
Abstract
Modulation of host signaling by the products of microbial activity in the gut may affect weight gain and fat formation.
Similar content being viewed by others


Minireview
The relationship between humans and the population of indigenous microorganisms in their intestines (the gut microbiota) is ancient and important. In a recent survey exploring the relationship between mammals and their microbiota it was found that individuals of the same species were more likely to have a similar gut microbiota than mammals of different species [1]. This observation held true regardless of the geographic separation between the two hosts. These results indicate that the composition of the microbiota is dependent more on the identity of the host than on geography and that host and microbiota have co-evolved for their mutual benefit [1]. In essence, we are a mosaic of millions of bacterial genomes that work in concert with the one human genome.
The bulk of our bacterial colleagues are located in the gastrointestinal tract, where the density of bacterial cells in the colon has been estimated at 1011-1012 cells/ml [2]. This close association is mutualistic in nature. The bacteria gain a nutrient-rich environment and humans gain a vast genetic repertoire of encoded physiological functions. Within this repertoire are many genes whose products may help humans adapt to changes in diet and lifestyle. With their short generation times and abilities to swap DNA, the bacteria in our gut adapt and evolve to meet the demands of their ever-changing world, and because their world is our world they serve to complement the human genome. The interactions between host and microbiota determine the success of this relationship. In a recent study published in the Proceedings of the National Academy of Sciences, Samuel and colleagues [3] demonstrate that short-chain fatty acids (SCFAs) produced by the microbiota signal through the host G-protein-coupled receptor (GPCR) Gpr41 and influence weight gain and adiposity.
The microbiota and energy harvesting
We know from studies in germ-free (GF) mouse models that the gut microbiota help to stimulate development of the innate immune system. GF mice also tend to be smaller and do not gain weight like conventionally raised mice. Studies in humans have revealed a shift in the overall community architecture of the gut microbiota in people who lose weight by following either a low-fat or a low-carbohydrate diet. The shift, as hosts lose adiposity, is marked by a reduction in the ratio of Firmicutes to Bacteroidetes [4]. In a mouse model under similar conditions, there are indications that the new microbial composition is less efficient at harvesting energy from nutrients [5]. These studies indicate a role for the gut microbiota in our ability to extract energy from the foods we eat and the ability to maintain a given weight. In fact, the genomes of the microbiota contain many genes related to the breakdown of complex polysaccharides that humans cannot process on their own [6]. The fermentation of carbohydrates by the gut microbiota results in the production of SCFAs.
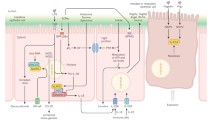
In examining the relationship between SCFA production by the microbiota and host signaling, Samuel and colleagues examined the effects of microbiota-derived SCFAs on the host GPCR Gpr41. Gpr41 is activated by the ligands propionate, butyrate, acetate and pentanoate, especially by the first two. In GF mice, there was no apparent difference in adiposity or weight gain (while on a standard polysaccharide-rich diet) between Gpr41-knockout GF mice and wild-type GF mice. However, when Gpr41-knockout and wild-type GF mice were colonized by the syntrophic partners Bacteroides thetaiotaomicron (Bt) and the archaeon Methanobrevibacter smithii (Ms) (in which one organism lives off the products of the other), Gpr41-knockout mice failed to gain as much weight and adiposity as wild-type mice. This difference in the responses of Bt/Ms-colonized wild-type and Bt/Ms-colonized Gpr41-knockout mice was also observed in conventionally raised mice.
Further analysis indicated that serum levels of the anorexigenic (appetite-suppressing) hormones leptin and peptide YY were lower in GF mice than in Bt/Ms-colonized mice, and lower in Bt/Ms-colonized Gpr41-knockout mice than in Bt/Ms-colonized wild-type mice. Leptin is derived from adipose tissue and is involved in regulating many different responses, including metabolic rate and eating behavior. Peptide YY, among other activities, inhibits gut motility. As they predicted, Samuel et al. [3] demonstrated enhanced gut motility in Bt/Ms-colonized Gpr41-knockout mice compared with Bt/Ms-colonized wild-type mice. Thus, one mechanism by which Gpr41 and the gut microbiota appear to mediate weight gain is to decrease food transit time in the small intestine and thus increase time for absorption of SCFAs. Interestingly, peptide YY levels were higher in colonized Gpr41-knockout mice than in GF Gpr41-knockout mice, suggesting that the gut microbiota may also induce peptide YY expression via a mechanism independent of Gpr41.
