
Abstract
Background
Many widely distributed coral reef fishes exhibit cryptic lineages across their distribution. Previous study revealed a cryptic lineage of Pomacentrus coelestis mainly distributed in the area of Micronesia. Herein, we attempted to use molecular and morphological approaches to descript a new species of Pomacentrus.
Results
The morphological comparisons have been conducted between cryptic species and P. coelestis. Pomacentrus micronesicus sp. nov. is characterized by 13 to 16 (typically 15) anal fin rays (vs. 13 to 15, typically 14 rays in P. coelestis) and 15 or 16 rakers on the lower limb of the first gill arch (vs. 13 or 14 rakers in P. coelestis). Divergence in cytochrome oxidase subunit I sequences of 4.3% is also indicative of species-level separation of P. micronesicus and P. coelestis.
Conclusions
P. micronesicus sp. nov. is described from the Marshall Islands, Micronesia on the basis of 21 specimens. Both morphological and genetic evidences support its distinction as a separate species from P. coelestis.
Similar content being viewed by others


Background
Marine organisms that are morphologically undistinguishable but genetically distinct are known as ‘cryptic species’ (Knowlton 2000). In the past decade, DNA sequencing has provided an independent means of testing the validity of existing taxonomic units, revealing cases of inappropriate synonymy and, consequently, the existence of numerous cryptic species. For instance, Steinke et al. (2009) examined 391 species of ornamental fishes and, based on mitochondrial (mt)DNA genetic divergence, concluded that three of 44 pomacentrids examined represented previously overlooked species, suggesting that cryptic speciation might be common in this family of marine fishes.
Pomacentrus coelestis (Jordan and Starks, 1901), also known as the neon damselfish, was described on the basis of a single specimen collected at Wakanoura in Kii, Japan and is widely distributed in the Indo-Pacific region (Allen 1991). Liu et al. (2012) conducted a phylogeographic study of P. coelestis across its distribution using both the mtDNA control region and microsatellite loci as genetic markers and revealed two deeply divergent clades; the first clade was shown to encompass the ‘true’ P. coelestis haplotype, which is widespread in the West Pacific, Indonesia, and the South Pacific, while the second clade included a cryptic species, distributed in a relatively narrow area between Micronesia and West Papua. This cryptic species could not be distinguished from P. coelestis in situ based on its color pattern and body shape (GR Allen, personal communication). In addition, several morphologically similar species, including Pomacentrus auriventris (Allen, 1991), Pomacentrus similis (Allen, 1991), Pomacentrus alleni (Burgess, 1981), and Pomacentrus caeruleus (Quoy and Gaimard, 1825) were suggested to compose a ‘P. coelestis complex’ (GR Allen, personal communication). The goals of the present study were to compare morphological measurements among species within this complex and discuss the phylogenetic relationship between the true P. coelestis and other congeners.
Methods
Methods for taking morphometric and meristic data followed that of Allen (2004). Proportional measurements are expressed as a percentage of the standard length (SL) or head length (HL), as shown in Table 1. Counts and proportions for the holotype are followed by value ranges for the paratypes (in parentheses). Type specimens were deposited at the National Museum of Marine Biology and Aquarium (NMMB-P, Pingtung, Taiwan), the National Museum of Natural History (Smithsonian Institution, USNM, Washington, DC, USA), and the Western Australian Museum (WAM, Perth, Australia). Institution codes followed those of Fricke and Eschmeyer (2012). Specimens for phylogenetic analyses were collected either from the field with barrier and hand nets or purchased from a local aquarium shop. Ten specimens of the new species used for sequencing were collected from Kwajalein Atoll, Marshall Islands, and P. coelestis was collected from northern Taiwan. Two specimens of P. alleni and P. auriventris were purchased from a local aquarium shop.
