria including those of the gut microbiome. The disruption of gut bacterial composition and function also makes bees more vulnerable, leading to various health effects and decreased resistance to pathogens, which may even result in the weakening or complete loss of bee colonies. Application of GLY (5–10 mg L–1) resulted in higher mortality of worker bees when exposed to the pathogen Serratia marcescens. Furthermore, GLY exposure (0.1, 1, or 10 mM for 5 days) decreased the expression of antimicrobial peptides (e.g., apidaecin, defensin, and hymenoptaecin in the exposed bees. In addition, GLY can reduce the abundance of beneficial gut bacteria leading to immune dysregulation [164, 165]. Co-exposure to GBH Roundup and parasitic Nosema microsporidia resulted in a significantly reduced survival rate and increased food consumption of honey bees [271]. However, based on modified acute oral toxicity tests in bumble bees, the interaction between GLY treatments (200 µg per worker) and the intensity of the parasite Crithidia bombi in the bumble bees tested did not result in adverse effects in the treated groups [272].
Recently, many concerns have been raised regarding altered susceptibility of bacteria species to a diverse range of antibiotics (e.g., ampicillin, ciprofloxacin, tetracycline). There is evidence to suggest that GLY may serve as one of the drivers for bacterial antibiotic resistance [273,274,275]. The co-occurrence of GLY and various co-formulants presented in GBHs at or below the recommended application concentrations altered the susceptibility of the potentially pathogenic bacteria Escherichia coli and Salmonella enterica to multiple antibiotics [273]. These results suggest the promotion of antibiotic resistance by GLY and other herbicide AIs [273, 276].
Interactions between GLY/GBH and other environmental pollutants are the most understudied and complex effects. ERA is based predominantly on the results of laboratory studies where test organisms are exposed to a range of concentrations of single compounds under standard conditions. However, in their natural settings, organisms rarely experience optimal conditions. Additionally, for most of their lifespan, they are forced to cope with suboptimal conditions and frequent exposure to severe environmental stress. The available studies that address these aspects show that these environmental pollutants may or may not alter the effects of GLY or GBHs.
Conclusions and recommendations
There is a widespread assumption that GLY/GBHs are specific and only affect target weed plants. In this review, we have compiled extensive evidence published since 2010 showing that GLY and GBHs have multiple effects on various non-target organisms in terrestrial ecosystems. Regardless of the fact that GLY may be less acutely toxic than many other herbicidal AIs, unintended side-effects on a range of organisms, including microorganisms, invertebrates, and vertebrates, are evident especially upon chronic exposure. The major mechanisms of these effects are oxidative stress, metabolic dysregulation, metabolome disturbances, endocrine disruptions and genotoxicity including direct DNA damage. After exposure, the consequences of these effects manifest themselves in various physiological and behavioral outcomes. The extremely high global use of GLY/GBH, exceeding 800 thousand tonnes annually since 2014 [43], increases the likelihood of exposure and increases risks to the environment and health. Although our review includes studies conducted worldwide, we evaluated them within the framework of the EU regulations. This is because the extension for regulatory approval for GLY is currently under consideration in 2023. Regulations in different countries vary widely [277, 278], and a discussion of their differences is beyond the scope of this review. Impacts result from ecosystem disruptions that are not a part of any regulatory review anywhere in the world.
Simultaneous exposure to GLY, GBHs, and the co-formulants contained in the GBHs can cause a wide range of ecotoxicological effects, including lethal/sublethal effects, and morphological and biochemical changes in exposed organisms [52, 98, 115]. In general, there is an urgent and great need for evaluations of the effects of GLY/GBH at low, realistic levels of exposure and long-term testing involving multiple generations of organisms. Most ecotoxicological studies have been conducted on only a few surrogate non-target species, ignoring the different life history characteristics and sensitivities of most species exposed to GLY/GBHs. Therefore, in order to be able to estimate the contribution of GLY/GBHs to biodiversity decline, the inclusion of more sensitive species in the risk assessment being conducted by the EFSA should be considered. In addition, risk assessment and ecotoxicological testing should be carried out in all cases for the pesticide formulations and not only for the corresponding AIs, since exposure is associated with the formulated products under real environmental conditions. In addition, mandatory full disclosure and strict regulation of co-formulants is needed as they affect the toxicity of commercial pesticide formulations and the fate of the AIs in the environment. However, shockingly Annex III of Regulation (EC) 1107/2009, which is supposed to contain the list of co-formulants not allowed in pesticide formulations, is still empty [32]. Given the broad range of adverse effects on various non-target organisms demonstrated, changes in official pesticide regulations, including strict regulation of co-formulants, and development of a unified approach to determine the environmental risks of mixtures of different chemical compounds is essential. In addition, the development and implementation of new effective guidelines for the detection of combined effects of pollutants are also needed [43, 44]. The identified hazards are of great importance for appropriate risk assessment of commercial pesticide formulations and point to the need for re-evaluation of current pesticide testing schemes and methods [42, 44].
As already highlighted, the result of ecosystem disturbances is not subject to regulatory review. Current regulation of pesticides is based on limited studies that evaluate the effects of a typical rate of application on a given organism in a specified experimental setting. The results of these studies meet the requirements of the current labels and uses, but do not take into account what happens to ecosystems and the organisms living within them when a GBH is sprayed two to three times a year, every year, for many years and now for decades [6]. Furthermore, the current regulation assumes that if one hectare can be treated without observable adverse effects, all hectares can be treated multiple times or for many years without problems.
This review includes studies that address terrestrial ecotoxicity of GLY, GBHs and co-formulants. The reviewed studies used a wide variety of metrics, endpoints, thresholds, and methods to estimate exposures and effects [8]. The use of higher doses than would be encountered in the environment was at times justified by simulating direct overspraying of species, or wrong dosage because of misleading label instructions [207]. It has been shown that even after several decades of GBH use, we still do not have a clear picture of their effects on terrestrial organisms and ecosystems. One of the main problems is that the full list of ingredients in a GBH is not made public and considered confidential business information. Given the drastic decline in biodiversity [279], and especially in Europe, where ERA is conducted according to the precautionary principle [280], business interests should not be more important than public and environmental health.
Our literature review has shown that there is still a great need for studies investigating the effects of co-formulants in GBHs. In the registration of GBHs, these co-formulants are still considered to be inert and chemically inactive. However, many studies have shown the high individual toxicity of the co-formulants and the increased combined toxicity of AIs and co-formulants compared to the individual toxicity of AIs [21,22,23]. Therefore, evidently various co-formulants are clearly chemically active and indeed in the vast majority of cases up to 1000-fold more toxic than the corresponding AI alone. In general, the results on the toxicity of GLY and GBH are difficult to extrapolate and compare because the species tested have different sensitivities, the study settings are different, and the compositions of the thousands of GBH is also vary. In addition, in most cases, the co-formulants in GBHs are not disclosed, and GBHs differ in composition from country to country, even if they have the same trade name.
A critical point is that many of the ecological impacts discussed occur because of the frequency and extent of GBH use. Current regulation is based on ERA that evaluates the effects of a typical or maximum rate of application on a surrogate organism in a given environment. The existing ERA is conducted for an AI or pesticide formulation applied once or a few times in a given crop. However, in current cropping systems, multiple pesticide applications are made throughout the season. The current ERA scheme addresses the effects of a pesticide on each group of organisms separately but crucially does not include interactions between trophic levels of different organism groups [73, 154]. The EU ERA focuses on environmental effects that may occur in semi-natural structures outside of agricultural fields. Currently, however, ERA does not mandate in-field risks, even though the prevailing scientific opinion states that biodiversity must be supported to some degree in order to provide important ecosystem services [154]. Therefore, many studies do not examine what happens to organisms and agroecosystems when a GBH is sprayed 2–3 times per year, every year, for many years, and now decades [6].
There are many knowledge gaps of the effects of GLY/GBH on terrestrial non-target organisms and ecosystems that we have identified in this literature review:
-
Multispecies and trophic interactions within agricultural fields and landscapes.
-
Detailed studies on the contribution of all ingredients of a GBH, including AIs, co-formulants, or other contaminants such as arsenic or lead.
-
Effects on soil microbiota composition and function stemming from the inhibition of the shikimate pathway and its impact on plant health.
-
Since GBHs have been in use for decades, there is a great need for long-term effects of high and low chronic exposure in species with short and long generation times.
-
Sequences and interactions of GBHs with other pesticides applied during the cropping year.
-
Carry-over effects of GBH applications to the following year and influences on GLY-tolerant crops and resistant weeds, or effects on the disease susceptibility of crop plants.
-
Interactions with other environmental pollutants such as agrochemicals, antibiotics, other chemicals, microplastics, parasites and global change factors.