Ever since the discovery of leptin and its effects on obesity, attempts to develop drugs that target its function have failed. GPCRs are an important class of drug targets; approximately 30% of pharmaceuticals in current use target this family of receptors [7]. The results reported by Samuel et al. [3] indicate that Gpr41 might be an attractive drug target for countering obesity. However, the desired mechanisms of such drugs are unclear, as increases in peptide YY levels also increase satiety, and have been linked to decreases in human obesity [8]. A more complete understanding of the downstream signaling and physiology controlled by Gpr41 will be a prerequisite for such a drug-targeting strategy.
Microbial modulation of host signaling
In a broader context, modulation of host signaling pathways is a common mechanism utilized by bacterial and viral pathogens. Manipulation of host signaling machinery often serves to promote the pathogen's own agenda. For example, Salmonella species utilize a guanine-exchange factor to induce membrane ruffling in host cells and promote their own uptake, and then deploy a GTPase-activating protein to downregulate membrane ruffling once they are inside the cell. Other pathogens modulate host signaling to prevent uptake by host cells, to block immune responses, or to direct cellular machinery for other specific purposes to the benefit of the pathogen [9]. Studies of bacterial and viral pathogens have taught us fundamental lessons about the regulation of signaling and normal physiology in eukaryotic cells. For example, studies of the Rous sarcoma virus protein Src led to the discovery of phosphorylation as a means of regulating signaling [10]. In a recent study, the Vibrio parahaemolyticus type III effector protein VopS was found to inhibit host-cell Rho GTPases by covalently attaching AMP to them [11]. This study of a bacterial pathogen provided the first evidence for the role of 'AMPylation' in the regulation of eukaryotic signaling.
Not all host-microbe conversations are private. Redundancy and use of shared or common language are important features of the signaling interactions between commensal microorganisms and their animal hosts. Toll-like receptors (TLRs) and Nod receptors provide major 'trunk lines' through which both pathogens and commensals interact with the host. TLRs are sentinels of the immune system, sensing the presence of many different types of microbial products. Bacterial lipopolysaccharide (LPS) and peptidoglycan are potent stimulators of the TLR system. Commensal LPS and peptidoglycan help maintain homeostasis in the gut epithelium and protect the gut mucosa from injury by stimulating the production of the protective molecules IL-6, TGF-β, KC-1 and heat-shock proteins [12]. Both pathogens and commensals often target elements of the host innate immune system in order to subvert host defenses. Commensals have been shown to induce expression of an antimicrobial protein, angiogenin-4, in order perhaps to reshape innate immunity in the gut [13]. In fact, Gpr43, another receptor for SCFAs, is known to be highly expressed on polymorpho-nuclear leukocytes, and SCFAs are known to attract and activate these cells [14]. The Gpr-microbiota signaling story may be relevant to various inflammatory diseases of the gut; strategies to interfere with Gpr signaling might prove useful for treating these disorders (as well as obesity).
Monotypic associations of a microbial symbiont with its host sometimes provide more dramatic examples of the effect of microbial signaling on the host. In the squid/Vibrio fischeri symbiosis, V. fischeri resides in the light organ of the host and provides luminescence. V. fischeri stimulates the expression of two squid genes: a putative LPS-binding protein and a receptor for peptidoglycan. Both of these proteins are required for the development of the squid's light organ [15]. In this example, symbiont signaling fundamentally alters the physiology of the host and induces the formation of an environment conducive to the symbiont.
The elegant work of Samuel et al. [3] illuminates one pathway through which the microbiota and host communicate. However, the complex mix of SCFAs and other by-products of bacterial metabolism in the gut, the diversity of associated potential host receptors, and the variation in expression of both along the length of the gut and among different types of host cells predict multiple levels of host-microbiota regulation and response. In a recent study of obese and lean twins, metagenomic analysis revealed the presence of a core microbiome, defined by a set of 'functional' microbial genes [16]. Further analysis of these data will undoubtedly lead to an array of new potential signaling factors. With everyone talking at once, the biggest challenge for us will be to learn how to listen.