DNA was isolated with the Genomic DNA extraction kit (Genomics BioScience and Technology, New Taipei City, Taiwan) from muscle tissue preserved in 95% ethanol according to the manufacturer’s recommendations. A partial fragment of the mtDNA cytochrome oxidase subunit I (COI) gene was amplified by a polymerase chain reaction (PCR) using Taq DNA polymerase (MDbio, Taipei, Taiwan) and FishF2 and FishR2 primers (Ward et al. 2005). Each 25-μl reaction contained 10 to 50 ng DNA, 10 mM Tris HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 1 U Taq DNA polymerase, 0.2 mM dNTPs, and 0.3 mM of each primer. The mixture was amplified with a cycling profile of 2 min at 94°C for the first cycle, followed by 34 cycles at 95°C for 30 s, 54°C for 30 s, and 70°C for 40 s.
Nucleotide sequences of the PCR products were determined using an ABI 377 automated sequencer. Sequences obtained in this study were submitted to the NCBI GenBank database. Sequences were aligned using CLUSTAL W (Thompson et al. 1994) followed by manual editing using Sequencher 4.2 (Gene Code, Ann Arbor, MI, USA). A Kimura 2-parameter (K2P) distance metric was employed for sequence comparisons between samples, as this distance is commonly used to represent genetic divergence between fish species (Hubert et al. 2008; Steinke et al. 2009). Then, the best-fit substitution model, K2P plus gamma (γ = 0.0619), was selected by MEGA5 (Tamura et al. 2011) based on the Bayesian information criterion. This model was used to construct a maximum likelihood (ML) tree with additional sequences downloaded from GenBank (P. auriventris, GenBank:HLC-12192-6 and P. alleni, GenBank:JQ418308-11). Bootstrap values for the ML trees were estimated using 1,000 replicates.
Additionally, Bayesian phylogenetic reconstructions were estimated with MrBayes 3.12 (Ronquist and Huelsenbeck 2003) using the K2P model and default priors. Subsequently, two duplicate runs of three heated and one cold Markov chain Monte Carlo chains were established, and each was initiated from a random tree and run for 106 generations. A convergence diagnostic was applied, and the stop probability was set to 0.01. Then, trees were sampled every 100 generations, and a consensus tree was built on all trees with the exclusion of the first 25% of sampled trees to allow for sufficient burn-in. Posterior probabilities were determined by constructing a 50% majority-rule tree of all sampled trees. The accession numbers were GenBank:JX232402-3 for P. auriventris, GenBank:JX232380-1 for P. alleni, GenBank:JX232382-91 for P. coelestis, and GenBank:JX232392-401 for the new species. The study complies with the current laws of the countries in which it was performed, and fishes used in this study have been treated based on the ethical justification for the use and treatment of fishes by the Journal of Fish Biology, 2006 (http://www.blackwell-synergy.com).
Results
Pomacentrus micronesicus sp. nov. Liu, Ho and DaiCommon name: Micronesian demselfish(Figures 1, 2, 3, 4 and Tables 1, 2, 3)

Photographs of the P. coelestis complex. (A) P. alleni, Weh Island, Sumatra, Indonesia. (B) P. coelestis, Nusa Kode, Komodo Islands, Indonesia. (C) P. auriventris, Raja Ampat Islands, Indonesia. (D) P. similis, Mergui Archipelago, Myanmar. (E) P. caeruleus, Andavadoaka, southwestern Madagascar. (F) P. micronesicus sp. nov., Chuuk, Federated States of Micronesia. Courtesy of GR Allen.

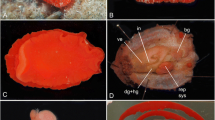
P. micronesicus sp. nov. (A) NMMB-P15602, holotype, 58.3 mm standard length (SL), right lateral view. (B) WAM P.33715-001, 55.4 mm SL, left lateral view. (C) X-ray film showing the anal fin elements of the holotype. Spines are shown in Roman numerals, and soft rays are indicated by a white dot at each base.

Underwater photograph of adult P. micronesicus sp. nov. (70 mm total length). Near Kwajalein Atoll, Marshall Islands at 5 to 8 m in depth. Courtesy of Jeanette Johnson.