In the absence of information on the above issues, which are vital for accurate ERA assessment of GLY/GBHs, we recommend invoking the precautionary principle enshrined in EU law and removal of this class of herbicides from the market. It is particularly important to do so since GLY/GBHs are applied on a massive scale and that impacts of toxicity can take many years to manifest.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- AChE:
-
Acetylcholinesterase
- ADI:
-
Acceptable daily intake
- AI:
-
Active ingredient
- AMF:
-
Arbuscular mycorrhizal fungi
- AMPA:
-
Aminomethylphosphonic acid
- APG:
-
Alkyl polyglucoside
- CFU:
-
Colony-forming units
- DNA:
-
Desoxyribonucleic acid
- EC:
-
European Commission
- EC50 :
-
50% Effective concentration
- EFSA:
-
European Food Safety Authority
- EPSP:
-
5-Enol-pyruvyl-shikimic acid-3-phosphate
- ERA:
-
Environmental risk assessment
- EU:
-
European Union
- GBH:
-
Glyphosate-based herbicide
- GLY:
-
Glyphosate
- GLY-IPA:
-
Glyphosate-isopropylammonium salt
- GST:
-
Glutathione-S-transferase
- GT:
-
Glyphosate-tolerant
- GM:
-
Genetically modified
- LC50 :
-
50% Lethal concentration
- LD50 :
-
50% Lethal dose
- MRL:
-
Maximum residue level
- POEA:
-
Polyethoxylated tallow amines
- ROS:
-
Reactive oxygen species
- SOM:
-
Soil organic matter
References
Székács A, Darvas B (2012) Forty years with glyphosate. In: Hasaneen M (ed) Herbicides—properties, synthesis and control of weeds. InTech, Rijeka, pp 247–284. https://doi.org/10.5772/32491
Casida JE (2017) Organophosphorus xenobiotic toxicology. Ann Rev Pharmacol Toxicol 57:309–327. https://doi.org/10.1146/annurev-pharmtox-010716-104926
Dill GM et al (2010) Glyphosate: Discovery, Development, Applications, and Properties. In: Glyphosate Resistance in Crops and Weeds; pp 1–33. https://doi.org/10.1002/9780470634394.ch1
Mesnage R, Szekács A, Zaller JG (2021) Herbicides: brief history, agricultural use, and potential alternatives for weed control. In: Mesnage R, Zaller J (eds) Herbicides: chemistry, efficacy, toxicology, and environmental impacts. Emerging issues in analytical chemistry. Elsevier, Amsterdam, pp 1–20. https://doi.org/10.1016/B978-0-12-823674-1.00002-X
Swanson N, Leu A, Abrahamson J, Wallet B (2014) Genetically engineered crops, glyphosate and the deterioration of health in the United States of America. J Organ Syst 9:6–37
Benbrook CM (2016) Trends in glyphosate herbicide use in the United States and globally. Env Sci Eur 28:3. https://doi.org/10.1186/s12302-016-0070-0
Martins-Gomes C, Silva TL, Andreani T, Silva AM (2022) Glyphosate vs. glyphosate-based herbicides exposure: a review on their toxicity. J Xenobiot 12:21–40. https://doi.org/10.3390/jox12010003
Benbrook CM, Benbrook R (2021) A minimum data set for tracking changes in pesticide use. In: Mesnage R, Zaller J (eds) Herbicides: chemistry, efficacy, toxicology, and environmental impacts Emerging issues in analytical chemistry. Elsevier, Amsterdam, pp 21–40
ReportLinker (2021) Global glyphosate market to reach $8.9 billion by 2026. GlobeNewswire, Los Angeles, USA https://www.globenewswire.com/news-release/2021/10/18/2315706/0/en/Global-Glyphosate-Market-to-Reach-8-9-Billion-by-2026.html. Accessed 14 June 2023
PAN (2023) Weed management: alternatives to the glyphosate use. Pesticide Action Network Europe, Belgium https://www.pan-europe.info/resources/reports/2023/03/weed-management-alternatives-use-glyphosate. Accessed 14 June 2023
Antier C, Kudsk P, Reboud X, Ulber L, Baret PV, Messéan A (2020) Glyphosate use in the European agricultural sector and a framework for its further monitoring. Sustainabil 12:5682
Herrmann KM, Weaver LM (1999) The shikimate pathway. Ann Rev Plant Physiol Plant Mol Biol 50:473–503. https://doi.org/10.1146/annurev.arplant.50.1.473
Boocock MR, Coggins JR (1983) Kinetics of 5-enolpyruvylshikimate-3-phosphate synthase inhibition by glyphosate. FEBS Lett 154:127–133. https://doi.org/10.1016/0014-5793(83)80888-6
Tohge T, Watanabe M, Hoefgen R, Fernie A (2013) Shikimate and phenylalanine biosynthesis in the green lineage. Front Plant Sci. https://doi.org/10.3389/fpls.2013.00062
Defarge N, Takács E, Lozano VL, Mesnage R, Vendômois JS, Seralini GE, Székács A (2016) Co-formulants in glyphosate-based herbicides disrupt aromatase activity in human cells below toxic levels. Int J Env Res Pub Health 13:264. https://doi.org/10.3390/ijerph13030264
Travlos I, Cheimona N, Bilalis D (2017) Glyphosate efficacy of different salt formulations and adjuvant additives on various weeds. Agronomy 7:60. https://doi.org/10.3390/agronomy7030060
Foy C (1987) Adjuvants: terminology, classification, and mode of action. In: Chow P, Grant C, Hinshalwood A, Simundson E (eds) Adjuvants and agrochemicals. CRC Press, Boca Raton, pp 1–15. https://doi.org/10.1201/9781351069489
Defarge N, Spiroux de Vendômois J, Séralini GE (2018) Toxicity of formulants and heavy metals in glyphosate-based herbicides and other pesticides. Toxicol Rep 5:156–163. https://doi.org/10.1016/j.toxrep.2017.12.025
Fishel F (2020) What are inert ingredients? Pesticide Information Office, Gainesville, USA https://edis.ifas.ufl.edu/publication/PI081. Accessed 14 June 2023
US EPA (2022) Use of term “Inert” in the label ingredients statement. Pesticide Registration (PR) Notice 97–6. Office of Prevention, Pesticides and Toxic Substances, Office of Pesticide Programs, U.S. Government Printing Office, Washington DC, USA. https://www.epa.gov/pesticide-registration/prn-97-6-use-term-inert-label-ingredients-statement. Accessed 14 June 2023.
Mesnage R (2021) Coformulants in commercial herbicides. In: Mesnage R, Zaller J (eds) Herbicides: chemistry, efficacy, toxicology, and environmental impacts. Emerging issues in analytical chemistry. Elsevier, Amsterdam, pp 87–112
Tsui MTK, Chu LM (2003) Aquatic toxicity of glyphosate-based formulations: comparison between different organisms and the effects of environmental factors. Chemosphere 52:1189–1197. https://doi.org/10.1016/S0045-6535(03)00306-0
Székács A (2017) Mechanism-related teratogenic, hormone modulant and other toxicological effects of veterinary and agricultural surfactants. Insights Vet Sci 1:24–31. https://doi.org/10.29328/journal.ivs.1001003
Mesnage R, Benbrook C, Antoniou MN (2019) Insight into the confusion over surfactant co-formulants in glyphosate-based herbicides. Food Chem Toxicol 128:137–145. https://doi.org/10.1016/j.fct.2019.03.053
Mesnage R et al (2022) Comparative toxicogenomics of glyphosate and roundup herbicides by mammalian stem cell-based genotoxicity assays and molecular profiling in Sprague-Dawley Rats. Toxicol Sci 186:83–101. https://doi.org/10.1093/toxsci/kfab143
Schmidt R et al (2023) Glyphosate effects on earthworms: active ingredients vs. commercial herbicides at different temperature and soil organic matter levels. Agrochemicals 2:1–16
European Commission (2016) Commission Implementing Regulation (EU) 2016/1313 of 1 August 2016 amending Implementation Regulation (EU) No 540/2011 as regards the conditions of approval of the active substance glyphosate. Off J Eur Union 208:1–3. https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32016R1313. Accessed 14 June 2023
Mesnage R et al (2021) Use of Shotgun metagenomics and metabolomics to evaluate the impact of glyphosate or Roundup MON 52276 on the gut microbiota and serum metabolome of Sprague-Dawley rats. Env Health Perspec 129:017005. https://doi.org/10.1289/EHP6990
Gower SA, Loux MM, Cardina J, Harrison SK (2002) Effect of planting date, residual herbicide, and postemergence application timing on weed control and grain yield in glyphosate-tolerant corn (Zea mays). Weed Techn 16:488–494
Whigham DK, Stoller EW (1979) Soybean desiccation by paraquat, glyphosate, and ametryn to accelerate harvest. Agron J 71:630–633
Székács A, Mörtl M, Darvas B (2015) Monitoring pesticide residues in surface and ground water in Hungary: surveys in 1990–2015. J Chem 2015:717948. https://doi.org/10.1155/712015/717948
EP (2009) Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 concerning the placing of plant protection products on the market and repealing Council Directives 79/117/EEC and 91/414/EEC. Off J Eur Union 309:1–50. https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2009:309:0001:0050:en:PDF. Accessed 14 June 2023
Klátyik S, Bohus P, Darvas B, Székács A (2017) Authorization and toxicity of veterinary drugs and plant protection products: residues of the active ingredients in food and feed and toxicity problems related to adjuvants. Front Vet Sci 4:146. https://doi.org/10.3389/fvets.2017.00146
Anton A, Fekete G, Darvas B, Székács A (2014) Environmental risk of chemical agriculture. In: Gruiz K, Meggyes T, Fenyvesi E (eds) Engineering tools for environmental risk management. Environmental deterioration and contamination - problems and their management, 1st edn. CRC Press, Boca Raton, pp 93–112. https://doi.org/10.1201/b17102
Robin DC, Marchand PA (2021) The slow decrease of active substance candidates for substitution in the framework of the European Pesticide Regulation (EC) No 1107/2009. Eur J Risk Regul: https://doi.org/10.1017/err.2021.20
European Commission (2019) Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic And Social Committee and the Committee of the Regions. The European Green Deal. European Commission, Brussels, Belgium, Document 52019DC0640. COM(2019) 640 final https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:52019DC0640. Accessed 14 June 2023.
European Commission (2022) Trends in harmonised risk indicators for the European Union. European Commission, Brussels, Belgium https://food.ec.europa.eu/plants/pesticides/sustainable-use-pesticides/harmonised-risk-indicators/trends-eu_en. Accessed 14 June 2023.
