
Abstract
Despite recent advances in tumor diagnosis and treatment technologies, the number of cancer cases and deaths worldwide continues to increase yearly, creating an urgent need to find new methods to prevent or treat cancer. Sulforaphane (SFN), as a member of the isothiocyanates (ITCs) family, which is the hydrolysis product of glucosinolates (GLs), has been shown to have significant preventive and therapeutic cancer effects in different human cancers. Early studies have shown that SFN scavenges oxygen radicals by increasing cellular defenses against oxidative damage, mainly through the induction of phase II detoxification enzymes by nuclear factor erythroid 2-related factor 2 (Nrf2). More and more studies have shown that the anticancer mechanism of SFN also includes induction of apoptotic pathway in tumor cells, inhibition of cell cycle progression, and suppression of tumor stem cells. Therefore, the application of SFN is expected to be a necessary new approach to treating cancer. In this paper, we review the multiple molecular mechanisms of SFN in cancer prevention and treatment in recent years, which can provide a new vision for cancer treatment.
Similar content being viewed by others


Introduction
According to epidemiologic data, there were 10.3 million cancer-related deaths worldwide in 2020 and 19.3 million new cancer diagnoses (Sung et al. 2021). By 2030, it is anticipated that there will be 21.6 million additional cases and 13 million fatalities (Fidler et al. 2018). Notably, despite significant advances in modern diagnostic techniques and therapeutic strategies, cancer mortality and morbidity remain high, and cancer is recognized as a significant challenge in all countries (Baig et al. 2019). Currently, practical methods for treating malignant tumors include surgery, radiotherapy, and chemotherapy agents. Among them, chemical drugs show beneficial therapeutic effects in cancer treatment. However, they also produce drug resistance and severe side effects, which finally lead to the failure of treatment (Yan et al. 2016; Wang et al. 2018). Therefore, searching for new drugs and strategies for cancer treatment remains an urgent problem.
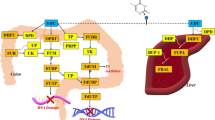
A naturally occurring active substance known as isothiocyanate (ITC) originates from cruciferous vegetables, including broccoli, cabbage, and kale (Lam et al. 2009). Sulforaphane (SFN), a member of the ITCs family, is present in broccoli as glucosinolates (GLs). When broccoli is damaged, e.g., by chewing or chopping resulting in cellular fragmentation, it activates endogenous black mustard enzymes that hydrolyze the GLs and convert them to SFN (Jed et al. 2001; Bones and Rossiter 2006) (Fig. 1). SFN is a biologically active small molecule compound with anti-inflammatory and antioxidant properties (Carlos-Reyes et al. 2019; Liebman and Le 2021). SFN has been shown to play a cancer chemopreventive role by inducing phase II detoxification and antioxidant enzymes through the Nrf2/ARE signaling pathway (Brooks et al. 2001). At the same time, SFN can inhibit phase I enzymes that activate procarcinogens, thereby interfering with the initiation stage of cancer (Langoue et al. 2000). In addition to its preventive effects on cancer, SFN has recently been shown to inhibit tumor growth and progression by modulating multiple pathways associated with cancer development (Briones-Herrera et al. 2018; Negrette-Guzman 2019; Rafiei et al. 2020). Furthermore, several studies have reported that SFN can also be used as a natural dietary supplement taken with some norm chemotherapeutic medicines to increase therapeutic effectiveness while reducing their adverse side effects (Bose et al. 2018; Mielczarek et al. 2019; Xu et al. 2019; Justin et al. 2020).

Conversion of glucosinolates to sulforaphane by hydrolysis of black mustard enzyme
For these reasons, we reviewed the progress of research on the complex molecular mechanisms of SFN in various cancers (Fig. 2; Table 1). A better understanding of the complexity of these mechanisms will provide possible opportunities for cancer therapy. We hope this review will open new horizons for applying SFN in cancer treatment and prevention.

Overview of the multiple molecular mechanisms of the plant natural compound sulforaphane for cancer prevention and treatment, such as inhibiting tumor cell proliferation, arresting the invasive migration ability of tumor cells, inducing programmed cell death, inhibiting tumor stem cells, and regulating tumor cell energy metabolism
Regulation of tumor cell proliferative capacity by SFN
It is generally recognized that cancer cells arise from normal cells with abnormal proliferation and survival signaling pathways. They accumulate mutations during proliferation, eventually forming malignant tumor cells that can replicate indefinitely (Rycaj and Tang 2015). One of the foremost aggressive characteristics of tumor cells is proliferation, and numerous studies have shown that SFN can prevent tumor cells from developing via various processes.
The PI3K/Akt signaling pathway coordinates many physiological processes in cells, including migration, metabolism, cell viability, and proliferation (Han et al. 2018). Meanwhile, the PI3K/Akt signaling pathway promotes tumorigenesis by regulating its downstream effectors (Wang et al. 2023). Micromolar SFN concentrations inhibit endometrial epithelial tumor cell growth in vivo and in vitro by inhibiting Akt, mTOR, and ribosomal protein S6 kinase (S6K) phosphorylation (Rai et al. 2020). In human osteosarcoma cells, Zhang et al. demonstrated that SFN suppressed cell proliferation in a concentration-related way. The mechanism is that SFN inhibits cytoplasmic histone deacetylase 6 (HDAC6) and promotes Akt acetylation, decreasing the kinase catalytic activity of Akt to inhibit mTOR (Zhang et al. 2021). In conclusion, SFN would prevent tumor cell growth by regulating Akt signaling.
SFN can regulate different cancer cells’ proliferation at cell cycle stages. In human ovarian cancer, Kan et al. revealed that the SFN-treated group had considerably higher Bax, P53, and P27 expression levels. Bcl-2, Cyclin-D1, cMyc, and Her2 expression, on the other hand, were reduced in vivo and in vitro (Kan et al. 2018). According to a different study, SFN significantly increased the expression of P53 and P21 in gastric cancer cells while inhibiting the S-phase of the cell cycle and promoting apoptosis (Wang et al. 2021b). Interestingly, SFN alters the CDK-cytolytic protein axis, P19 and P27, and the expression of CD44 variations in prostate cancer cells, prompting cell cycle arrest in the S and G2/M phases and suppressing growth and proliferation (Rutz et al. 2020). SFN’s impact on cervical cancer cells was investigated by Cheng et al. Cyclin-B1 expression, CDC25C dephosphorylation, and remodeling of the GADD45/CDC2 complex were all restricted by SFN, which also caused a cell cycle halt in the G2/M phase and reduced proliferation (Cheng et al. 2016). Thus, SFN may inhibit key antitumor factors in vivo, such as P53 and P27 (Psyrri et al. 2005; Kandoth et al. 2013).
Of note, oxidative stress is upregulated in human cancers associated with elevated reactive oxygen species (ROS) and promoting DNA damage and mutation (Obtulowicz et al. 2010). In malignant gliomas, SFN inhibits tumor cell growth by inducing mitochondrial ROS accumulation and DNA damage, suppressing malignant glioma growth in vivo and in vitro (Bijangi-Vishehsaraei et al. 2017). Two studies in mice have shown that cellular senescence, an irreversible blockage of cell proliferation, is feasible as a tumor suppression strategy (Baker et al. 2016; Demaria et al. 2017). Senescence can inhibit tumorigenesis by limiting the malignant transformation of precancerous cells and blocking the proliferation of cancer cells. SFN inhibits esophageal cancer cells proliferation, triggering senescence of esophageal cancer cells by promoting ROS accumulation to induce DNA damage, modulating the mTOR/TEF3 axis to interfere with autophagy, and promoting lysosomal abnormalities (Zheng et al. 2020). Research in prostate cancer cells found that SFN induced more pronounced DNA double-strand breaks in cancer cells than in normal cells, mainly because DNA repair is more efficient in normal cells than in cancer cells, allowing SFN to inhibit cancer cell proliferation selectively in vitro (Hac et al. 2020). In summary, SFN can inhibit tumor cell proliferation by inhibiting Akt signaling, blocking the cell cycle, and inducing cellular DNA damage, providing new ideas for tumor treatment.