References
Ley RE, Hamady M, Lozupone C, Turnbaugh PJ, Ramey RR, Bircher JS, Schlegel ML, Tucker TA, Schrenzel MD, Knight R, Gordon JI: Evolution of mammals and their gut microbes. Science. 2008, 320: 1647-1651. 10.1126/science.1155725.
Ley RE, Peterson DA, Gordon JI: Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell. 2006, 124: 837-848. 10.1016/j.cell.2006.02.017.
Samuel BS, Shaito A, Motoike T, Rey FE, Backhed F, Manchester JK, Hammer RE, Williams SC, Crowley J, Yanagisawa M, Gordon JI: Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proc Natl Acad Sci USA. 2008, 105: 16767-16772. 10.1073/pnas.0808567105.
Ley RE, Turnbaugh PJ, Klein S, Gordon JI: Microbial ecology: human gut microbes associated with obesity. Nature. 2006, 444: 1022-1023. 10.1038/4441022a.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI: An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006, 444: 1027-1031. 10.1038/nature05414.
Hooper LV, Midtvedt T, Gordon JI: How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu Rev Nutr. 2002, 22: 283-307. 10.1146/annurev.nutr.22.011602.092259.
Drews J: Drug discovery: a historical perspective. Science. 2000, 287: 1960-1964. 10.1126/science.287.5460.1960.
Gardiner JV, Jayasena CN, Bloom SR: Gut hormones: a weight off your mind. J Neuroendocrinol. 2008, 20: 834-841. 10.1111/j.1365-2826.2008.01729.x.
Patel JC, Galan JE: Manipulation of the host actin cytoskeleton by Salmonella - all in the name of entry. Curr Opin Microbiol. 2005, 8: 10-15. 10.1016/j.mib.2004.09.001.
Butel JS: Viral carcinogenesis: revelation of molecular mechanisms and etiology of human disease. Carcinogenesis. 2000, 21: 405-426. 10.1093/carcin/21.3.405.
Yarbrough M, Li Y, Kinch LN, Grishin NV, Ball HL, Orth K: AMPylation of Rho GTPases by Vibrio VopS disrupts effector binding and downstream signaling. Science. 2009, 323: 269-272. 10.1126/science.1166382.
Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S, Medzhitov R: Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell. 2004, 118: 229-241. 10.1016/j.cell.2004.07.002.
Hooper LV, Stappenbeck TS, Hong CV, Gordon JI: Angiogenins: a new class of microbicidal proteins involved in innate immunity. Nat Immunol. 2003, 4: 269-273. 10.1038/ni888.
Le Poul E, Loison C, Struyf S, Springael JY, Lannoy V, Decobecq ME, Brezillon S, Dupriez V, Vassart G, Van Damme J, Parmentier M, Detheux M: Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J Biol Chem. 2003, 278: 25481-25489. 10.1074/jbc.M301403200.
Chun CK, Troll JV, Koroleva I, Brown B, Manzella L, Snir E, Almabrazi H, Scheetz TE, Bonaldo Mde F, Casavant TL, Soares MB, Ruby EG, McFall-Ngai MJ: Effects of colonization, luminescence, and autoinducer on host transcription during development of the squid-vibrio association. Proc Natl Acad Sci USA. 2008, 105: 11323-11328. 10.1073/pnas.0802369105.
Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, Sogin ML, Jones WJ, Roe BA, Affourtit JP, Egholm M, Henrissat B, Heath AC, Knight R, Gordon JI: A core gut microbiome in obese and lean twins. Nature. 2008
Acknowledgements
JED is supported by NIH Postdoctoral Training Grant 2 T32 AI007328-21; DAR is supported by an NIH Director's Pioneer Award and a Doris Duke Charitable Trust Distinguished Clinical Scientist Award.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dinalo, J.E., Relman, D.A. Cross-talk in the gut. Genome Biol 10, 203 (2009). https://doi.org/10.1186/gb-2009-10-1-203
Published:
DOI: https://doi.org/10.1186/gb-2009-10-1-203