Heap I, Duke S (2018) Overview of glyphosate-resistant weeds worldwide. Pest Manag Sci 74:1040–1049. https://doi.org/10.1002/ps.4760
Székács A (2021) Herbicide mode of action. In: Mesnage R, Zaller JG (eds) Herbicides: chemistry, efficacy, toxicology, and environmental impacts. Emerging issues in analytical chemistry. Elsevier, Amsterdam, pp 41–86
Gulickson G (2022) New Bayer crop herbicide with new mode of action coming by 2030. Successful Farming, https://www.agriculture.com/news/technology/new-bayer-row-crop-herbicide-with-new-mode-of-action-coming-by-2030-0. Accessed 14 June 2023
Defarge N, Otto M, Hilbeck A (2023) A Roundup herbicide causes high mortality and impairs development of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). Sci Tot Environ 865:161158. https://doi.org/10.1016/j.scitotenv.2022.161158
Panizzi S, Suciu NA, Trevisan M (2017) Combined ecotoxicological risk assessment in the frame of European authorization of pesticides. Sci Tot Environ 580:136–146. https://doi.org/10.1016/j.scitotenv.2016.10.154
Székács A, Darvas B (2018) Re-registration challenges of glyphosate in the European Union. Front Environ Sci. https://doi.org/10.3389/fenvs.2018.00078
Handford CE, Elliott CT, Campbell K (2015) A review of the global pesticide legislation and the scale of challenge in reaching the global harmonization of food safety standards. Integr Environ Assess Manag 11:525–536. https://doi.org/10.1002/ieam.1635
van den Berg H et al (2020) Pesticide lifecycle management in agriculture and public health: Where are the gaps? Sci Tot Environ 742:140598. https://doi.org/10.1016/j.scitotenv.2020.140598
Kanissery R, Gairhe B, Kadyampakeni D, Batuman O, Alferez F (2019) Glyphosate: its environmental persistence and impact on crop health and nutrition. Plants. https://doi.org/10.3390/plants8110499
Duke SO, Powles SB (2008) Glyphosate: a once-in-a-century herbicide. Pest Manag Sci 64:319–325
Borggaard OK, Gimsing AL (2008) Fate of glyphosate in soil and the possibility of leaching to ground and surface waters: a review. Pest Manag Sci 64:441–456
Helander M, Saloniemi I, Saikkonen K (2012) Glyphosate in northern ecosystems. Trends Plant Sci 17:569–574. https://doi.org/10.1016/j.tplants.2012.05.008
Hébert M-P, Fugère V, Gonzalez A (2019) The overlooked impact of rising glyphosate use on phosphorus loading in agricultural watersheds. Front Ecol Environ 17:48–56. https://doi.org/10.1002/fee.1985
Maderthaner M et al (2020) Commercial glyphosate-based herbicides effects on springtails (Collembola) differ from those of their respective active ingredients and vary with soil organic matter content. Env Sci Poll Res 27:17280–17289. https://doi.org/10.1007/s11356-020-08213-5
Zaller JG et al (2021) Effects of glyphosate-based herbicides and their active ingredients on earthworms, water infiltration and glyphosate leaching are influenced by soil properties. Env Sci Eur 33:51. https://doi.org/10.1186/s12302-021-00492-0
Tush D, Meyer MT (2016) Polyoxyethylene tallow amine, a glyphosate formulation adjuvant: soil adsorption characteristics, degradation profile, and occurrence on selected soils from agricultural fields in Iowa, Illinois, Indiana, Kansas, Mississippi, and Missouri. Env Sci Technol 50:5781–5789. https://doi.org/10.1021/acs.est.6b00965
Tush D, Maksimowicz MM, Meyer MT (2018) Dissipation of polyoxyethylene tallow amine (POEA) and glyphosate in an agricultural field and their co-occurrence on streambed sediments. Sci Tot Environ 636:212–219. https://doi.org/10.1016/j.scitotenv.2018.04.246
Kjær J, Olsen P, Ullum M, Grant R (2005) Leaching of glyphosate and amino-methylphosphonic acid from Danish agricultural field sites. J Environm Qual 34:608–620. https://doi.org/10.2134/jeq2005.0608
EP (2005) Regulation (EC) No 396/2005 of the European Parliament and of the Council of 23 February 2005 on maximum residue levels of pesticides in or on food and feed of plant and animal origin and amending Council Directive 91/414/EEC Text with EEA relevance. 16.3.2005. Off J Eur Union L70:1–16; https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32005R0396&qid=1686732142705. Accessed 14 June 2023
Silva V, Montanarella L, Jones A, Fernandez-Ugalde O, Mol HGJ, Ritsema CJ, Geissen V (2018) Distribution of glyphosate and aminomethylphosphonic acid (AMPA) in agricultural topsoils of the European Union. Sci Tot Environ 621:1352–1359. https://doi.org/10.1016/j.scitotenv.2017.10.093
Primost JE, Marino DJG, Aparicio VC, Costa JL, Carriquiriborde P (2017) Glyphosate and AMPA, “pseudo-persistent” pollutants under real-world agricultural management practices in the Mesopotamic Pampas agroecosystem, Argentina. Env Poll 229:771–779. https://doi.org/10.1016/j.envpol.2017.06.006
Kruse-Plaß M, Hofmann F, Wosniok W, Schlechtriemen U, Kohlschütter N (2021) Pesticides and pesticide-related products in ambient air in Germany. Env Sci Eur 33:114. https://doi.org/10.1186/s12302-021-00553-4
Zaller JG et al (2022) Pesticides in ambient air, influenced by surrounding land use and weather, pose a potential threat to biodiversity and human. Sci Tot Environ. 838:156012. https://doi.org/10.1016/j.scitotenv.2022.156012
Sabzevari S, Hofman J (2022) A worldwide review of currently used pesticides’ monitoring in agricultural soils. Sci Tot Environ 812:152344. https://doi.org/10.1016/j.scitotenv.2021.152344
Maqueda C, Undabeytia T, Villaverde J, Morillo E (2017) Behaviour of glyphosate in a reservoir and the surrounding agricultural soils. Sci Tot Environ 593–594:787–795
Grandcoin A, Piel S, Baurès E (2017) AminoMethylPhosphonic acid (AMPA) in natural waters: its sources, behavior and environmental fate. Weed Res 117:187–197. https://doi.org/10.1016/j.watres.2017.03.055
Knepper TP, Berna JL (2003) Chapter 1 surfactants: properties, production, and environmental aspects. In: Comprehensive analytical chemistry, vol 40. Elsevier, pp 1–49. doi:https://doi.org/10.1016/S0166-526X(03)40004-4
Bergé A, Wiest L, Baudot R, Giroud B, Vulliet E (2018) Occurrence of multi-class surfactants in urban wastewater: contribution of a healthcare facility to the pollution transported into the sewerage system. Env Sci Poll Res 25:9219–9229. https://doi.org/10.1007/s11356-017-0470-8
Mie A, Rudén C (2022) What you don’t know can still hurt you - underreporting in EU pesticide regulation. Env Health 21:79. https://doi.org/10.1186/s12940-022-00891-7
Simões T et al (2018) An integrative omics approach to unravel toxicity mechanisms of environmental chemicals: effects of a formulated herbicide. Sci Rep 8:11376. https://doi.org/10.1038/s41598-018-29662-6
Niemeyer JC, de Santo FB, Guerra N, Ricardo Filho AM, Pech TM (2018) Do recommended doses of glyphosate-based herbicides affect soil invertebrates? Field and laboratory screening tests to risk assessment. Chemosphere 198:154–160. https://doi.org/10.1016/j.chemosphere.2018.01.127
Nuutinen V, Hagner M, Jalli H, Jauhiainen L, Rämö S, Sarikka I, Uusi-Kämppä J (2020) Glyphosate spraying and earthworm Lumbricus terrestris L. activity: Evaluating short-term impact in a glasshouse experiment simulating cereal post-harvest. Europ J Soil Biol 96:103148. https://doi.org/10.1016/j.ejsobi.2019.103148
Pochron S, Simon L, Mirza A, Littleton A, Sahebzada F, Yudell M (2020) Glyphosate but not Roundup® harms earthworms (Eisenia fetida). Chemosphere 241:125017. https://doi.org/10.1016/j.chemosphere.2019.125017
Turner J (2021) The Pesticide Manual. 19th edition edn. The British Crop Protection Council, Brighton, UK
Vereecken H (2005) Mobility and leaching of glyphosate: a review. Pest Manag Sci 61:1139–1151. https://doi.org/10.1002/ps.1122
Brühl CA, Zaller JG (2021) Indirect herbicide effects on biodiversity, ecosystem functions, and interactions with global changes. In: Mesnage R, Zaller JG (eds) Herbicides: chemistry, efficacy, toxicology, and environmental impacts. Emerging issues in analytical chemistry. Elsevier, Amsterdam, pp 231–272
Zaller JG, Brühl CA (2021) Direct herbicide effects on terrestrial nontarget organisms belowground and aboveground. In: Mesnage R, Zaller JG (eds) Herbicides: chemistry, efficacy, toxicology, and environmental impacts. Emerging issues in analytical chemistry. Elsevier, Amsterdam, pp 181–230
European Parliament, European Council (2006) Regulation (EC) No 1907/2006 of the European Parliament and of the Council of 18 December 2006 concerning the Registration, Evaluation, Authorisation and Restriction of Chemicals (REACH), establishing a European Chemicals Agency, amending Directive 1999/45/EC and repealing Council Regulation (EEC) No 793/93 and Commission Regulation (EC) No 1488/94 as well as Council Directive 76/769/EEC and Commission Directives 91/155/EEC, 93/67/EEC, 93/105/EC and 2000/21/EC. Off J Eur Union L396:1–520. https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:02006R1907-20140410. Accessed 14 June 2023
Liess M, Henz S, Shahid N (2020) Modeling the synergistic effects of toxicant mixtures. Env Sci Eur 32:119. https://doi.org/10.1186/s12302-020-00394-7
Peluso J, Furió Lanuza A, Pérez Coll CS, Aronzon CM (2022) Synergistic effects of glyphosate- and 2,4-D-based pesticides mixtures on Rhinella arenarum larvae. Env Sci Poll Res 29:14443–14452. https://doi.org/10.1007/s11356-021-16784-0