Regulation of tumor cell migratory and invasive capacity by SFN
Another of the primary characteristics of many malignant tumors is migration and invasion. Cancer metastatic cells separate from the original tumor location, infiltrate mesenchymal tissues, and enter the bloodstream to start new cancer cell colonies in distant organs. Vascular endothelial growth factor (VEGF) is a crucial factor in encouraging angiogenesis, and the release of VEGF by tumor cells regulates angiogenesis in the formation of malignancies (Ahmad and Nawaz 2022). Under hypoxic conditions, VEGF is a critical protein that functions downstream of hypoxia-inducible factor-1 (HIF-1α). HIF-1α-induced up-regulation of VEGF expression activated by a hypoxic environment was inhibited by SFN, resulting in decreased migration of gastric cancer cell lines AGS and HCT116 under hypoxic conditions (Kim et al. 2015).
Elevated matrix metalloproteinase (MMP) expression in tumor cells represents higher invasion and metastasis. According to research on melanoma in vivo and in vitro, SFN therapy reduced MMP-2 and MMP-9 levels while increasing the levels of TIMP3, an MMPs inhibitor (Fisher et al. 2016). Similarly, SFN reduced invasion in gastric cancer cells by blocking the AP-1 and NF-κB signaling pathways, activated by ROS, and by reducing the expression of MMP-9 stimulated by nicotine (Li et al. 2022). Related experiments in human breast cancer cells and nude mice also demonstrated that SFN inhibited breast cancer development by inhibiting NF-κB and suppressing MMP-9 expression (Zhou et al. 2022). Additionally, because glioblastoma is a highly vascularized malignancy, SFN quickly crosses the blood-brain barrier (BBB) and builds up in the central nervous system. These effects include lowering MMP-9 release, preventing angiogenesis and invasion of glioblastoma cells, and overcoming chemoresistance (Li et al. 2014; Zhang et al. 2016).
Epithelial cells lose connection and polarity, restructure the cytoskeleton, acquire motility, and transition into the mesenchymal cellular morphology to develop an invasive phenotype. This process is known as epithelial-mesenchymal transition (EMT) (Lamouille et al. 2014). SFN inhibits the onset of EMT and invasion in advanced human prostate cancer cells by activating ERK1/2 to upregulate E-cadherin and down-regulate CD44v6 in vitro (Peng et al. 2015). Meanwhile, SFN treatment also decreased CD44v6 and YAP1 levels and their downstream genes, reducing cutaneous squamous carcinoma cell and NSG mice models’ properties and EMT markers, inhibiting cell sphere formation, invasion, and migration (Chen et al. 2023).
The processing, localization, and regulation of specific miRNAs are closely related to cancer migration and invasion (Ohtsuka et al. 2015). SFN suppresses the EMT process and the ability to metastasize non-small cell lung cancer in vivo and in vitro by altering the expression of the miR-616-5p/GSK3β/β-catenin pathway. This leads to an increase in the face of E-cadherin and a decrease in the expression of β-cadherin, N-cadherin, and vimentin (Wang et al. 2017). SFN induces overexpression of novel tumor suppressors miR-135b-5p and RASAL2 in highly aggressive pancreatic cancer cell lines and effectively inhibits tumor cell growth in vitro and in vivo (Yin et al. 2019). Similarly, SFN promoted the maturation of miR-29a-3p and inhibited the expression of COL3A1 and COL5A1 in gastric cancer cells and nude BALB/c mice. It leads to the inactivation of the downstream Wnt/β-catenin pathway, thus inhibiting gastric cancer progression (Han et al. 2021). Not only that, Zhang et al. investigated that SFN inhibited the expression of Sirt1 and CD44ICD by inducing the expression of miR-199a-5p, which in turn blocked the invasion of skin squamous carcinoma cells (Zhang et al. 2023). Therefore, developing therapies that target miRNA to inhibit cancer development and progression is an ideal strategy.
Cancer cell migration and invasion are critical reflections of the degree of tumor malignancy, and often distally migrating tumors represent a worse prognosis and a high mortality rate. Invasion and migration of cancer cells are mediated by several signaling molecules. Here, we stress the significance of SFN’s potential benefit in reducing cancer cell invasion and metastasis.
Regulation of tumor cell programmed cell death by SFN
One of the many cell death pathways, programmed cell death (PCD), is one of the key players in many physiological and pathological states. PCD allows damaged, malignant, or no longer needed cells to be lysed and removed to maintain homeostasis within the organism (Chen et al. 2020). When PCD regulation is disturbed, it may lead to cancer, and autoimmune diseases (Fuchs and Steller 2011).
Apoptosis is an essential regulatory death mechanism in the organism by which it maintains homeostasis of its internal environment and removes unwanted, damaged, and infected cells. Most current chemotherapeutic agents induce cancer cell death through apoptosis (Carneiro and El-Deiry 2020). SFN treatment increases the levels of cleaved Caspase-3 and Bax, decreases the levels of Bcl-2, generates ROS to induce apoptosis in glioblastoma cells, and exerts anticancer effects by inhibiting the activation of the STAT3 signaling pathway in cancer cells (Miao et al. 2017). SFN significantly promotes apoptosis in glioblastoma cell lines through a mitochondria-dependent manner, manifested by cysteine asparaginase activation and DNA breaks (Sita et al. 2021). Histone deacetylase (HDAC) is overexpressed in multiple cancer subtypes. SFN affects human telomere reverse transcriptase (hTERT) mRNA levels by regulating colorectal cancer cells histone deacetylase 1 (HDAC1), which reduces hTERT protein expression and enzyme activity and induces apoptosis (Martin et al. 2018). SFN might decrease the growth of all four bladder cancer cell lines, cause apoptosis without producing toxicity, and make tumor cells more sensitive to the chemotherapy drug cisplatin, according to research by Xie et al. (Xie et al. 2022). Survivin is a critical anti-apoptotic and mitotic regulatory protein undetectable in most normal adult tissues but overexpressed in tumors. SFN inhibited prostate cancer cells growth by reducing Survivin protein synthesis by inhibiting the mTOR-S6K1-S6 signaling pathway. At the same time, SFN-induced block in protein synthesis enables cells to maintain ATP at the control cells’ level (Wiczk et al. 2012). Overall, SFN can promote apoptosis of tumor cells in various ways, exerting an inhibitory effect on cancer.
Autophagy is the process by which cells degrade substances, such as proteins, lipids, and organelles in the cytoplasm by capturing them and forming autophagosomes, which fuse with lysosomes. Autophagy is a building block and an essential energy source for cellular self-repair and maintaining homeostasis. Autophagy in cancer provides metabolic substrates for established tumors and promotes their growth (Khayati et al. 2022). It was discovered that SFN therapy boosted the levels of the crucial autophagy regulators HSP90AA1 and UVRAG as well as the lysosome-associated membrane protein 2 (LAMP2) mRNA and protein in human prostate cancer cells (Hahm et al. 2020). Additionally, SFN reduced the proliferation of TNBC cells in vitro and in vivo and triggered autophagy in a dose- and time-varying way, resulting in autophagosome development, up-regulation of Beclin1 expression, and increased synthesis of LC3-II in cancer cells (Yang et al. 2018).
An iron-dependent programmed cell death process known as ferroptosis is connected to growth blockade in many cancer cells (Zhao et al. 2020). It was found that ferroptosis induced by 50 µM SFN in acute myeloid leukemia cells were accompanied by decreased Glutathione Peroxidase 4 (GPX4) expression and increased lipid peroxidation, which provided a new extension of SFN potential as an anticancer agent (Greco et al. 2021). Iida et al. researched explored how SFN caused ferroptosis in small-cell lung cancer cells by upregulating the levels of Fe2+ and lipid peroxidation and downregulating the expression of glutamate anti-transporter protein xCT (SLC7A11) and total glutathione (GSH) (Iida et al. 2021). Meanwhile, SFN also increased the opening of mitochondrial permeability transition pore in gastric cancer cells. It promotes the entry of iron into mitochondria, leading to Fe2+ accumulation, mitochondrial dysfunction, and ferroptosis in gastric cancer cells (Wen et al. 2023).
Overall, PCD is an essential mechanism for maintaining homeostasis in the internal environment of multicellular organisms, and its role in cancer treatment is becoming more and more prominent. SFN has shown its unique characteristics in the multi-pathway regulation of PCD in cancer cells, which makes SFN a possible new strategy to inhibit tumor drug resistance.