Maximum likelihood phylogenic tree of the four morphologically similar species of the P. coelestis complex. P. coelestis, P. auriventris, P. micronesicus sp. nov., and P. alleni, based on mtDNA COI sequences. Numbers below the nodes represent support based on 1,000 bootstraps, and numbers above the nodes represent clade credibility values from the Bayesian analysis.
Holotype
NMMB-P15602 (58.3 mm), Kwajalein Atoll, Marshall Islands (9°11′2.87″N, 167°25′4.88″E), 5 to 10 m, hand net, 2 August 2009, collected by S Johnson.
Paratypes
CAS 235015–6 (formerly NMMB-P15605-6, 37.8 to 51.7 mm), NMMB-P15591-601, 15603–4, 15607 (14: 37.8 to 58.4 mm), USNM 410087 (2: 46.8 and 57.0 mm), WAM P.33715-001 (2: 49.8 and 55.4 mm), all collected with the holotype by S Johnson.
Comparative materials
P. alleni
WAM P.26506-009 (paratypes, 4: 39.5 to 44.1 mm), 8°40′N, 97°38′E, Thailand, Similan Island, Ko Bangu, Andaman Sea, 12 February 1979, collected by GR Allen.
P. auriventris
WAM P.29708-005 (8: 30.7 to 45.6 mm), 8°30′S, 122°20′E, Pulau Besar, Flores, Indonesia, 8 to 12 m, 15 April 1988, collected by GR Allen.
P. caeruleus
WAM P.27329-001 (4: 39.6 to 51.2 mm), 5°25′S, 71°44′E, Peros Banhos, Chagos Archipelago, 18 to 24 m, March 1979, collected by R Winterbottom et al., unpublished work, (3: 40.1 to 44.3 mm), Madagascar.
P. coelestis
NMMB-P15608-15 (8: 44.7 to 59.6 mm), 25°00′45″N, 121°59′23″E, off Mao-ao, Ilan, northeastern Taiwan, 5 to 10 m, July 2009, collected by S-Y Liu. NMMB-P15616 (4: 29.1 to 37.3 mm), Tanegashima, Japan, western Pacific Ocean, 5 to 10 m, July 2002, collected by T. Kokita, 30.43′N, 130.59′E. WAM P.31418-001 (4: 23.4 to 36.6 mm), 5°10′18″S, 145°51′24″E, southern tip of Massas Island, Papua New Guinea, 18.3 m, 6 November 1978, collected by V Springer et al. WAM P.28035-014 (5: 24.4 to 51.5 mm), 17°6′S, 119°38′E, outer reef northeast side near shipwreck, Mermaid Reef, Rowley Shoals, northwest Australia, 10 to 12 m, 13 August 1983, collected by GR Allen. WAM P.27453-003 (2: 47.6 and 49.6 mm), 30°2′S, 142°2′E, reef on west side above drop-off, 500 m offshore, One Tree Island, Queensland, northeast Australia, 8 to 13 m, 7 December 1966, collected by V Springer et al. WAM P.25129-003 (2: 35.9 and 47.4 mm), 17°40′S, 168°25′E, Efate, off Erakor Island, New Hebrides, Vanuatu, 13 m, 23 June 1973, collected by GR Allen and D Popper. WAM P.31065-010 (23: 20.5 to 34.4 mm), 1°52′S, 136°33′E, south coast of Yapen Island near Warironi Village, West Papua, Indonesia, 2 to 3 m, 9 July 1995.
P. similis
WAM P.25525-001 (2: 43.2 and 52.7 mm), 6°11′N, 81°28′E, Great Basses Reef, Sri Lanka, 2 to 10 m, 15 November 1975, collected by GR Allen.
Diagnosis
Dorsal fin rays XIII, 13 to 15 (typically XIII, 14); anal fin rays II, 13 to 16 (typically II, 15); pectoral fin rays 16 to 18 (typically 17 or 18); tubed lateral line scales 16 to 18 (typically 17 or 18); gill rakers 5 to 7 + 15 or 16 = 20 to 23 (typically 21); body depth 2.5 to 2.8 in proportion to SL in specimens longer than 30 mm SL; caudal fin deeply forked with a relatively long filamentous upper lobe.