Soil Association (2016) The impact of glyphosate on soil health: the evidence to date. Soil Association, Bristol, UK https://www.soilassociation.org/media/7202/glyphosate-and-soil-health-full-report.pdf. Accessed 14 June 2023
Richmond ME (2018) Glyphosate: a review of its global use, environmental impact, and potential health effects on humans and other species. J Env Stud Sci 8:416–434. https://doi.org/10.1007/s13412-018-0517-2
Wagner N, Reichenbecher W, Teichmann H, Tappeser B, Lötters S (2013) Questions concerning the potential impact of glyphosate-based herbicides on amphibians. Env Toxicol Chem 32:1688–1700. https://doi.org/10.1002/etc.2268
Jacoby R, Peukert M, Succurro A, Koprivova A, Kopriva S (2017) The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Front Plant Sci. https://doi.org/10.3389/fpls.2017.01617
Panettieri M, Lazaro L, López-Garrido R, Murillo JM, Madejón E (2013) Glyphosate effect on soil biochemical properties under conservation tillage. Soil Till Res 133:16–24. https://doi.org/10.1016/j.still.2013.05.007
Nourouzi MM, Chuah TG, Choong TSY, Lim CJ (2011) Glyphosate utilization as the source of carbon: isolation and Identification of new Bacteria. E-J Chem 8:614109. https://doi.org/10.1155/2011/614109
Singh S et al (2020) Herbicide glyphosate: toxicity and microbial degradation. Int J Environ Res Public Health. https://doi.org/10.3390/ijerph17207519
Haney RL, Senseman SA, Hons FM (2002) Effect of Roundup ultra on microbial activity and biomass from selected soils. J Environ Qual 31:730–735. https://doi.org/10.2134/jeq2002.7300
Mandl K, Cantelmo C, Gruber E, Faber F, Friedrich B, Zaller JG (2018) Effects of glyphosate-, glufosinate- and flazasulfuron-based herbicides on soil microorganisms in a Vineyard. Bull Environ Contam Toxicol 101:562–569. https://doi.org/10.1007/s00128-018-2438-x
Zaller JG et al (2018) Herbicides in vineyards reduce grapevine root mycorrhization and alter soil microorganisms and the nutrient composition in grapevine roots, leaves, xylem sap and grape juice. Env Sci Poll Res 25:23215–23226. https://doi.org/10.1007/s11356-018-2422-3
Newman MM, Hoilett N, Lorenz N, Dick RP, Liles MR, Ramsier C, Kloepper JW (2016) Glyphosate effects on soil rhizosphere-associated bacterial communities. Sci Tot Environ 543:155–160. https://doi.org/10.1016/j.scitotenv.2015.11.008
Allegrini M, Zabaloy M, del Gómez VE (2015) Ecotoxicological assessment of soil microbial community tolerance to glyphosate. Sci Tot Environ 533:60–68. https://doi.org/10.1016/j.scitotenv.2015.1006.1096
Kepler R et al (2020) Soil microbial communities in diverse agroecosystems exposed to the herbicide glyphosate. Appl Environ Microbiol 86:e01744-e11719. https://doi.org/10.1128/AEM.01744-01719
Zobiole LHS, Kremer RJ, Oliveira RS Jr, Constantin J (2011) Glyphosate affects micro-organisms in rhizospheres of glyphosate-resistant soybeans. J Appl Microbiol 110:118–127. https://doi.org/10.1111/j.1365-2672.2010.04864.x
Ekwomadu TI, Akinola SA, Mwanza M (2021) Fusarium mycotoxins, their metabolites (free, emerging, and masked), food safety concerns, and health impacts. Int J Environ Res Public Health. https://doi.org/10.3390/ijerph182211741
Ferrigo D, Raiola A, Causin R (2016) Fusarium toxins in cereals: occurrence, legislation, factors promoting the appearance and their management. Molecules 21:627. https://doi.org/10.3390/molecules21050627
Bruckner A, Schmerbauch A, Ruess L, Heigl F, Zaller J (2019) Foliar Roundup application has minor effects on the compositional and functional diversity of soil microorganisms in a short-term greenhouse experiment. Ecotox Env Safety 174:506–513. https://doi.org/10.1016/j.ecoenv.2019.02.073
Banks ML, Kennedy AC, Krerner RJ, Eivazi F (2014) Soil microbial community response to surfactants and herbicides in two soils. Appl Soil Ecol 74:12–20. https://doi.org/10.1016/j.apsoil.2013.08.018
Arango L et al (2014) Effects of glyphosate on the bacterial community associated with roots of transgenic Roundup Ready® soybean. Europ J Soil Biol 63:41–48. https://doi.org/10.1016/j.ejsobi.2014.05.005
Zabaloy M, Garland J, Gomez M (2012) An integrated approach to evaluate the impacts of the herbicides glyphosate, 2,4-D and metsulfuron-methyl on soil microbial communities in the Pampas region, Argentina. Appl Soil Ecol 40:1–12. https://doi.org/10.1016/j.apsoil.2008.1002.1004
Dennis PG, Kukulies T, Forstner C, Orton TG, Pattison AB (2018) The effects of glyphosate, glufosinate, paraquat and paraquat-diquat on soil microbial activity and bacterial, archaeal and nematode diversity. Sci Rep 8:2119. https://doi.org/10.1038/s41598-018-20589-6
Aristilde L, Reed ML, Wilkes RA, Youngster T, Kukurugya MA, Katz V, Sasaki CRS (2017) Glyphosate-induced specific and widespread perturbations in the metabolome of soil Pseudomonas species. Front Environ Sci. https://doi.org/10.3389/fenvs.2017.00034
Nicolas V, Oestreicher N, Vélot C (2016) Multiple effects of a commercial Roundup formulation on the soil filamentous fungus Aspergillus nidulans at low doses: evidence of an unexpected impact on energetic metabolism. Env Sci Poll Res Internat 23:14393–14404. https://doi.org/10.1007/s11356-016-6596-2
Carranza C, Barberis C, Chiacchiera S, Magnoli C (2017) Assessment of growth of Aspergillus spp. from agricultural soils in the presence of glyphosate. Rev Argent Microbiol 49:384–393. https://doi.org/10.1016/j.ram.2016.1011.1007
Nivelle E et al (2018) Effects of glyphosate application and nitrogen fertilization on the soil and the consequences on aboveground and belowground interactions. Geoderma 311:45–57. https://doi.org/10.1016/j.geoderma.2017.10.002
Vázquez MB, Moreno MV, Amodeo MR, Bianchinotti MV (2021) Effects of glyphosate on soil fungal communities: a field study. Rev Argentina Microbiol 53:349–358. https://doi.org/10.1016/j.ram.2020.10.005
Mendonca CM, Reed ML, Kukurugya MA, Aristilde L (2019) Adverse metabolic outcomes in soil pseudomonas species exposed to polyethoxylated tallow amine and glyphosate. Env Sci Technol Lett 6:448–455. https://doi.org/10.1021/acs.estlett.9b00363
Saunders LE, Pezeshki R (2015) Glyphosate in Runoff Waters and in the Root-Zone: A Review. Toxics 3:462–480. https://doi.org/10.3390/toxics3040462
Sesin V, Davy CM, Stevens KJ, Hamp R, Freeland JR (2021) Glyphosate toxicity to native nontarget macrophytes following three different routes of incidental exposure. Integr Environ Assess Manag 17:597–613. https://doi.org/10.1002/ieam.4350
Ferreira MF, Torres C, Bracamonte E, Galetto L (2017) Effects of the herbicide glyphosate on non-target plant native species from Chaco forest (Argentina). Ecotox Env Safety 144:360–368. https://doi.org/10.1016/j.ecoenv.2017.06.049
Cederlund H (2017) Effects of spray drift of glyphosate on nontarget terrestrial plants—a critical review. Env Toxicol Chem 36:2879–2886. https://doi.org/10.1002/etc.3925
White AL, Boutin C (2007) Herbicidal effects on nontarget vegetation: investigating the limitations of current pesticide registration guidelines. Env Toxicol Chem 26:2634–2643. https://doi.org/10.1897/06-553.1
Whitehead SR, Turcotte MM, Poveda K (2017) Domestication impacts on plant–herbivore interactions: a meta-analysis. Philos Trans Roy Soc B Biol Sci 372:20160034. https://doi.org/10.1098/rstb.2016.0034
Ke M et al (2021) Leaf metabolic influence of glyphosate and nanotubes on the Arabidopsis thaliana phyllosphere. J Env Sci 106:66–75. https://doi.org/10.1016/j.jes.2021.01.002
Saikkonen K, Nissinen R, Helander M (2020) Toward comprehensive plant microbiome research. Front Ecol Evol. https://doi.org/10.3389/fevo.2020.00061
Fuchs B, Laihonen M, Muola A, Saikkonen K, Dobrev PI, Vankova R, Helander M (2022) A Glyphosate-based herbicide in soil differentially affects hormonal homeostasis and performance of non-target crop plants. Front Plant Sci. https://doi.org/10.3389/fpls.2021.787958
Fuchs B, Saikkonen K, Helander M (2021) Glyphosate-modulated biosynthesis driving plant defense and species interactions. Trends Plant Sci 26:312–323. https://doi.org/10.1016/j.tplants.2020.11.004
Räsch A, Hunsche M, Mail M, Burkhardt J, Noga G, Pariyar S (2018) Agricultural adjuvants may impair leaf transpiration and photosynthetic activity. Plant Physiol Biochem 132:229–237. https://doi.org/10.1016/j.plaphy.2018.08.042
Seralini G-E, Jungers G (2021) Endocrine disruptors also function as nervous disruptors and can be renamed endocrine and nervous disruptors (ENDs). Toxicol Rep 8:1538–1557. https://doi.org/10.1016/j.toxrep.2021.07.014
Jungers G, Portet-Koltalo F, Cosme J, Seralini G-E (2022) Petroleum in pesticides: a need to change regulatory toxicology. Toxics 10:670
Tremearne T, Jacob K (1941) Arsenic in natural phosphates and phosphate fertilizer. United States Department of Agriculture, Washington. Techn Bull 781:41 pp
Marshall EJP, Brown VK, Boatman ND, Lutman PJW, Squire GR, Ward LK (2003) The role of weeds in supporting biological diversity within crop fields. Weed Res 43:77–89. https://doi.org/10.1046/j.1365-3180.2003.00326.x
Andreasen C, Streibig JC (2011) Evaluation of changes in weed flora in arable fields of Nordic countries—based on Danish long-term surveys. Weed Res 51:214–226. https://doi.org/10.1111/j.1365-3180.2010.00836.x