Combination of SFN with other anticancer drugs
Combination chemotherapy using two or more drugs with different mechanisms of action reduces drug resistance and normal cytotoxicity and is more effective than monotherapy (Zhang et al. 2015). Combination drugs have the advantage of synergistically affecting multiple survival pathways of tumor cells and can inhibit tumor heterogeneity and drug resistance (Yamada et al. 2016). SFN has anticancer activity and may also enhance the efficacy of other anticancer drugs (Table 2), such as cisplatin, clofarabine, and withaferin A in vitro (Wang et al. 2016; Royston et al. 2017).
In breast cancer cell lines, a two-drug combination of SFN with Biochanin A or Withaferin A induces apoptosis and inhibits cell cycle progression at lower doses (Royston et al. 2018; Li et al. 2023). In addition, the triple-agent combination of SFN, Genistein, and Nodium butyrate also showed a high degree of inhibition of breast cancer progression in vitro. The treatment resulted in a significant decrease cancer cell survival and a significant increase in apoptosis and necrosis rates with the combination treatment compared to therapy alone and the control group (Sharma and Tollefsbol 2022). Compared to control and monotherapy, treating colorectal cell lines with SFN and Salinomycin resulted in a reduction in cell migration and invasion, a boost in the number of apoptotic cells, and inhibited proliferation of colorectal cell lines in vitro and in vivo (Liu et al. 2020). The combination of SFN and the proprietary extract Fernblock® XP (FB) inhibits the production of migratory MMPs in melanoma cells and suppresses melanoma cell growth more effectively than the drug alone (Serini et al. 2020).
The combination of SFN with platinum-based drugs enhances tumor cell killing. By increasing the expression of miR-30a-3p to cause DNA damage and intracellular cisplatin concentration in cisplatin-resistant ovarian cancer cells A2780/CP70 and IGROV1-R10, Gong et al. found that SFN increased the sensitivity of ovarian cancer cells to cisplatin in vitro and in vivo (Gong et al. 2020). Combination treatment of SFN with cisplatin decreases cholangiocarcinoma cells survival, increases the cytotoxicity of cisplatin, and promotes apoptosis in a time-dependent manner (Rackauskas et al. 2017). Multidrug combination of SFN and FOLFOX (5-fluorouracil + oxaliplatin + calcium folinate) promotes the expression of multidrug resistance protein 2 (MRP2) and Bax/Bcl-2 mRNA in colorectal CX-1 cells, which is approximately 2-fold higher than that of the single-agent group (Čižauskaitė et al. 2022).
Moreover, Gasparello et al. developed a “combination therapy” based on SFN and peptide nucleic acid (PNA) targeting miRNAs. SFN and PNA-a15b (targeting miR-15b-5p) collaborative therapy induce a higher level of cellular apoptosis in glioblastoma U251 cells than using the drugs alone (Gasparello et al. 2022b). Similar findings were validated in colorectal cancer, where combination therapy of SFN with PNAs (R8-PNA-a15b, R8-PNA-a425, and R8-PNA-a584) significantly induced more apoptosis in colorectal cancer cells than either compound alone (Gasparello et al. 2020, 2022a). It suggests that combination therapy using PNA and SFN targeting tumor-associated miRNAs is a promising anticancer strategy.
In recent years, the search for natural small-molecule compounds that can effectively fight cancer has shown increasing interest. A promising anticancer method for increasing efficacy and reducing adverse effects is combining various medications with SFN.
Other regulatory of tumors by SFN
-
Regulatory of tumor stem cells by SFN.
Cancer stem cells (CSCs) have the potential for self-renewal and differentiation. They can drive cancer to regenerate repeatedly at primary and metastatic sites, employing various strategies to resist drug therapy and cell death (Batlle and Clevers 2017; Liu et al. 2019). CSCs use many of the same signaling pathways as conventional stem cells, such as the Notch, Hedgehog, and Wnt/β-catenin pathways (Song et al. 2011; Takebe et al. 2011). Since conventional therapies have shown little success in suppressing CSCs (Ehmsen et al. 2019; Molina-Pena et al. 2020; Madsen et al. 2022), finding new drugs to remove CSCs and inhibit their metastasis and drug resistance is necessary.
Several studies in recent years have found that SFN can inhibit the self-renewal of many types of CSCs (Ge et al. 2019; Wang et al. 2021a). Studies in colorectal cancer stem cell and nude mice models have shown that SFN all display a better inhibitory effect. The mechanism may regulate the expression of many stem cell markers by inhibiting ΔNp63α and upregulating ZO-1 (Chen et al. 2022). It’s interesting to note that SFN suppresses tumor stem cells by acting on miRNAs. For instance, SFN reduced the activity of oral squamous cell carcinoma CSCs and lung cancer stem cells by upregulating miR-19 and miR-200 C expression and downregulating GSK3 activation of the Wnt/β-catenin pathway. Reducing the expression of lung CSCs markers and the ability to invade and form colonies inhibited tumor growth in vivo and in vitro (Liu et al. 2017; Zhu et al. 2017). SFN significantly halts tumor progression by interfering with tumor stem cell growth, making it an ideal natural compound for cancer prevention.
-
Regulation of tumor cell energy metabolism by SFN.
A characteristic feature of malignant tumor cells is their reliance on glycolysis for the energy they need to survive and thrive. Tumor cells shift their primary energy source from oxidative phosphorylation to aerobic glycolysis, a shift referred to as the Warburg effect (Schell et al. 2017).
Xia et al. showed that SFN specifically inhibited hypoxia-induced glycolysis by decreasing HIF-1α protein levels and inhibiting nuclear translocation of HIF-1α, resulting in a blocked proliferation of bladder cancer cells in vivo and in vitro (Xia et al. 2019). Another study showed that SFN also inhibited glucokinase 2 (HK2) and pyruvate dehydrogenase (PDH), decreased glycolysis and mitochondrial oxidative phosphorylation, blocked the Akt1/HK2 axis, and reduced aerobic glycolysis with abnormal glucose transport in bladder cancer cells and BBN-induced bladder tumor mouse (Huang et al. 2022). In prostate cancer, SFN has multiple mechanisms to inhibit glycolysis. SFN can inhibit androgen and Tip60-induced glycolysis in prostate cancer cells, increase the activities of HK and Pyruvate kinase (PK), and affect the proliferation and metabolism of tumor cells in vitro (Carrasco-Pozo et al. 2019). By lowering levels of the regulator of fatty acid production SREBP1, SFN can also slow the growth of prostate cancer cells and inhibition of cancer progression in the TRAMP mice model (Singh et al. 2018). Another study conducted in both prostate cancer cells and TRAMP mice have demonstrated that SFN treatment can effectively inhibit the growth of prostate cancer by reducing plasma lactate levels in the prostate and down-regulating the levels of glycolysis-related proteins HK2, pyruvate kinase isozymes M2 (PKM2), and lactate dehydrogenase A (LDHA) (Singh et al. 2019). Thus, SFN plays a significant role in targeting and improving energy metabolism in cancer patients.
The future perspective and challenges for clinical translation of SFN
Improving the stability of SFN in clinical applications
In recent years, the number of publications and patents on SFN inhibits cancer progression has been climbing yearly, indicating that more and more scientists worldwide are beginning to explore this fantastic small molecule compound with the hope of using it in future clinical applications. However, the problem also arises that some contradictory mechanisms inevitably appear in many SFN anticancer mechanisms. Therefore, it is imperative to fully elucidate the molecular targets of SFN and accurately characterize its efficacy. In previous studies of animal tumor models, one of the most common methods of SFN administration is injection, which is usually administered orally in clinical trials. The gap between animal studies and clinical trial scenarios requires more in-depth analyses of the anticancer effects of SFN. These are likewise impediments to the translation of SFN from basic research to clinical application. According to clinicaltrials.gov (http://ClinicalTrials.gov), SFN has conducted several cancer-related clinical trials, such as NCT03517995 (Bladder cancer, Phase 2), NCT04046653 (Prostate cancer, Not Applicable), NCT03232138 (Lung cancer, Phase 2), NCT02404428 (Prostate cancer, Not Applicable), NCT01879878 (Advanced pancreatic cancer, Not Applicable), NCT01568996 (Melanoma, Early Phase), NCT02404428 (Prostate cancer, Not Applicable), NCT01879878 (Advanced pancreatic cancer, Not Applicable), NCT01568996 (Melanoma, Early Phase 1), and so on. These clinical studies show that SFN has a favorable safety and efficacy profile in treating various cancers.