Description
Morphometric and metric data are provided in Table 1. The following data are provided for the holotype, and the corresponding measurements for the paratypes are given in parentheses.
Dorsal fin rays XIII, 14 (XIII, 13 to 15); anal fin rays II, 14 (II, 13 to 16); all dorsal and anal fin soft rays branched, the last to the base; 2 uppermost and lowermost pectoral fin rays unbranched; pelvic fin rays I, 5; principal caudal fin rays 15, median 13 with branched upper and lower procurrent caudal fin rays, posterior 2 segmented; scales in longitudinal series 26 (25 or 26); tubed lateral line scales 18 (17 or 18); posterior midlateral scales with a pore or deep pit 9 (8 to 10) in continuous series; scale rows below origin of the 1st dorsal fin 2.5; scale rows above lateral line to the base of the middle dorsal spine 1.5; scale rows below lateral line to the origin of anal fin 9; gill rakers 6 + 15 (5 to 7 + 15 or 16); total gill rakers 21 (20 to 23, mainly 21).
Body ovate, slightly elongated, depth 2.7 (2.5 to 2.8) in SL and compressed, width 2.5 (1.9 to 2.7) in body depth; head length 3.5 (3.3 to 3.6) in SL; dorsal and ventral profiles of the head evenly rounded from dorsal and pelvic fin origins to snout; snout shorter than orbit, length 4.4 (3.7 to 4.8) in HL; caudal peduncle depth 2.2 (2.0 to 2.3) in HL; caudal peduncle length 2.1 (1.9 to 2.4) in HL.
Mouth terminal, small and oblique, forming an angle of about 40° to 60° to horizontal axis of the head and body; maxilla not reaching vertical at the anterior margin of orbit, upper jaw length 3.4 (3.2 to 3.6) in HL; teeth biserial, at least at the front of jaws, consisting of the outer row of larger incisiform to conical teeth and slender buttress teeth behind and in the spaces between the outer row teeth, 38 (35 to 40) in the outer row of upper jaw and 36 (34 to 39) in the outer row of lower jaw. Tongue triangular with rounded tip, papillose on dorsal surface. Gill rakers long and slender, the longest on the lower limb near angle about equal to or slightly shorter than the length of the longest gill filaments.
Nostrils 2, anterior nostril rounded with slightly raised rim, close to the upper lip; posterior nostril rounded without rim, directly anterior to the middle of the eye and about midway between the anterior edge of the eye and upper lip. Opercle ending posteriorly in a flat spine, tip acute but short, barely projecting from beneath membrane; preopercle serrations well developed, 22 on the left side and 18 on the right side of holotype (14 to 26); preorbital with a small spine posteriorly, separated by a notch (poorly developed in specimens of < 50 mm SL) from suborbital series; lower edge of suborbital serrate in specimens of > 50 mm SL and smooth in others.
Scales finely ctenoid; head scaled except the lips; tip of the snout, preorbital and suborbital; a scaly sheath at the base of dorsal and anal fins, averaging about one half the pupil width at the base of spinous portion of dorsal fin and slightly less at the base of anal fin; a column of scales on each membrane of dorsal and anal fins, narrowing distally, those on the spinous portion of dorsal fin progressively longer, reaching about two third distance to spine tips at the end of spinous dorsal fin and the first one half of soft dorsal fin then gradually shorter on the remainder of fin; small scales on caudal fin extending about two third to three fourth distance to posterior margin; small scales on the basal one fourth of pectoral fins; a cluster of several scales forming the median process, extending posteriorly from between the base of pelvic fins, its length slightly more than one half that of the pelvic spine, the last scale enlarged and triangular; axillary scale above the base of pelvic spine about one half length of the pelvic fin.