Semmens BX et al (2016) Quasi-extinction risk and population targets for the Eastern, migratory population of monarch butterflies (Danaus plexippus). Sci Rep 6:23265. https://doi.org/10.1038/srep23265
Pleasants JM, Zalucki MP, Oberhauser KS, Brower LP, Taylor OR, Thogmartin WE (2017) Interpreting surveys to estimate the size of the monarch butterfly population: pitfalls and prospects. PLoS ONE 12:e0181245. https://doi.org/10.1371/journal.pone.0181245
Yeates GW, Bongers T, De Goede RG, Freckman DW, Georgieva SS (1993) Feeding habits in soil nematode families and genera-an outline for soil ecologists. J Nematol 25:315–331
Zhao J, Neher D, Fu S, Za Li, Wang K-L (2013) Non-target effects of herbicides on soil nematode assemblages. Pest Manag Sci. https://doi.org/10.1002/ps.3505
Mbatyoti A, Daneel MS, Swart A, de Waele D, Fourie H (2018) Terrestrial non-parasitic nematode assemblages associated with glyphosate-tolerant and conventional soybean-based cropping systems. J Nematol 50:243–260. https://doi.org/10.21307/jofnem-2018-028
Hagner M, Mikola J, Saloniemi I, Saikkonen K, Helander M (2019) Effects of a glyphosate-based herbicide on soil animal trophic groups and associated ecosystem functioning in a northern agricultural field. Sci Rep 9:13. https://doi.org/10.1038/s41598-019-44988-5
Queirós L, Pereira JL, Gonçalves FJM, Pacheco M, Aschner M, Pereira P (2019) Caenorhabditis elegans as a tool for environmental risk assessment: emerging and promising applications for a “nobelized worm.” Crit Rev Toxicol 49:411–429. https://doi.org/10.1080/10408444.2019.1626801
Leung MC, Williams PL, Benedetto A, Au C, Helmcke KJ, Aschner M, Meyer JN (2008) Caenorhabditis elegans: an emerging model in biomedical and environmental toxicology. Toxicol Sci 106:5–28. https://doi.org/10.1093/toxsci/kfn121
McVey KA, Snapp IB, Johnson MB, Negga R, Pressley AS, Fitsanakis VA (2016) Exposure of C. elegans eggs to a glyphosate-containing herbicide leads to abnormal neuronal morphology. Neurotox Teratol 55:23–31. https://doi.org/10.1016/j.ntt.2016.03.002
Kronberg MF, Clavijo A, Moya A, Rossen A, Calvo D, Pagano E, Munarriz E (2018) Glyphosate-based herbicides modulate oxidative stress response in the nematode Caenorhabditis elegans. Comp Biochem Physiol C 214:1–8. https://doi.org/10.1016/j.cbpc.2018.08.002
García-Espiñeira M, Tejeda-Benitez L, Olivero-Verbel J (2018) Toxicity of atrazine- and glyphosate-based formulations on Caenorhabditis elegans. Ecotox Env Safety 156:216–222. https://doi.org/10.1016/j.ecoenv.2018.02.075
Naraine AS, Aker R, Sweeney I, Kalvey M, Surtel A, Shanbhag V, Dawson-Scully K (2022) Roundup and glyphosate’s impact on GABA to elicit extended proconvulsant behavior in Caenorhabditis elegans. Sci Rep 12:13655. https://doi.org/10.1038/s41598-022-17537-w
Mbatyoti A, Daneel MS, Swart A, Marais M, De Waele D, Fourie H (2019) Case study of effect of glyphosate application on plant-parasitic nematodes associated with a soybean–maize rotation system in South Africa. South Afric J Plant Soil 36:389–392. https://doi.org/10.1080/02571862.2019.1618505
Mbatyoti A, De Beer A, Daneel MS, Swart A, Marais M, De Waele D, Fourie H (2021) The host status of glyphosate-tolerant soybean genotypes to Meloidogyne incognita and Pratylenchus infection. Trop Plant Pathol 46:336–349. https://doi.org/10.1007/s40858-020-00416-y
Hopkin SP (1997) Biology of the springtails (Insecta: Collembola). Oxford University Press, Oxford, New York
Sanchez-Bayo F, Goka K (2016) Impacts of pesticides on honey bees. In: Chambo ED (ed) Beekeeping and bee conservation—advances in research. IntechOpen, London. https://doi.org/10.5772/62487
Sánchez-Bayo F, Goulson D, Pennacchio F, Nazzi F, Goka K, Desneux N (2016) Are bee diseases linked to pesticides?—A brief review. Env Int 89–90:7–11. https://doi.org/10.1016/j.envint.2016.01.009
Sánchez-Bayo F, Wyckhuys KAG (2019) Worldwide decline of the entomofauna: a review of its drivers. Biol Conserv 232:8–27. https://doi.org/10.1016/j.biocon.2019.01.020
Michalko R, Pekar S, Entling MH (2019) An updated perspective on spiders as generalist predators in biological control. Oecologia 189:21–36. https://doi.org/10.1007/s00442-018-4313-1
Santos MJG, Ferreira MFL, Cachada A, Duarte AC, Sousa JP (2012) Pesticide application to agricultural fields: effects on the reproduction and avoidance behaviour of Folsomia candida and Eisenia andrei. Ecotoxicol 21:2113–2122. https://doi.org/10.1007/s10646-012-0963-7
Haddaway NR et al (2017) How does tillage intensity affect soil organic carbon? A systematic review. Env Evidence 6:30. https://doi.org/10.1186/s13750-017-0108-9
Ximenes RL, Gomes AAA, Ximenes TSDS, Pires MSG (2020) Behavioral Analysis of Folsomia Candida (Collembola) with herbicide using electronic and computational instrumentation: bioassays. Soil Sediment Contam 29:545–556. https://doi.org/10.1080/15320383.2020.1748568
Fiera C et al (2020) Tillage intensity and herbicide application influence surface-active springtail (Collembola) communities in Romanian vineyards. Agric Ecosyst Environ 300:107006. https://doi.org/10.1016/j.agee.2020.107006
Haughton AJ, Bell JR, Wilcox A, Boatman ND (2001) The effect of the herbicide glyphosate on non-target spiders: Part I. Direct effects on Lepthyphantes tenuis under laboratory conditions. Pest Manag Sci 57:1033–1036. https://doi.org/10.1002/ps.388
Evans SC, Shaw EM, Rypstra AL (2010) Exposure to a glyphosate-based herbicide affects agrobiont predatory arthropod behaviour and long-term survival. Ecotoxicol 19:1249–1257. https://doi.org/10.1007/s10646-010-0509-9
Tahir HM, Basheer T, Ali S, Yaqoob R, Naseem S, Khan SY (2019) Effect of pesticides on biological control potential of Neoscona theisi (Araneae: Araneidae). J Insect Sci. https://doi.org/10.1093/jisesa/iez024
Niedobová J, Skalský M, Ouředníčková J, Michalko R, Bartošková A (2019) Synergistic effects of glyphosate formulation herbicide and tank-mixing adjuvants on Pardosa spiders. Env Poll 249:338–344. https://doi.org/10.1016/j.envpol.2019.03.031
de Aguiar LM, Figueira FH, Gottschalk MS, da Rosa CE (2016) Glyphosate-based herbicide exposure causes antioxidant defence responses in the fruit fly Drosophila melanogaster. Comp Biochem Physiol C 185–186:94–101. https://doi.org/10.1016/j.cbpc.2016.03.006
Stahlschmidt ZR, Whitlock J, Vo C, Evalen P, D B, (2022) Pesticides in a warmer world: effects of glyphosate and warming across insect life stages. Env Poll 307:119508. https://doi.org/10.1016/j.envpol.2022.119508
Smith DFQ et al (2021) Glyphosate inhibits melanization and increases susceptibility to infection in insects. PLOS Biol 19:e3001182. https://doi.org/10.1371/journal.pbio.3001182
Baglan H, Lazzari CR, Guerrieri FJ (2018) Glyphosate impairs learning in Aedes aegypti mosquito larvae at field-realistic doses. J Exp Biol. https://doi.org/10.1242/jeb.187518
McEwen P, New T, Whittington A (eds) (2001) Lacewings in the crop environment. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511666117
Goulson D (2019) The insect apocalypse, and why it matters. Curr Biol 29:R967–R971
Brühl CA, Zaller JG (2019) Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front Environ Sci 7:177. https://doi.org/10.3389/fenvs.2019.00177doi:10.3389/fenvs.2019.00177
Böhme F, Bischoff G, Zebitz CPW, Rosenkranz P, Wallner K (2018) Pesticide residue survey of pollen loads collected by honeybees (Apis mellifera) in daily intervals at three agricultural sites in South Germany. PLoS ONE 13:e0199995. https://doi.org/10.1371/journal.pone.0199995
David A, Botías C, Abdul-Sada A, Nicholls E, Rotheray EL, Hill EM, Goulson D (2016) Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Env Int 88:169–178. https://doi.org/10.1016/j.envint.2015.12.011
Battisti L et al (2021) Is glyphosate toxic to bees? A meta-analytical review. Sci Tot Environ 767:145397. https://doi.org/10.1016/j.scitotenv.2021.145397
Tan S, Li G, Liu Z, Wang H, Guo X, Xu B (2022) Effects of glyphosate exposure on honeybees. Env Toxicol Pharm 90:103792. https://doi.org/10.1016/j.etap.2021.103792
Farina WM, Balbuena MS, Herbert LT, Mengoni Goñalons C, Vázquez DE (2019) Effects of the herbicide glyphosate on honey bee sensory and cognitive abilities: individual impairments with implications for the hive. Insects 10:31635293. https://doi.org/10.3390/insects10100354
Thompson HM, Levine SL, Doering J, Norman S, Manson P, Sutton P, von Mérey G (2014) Evaluating exposure and potential effects on honeybee brood (Apis mellifera) development using glyphosate as an example. Integr Environ Assess Manag 10:463–470. https://doi.org/10.1002/ieam.1529
Zioga E, White B, Stout JC (2022) Glyphosate used as desiccant contaminates plant pollen and nectar of non-target plant species. Heliyon 8:e12179. https://doi.org/10.1016/j.heliyon.2022.e12179
Ledoux ML, Hettiarachchy N, Yu X, Howard L, Lee S-O (2020) Penetration of glyphosate into the food supply and the incidental impact on the honey supply and bees. Food Control 109:106859. https://doi.org/10.1016/j.foodcont.2019.106859
Abraham J, Benhotons GS, Krampah I, Tagba J, Amissah C, Abraham JD (2018) Commercially formulated glyphosate can kill non-target pollinator bees under laboratory conditions. Entomol Exp Appl 166:695–702. https://doi.org/10.1111/eea.12694