Even though impressive results have been achieved, the relatively low stability of SFN and its sensitivity to water, heat, and alkaline conditions remain significant challenges for clinical translations in the future. It makes the conditions for the production and preservation of SFN more stringent in the pharmaceutical industry or late-stage applications. Also, it hinders the translation of SFN from basic research to clinical applications. Furthermore, the bioavailability decreases after the oral administration of SFN (Soni et al. 2018). To overcome these problems, researchers have focused on improving the stability and efficiency of SFN using micro- and nanotechnology approaches. Nanotherapeutic agents can increase the bioactive concentration and improve bioavailability compared to conventional drugs (Balakumar et al. 2013). Several micro- and nano-encapsulation technologies and wall materials (e.g., hydroxypropyl-β-cyclodextrin and β-cyclodextrin), or micro- and nanocapsule technologies have been used to improve the gastrointestinal stability and bioavailability of SFN (Wu et al. 2014; Simoes et al. 2020; Yepes-Molina and Carvajal 2021). In the results of a study on human breast cancer cells, gold-coated iron oxide nanoparticles significantly improved SFN stability and activity (Manjili et al. 2016). It is also good to develop synthetic analogs of SFN and test their efficacy against cancer. Shi et al. synthesized and evaluated SFN analogs containing heterocyclic molecules. On breast cancer cells, MCF-7, SUM-159, and leukemia stem cell-like cells, KG-1a, analogs 3d, 8d, and 9d considerably outperformed SFN alone regarding their inhibitory actions (Shi et al. 2016). Georgikou et al. evaluated the cytotoxicity of seven chemically synthesized SFN analogs in three pancreatic cancer cell lines. Significant oncogenic effects of the analogs SF102 and SF134 were identified. These therapeutic effects were consistently evaluated in other tumor cell lines, suggesting that SFN analogs have potential applications in cancer therapy (Georgikou et al. 2020).
Determining the optimal dose of SFN in clinical applications
Determining the most appropriate SFN dosage is critical to accurately guiding clinical dosing. Although most plant-derived compounds are generally safe, it is essential to analyze the toxicity of a drug before clinical trials. SFN is currently used in most preclinical studies at concentrations ranging from 0 to 40 µM, and no significant toxicity has been found. The U.S. Food and Drug Administration has limited the use of SFN to a dose of 200 µmoles in some clinical trials, and still only mild side effects such as grade 2 constipation, nausea, headache, and gastrointestinal discomfort have been observed. Still, more extreme doses of SFN have not yet been tested (Alumkal et al. 2015; Tahata et al. 2018; Zhang et al. 2020). The reported cancer progression inhibitory properties of SFN in recent years have far outweighed these mild side effects. Meanwhile, although SFN is significantly cytotoxic to cancer cells, it is relatively safe for normal cells. It has been reported that SFN inhibited the viability of about 75% of cancer cells after treating HepG2 cells with 32 µM SFN for 96 h. No significant toxicity was observed after treating human hepatocytes with 50 µM SFN for 48 h (Gross-Steinmeyer et al. 2010; Dos Santos et al. 2020). Overall, using SFN in a relatively narrow dose range is considered safe while maintaining SFN’s inhibitory effect on cancer cells. In previous studies, SFN was usually administered in a single daily dose. However, SFN is metabolized rapidly in the human body, and its half-life is very short. So far, reports on the dose-response of SFN are still scarce, and the effective dose range of SFN needs to be further determined. More high-quality studies on the dose response of SFN cancer-suppressing administration are required to provide a more accurate basis for developing rational SFN dosage regimens in clinical trials. In addition, researchers need to fill many data gaps, such as dosing frequency, contraindications, and potential adverse side effects. It can provide more detailed and accurate information for the optimal design of clinical trials, which would otherwise require a lot of time and multiple clinical trials to correct these deficiencies.
Conclusions
Cancer has become one of humanity’s most prominent medical problems, with expensive and time-consuming treatments, low survival rates, high side effects, and a considerable burden on low and middle-income populations. Based on these problems, there is an urgent need for a simple, safe, environmentally friendly, and cost-effective novel bioactive substance that exerts maximum therapeutic efficacy while producing minimum side effects. Increasing evidence suggests that SFN exerts anti-proliferative, anti-migratory invasive, pro-apoptotic, inhibitory cell stemness and regulates energy metabolism by targeting multiple signaling pathways, including PI3K, MAPK, Wnt/β-catenin, and NF-κB, among others. Additionally, SFN, a natural dietary supplement, can effectively combine with other anticancer medications. Some preclinical studies have also shown that SFN, in combination with other anticancer drugs, can reduce the side effects of chemotherapy while exerting synergistic anticancer effects. Although the application potential, pharmacokinetic profile, and toxicity characteristics of SFN against different cancers in vivo and in vitro need to be analyzed more comprehensively and accurately, its favorable anticancer activity provides a new direction for the prevention or treatment of various cancers. Hopefully, this review will shed light on the potential value of SFN applications in cancer prevention and treatment and guide future exploration of SFN’s unknown biological functions and possible mechanisms. It would make it easier to get SFN clinically approved to treat cancer, either solely or in cooperation with other chemotherapeutic drugs.
-
Abbreviations:
ITCs | Isothiocyanates |
|---|---|
GLs | Glucosinolates |
SFN | Sulforaphane |
Nrf2 | Nuclear factor erythroid 2-related factor 2 |
GSH | Glutathione |
ARE | Antioxidant response element |
PI3K | Phosphoinositide 3-kinase |
Akt | Protein kinase B |
AMPK | Adenosine 5′-monophosphate-activated protein kinase |
HDAC | Histone deacetylase |
mTOR | Mechanistic target of rapamycin |
S6K | Ribosomal protein S6 kinase |
Bax | Bcl-2 related X protein |
Bcl-2 | B-cell lymphoma-2 |
Her2 | Human epidermal growth factor receptor 2 |
CDK | Cyclin-dependent kinase |
CDC2 | Cell division cycle gene 2 |
CDC25C | Cell division cycle 25 homolog C |
GADD45β | Growth arrest and DNA damage-inducible protein beta |
ROS | Reactive oxygen species |
TFE3 | Transcription factor binding to IGHM enhancer 3 |
VEGF | Vascular endothelial growth factor |
MMP | Matrix metallo proteinase |
AP-1 | Activator protein-1 |
NF-κB | Nuclear factor kappa B |
BBB | Blood-brain barrier |
EMT | Epithelial-mesenchymal transition |
ERK | Extracellular signal-regulated kinase |
E-cadherin | Epithelial cadherin |
N-cadherin | Neural cadherin |
YAP1 | Yes-associated protein 1 |
RASAL2 | RAS protein activator like 2 |
GSK3β | Glycogen synthase kinase3β |
COL3A1 | Collagen type III alpha 1 |
COL5A1 | Collagen type V alpha 1 |
Sirt1 | Silent mating type information regulation 2 homolog- 1 |
PCD | Programmed cell death |
STAT3 | Signal transducer and activator of transcription 3 |
hTERT | Human telomere reverse transcriptase |
LAMP2 | Lysosome-associated membrane protein 2 |
HSP90AA1 | Heat shock protein 90 alpha family class a member 1 |
UVRAG | Ultraviolet radiation resistance-associated gene |
LC3-II | Protein light chain 3 II |
GPX4 | Glutathione peroxidase 4 |
SLC7A11 | Solute carrier family 7 member 11 |
CSCs | Cancer stem cells |
ZO-1 | Zonula occludens-1 |
HIF-1α | Hypoxia inducible factor-1α |
HK | Hexokinase |
PK | Pyruvate kinase |
PDH | Pyruvate dehydrogenase |
LDHA | Lactate dehydrogenase A |
TIP60 | Tat-interacting protein 60 |
SREBP1 | Sterol regulatory element binding protein-1 |
ECGC | Epigallocatechin gallate |
PNA | Peptide nucleic acid |
Data availability
Not applicable.
References
Ahmad A, Nawaz MI. Molecular mechanism of VEGF and its role in pathological angiogenesis. J Cell Biochem. 2022;123(12):1938–65. https://doi.org/10.1002/jcb.30344.
Alumkal JJ, Slottke R, Schwartzman J, Cherala G, Munar M, Graff JN, et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Invest New Drugs. 2015;33(2):480–9. https://doi.org/10.1007/s10637-014-0189-z.
Baig MH, Adil M, Khan R, Dhadi S, Ahmad K, Rabbani G, et al. Enzyme targeting strategies for prevention and treatment of cancer: implications for cancer therapy. Semin Cancer Biol. 2019;56:1–11. https://doi.org/10.1016/j.semcancer.2017.12.003.
Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, et al. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature. 2016;530(7589):184–9. https://doi.org/10.1038/nature16932.
Balakumar K, Raghavan CV, selvan NT, prasad RH, Abdu S. Self nanoemulsifying drug delivery system (SNEDDS) of rosuvastatin calcium: design, formulation, bioavailability and pharmacokinetic evaluation. Colloids Surf B Biointerfaces. 2013;112:337–43. https://doi.org/10.1016/j.colsurfb.2013.08.025.
Batlle E, Clevers H. Cancer stem cells revisited. Nat Med. 2017;23(10):1124–34. https://doi.org/10.1038/nm.4409.
Bijangi-Vishehsaraei K, Reza Saadatzadeh M, Wang H, Nguyen A, Kamocka MM, Cai W, et al. Sulforaphane suppresses the growth of glioblastoma cells, glioblastoma stem cell-like spheroids, and tumor xenografts through multiple cell signaling pathways. J Neurosurg. 2017;127(6):1219–30. https://doi.org/10.3171/2016.8.JNS161197.
Bones AM, Rossiter JT. The enzymic and chemically induced decomposition of glucosinolates. Phytochemistry. 2006;67(11):1053–67. https://doi.org/10.1016/j.phytochem.2006.02.024.
Bose C, Awasthi S, Sharma R, Benes H, Hauer-Jensen M, Boerma M, et al. Sulforaphane potentiates anticancer effects of doxorubicin and attenuates its cardiotoxicity in a breast cancer model. PLoS ONE. 2018;13(3). https://doi.org/10.1371/journal.pone.0193918. e0193918.
Briones-Herrera A, Eugenio-Perez D, Reyes-Ocampo JG, Rivera-Mancia S, Pedraza-Chaverri J. New highlights on the health-improving effects of sulforaphane. Food Funct. 2018;9(5):2589–606. https://doi.org/10.1039/c8fo00018b.
Brooks JD, Paton VG, Vidanes G. Potent induction of phase 2 enzymes in human prostate cells by sulforaphane. Cancer Epidemiol Biomarkers Prev. 2001;10(9):949–54. https://doi.org/10.1023/A:1011247024979.
Carlos-Reyes A, Lopez-Gonzalez JS, Meneses-Flores M, Gallardo-Rincon D, Ruiz-Garcia E, Marchat LA, et al. Dietary compounds as Epigenetic Modulating agents in Cancer. Front Genet. 2019;10:79. https://doi.org/10.3389/fgene.2019.00079.
Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020;17(7):395–417. https://doi.org/10.1038/s41571-020-0341-y.
Carrasco-Pozo C, Tan KN, Rodriguez T, Avery VM. The Molecular effects of Sulforaphane and Capsaicin on Metabolism upon Androgen and Tip60 activation of androgen receptor. Int J Mol Sci. 2019;20(21):5384. https://doi.org/10.3390/ijms20215384.
Chen K, Demarco B, Broz P. Beyond inflammasomes: emerging function of gasdermins during apoptosis and NETosis. EMBO J. 2020;39(2):e103397. https://doi.org/10.15252/embj.2019103397.
Chen Y, Wang M, Wu J, Zhu J, Xie C, Li X, et al. DeltaNp63alpha mediates sulforaphane suppressed colorectal cancer stem cell properties through transcriptional regulation of Nanog/Oct4/Sox2. J Nutr Biochem. 2022;107:109067. https://doi.org/10.1016/j.jnutbio.2022.109067.
Chen X, Adhikary G, Ma E, Newland JJ, Naselsky W, Xu W, et al. Sulforaphane inhibits CD44v6/YAP1/TEAD signaling to suppress the cancer phenotype. Mol Carcinog. 2023;62(2):236–48. https://doi.org/10.1002/mc.23479.
Cheng Y, Tsai C, Hsu Y. Sulforaphane, a Dietary Isothiocyanate, induces G(2)/M arrest in Cervical Cancer cells through CyclinB1 downregulation and GADD45beta/CDC2 Association. Int J Mol Sci. 2016;17(9):1530. https://doi.org/10.3390/ijms17091530.
Čižauskaitė A, Šimčikas D, Schultze D, Kallifatidis G, Bruns H, Čekauskas A, et al. Sulforaphane has an additive anticancer effect to FOLFOX in highly metastatic human colon carcinoma cells. Oncol Rep. 2022;48(5):205. https://doi.org/10.3892/or.2022.8420.
Demaria M, O’Leary MN, Chang J, Shao L, Liu S, Alimirah F, et al. Cellular Senescence promotes adverse effects of Chemotherapy and Cancer Relapse. Cancer Discov. 2017;7(2):165–76. https://doi.org/10.1158/2159-8290.CD-16-0241.
Dos Santos P, Machado ART, De Grandis RA, Ribeiro DL, Tuttis K, Morselli M, et al. Transcriptome and DNA methylation changes modulated by sulforaphane induce cell cycle arrest, apoptosis, DNA damage, and suppression of proliferation in human liver cancer cells. Food Chem Toxicol. 2020;136:111047. https://doi.org/10.1016/j.fct.2019.111047.
Ehmsen S, Pedersen MH, Wang G, Terp MG, Arslanagic A, Hood BL, et al. Increased cholesterol biosynthesis is a key characteristic of breast Cancer stem cells influencing patient outcome. Cell Rep. 2019;27(13):3927–e39383926. https://doi.org/10.1016/j.celrep.2019.05.104.
Fidler MM, Bray F, Soerjomataram I. The global cancer burden and human development: a review. Scand J Public Health. 2018;46(1):27–36. https://doi.org/10.1177/1403494817715400.
Fisher ML, Adhikary G, Grun D, Kaetzel DM, Eckert RL. The Ezh2 polycomb group protein drives an aggressive phenotype in melanoma cancer stem cells and is a target of diet derived sulforaphane. Mol Carcinog. 2016;55(12):2024–36. https://doi.org/10.1002/mc.22448.
Fuchs Y, Steller H. Programmed cell death in animal development and disease. Cell. 2011;147(4):742–58. https://doi.org/10.1016/j.cell.2011.10.033.
Gasparello J, Gambari L, Papi C, Rozzi A, Manicardi A, Corradini R, et al. High levels of apoptosis are Induced in the human Colon cancer HT-29 cell line by Co-administration of Sulforaphane and a peptide nucleic acid targeting miR-15b-5p. Nucleic Acid Ther. 2020;30(3):164–74. https://doi.org/10.1089/nat.2019.0825.
Gasparello J, Papi C, Zurlo M, Gambari L, Manicardi A, Rozzi A, et al. MicroRNAs mir-584-5p and mir-425-3p are Up-Regulated in plasma of Colorectal Cancer (CRC) patients: targeting with inhibitor peptide nucleic acids is Associated with induction of apoptosis in Colon Cancer cell lines. Cancers (Basel). 2022a;15(1):128. https://doi.org/10.3390/cancers15010128.
Gasparello J, Papi C, Zurlo M, Gambari L, Rozzi A, Manicardi A, et al. Treatment of human glioblastoma U251 cells with sulforaphane and a peptide nucleic acid (PNA) targeting miR-15b-5p: synergistic effects on induction of apoptosis. Molecules. 2022b;27(4):1299. https://doi.org/10.3390/molecules27041299.
Ge M, Zhang L, Cao L, Xie C, Li X, Li Y, et al. Sulforaphane inhibits gastric cancer stem cells via suppressing sonic hedgehog pathway. Int J Food Sci Nutr. 2019;70(5):570–8. https://doi.org/10.1080/09637486.2018.1545012.
Georgikou C, Buglioni L, Bremerich M, Roubicek N, Yin L, Gross W, et al. Novel broccoli sulforaphane-based analogues inhibit the progression of Pancreatic Cancer without Side effects. Biomolecules. 2020;10(5):769. https://doi.org/10.3390/biom10050769.
Gong T, Liu X, Zhan Z, Wu Q. Sulforaphane enhances the cisplatin sensitivity through regulating DNA repair and accumulation of intracellular cisplatin in ovarian cancer cells. Exp Cell Res. 2020;393(2):112061. https://doi.org/10.1016/j.yexcr.2020.112061.
Greco G, Schnekenburger M, Catanzaro E, Turrini E, Ferrini F, Sestili P, et al. Discovery of Sulforaphane as an inducer of ferroptosis in U-937 leukemia cells: expanding its Anticancer potential. Cancers (Basel). 2021;14(1):76. https://doi.org/10.3390/cancers14010076.