Origin of dorsal fin over the second or third lateral line scale; predorsal distance 3.3 (3.0 to 3.3) in SL; preanal distance 1.6 (1.5 to 1.7) in SL; prepelvic distance 2.7 (2.2 to 2.8) in SL; the base of the soft portion of dorsal fin containing about 2.4 times the base of spinous portion; dorsal fin spines gradually increasing in length to the last spine; first dorsal spine 4.0 (3.5 to 5.3) in HL; seventh dorsal spine 2.4 (1.9 to 2.7) in HL; last dorsal spine 1.9 (1.6 to 2.1) in HL; membranes of spinous portion of dorsal fin incised; fifth, sixth, or seventh dorsal soft ray longest, 1.2 (1.1 to 1.6) in HL; first anal spine 3.3 (3.3 to 4.9) in HL; second anal spine 2.2 (1.8 to 2.3) in HL; longest (11th) anal soft ray 1.3 (1.1 to 1.6) in HL; fourth pectoral ray longest, 1.1 (1.0 to 1.2) in HL; pelvic fin spine 2.1 (1.7 to 2.1) in HL; pelvic fin length 1.2 (0.9 to 1.1) in HL; caudal fin deeply forked, both lobes filamentous, upper lobe slightly longer than lower lobe, upper lobe length 0.6 (0.6 to 1.0) in HL; caudal concavity very deep, 1.3 (1.3 to 2.9) in HL.
Etymology
This species was named after the predominant area (Micronesia) where it was found.
Coloration in alcohol
The color was uniformly brownish to creamy blue (Figure 2A,B).
Coloration when alive (from underwater photographs)
Holotype is metallic blue anteriorly and dorsally with a light yellow ventral portion. The dorsal fin is metallic blue, and the soft ray of pectoral fin is light blue. The pectoral fin is transparent to light blue, and the pelvic fin is light blue to yellow with a blue anterior margin. The anal fin is light blue to bright yellow with a blue margin and many small blue spots. Finally, the caudal fin is light blue to yellowish, with a few blue spots on the anterior portion (Figure 2).
Distribution
Liu et al. (2012) collected samples of P. coelestis across its distribution and found two deeply divergent groups, the Pacific (true P. coelestis) and Micronesian (P. micronesicus sp. nov.) clades, and also suggested that the haplotype of P. micronesicus sp. nov. is mainly distributed in Micronesia and is also found in Indonesia at West Papua (Raja Ampat Islands) and Bali, which indicates that it may have a distribution from Palau to the Marshall Island and south to Bali, inhabiting depths of 5 to 20 m.
Comparison
P. micronesicus sp. nov. can be distinguished from its sibling species, P. coelestis, in typically having 15 rays on the anal fin (vs. 14 rays in P. coelestis, Figure 2C), 15 or 16 rakers on the lower limb of the first gill arch (vs. 13 or 14 rakers), more total gill rakers on the first gill arch (20 to 23 vs. 18 to 21), and a lower body depth (Table 1). Moreover, the caudal fin usually possesses a filamentous upper lobe that is deeply concave in P. micronesicus sp. nov., but that was not frequently observed in P. coelestis. On the other hand, P. auriventris has II, 14 anal fin rays, 13 gill rakers on the lower limb, and 18 or 19 total gill rakers on the first gill arch. P. alleni has II, 14 anal fin rays, 12 or 13 gill rakers in the upper limb, and 17 or 18 total gill rakers in the first gill arch (Table 2).
Remarks
It is notable that 23 specimens from one jar of P. coelestis (WAM P.31065-010) collected on the southern coast of Yapen Island, West Papua, Indonesia had mostly II, 15 anal fin rays instead of II, 14 in specimens collected from other localities. The majority of the anal fin ray count was similar to that of P. micronesicus sp. nov. However, Yapen (northern Cenderawasih Bay) specimens shared a similar number of gill rakers as P. coelestis (i.e., mainly 13 or 14 in the lower limb and 19 in total) and differed from the type series of P. micronesicus sp. nov. (typically with 15 in the lower limb and 21 in total). Cenderawasih Bay was suggested as a place where an extraordinary amount of endemism has arisen due to its complex geological history (Allen and Erdmann 2012). In this study, those specimens are temporary identified as P. coelestis. However, a more thorough investigation of this population may show it to represent a different species.