Motta EVS, Raymann K, Moran NA (2018) Glyphosate perturbs the gut microbiota of honey bees. PNAS 115:10305–10310. https://doi.org/10.1073/pnas.1803880115
Motta EVS et al (2020) Oral or topical exposure to glyphosate in herbicide formulation impacts the gut microbiota and survival rates of honey bees. Appl Environ Microbiol. https://doi.org/10.1128/aem.01150-20
Motta EVS, Moran NA (2020) Impact of glyphosate on the honey bee gut microbiota: effects of intensity, duration, and timing of exposure. mSystems 5:e00268-e1220. https://doi.org/10.1128/mSystems.00268-20
Blot N, Veillat L, Rouzé R, Delatte H (2019) Glyphosate, but not its metabolite AMPA, alters the honeybee gut microbiota. PLoS ONE 14:e0215466. https://doi.org/10.1371/journal.pone.0215466
Motta EVS, Moran NA (2023) The effects of glyphosate, pure or in herbicide formulation, on bumble bees and their gut microbial communities. Sci Tot Environ 872:162102. https://doi.org/10.1016/j.scitotenv.2023.162102
da Silva PC, Gonçalves B, Franceschinelli E, Brito P (2022) Glyphosate-based herbicide causes cellular alterations to gut epithelium of the neotropical stingless bee Melipona quadrifasciata quadrifasciata (Hymenoptera: Meliponini). Neotrop Entomol 51:860–868. https://doi.org/10.1007/s13744-022-01001-5
Motta EVS, Powell JE, Moran NA (2022) Glyphosate induces immune dysregulation in honey bees. Animal Microb 4:16. https://doi.org/10.1186/s42523-022-00165-0
Straw EA, Carpentier EN, Brown MJF (2021) Roundup causes high levels of mortality following contact exposure in bumble bees. J Appl Ecol 58:1167–1176. https://doi.org/10.1111/1365-2664.13867
Thompson LJ, Smith S, Stout JC, White B, Zioga E, Stanley DA (2022) Bumblebees can be exposed to the herbicide glyphosate when foraging. Env Toxicol Chem 41:2603–2612. https://doi.org/10.1002/etc.5442
Graffigna S, Marrero HJ, Torretta JP (2021) Glyphosate commercial formulation negatively affects the reproductive success of solitary wild bees in a Pampean agroecosystem. Apidologie 52:272–281. https://doi.org/10.1007/s13592-020-00816-8
Vázquez DE, Ilina N, Pagano EA, Zavala JA, Farina WM (2018) Glyphosate affects the larval development of honey bees depending on the susceptibility of colonies. PLoS ONE 13:e0205074. https://doi.org/10.1371/journal.pone.0205074
Wang B, Habermehl C, Jiang L (2022) Metabolomic analysis of honey bee (Apis mellifera L.) response to glyphosate exposure. Mol Omics 18:635–642. https://doi.org/10.1039/D2MO00046F
Balbuena MS, Tison L, Hahn M-L, Greggers U, Menzel R, Farina WM (2015) Effects of sublethal doses of glyphosate on honeybee navigation. J Exp Biol 218:2799–2805. https://doi.org/10.1242/jeb.117291
Zgurzynski MI, Lushington GH (2019) Glyphosate Impact on Apis mellifera navigation: a combined behavioral and cheminformatics study. Pharmacol Toxicol 7:806–824
Herbert LT, Vázquez DE, Arenas A, Farina WM (2014) Effects of field-realistic doses of glyphosate on honeybee appetitive behaviour. J Exper Biol 217:3457–3464. https://doi.org/10.1242/jeb.109520
Helander M et al (2023) Field-realistic acute exposure to glyphosate-based herbicide impairs fine-color discrimination in bumblebees. Sci Tot Environ 857:159298. https://doi.org/10.1016/j.scitotenv.2022.159298
Luo Q-H et al (2021) Effects of a commercially formulated glyphosate solutions at recommended concentrations on honeybee (Apis mellifera L.) behaviours. Sci Rep 11:2115. https://doi.org/10.1038/s41598-020-80445-4
Vázquez DE, Balbuena MS, Chaves F, Gora J, Menzel R, Farina WM (2020) Sleep in honey bees is affected by the herbicide glyphosate. Sci Rep 10:10516. https://doi.org/10.1038/s41598-020-67477-6
Weidenmüller A, Meltzer A, Neupert S, Schwarz A, Kleineidam C (2022) Glyphosate impairs collective thermoregulation in bumblebees. Science 376:1122–1126. https://doi.org/10.1126/science.abf7482
Hladik ML, Vandever M, Smalling KL (2016) Exposure of native bees foraging in an agricultural landscape to current-use pesticides. Sci Tot Environ 542:469–477. https://doi.org/10.1016/j.scitotenv.2015.10.077
Trouvé R, Drapela T, Frank T, Hadacek F, Zaller JG (2014) Herbivory of an invasive slug in a model grassland community can be affected by earthworms and mycorrhizal fungi. Biol Fertil Soils 50:13–23. https://doi.org/10.1007/s00374-013-0827-1
Zaller JG et al (2013) Herbivory of an invasive slug is affected by earthworms and the composition of plant communities. BMC Ecol 13:20
Druart C, Millet M, Scheifler R, Delhomme O, Vaufleury Ad (2011) Glyphosate and glufosinate-based herbicides: fate in soil, transfer to, and effects on land snails. J Soils Sediments 11:1373–1384
Druart C, Millet M, Scheifler R, Delhomme O, Raeppel C, de Vaufleury A (2011) Snails as indicators of pesticide drift, deposit, transfer and effects in the vineyard. Sci Tot Environ 409:4280–4288. https://doi.org/10.1016/j.scitotenv.2011.07.006
Ogeleka DF, Onwuemene CJ, Okieimen FE (2017) Toxicity potential of Grassate® a non-selective herbicide on snails (Achachatina marginata) and earthworms (Aporrectodea longa). Chem Ecol 33:447–463. https://doi.org/10.1080/02757540.2017.1320393
Baurand PE, Capelli N, de Vaufleury A (2015) Genotoxicity assessment of pesticides on terrestrial snail embryos by analysis of random amplified polymorphic DNA profiles. J Hazard Mat 298:320–327. https://doi.org/10.1016/j.jhazmat.2015.05.051
Dörler D et al (2021) Experimental indications of gardeners’ anecdotes that snails interfere with invasive slugs. PeerJ 9:e11309. https://doi.org/10.7717/peerj.11309
Dörler D, Scheucher A, Zaller JG (2019) Efficacy of chemical and biological slug control measures in response to watering and earthworms. Sci Rep 9:2954. https://doi.org/10.1038/s41598-019-39585-5
Wurst S, Sonnemann I, Zaller JG (2018) Soil macro-invertebrates—their impact on plants and associated aboveground communities in temperate regions. In: Ohgushi T, Wurst S, Johnson SN (eds) Aboveground-belowground community ecology, vol 234. Springer, Tokyo, pp 175–200. https://doi.org/10.1007/978-3-319-91614-9
Didden W, Rombke J (2001) Enchytraeids as indicator organisms for chemical stress in terrestrial ecosystems. Ecotox Env Safety 50:25–43. https://doi.org/10.1006/eesa.2001.2075
Pelosi C, Barot S, Capowiez Y, Hedde M, Vandenbulcke F (2014) Pesticides and earthworms. A review. Agron Sustain Develop 34:199–228
Correia FV, Moreira JC (2010) Effects of glyphosate and 2,4-D on earthworms (Eisenia foetida) in laboratory tests. Bull Environ Contam Toxicol 85:264–268
Piola L, Fuchs J, Oneto ML, Basack S, Kesten E, Casabé N (2013) Comparative toxicity of two glyphosate-based formulations to Eisenia andrei under laboratory conditions. Chemosphere 91:545–551
Casabé N et al (2007) Ecotoxicological assessment of the effects of glyphosate and chlorpyrifos in an Argentine soya field. J Soils Sediments 8:1–8
Muangphra P, Kwankua W, Gooneratne R (2014) Genotoxic effects of glyphosate or paraquat on earthworm coelomocytes. Env Toxicol 29:612–620. https://doi.org/10.1002/tox.21787
Pochron S et al (2019) Temperature and body mass drive earthworm (Eisenia fetida) sensitivity to a popular glyphosate-based herbicide. Appl Soil Ecol 139:32–39. https://doi.org/10.1016/j.apsoil.2019.03.015
Pochron ST et al (2021) Earthworms Eisenia fetida recover from Roundup exposure. Appl Soil Ecol 158:103793. https://doi.org/10.1016/j.apsoil.2020.103793
Lescano MR, Masin CE, Rodríguez AR, Godoy JL, Zalazar CS (2020) Earthworms to improve glyphosate degradation in biobeds. Env Sci Poll Res 27:27023–27031. https://doi.org/10.1007/s11356-020-09002-w
Santadino M, Coviella C, Momo F (2014) Glyphosate sublethal effects on the population dynamics of the earthworm Eisenia fetida (Savigny, 1826). Water Air Soil Poll 225:2207. https://doi.org/10.1007/s11270-014-2207-3
Stellin F, Gavinelli F, Stevanato P, Concheri G, Squartini A, Paoletti MG (2017) Effects of different concentrations of glyphosate (Roundup 360®) on earthworms (Octodrilus complanatus, Lumbricus terrestris and Aporrectodea caliginosa) in vineyards in the North-East of Italy. Appl Soil Ecol. doi:https://doi.org/10.1016/j.apsoil.2017.07.028
Owagboriaye F (2020) Wormcasts produced by three earthworm species (Alma millsoni, Eudrilus eugeniae and Libyodrilus violaceus) exposed to a glyphosate-based herbicide reduce growth, fruit yield and quality of tomato (Lycopersicon esculentum). Chemosphere 250:126270
Zaller JG, Heigl F, Ruess L, Grabmaier A (2014) Glyphosate herbicide affects belowground interactions between earthworms and symbiotic mycorrhizal fungi in a model ecosystem. Sci Rep 4:5634. https://doi.org/10.1038/srep05634
Hackenberger DK, Stjepanović N, Lončarić Ž, Hackenberger BK (2018) Acute and subchronic effects of three herbicides on biomarkers and reproduction in earthworm Dendrobaena veneta. Chemosphere 208:722–730. https://doi.org/10.1016/j.chemosphere.2018.06.047
Gaupp-Berghausen M, Hofer M, Rewald B, Zaller JG (2015) Glyphosate-based herbicides reduce the activity and reproduction of earthworms and lead to increased soil nutrient concentrations. Sci Rep 5:12886. https://doi.org/10.1038/srep12886
Owagboriaye F et al (2021) Impacts of a glyphosate-based herbicide on the gut microbiome of three earthworm species (Alma millsoni, Eudrilus eugeniae and Libyodrilus violaceus): a pilot study. Toxicol Rep 8:753–758. https://doi.org/10.1016/j.toxrep.2021.03.021