Gross-Steinmeyer K, Stapleton PL, Tracy JH, Bammler TK, Strom SC, Eaton DL. Sulforaphane- and phenethyl isothiocyanate-induced inhibition of aflatoxin B1-mediated genotoxicity in human hepatocytes: role of GSTM1 genotype and CYP3A4 gene expression. Toxicol Sci. 2010;116(2):422–32. https://doi.org/10.1093/toxsci/kfq135.
Hac A, Brokowska J, Rintz E, Bartkowski M, Wegrzyn G, Herman-Antosiewicz A. Mechanism of selective anticancer activity of isothiocyanates relies on differences in DNA damage repair between cancer and healthy cells. Eur J Nutr. 2020;59(4):1421–32. https://doi.org/10.1007/s00394-019-01995-6.
Hahm ER, Singh KB, Kim SH, Powolny AA, Singh SV. The role of lysosome-associated membrane protein 2 in prostate Cancer Chemopreventive mechanisms of Sulforaphane. Cancer Prev Res (Phila). 2020;13(8):661–72. https://doi.org/10.1158/1940-6207.CAPR-20-0054.
Han F, Li C, Cai Z, Zhang X, Jin G, Zhang W, et al. The critical role of AMPK in driving akt activation under stress, tumorigenesis and drug resistance. Nat Commun. 2018;9(1):4728. https://doi.org/10.1038/s41467-018-07188-9.
Han S, Wang Z, Liu J, Wang H, Yuan Q. miR-29a-3p-dependent COL3A1 and COL5A1 expression reduction assists sulforaphane to inhibit gastric cancer progression. Biochem Pharmacol. 2021;188:114539. https://doi.org/10.1016/j.bcp.2021.114539.
Huang L, He C, Zheng S, Wu C, Ren M, Shan Y. AKT1/HK2 Axis-mediated glucose metabolism: a novel therapeutic target of sulforaphane in bladder Cancer. Mol Nutr Food Res. 2022;66(3). https://doi.org/10.1002/mnfr.202100738. e2100738.
Iida Y, Okamoto-Katsuyama M, Maruoka S, Mizumura K, Shimizu T, Shikano S, et al. Effective ferroptotic small-cell lung cancer cell death from SLC7A11 inhibition by sulforaphane. Oncol Lett. 2021;21(1):71. https://doi.org/10.3892/ol.2020.12332.
Jed W, Fahey, Amy T, et al. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry. 2001;56(1):5–51. https://doi.org/10.1016/s0031-9422(00)00316-2.
Justin S, Rutz J, Maxeiner S, Chun FK, Juengel E, Blaheta RA. Bladder Cancer Metastasis Induced by Chronic Everolimus Application can be counteracted by Sulforaphane in Vitro. Int J Mol Sci. 2020;21(15). https://doi.org/10.3390/ijms21155582.
Kan S, Wang J, Sun G. Sulforaphane regulates apoptosis- and proliferation-related signaling pathways and synergizes with cisplatin to suppress human ovarian cancer. Int J Mol Med. 2018;42(5):2447–58. https://doi.org/10.3892/ijmm.2018.3860.
Kandoth C, McLellan MD, Vandin F, Ye K, Niu B, Lu C, et al. Mutational landscape and significance across 12 major cancer types. Nature. 2013;502(7471):333–9. https://doi.org/10.1038/nature12634.
Khayati K, Bhatt V, Lan T, Alogaili F, Wang W, Lopez E, et al. Transient systemic autophagy inhibition is selectively and irreversibly deleterious to Lung Cancer. Cancer Res. 2022;82(23):4429–43. https://doi.org/10.1158/0008-5472.CAN-22-1039.
Kim D, Sung B, Kang Y, Hwang S, Kim M, Yoon J, et al. Sulforaphane inhibits hypoxia-induced HIF-1alpha and VEGF expression and migration of human colon cancer cells. Int J Oncol. 2015;47(6):2226–32. https://doi.org/10.3892/ijo.2015.3200.
Lam TK, Gallicchio L, Lindsley K, Shiels M, Hammond E, Tao XG, et al. Cruciferous vegetable consumption and lung cancer risk: a systematic review. Cancer Epidemiol Biomarkers Prev. 2009;18(1):184–95. https://doi.org/10.1158/1055-9965.EPI-08-0710.
Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–96. https://doi.org/10.1038/nrm3758.
Langoue TS, Furge LL, Kerriguy N, Nakamura K, Guillouzo A, Guengerich FP. Inhibition of human cytochrome P450 enzymes by 1,2-dithiole-3-thione, oltipraz and its derivatives, and sulforaphane. Chem Res Toxicol. 2000;13(4):245. https://doi.org/10.1021/tx990189w.
Li C, Zhou Y, Peng X, Du L, Tian H, Yang G, et al. Sulforaphane inhibits invasion via activating ERK1/2 signaling in human glioblastoma U87MG and U373MG cells. PLoS ONE. 2014;9(2):e90520. https://doi.org/10.1371/journal.pone.0090520.
Li S, Khoi PN, Yin H, Sah DK, Kim NH, Lian S, et al. Sulforaphane suppresses the Nicotine-Induced expression of the Matrix Metalloproteinase-9 via inhibiting ROS-Mediated AP-1 and NF-κB signaling in human gastric Cancer cells. Int J Mol Sci. 2022;23(9):5172. https://doi.org/10.3390/ijms23095172.
Li J, Xu J, Sun Y, Fu R, Ye D. An insight on synergistic anti-cancer efficacy of biochanin A and sulforaphane combination against breast Cancer. Appl Biochem Biotechnol. 2023. https://doi.org/10.1007/s12010-023-04584-w.
Liebman SE, Le TH. Eat your broccoli: oxidative stress, NRF2, and sulforaphane in chronic kidney disease. Nutrients. 2021;13(1):266. https://doi.org/10.3390/nu13010266.
Liu C, Peng C, Liao Y, Lu M, Tsai M, Yeh J, et al. Sulforaphane targets cancer stemness and tumor initiating properties in oral squamous cell carcinomas via miR-200c induction. J Formos Med Assoc. 2017;116(1):41–8. https://doi.org/10.1016/j.jfma.2016.01.004.
Liu Z, Zhang W, Phillips JB, Arora R, McClellan S, Li J, et al. Immunoregulatory protein B7-H3 regulates cancer stem cell enrichment and drug resistance through MVP-mediated MEK activation. Oncogene. 2019;38(1):88–102. https://doi.org/10.1038/s41388-018-0407-9.
Liu F, Lv R, Liu Y, Hao Q, Liu S, Zheng Y, et al. Salinomycin and Sulforaphane Exerted Synergistic Antiproliferative and Proapoptotic effects on Colorectal Cancer cells by inhibiting the PI3K/Akt signaling pathway in vitro and in vivo. Onco Targets Ther. 2020;13:4957–69. https://doi.org/10.2147/OTT.S246706.
Madsen KL, Gerke O, Hoilund-Carlsen PF, Olsen BB. Cisplatin-resistant CD44(+) Lung Cancer cells are sensitive to Auger electrons. Int J Mol Sci. 2022;23(13). https://doi.org/10.3390/ijms23137131.
Manjili H, Ma’mani K, Tavaddod L, Mashhadikhan S, Shafiee M, A., and, Naderi-Manesh H. D, L-Sulforaphane loaded Fe3O4@ Gold Core Shell nanoparticles: a potential sulforaphane delivery system. PLoS ONE. 2016;11(3). https://doi.org/10.1371/journal.pone.0151344. e0151344.
Martin SL, Kala R, Tollefsbol TO. Mechanisms for the inhibition of Colon cancer cells by sulforaphane through epigenetic modulation of MicroRNA-21 and human telomerase reverse transcriptase (hTERT) down-regulation. Curr Cancer Drug Targets. 2018;18(1):97–106. https://doi.org/10.2174/1568009617666170206104032.
Miao Z, Yu F, Ren Y, Yang J. d,l-Sulforaphane induces ROS-Dependent apoptosis in human gliomablastoma cells by inactivating STAT3 signaling pathway. Int J Mol Sci. 2017;18(1):72. https://doi.org/10.3390/ijms18010072.