Phylogenetic analyses
A 597-bp fragment of the mtDNA COI gene was amplified from 24 samples of 4 morphologically similar species within the P. coelestis complex and compared to 9 additional sequences downloaded from the NCBI GenBank database (5 sequences from P. auriventris and 4 from P. alleni). Among these sequences, 60 variable sites were found. Nucleotide frequencies were 23.06% for A, 28.46% for T, 29.65% for C, and 18.82% for G. Two independently generated trees (ML and Bayesian inference) revealed similar tree topologies. Three well-supported major clades were observed (Figure 4), one including P. coelestis and P. auriventris, one including P. mironesicus sp. nov., and the final including P. alleni. In addition, the average K2P genetic distance between the four species was 3.8%. The K2P genetic distance between the two most morphologically similar species, P. mironesicus sp. nov. and P. coelestis, was 4.3%, which was higher than the average divergence between species within the complex (Table 3).
Discussion
Many coral reef fishes possess vivid coloration, and this trait is very important to reef fish identification, especially among closely related species (Randall 1998; Lieske and Meyers 2001). Within the P. coelestis species complex, only two species with diagnostic color patterns can be differentiated from P. coelestis: P. alleni, which has a black stripe along the basal region of the caudal fin, and P. auriventris, which possesses yellow coloration over the lower one half of its body. Based on their identical color patterns as observed in situ, it was expected that P. coelestis and P. mironesicus sp. nov. could be a recently derived species pair. However, results of the molecular phylogenetic analyses showed that P. auriventris is more closely related to P. coelestis (Figure 3). Moreover, as P. alleni is found only in the eastern Indian Ocean, it could represent the ancestral clade of the phylogenetic tree, suggesting that this species complex might have originated in the Indian Ocean. However, a more complete phylogeny and analyses are needed to support this hypothesis.
Recently, mtDNA has frequently been used as genetic markers to support the divergence of species pairs in the Pomacentridae (Allen et al. 2010; Allen and Drew 2012). The average genetic distance of COI between P. micronesicus sp. nov. and three other closely related species was 5.7%, which is on average higher than that reported for many recognized species pairs (Rocha and Bowen 2008; Steinke et al. 2009). In addition to these gene-based findings, several diagnostic morphological traits were found to distinguish P. micronesicus sp. nov. from P. coelestis. First, P. micronesicus sp. nov. typically possesses 15 anal fin rays versus only 14 in the latter species. Second, P. micronesicus sp. nov. possesses 15 or 16 gill rakers on the lower limb of the first gill arch versus only 13 or 14 in the latter species. Therefore, both morphological and genetic data support the conclusion that P. mironesicus sp. nov. is a valid species.
Conclusions
P. micronesicus sp. nov. is described from the Marshall Islands, Micronesia on the basis of 21 specimens. It is characterized by 13 to 16 (typically 15) anal fin rays (vs. 13 to 15, typically 14 rays in P. coelestis) and 15 or 16 rakers on the lower limb of the first gill arch (vs. 13 or 14 rakers in P. coelestis). Divergence in cytochrome oxidase subunit I sequences of 4.3% is also indicative of species-level separation of P. micronesicus and P. coelestis. Both morphological and genetic evidences support its distinction as a separate species from P. coelestis.
References
Allen GR: Damselfishes of the world. Melle: Mergus Publishers; 1991.
Allen GR: Pomacentrus aurifrons , a new species of damselfish (Pomacentridae) from Melanesia. Zootaxa 2004, 399: 1–7.
Allen GR, Drew J: A new species of damselfish ( Pomacentrus : Pomacentridae) from Fiji and Tonga. Aqua Int J Ichthyol 2012, 18: 171–180.