Römbke J, Schmelz RM, Pélosi C (2017) Effects of organic pesticides on Enchytraeids (Oligochaeta) in agroecosystems: laboratory and higher-tier tests. Front Environ Sci 5:20. https://doi.org/10.3389/fenvs.2017.00020
Bart S, Amossé J, Lowe CN, Mougin C, Péry ARR, Pelosi C (2018) Aporrectodea caliginosa, a relevant earthworm species for a posteriori pesticide risk assessment: current knowledge and recommendations for culture and experimental design. Env Sci Poll Res 25:33867–33881. https://doi.org/10.1007/s11356-018-2579-9
Pelosi C, Joimel S, Makowski D (2013) Searching for a more sensitive earthworm species to be used in pesticide homologation tests—a meta-analysis. Chemosphere 90:895–900. https://doi.org/10.1016/j.chemosphere.2012.09.034
FAO, ITPS (2017) Global assessment of the impact of plant protection products on soil functions and soil ecosystems. Food and Agriculture Organization of the United Nations, Rome
Fryday S, Thompson H (2012) Toxicity of pesticides to aquatic and terrestrial life stages of amphibians and occurrence, habitat use and exposure of amphibian species in agricultural environments. EFSA Supporting Publications EN-343:348pp
Brühl CA, Schmidt T, Pieper S, Alscher A (2013) Terrestrial pesticide exposure of amphibians: an underestimated cause of global decline? Sci Rep 3:1135. https://doi.org/10.1038/srep01135
EFSA et al (2018) Scientific Opinion on the state of the science on pesticide risk assessment for amphibians and reptiles. EFSA J 16:e05125. https://doi.org/10.2903/j.efsa.2018.5125
Carpenter JK, Monks JM, Nelson N (2016) The effect of two glyphosate formulations on a small, diurnal lizard (Oligosoma polychroma). Ecotoxicol 25:548–554. https://doi.org/10.1007/s10646-016-1613-2
Paranjpe DA, Medina D, Nielsen E, Cooper RD, Paranjpe SA, Sinervo B (2014) Does thermal ecology influence dynamics of side-blotched lizards and their micro-parasites? Integrat Compar Biol 54:108–117. https://doi.org/10.1093/icb/icu069
Preest Marion R, Cree A (2008) Corticosterone treatment has subtle effects on thermoregulatory behavior and raises metabolic rate in the New Zealand Common Gecko, Hoplodactylus maculatus. Physiol Biochem Zool 81:641–650. https://doi.org/10.1086/590371
Wang Z et al (2023) Consolidation of temperature-dependent toxicity and thermoregulatory behavior into risk assessments of insecticides under thermal scenarios: a prospective study on Eremias argus. Env Int 172:107742. https://doi.org/10.1016/j.envint.2023.107742
Schaumburg L, Siroski P, Poletta G, Mudry M (2016) Genotoxicity induced by Roundup® (Glyphosate) in tegu lizard (Salvator merianae) embryos. Pestic Biochem Physiol 130:71–78. https://doi.org/10.1016/j.pestbp.2015.1011.1009
Verderame M, Scudiero R (2019) How glyphosate impairs liver condition in the field lizard Podarcis siculus (Rafinesque-Schmaltz, 1810): histological and molecular evidence. BioMed Res Int 2019:4746283. https://doi.org/10.1155/2019/4746283
Verderame M, Chianese T, Rosati L, Scudiero R (2022) Molecular and histological effects of glyphosate on testicular tissue of the lizard Podarcis siculus. Int J Mol Sci 23:4850
Weir S, Yu S, Knox A, Talent L, Monks J, Salice C (2016) Acute toxicity and risk to lizards of rodenticides and herbicides commonly used in New Zealand. NZ J Ecol Res 40:342–350. https://doi.org/10.20417/nzjecol.20440.20443
Lajmanovich RC et al (2019) First evaluation of novel potential synergistic effects of glyphosate and arsenic mixture on Rhinella arenarum (Anura: Bufonidae) tadpoles. Heliyon. https://doi.org/10.1016/j.heliyon.2019.e02601
Rissoli RZ, Abdalla FC, Costa MJ, Rantin FT, McKenzie DJ, Kalinin AL (2016) Effects of glyphosate and the glyphosate based herbicides Roundup Original® and Roundup Transorb® on respiratory morphophysiology of bullfrog tadpoles. Chemosphere 156:37–44. https://doi.org/10.1016/j.chemosphere.2016.04.083
Baier F, Jedinger M, Gruber E, Zaller JG (2016) Temperature-dependence of glyphosate-based herbicide’s effects on egg and tadpole growth of Common Toads. Front Environ Sci 4:51. https://doi.org/10.3389/fenvs.2016.00051
Relyea RA (2012) New effects of Roundup on amphibians: Predators reduce herbicide mortality; herbicides induce antipredator morphology. Ecol Appl 22:634–647
Lopes A et al (2021) Evaluation of the genotoxic, mutagenic, and histopathological hepatic effects of polyoxyethylene amine (POEA) and glyphosate on Dendropsophus minutus tadpoles. Env Poll 289:117911. https://doi.org/10.1016/j.envpol.2021.117911
Pérez-Iglesias JM, Franco-Belussi L, Moreno L, Tripole S, de Oliveira C, Natale GS (2016) Effects of glyphosate on hepatic tissue evaluating melanomacrophages and erythrocytes responses in neotropical anuran Leptodactylus latinasus. Env Sci Poll Res 23:9852–9861. https://doi.org/10.1007/s11356-016-6153-z
Lajmanovich RC et al (2015) Harmful effects of the dermal intake of commercial formulations containing chlorpyrifos, 2,4-D, and glyphosate on the common toad Rhinella arenarum (Anura: Bufonidae). Water Air Soil Poll 226:427. https://doi.org/10.1007/s11270-015-2695-9
Moore LJ, Fuentes L, Rodgers JH, Bowerman WW, Yarrow GK, Chao WY, Bridges WC (2012) Relative toxicity of the components of the original formulation of Roundup® to five North American anurans. Ecotox Env Safety 78:128–133. https://doi.org/10.1016/j.ecoenv.2011.11.025
Panzacchi S et al (2018) The Ramazzini Institute 13-week study on glyphosate-based herbicides at human-equivalent dose in Sprague Dawley rats: study design and first in-life endpoints evaluation. Env Health 17:52. https://doi.org/10.1186/s12940-018-0393-y
Aitbali Y, Ba-M’hamed S, Elhidar N, Nafis A, Soraa N, Bennis M (2018) Glyphosate based- herbicide exposure affects gut microbiota, anxiety and depression-like behaviors in mice. Neurotox Teratol 67:44–49. https://doi.org/10.1016/j.ntt.2018.04.002
Mesnage R et al (2022) Glyphosate and its formulations Roundup Bioflow and RangerPro alter bacterial and fungal community composition in the rat caecum microbiome. Front Microbiol 13:888853. https://doi.org/10.3389/fmicb.2022.888853
Hu J et al (2021) Low-dose exposure of glyphosate-based herbicides disrupt the urine metabolome and its interaction with gut microbiota. Sci Rep 11:3265. https://doi.org/10.1038/s41598-021-82552-2
Sanam N et al (2019) Effect of glyphosate on hematological and biochemical parameters of Rabbit (Oryctolagus cuniculus). Pure Appl Biol 8:78–92
Pham TH, Derian L, Kervarrec C, Kernanec P-Y, Jégou B, Smagulova F, Gely-Pernot A (2019) Perinatal exposure to glyphosate and a glyphosate-based herbicide affect spermatogenesis in mice. Toxicol Sci 169:260–271. https://doi.org/10.1093/toxsci/kfz039
Ganesan S, Keating AF (2020) Ovarian mitochondrial and oxidative stress proteins are altered by glyphosate exposure in mice. Toxicol Appl Pharmacol 402:115116. https://doi.org/10.1016/j.taap.2020.115116
Milesi MM, Lorenz V, Pacini G, Repetti MR, Demonte LD, Varayoud J, Luque EH (2018) Perinatal exposure to a glyphosate-based herbicide impairs female reproductive outcomes and induces second-generation adverse effects in Wistar rats. Arch Toxicol 92:2629–2643. https://doi.org/10.1007/s00204-018-2236-6
Kubsad D, Nilsson EE, King SE, Sadler-Riggleman I, Beck D, Skinner MK (2019) Assessment of glyphosate induced epigenetic transgenerational inheritance of pathologies and sperm epimutations: generational toxicology. Sci Rep 9:6372. https://doi.org/10.1038/s41598-019-42860-0
Luaces JP, Rossi LF, Chirino MG, Browne M, Merani MS, Mudry MD (2017) Genotoxic effects of Roundup Full II® on lymphocytes of Chaetophractus villosus (Xenarthra, Mammalia): In vitro studies. PLoS ONE 12:e0182911. https://doi.org/10.1371/journal.pone.0182911
Rossi LF, Luaces JP, Palermo AM, Merani MS, Mudry MD (2018) Cytogenetic damage in peripheral blood cultures of Chaetophractus villosus exposed in vivo to a glyphosate formulation (Roundup). Ecotox Env Safety 157:121–127. https://doi.org/10.1016/j.ecoenv.2018.03.046
Ruuskanen S, Rainio MJ, Uusitalo M, Saikkonen K, Helander M (2020) Effects of parental exposure to glyphosate-based herbicides on embryonic development and oxidative status: a long-term experiment in a bird model. Sci Rep. https://doi.org/10.1038/s41598-020-63365-1
Ruuskanen S, Rainio MJ, Kuosmanen V, Laihonen M, Saikkonen K, Saloniemi I, Helander M (2020) Female preference and adverse developmental effects of glyphosate-based herbicides on ecologically relevant traits in Japanese Quails. Env Sci Technol 54:1128–1135. https://doi.org/10.1021/acs.est.9b07331
Ruuskanen S et al (2020) Glyphosate-based herbicides influence antioxidants, reproductive hormones and gut microbiome but not reproduction: a long-term experiment in an avian model. Env Poll 266:115108. https://doi.org/10.1016/j.envpol.2020.115108
Serra L et al (2021) Chronic dietary exposure of roosters to a glyphosate-based herbicide increases seminal plasma glyphosate and AMPA concentrations, alters sperm parameters, and induces metabolic disorders in the progeny. Toxics. https://doi.org/10.3390/toxics9120318
Shehata AA, Schrodl W, Aldin AA, Hafez HM, Krüger M (2013) The effect of glyphosate on potential pathogens and beneficial members of poultry microbiota in vitro. Curr Microbiol 66:350–358. https://doi.org/10.1007/s00284-012-0277-2