Mielczarek L, Krug P, Mazur M, Milczarek M, Chilmonczyk Z, Wiktorska K. In the triple-negative breast cancer MDA-MB-231 cell line, sulforaphane enhances the intracellular accumulation and anticancer action of doxorubicin encapsulated in liposomes. Int J Pharm. 2019;558:311–8. https://doi.org/10.1016/j.ijpharm.2019.01.008.
Molina-Pena R, Tudon-Martinez JC, Aquines-Gutierrez O. A Mathematical Model of Average dynamics in a stem cell Hierarchy suggests the combinatorial targeting of Cancer Stem cells and progenitor cells as a potential strategy against Tumor Growth. Cancers (Basel). 2020;12(9). https://doi.org/10.3390/cancers12092590.
Negrette-Guzman M. Combinations of the antioxidants sulforaphane or curcumin and the conventional antineoplastics cisplatin or doxorubicin as prospects for anticancer chemotherapy. Eur J Pharmacol. 2019;859:172513. https://doi.org/10.1016/j.ejphar.2019.172513.
Obtulowicz T, Winczura A, Speina E, Swoboda M, Janik J, Janowska B, et al. Aberrant repair of etheno-DNA adducts in leukocytes and colon tissue of colon cancer patients. Free Radic Biol Med. 2010;49(6):1064–71. https://doi.org/10.1016/j.freeradbiomed.2010.06.027.
Ohtsuka M, Ling H, Doki Y, Mori M, Calin GA. MicroRNA Processing and Human Cancer. J Clin Med. 2015;4(8):1651–67. https://doi.org/10.3390/jcm4081651.
Peng X, Zhou Y, Tian H, Yang G, Li C. Sulforaphane inhibits invasion by phosphorylating ERK1/2 to regulate E-cadherin and CD44v6 in human prostate cancer DU145 cells. Oncol Rep. 2015. https://doi.org/10.3892/or.2015.4098.
Psyrri A, Bamias A, Yu Z, Weinberger PM, Kassar M, Markakis S, et al. Subcellular localization and protein levels of cyclin-dependent kinase inhibitor p27 independently predict for survival in epithelial ovarian cancer. Clin Cancer Res. 2005;11(23):8384–90. https://doi.org/10.1158/1078-0432.CCR-05-1270.
Rackauskas R, Zhou D, Uselis S, Strupas K, Herr I, Schemmer P. Sulforaphane sensitizes human cholangiocarcinoma to cisplatin via the downregulation of anti-apoptotic proteins. Oncol Rep. 2017;37(6):3660–6. https://doi.org/10.3892/or.2017.5622.
Rafiei H, Ashrafizadeh M, Ahmadi Z. MicroRNAs as novel targets of sulforaphane in cancer therapy: the beginning of a new tale? Phytother Res. 2020;34(4):721–8. https://doi.org/10.1002/ptr.6572.
Rai R, Gong Essel K, Mangiaracina Benbrook D, Garland J, Zhao D, Y., and, Chandra V. Preclinical efficacy and involvement of AKT, mTOR, and ERK Kinases in the mechanism of sulforaphane against Endometrial Cancer. Cancers (Basel). 2020;12(5):1273. https://doi.org/10.3390/cancers12051273.
Royston KJ, Udayakumar N, Lewis K, Tollefsbol TO. A Novel combination of withaferin A and sulforaphane inhibits epigenetic Machinery, Cellular viability and induces apoptosis of breast Cancer cells. Int J Mol Sci. 2017;18(5):1092. https://doi.org/10.3390/ijms18051092.
Royston KJ, Paul B, Nozell S, Rajbhandari R, Tollefsbol TO. Withaferin A and sulforaphane regulate breast cancer cell cycle progression through epigenetic mechanisms. Exp Cell Res. 2018;368(1):67–74. https://doi.org/10.1016/j.yexcr.2018.04.015.
Rutz J, Thaler S, Maxeiner S, Chun FK, Blaheta RA. Sulforaphane reduces prostate Cancer Cell Growth and Proliferation in Vitro by modulating the cdk-cyclin Axis and expression of the CD44 variants 4, 5, and 7. Int J Mol Sci. 2020;21(22):8724. https://doi.org/10.3390/ijms21228724.
Rycaj K, Tang DG. Cell-of-origin of Cancer versus Cancer Stem cells: assays and interpretations. Cancer Res. 2015;75(19):4003–11. https://doi.org/10.1158/0008-5472.CAN-15-0798.
Schell JC, Wisidagama DR, Bensard C, Zhao H, Wei P, Tanner J, et al. Control of intestinal stem cell function and proliferation by mitochondrial pyruvate metabolism. Nat Cell Biol. 2017;19(9):1027–36. https://doi.org/10.1038/ncb3593.
Serini S, Guarino R, Vasconcelos O, Celleno R, L., and, Calviello G. (2020). The Combination of Sulforaphane and Fernblock((R)) XP Improves Individual Beneficial Effects in Normal and Neoplastic Human Skin Cell Lines. Nutrients 12(6), 1608. https://doi.org/10.3390/nu12061608.
Sharma M, Tollefsbol TO. Combinatorial epigenetic mechanisms of sulforaphane, genistein and sodium butyrate in breast cancer inhibition. Exp Cell Res. 2022;416(1):113160. https://doi.org/10.1016/j.yexcr.2022.113160.
Shi Y, Dai D, Li J, Dong Y, Jiang Y, Li H, et al. Sulforaphane analogues with Heterocyclic moieties: syntheses and Inhibitory activities against Cancer Cell lines. Molecules. 2016;21(4):514. https://doi.org/10.3390/molecules21040514.
Simoes BM, Santiago-Gomez A, Chiodo C, Moreira T, Conole D, Lovell S, et al. Targeting STAT3 signaling using stabilised sulforaphane (SFX-01) inhibits endocrine resistant stem-like cells in ER-positive breast cancer. Oncogene. 2020;39(25):4896–908. https://doi.org/10.1038/s41388-020-1335-z.
Singh KB, Kim SH, Hahm ER, Pore SK, Jacobs BL, Singh SV. Prostate cancer chemoprevention by sulforaphane in a preclinical mouse model is associated with inhibition of fatty acid metabolism. Carcinogenesis. 2018;39(6):826–37. https://doi.org/10.1093/carcin/bgy051.
Singh KB, Hahm ER, Alumkal JJ, Foley LM, Hitchens TK, Shiva SS, et al. Reversal of the Warburg phenomenon in chemoprevention of prostate cancer by sulforaphane. Carcinogenesis. 2019;40(12):1545–56. https://doi.org/10.1093/carcin/bgz155.
Sita G, Graziosi A, Hrelia P, Morroni F. Sulforaphane causes cell cycle arrest and apoptosis in human glioblastoma U87MG and U373MG cell lines under hypoxic conditions. Int J Mol Sci. 2021;22(20):11201. https://doi.org/10.3390/ijms222011201.
Song Z, Yue W, Wei B, Wang N, Li T, Guan L, et al. Sonic hedgehog pathway is essential for maintenance of cancer stem-like cells in human gastric cancer. PLoS ONE. 2011;6(3). https://doi.org/10.1371/journal.pone.0017687. e17687.
Soni K, Rizwanullah M, Kohli K. Development and optimization of sulforaphane-loaded nanostructured lipid carriers by the Box-Behnken design for improved oral efficacy against cancer: in vitro, ex vivo and in vivo assessments. Artif Cells Nanomed Biotechnol. 2018;46(sup1):15–31. https://doi.org/10.1080/21691401.2017.1408124.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer statistics 2020: GLOBOCAN estimates of incidence and Mortality Worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49. https://doi.org/10.3322/caac.21660.
Tahata S, Singh SV, Lin Y, Hahm ER, Beumer JH, Christner SM, et al. Evaluation of Biodistribution of sulforaphane after administration of oral broccoli sprout Extract in Melanoma patients with multiple atypical Nevi. Cancer Prev Res (Phila). 2018;11(7):429–38. https://doi.org/10.1158/1940-6207.CAPR-17-0268.
Takebe N, Harris PJ, Warren RQ, Ivy SP. Targeting cancer stem cells by inhibiting wnt, notch, and hedgehog pathways. Nat Rev Clin Oncol. 2011;8(2):97–106. https://doi.org/10.1038/nrclinonc.2010.196.
Wang X, Li Y, Dai Y, Liu Q, Ning S, Liu J, et al. Sulforaphane improves chemotherapy efficacy by targeting cancer stem cell-like properties via the miR-124/IL-6R/STAT3 axis. Sci Rep. 2016;6:36796. https://doi.org/10.1038/srep36796.