Allen GR, Erdmann MV: Reef fishes of the East Indies. Perth: Tropical Reef Research; 2012.
Allen GR, Drew J, Fenner D: Amphiprion pacificus , a new species of anemonefish (Pomacentridae) from Fiji, Tonga, Samoa, and Wallis Island. Aqua Int J Ichthyol 2010, 16: 129–138.
Fricke R, Eschmeyer WN: A guide to fish collections in the catalog of fishes database. 2012. . Accessed 20 Apr 2012 http://researcharchive.calacademy.org/research/ichthyology/catalog/collections.asp
Hubert N, Hanner R, Holm E, Mandrak NE, Taylor E, Burridge M, Watkinson D, Dumont P, Curry A, Bentzen P, Zhang J, April J, Bernatchez L: Identifying Canadian freshwater fishes through DNA barcodes. PLoS One 2008, 3: e2490. 10.1371/journal.pone.0002490
Knowlton N: Molecular genetic analyses of species boundaries in the sea. Hydrobiologia 2000, 420: 73–90. 10.1023/A:1003933603879
Lieske E, Meyers R: Coral reef fishes: Indo-Pacific and Caribbean. Princeton: Princeton University Press; 2001.
Liu S-YV, Dai C-F, Allen GR, Erdmann MV: Phylogeography of the neon damselfish ( Pomacentrus coelestis ) indicates a cryptic species and different species origin in the West Pacific Ocean. Mar Ecol Prog Ser 2012, 458: 155–167.
Randall JE: Zoogeography of shore fishes of the Indo-Pacific region. Zool Stud 1998, 37: 227–268.
Rocha LA, Bowen BW: Speciation in coral reef fishes. J Fish Biol 2008, 72: 1101–1121. 10.1111/j.1095-8649.2007.01770.x
Ronquist F, Huelsenbeck JP: MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19: 1572–1574. 10.1093/bioinformatics/btg180
Steinke D, Zemlak TS, Hebert PDN: Barcoding Nemo: DNA-based identifications for the ornamental fish trade. PLoS One 2009, 4: e6300. 10.1371/journal.pone.0006300
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S: MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 2011, 28: 2731–2739. 10.1093/molbev/msr121
Thompson JD, Higgins DG, Gibson TJ: CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucl Acid Res 1994, 22: 4673–4680. 10.1093/nar/22.22.4673
Ward RD, Zemlak TS, Innes BH, Last PR, Hebert PDN: DNA barcoding Australia’s fish species. Philos Trans Royal Soc B 2005, 360: 1847–1857. 10.1098/rstb.2005.1716
Acknowledgments
We would like to thank Dr. G. Allen for his support by collecting fish, sharing information, and providing underwater photographs. We specially thank S. Morrison (WAM), S. Smith (USNM), Dr. L.A. Rocha (CAS), and S.-I. Wang (NMMB-P) for the curatorial assistance. We are also grateful to Mr. Scott Johnson and Mrs. Jeanette Johnson for their assistance with fish collection and photography at Kwajalein Atoll, Marshall Islands. Finally, we would like to thank Dr. Anderson Mayfield for his assistance with the English proofreading and two anonymous referees for the critical comments. This study was supported by a grant (NSC99-2811-M-002-151) from the National Science Council of Taiwan to CFD.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
S-YVL conceived and designed the experiments. S-YVL and H-CHH performed the experiments, analyzed the data, and wrote the paper. C-FD made intellectual contributions and financial support. All authors read and approved the final manuscript.
Shang-Yin Vanson Liu, Hsuan-Ching Hans Ho contributed equally to this work.
An erratum to this article is available at http://dx.doi.org/10.1186/s40555-014-0046-y.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Liu, SY.V., Ho, HC.H. & Dai, CF. A new species of Pomacentrus (Actinopterygii: Pomacentridae) from Micronesia, with comments on its phylogenetic relationships. Zool. Stud. 52, 6 (2013). https://doi.org/10.1186/1810-522X-52-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1810-522X-52-6