van Hoesel W et al (2017) Single and combined effects of pesticide seed dressings and herbicides on earthworms, soil microorganisms, and litter decomposition. Front Plant Sci 8:215. https://doi.org/10.3389/fpls.2017.00215
Pankey JH, Griffin JL, Leonard BR, Miller DK, Downer RG, Costello RW (2004) Glyphosate-insecticide combination effects on weed and insect control in cotton. Weed Techn 18:698–703
Almasri H et al (2021) Toxicological status changes the susceptibility of the honey bee Apis mellifera to a single fungicidal spray application. Env Sci Poll Res 28:42807–42820. https://doi.org/10.1007/s11356-021-13747-3
Almasri H et al (2020) Mixtures of an insecticide, a fungicide and a herbicide induce high toxicities and systemic physiological disturbances in winter Apis mellifera honey bees. Ecotox Env Safety 203:111013. https://doi.org/10.1016/j.ecoenv.2020.111013
Hao Y, Zhang Y, Xu W, Gao J, Tao L (2019) Synergistic effects of adjuvant A-134 on the herbicidal effects of glyphosate. J Pestic Sci 44:249–254. https://doi.org/10.1584/jpestics.D19-030
Castro MJL, Ojeda C, Cirelli AF (2014) Advances in surfactants for agrochemicals. Env Chem Lett 12:85–95. https://doi.org/10.1007/s10311-013-0432-4
Carvalho WF et al (2018) Evaluation of genotoxic and mutagenic effects of glyphosate roundup Original® in Dendropsophus minutus Peters, 1872 Tadpoles. South Am J Herpetol 13(220–229):210
Song HY, Kim YH, Seok SJ, Gil HW, Hong SY (2012) In vitro cytotoxic effect of glyphosate mixture containing surfactants. J Korean Med Sci 27:711–715. https://doi.org/10.3346/jkms.2012.27.7.711
Peillex C, Pelletier M (2020) The impact and toxicity of glyphosate and glyphosate-based herbicides on health and immunity. J Immunotoxicol 17:163–174. https://doi.org/10.1080/1547691X.2020.1804492
Toy A, Uhing E (1964) Aminomethylenephosphinic acids, salts thereof, and process for their production. United States Patent Office, Patent 3(160):632
Mertens M, Höss S, Neumann G, Afzal J, Reichenbecher W (2018) Glyphosate, a chelating agent—relevant for ecological risk assessment? Env Sci Poll Res 25:5298–5317. https://doi.org/10.1007/s11356-017-1080-1
Zhou CF, Wang YJ, Yu YC, Sun RJ, Zhu XD, Zhang HL, Zhou DM (2012) Does glyphosate impact on Cu uptake by, and toxicity to, the earthworm Eisenia fetida? Ecotoxicol 21:2297–2305. https://doi.org/10.1007/s10646-012-0986-0
Zhou CF, Wang YJ, Li CC, Sun RJ, Yu YC, Zhou DM (2013) Subacute toxicity of copper and glyphosate and their interaction to earthworm (Eisenia fetida). Env Poll 180:71–77. https://doi.org/10.1016/j.envpol.2013.05.016
Szabó R, Csonka D, Major L, Lehel J, Budai P (2020) Toxicity test of individual and combined toxic effects of glyphosate herbicide and heavy metals on chicken embryos. Agrofor Int J 5:3. https://doi.org/10.7251/AGRENG2003064S
EFSA (2015) Conclusion on the peer review of the pesticide risk assessment of the active substance glyphosate. EFSA J 13:4302. https://doi.org/10.2903/j.efsa.2015.4302
EFSA (2015) Final addendum to the Renewal Assessment Report - public version. Risk assessment provided by the rapporteur Member State Germany and co-rapporteur Member State Slovakia for the active substance Glyphosate according to the procedure for the renewal of the inclusion of a second group of active substances in Annex I to Council Directive 91/414/EEC laid down in Commission regulation (EU) No. 1141/2010, October 2015. ECHA, http://registerofquestions.efsa.europa.eu/roqFrontend/outputLoader?output=ON-4302. Accessed 14 June 2023
Eker S, Ozturk L, Yazici A, Erenoglu B, Romheld V, Cakmak I (2006) Foliar-applied glyphosate substantially reduced uptake nd transport of iron and manganese in sunflower (Helianthus annuus L.) plants. J Ag Food Chem. https://doi.org/10.1021/jf0625196
Tesfamariam T, Bott S, Cakmak I, Römheld V, Neumann G (2009) Glyphosate in the rhizosphere—role of waiting times and different glyphosate binding forms in soils for phytotoxicity to non-target plants. Europ J Agron 31:126–132. https://doi.org/10.1016/j.eja.2009.03.007
Jayasumana C, Gunatilake S, Siribaddana S (2015) Simultaneous exposure to multiple heavy metals and glyphosate may contribute to Sri Lankan agricultural nephropathy. BMC Nephrol 16:103. https://doi.org/10.1186/s12882-015-0109-2
Liu H et al (2019) Interactive effects of microplastics and glyphosate on the dynamics of soil dissolved organic matter in a Chinese loess soil. CATENA. https://doi.org/10.1016/j.catena.2019.104177
Yang X et al (2018) Influence of microplastic addition on glyphosate decay and soil microbial activities in Chinese loess soil. Env Poll 242:338–347. https://doi.org/10.1016/j.envpol.2018.07.006
Yang X et al (2019) Biogenic transport of glyphosate in the presence of LDPE microplastics: a mesocosm experiment. Env Poll 245:829–835. https://doi.org/10.1016/j.envpol.2018.11.044
van Bruggen A, Finckh M, He M, Ritsema C, Knuth D, Harkes P, Geissen V (2021) Indirect effects of the herbicide glyphosate on plant, animal and human health through its effects on microbial communities. Front Environ Sci 9:763917. https://doi.org/10.3389/fenvs.2021.763917
Faita MR, Cardozo MM, Amandio DTT, Orth AI, Nodari RO (2020) Glyphosate-based herbicides and Nosema sp. microsporidia reduce honey bee (Apis mellifera L.) survivability under laboratory conditions. J Apicult Res 59:332–342. https://doi.org/10.1080/00218839.2020.1736782
Straw EA, Brown MJF (2021) No evidence of effects or interaction between the widely used herbicide, glyphosate, and a common parasite in bumble bees. PeerJ 9:e12486. https://doi.org/10.7717/peerj.12486
Kurenbach B, Gibson PS, Hill AM, Bitzer AS, Silby MW, Godsoe W, Heinemann JA (2017) Herbicide ingredients change Salmonella enterica sv. Typhimurium and Escherichia coli antibiotic responses. Microbiol 163:1791–1801. https://doi.org/10.1099/mic.0.000573
Kurenbach B, Marjoshi D, Amabile-Cuevas CF, Ferguson GC, Godsoe W, Gibson P (2015) Sublethal exposure to commercial formulations of the herbicides dicamba, 2,4-dichlorophenoxyacetic acid, and glyphosate cause changes in antibiotic susceptibility in Escherichia coli and Salmonella enterica serovar typhimurium. MBio. https://doi.org/10.1128/mBio.00009-15
Raoult D, Hadjadj L, Baron SA, Rolain JM (2021) Role of glyphosate in the emergence of antimicrobial resistance in bacteria? J Antimicrob Chemother 76:1655–1657. https://doi.org/10.1093/jac/dkab102
Van Bruggen AHC, He MM, Shin K, Mai V, Jeong KC, Finckh MR, Morris JG (2018) Environmental and health effects of the herbicide glyphosate. Sci Tot Environ 616–617:255–268. https://doi.org/10.1016/j.scitotenv.2017.10.309
Donley N (2019) The USA lags behind other agricultural nations in banning harmful pesticides. Env Health 18:44. https://doi.org/10.1186/s12940-019-0488-0
Zaller JG (2020) Daily poison. Pesticides—an underestimated danger. Springer Nature, Cham. https://doi.org/10.1007/978-3-030-50530-1
IPBES (2019) Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, IPBES secretariat, Bonn, Germany; https://doi.org/10.5281/zenodo.3831673
Carvalho FP (2020) Glyphosate, the herbicide that become a nightmare and the Precautionary Principle. Int J Env Stud 77:1012–1023. https://doi.org/10.1080/00207233.2020.1773682
Acknowledgements
GBHs mentioned in this review
Agpro Glyphosate 360; Atanor; Bypass; Crucial; Glialka Star; Glifosato Atanor II; Glyphogan; Grassate; Medallon; MON 8750; Montana; RangerPro; Rodeo XL; Roundup; Roundup 3 Plus; Roundup 360; Roundup Alphée; Roundup Bioflow; Roundup Classic; Roundup ControlMax; Roundup FG; Roundup Flash; RoundUp Flex; Roundup Full; Roundup Full II; Roundup Gold; Roundup Grands Travaux plus; Roundup Klasik Pro; Roundup LB Plus; Roundup Original; Roundup PowerFlex; Roundup PowerMax; Roundup ProActive; Roundup R450; Roundup Ready-To-Use; RoundupReady-To-Use III; Roundup Speed; Roundup Super Concentrate; Roundup UltraMax; Roundup WeatherMax; Sunphosate 360 SL; Touchdown; Touchdown Quattro; Trop; Weedol; Yates Roundup Weedkiller; Zapp.
We thank the four anonymous reviewers whose comments helped improve this paper.
Funding
Open access funding provided by University of Natural Resources and Life Sciences Vienna (BOKU). This work was supported by Erasmus + project Hort4EUGreen (2020-1-RO01-KA203-080398).
Author information
Authors and Affiliations
Contributions
SzK: conceptualization; writing—original draft; review and editing. GS: conceptualization; writing—original draft. MO: writing—original draft. RM: supervision; writing—review and editing. JGZ: supervision; writing—review and editing. ASz: supervision; writing—review and editing. MNA: writing, review and editing. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
R.M. has served as a consultant on glyphosate risk assessment issues as part of litigation in the United States over glyphosate health effects. The other authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Klátyik, S., Simon, G., Oláh, M. et al. Terrestrial ecotoxicity of glyphosate, its formulations, and co-formulants: evidence from 2010–2023. Environ Sci Eur 35, 51 (2023). https://doi.org/10.1186/s12302-023-00758-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12302-023-00758-9