Wang D, Zou Y, Zhuang X, Chen S, Lin Y, Li W, et al. Sulforaphane suppresses EMT and metastasis in human lung cancer through miR-616-5p-mediated GSK3β/β-catenin signaling pathways. Acta Pharmacol Sin. 2017;38(2):11. https://doi.org/10.1038/aps.2016.122.
Wang X, Yan Y, Chen X, Zeng S, Qian L, Ren X, et al. The Antitumor activities of Marsdenia tenacissima. Front Oncol. 2018;8:473. https://doi.org/10.3389/fonc.2018.00473.
Wang F, Sun Y, Huang X, Qiao C, Zhang W, Liu P, et al. Sulforaphane inhibits self-renewal of lung cancer stem cells through the modulation of sonic hedgehog signaling pathway and polyhomeotic homolog 3. AMB Express. 2021a;11(1):121. https://doi.org/10.1186/s13568-021-01281-x.
Wang Y, Wu H, Dong N, Su X, Duan M, Wei Y, et al. Sulforaphane induces S-phase arrest and apoptosis via p53-dependent manner in gastric cancer cells. Sci Rep. 2021b;11(1):2504. https://doi.org/10.1038/s41598-021-81815-2.
Wang H, Chen M, Yang C, Hu H, Jiang Y, Yang F, et al. SNRPD1 inhibition suppresses the proliferation of hepatocellular carcinoma and promotes autophagy through the PI3K/AKT/mTOR/4EBP1 pathway. Arch Biochem Biophys. 2023;743:109661. https://doi.org/10.1016/j.abb.2023.109661.
Wen J, Yang F, Fang C, Chen H, Yang L. Sulforaphane triggers iron overload-mediated ferroptosis in gastric carcinoma cells by activating the PI3K/IRP2/DMT1 pathway. Hum Exp Toxicol. 2023;42:9603271231177295. https://doi.org/10.1177/09603271231177295.
Wiczk A, Hofman D, Konopa G, Herman-Antosiewicz A. Sulforaphane, a cruciferous vegetable-derived isothiocyanate, inhibits protein synthesis in human prostate cancer cells. Biochim Biophys Acta. 2012;1823(8):1295–305. https://doi.org/10.1016/j.bbamcr.2012.05.020.
Wu Y, Zou L, Mao J, Huang J, Liu S. Stability and encapsulation efficiency of sulforaphane microencapsulated by spray drying. Carbohydr Polym. 2014;102:497–503. https://doi.org/10.1016/j.carbpol.2013.11.057.
Xia Y, Kang T, Jung Y, Zhang C, Lian S. Sulforaphane inhibits nonmuscle invasive bladder Cancer cells proliferation through suppression of HIF-1alpha-Mediated glycolysis in Hypoxia. J Agric Food Chem. 2019;67(28):7844–54. https://doi.org/10.1021/acs.jafc.9b03027.
Xie H, Rutz J, Maxeiner S, Grein T, Thomas A, Juengel E, et al. Plant-derived sulforaphane suppresses growth and proliferation of drug-sensitive and drug-resistant bladder Cancer Cell Lines in Vitro. Cancers (Basel). 2022;14(19):4682. https://doi.org/10.3390/cancers14194682.
Xu Y, Han X, Li Y, Min H, Zhao X, Zhang Y, et al. Sulforaphane mediates glutathione depletion via Polymeric nanoparticles to restore cisplatin chemosensitivity. ACS Nano. 2019;13(11):13445–55. https://doi.org/10.1021/acsnano.9b07032.
Yamada R, Suda H, Sadanari H, Matsubara K, Tuchida Y, Murayama T. Synergistic effects by combination of ganciclovir and tricin on human cytomegalovirus replication in vitro. Antiviral Res. 2016;125:79–83. https://doi.org/10.1016/j.antiviral.2015.11.008.
Yan Y, Xu Z, Dai S, Qian L, Sun L, Gong Z. Targeting autophagy to sensitive glioma to temozolomide treatment. J Exp Clin Cancer Res. 2016;35:23. https://doi.org/10.1186/s13046-016-0303-5.
Yang F, Wang F, Liu Y, Wang S, Li X, Huang Y, et al. Sulforaphane induces autophagy by inhibition of HDAC6-mediated PTEN activation in triple negative breast cancer cells. Life Sci. 2018;213:149–57. https://doi.org/10.1016/j.lfs.2018.10.034.
Yepes-Molina L, Carvajal M. Nanoencapsulation of sulforaphane in broccoli membrane vesicles and their in vitro antiproliferative activity. Pharm Biol. 2021;59(1):1490–504. https://doi.org/10.1080/13880209.2021.1992450.
Yin L, Xiao X, Georgikou C, Luo Y, Liu L, Gladkich J, et al. Sulforaphane induces miR135b-5p and its target gene, RASAL2, thereby inhibiting the progression of pancreatic Cancer. Mol Ther Oncolytics. 2019;14:74–81. https://doi.org/10.1016/j.omto.2019.03.011.
Zhang J, Desale SS, Bronich TK. Polymer-based vehicles for therapeutic peptide delivery. Therapeutic Delivery. 2015;6(11):1279–96. https://doi.org/10.4155/tde.15.71.
Zhang Z, Li C, Shang L, Zhang Y, Zou R, Zhan Y, et al. Sulforaphane induces apoptosis and inhibits invasion in U251MG glioblastoma cells. Springerplus. 2016;5:235. https://doi.org/10.1186/s40064-016-1910-5.
Zhang Z, Garzotto M, Davis EW 2nd, Mori M, Stoller WA, Farris PE, et al. Sulforaphane Bioavailability and Chemopreventive Activity in men presenting for Biopsy of the prostate gland: a Randomized Controlled Trial. Nutr Cancer. 2020;72(1):74–87. https://doi.org/10.1080/01635581.2019.1619783.
Zhang Y, Gilmour A, Ahn YH, de la Vega L, Dinkova-Kostova AT. The isothiocyanate sulforaphane inhibits mTOR in an NRF2-independent manner. Phytomedicine. 2021;86:153062. https://doi.org/10.1016/j.phymed.2019.153062.
Zhang Y, Ai P, Chen S, Lei S. Sulforaphane suppresses skin squamous cell carcinoma cells proliferation through miR-199a-5p/Sirt1/CD44ICD signaling pathway. Immunopharmacol Immunotoxicol. 2023;45(1):52–60. https://doi.org/10.1080/08923973.2022.2112221.
Zhao Y, Li Y, Zhang R, Wang F, Wang T, Jiao Y. The role of Erastin in Ferroptosis and its prospects in Cancer Therapy. Onco Targets Ther. 2020;13:5429–41. https://doi.org/10.2147/OTT.S254995.
Zheng K, Ma J, Wang Y, He Z, Deng K. Sulforaphane inhibits autophagy and induces exosome-mediated paracrine senescence via regulating mTOR/TFE3. Mol Nutr Food Res. 2020;64(14):e1901231. https://doi.org/10.1002/mnfr.201901231.
Zhou T, Zhou M, Tong C, Zhuo M. Cauliflower bioactive compound sulforaphane inhibits breast cancer development by suppressing NF-κB /MMP-9 signaling pathway expression. Cell Mol Biol (Noisy-le-grand). 2022;68(4):134–43. https://doi.org/10.14715/cmb/2022.68.4.17.
Zhu J, Wang S, Chen Y, Li X, Jiang Y, Yang X, et al. miR-19 targeting of GSK3beta mediates sulforaphane suppression of lung cancer stem cells. J Nutr Biochem. 2017;44:80–91. https://doi.org/10.1016/j.jnutbio.2017.02.020.
Acknowledgements
Not applicable.
Funding
This study was supported by the Key Research and Development Program of Guangdong Province (2022B1111080003), the National Natural Science Foundation of China (22238001, 22078011), Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project (TSBICIP-KJGG-009).
Author information
Authors and Affiliations
Contributions
PL and BZ are responsible for writing the manuscript and drawing the figures. YL revised the manuscript. QY designed the study and revised the manuscript. All authors contributed to the article and approved the submitted version.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, P., Zhang, B., Li, Y. et al. Potential mechanisms of cancer prevention and treatment by sulforaphane, a natural small molecule compound of plant-derived. Mol Med 30, 94 (2024). https://doi.org/10.1186/s10020-024-00842-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10020-024-00842-7