
Abstract
Background
The putative H. pylori pathogenicity-associated factor dupA has been associated with IL-8 induction in vitro, and duodenal ulcer (DU) and gastric cancer (GC) development in certain populations, but this association is inconsistent between studies. We aimed to investigate dupA prevalence in clinical isolates from Sweden, Australia and from ethnic Chinese, Indians and Malays resident in Malaysia and Singapore and to examine the association with DU and GC. In addition we investigated the sequence diversity between isolates from these diverse groups and compared the level of IL-8 secretion in isolates possessing and lacking dupA.
Methods
PCR primers were designed to amplify over the C/T insertion denoting a continuous dupA. PCR products from 29 clinical isolates were sequenced and compared with sequences from three additional strains obtained from GenBank. Clinical isolates from 21 Malaysian patients (8 dupA-positive, 14 dupA-negative) were assessed for their ability to induce IL-8 in AGS cells in vitro. Statistical analysis was performed using Fisher's exact test.
Results
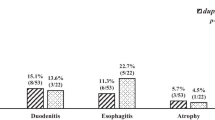
The prevalence of dupA in isolates from Swedish functional dyspepsia (FD) control patients (65%, 13/20) was higher and in isolates from Indian FD patients (7.1%, 3/42) was lower as compared with isolates from Chinese (28.9%, 13/49, P = 0.005, P = 0.025), Malay (35.7%, 5/14, P = 0.16, P = 0.018) and Australian (37.8%, 17/45, P = 0.060, P < 0.001) FD patients. dupA was associated with DU and GC development in Chinese with 62.5% (10/16) and 54.6% (12/22) of isolates possessing dupA respectively as compared with FD controls (28.9%) (P = 0.015, P = 0.032). No significant difference in prevalence of dupA between FD controls, DU (63.6%, 7/11) and GC (61.9%, 13/21) cases (P = 1.000) was observed in the Swedish population. Sequence analysis revealed a pairwise variation of 1.9% and all isolates possessed the C/T insertion. The average IL-8 induction was 1330 pg/mL for dupA-positive isolates and 1378 pg/mL for dupA-negative isolates.
Conclusion
Although dupA is highly conserved when present, we identified no consistent association between dupA and DU or GC development across the ethnic groups investigated, with the dupA prevalence in control groups varying significantly. Our results would suggest that in the clinical isolates investigated dupA is not associated with IL-8 induction in vitro.
Similar content being viewed by others


Introduction
Although Helicobacter pylori infection invariably results in gastritis, a significant minority of those infected will progress to more severe gastroduodenal pathologies, including Duodenal Ulcer (DU) and Gastric Cancer (GC). The factors resulting in progression to severe H. pylori- related disease are poorly understood, however there is compelling evidence to suggest a co-contribution of bacterial virulence factors, host genetics and environmental stimuli [1].
To date a number of H. pylori pathogenicity-associated factors, including flagella, adhesins, urease, the vacuolating cytotoxin and the cag pathogenicity island (cag PAI) have been associated with the development of more serious H pylori-related disease outcomes. The cag PAI encodes a type IV secretion system (TFSS), homologous to the well-studied virB/D4 TFSS of the plant pathogen Agrobacterium tumefaciens, as well as the effector protein CagA [2, 3]. Translocation of CagA by the TFSS into gastric epithelial cells has been shown to drive deregulation of intracellular signaling pathways resulting in a myriad of cellular effects such as increased motility and elongation [4]. In a process independent of CagA, the TFSS also induces the production of the chemo-attractant and pro-inflammatory cytokine IL-8 by facilitating the entry of peptidoglycan into the host epithelial cell [5]. In vitro studies have demonstrated that the virB4 homologous ATPase, CagE (HP0544, JHP0492), is essential for IL-8 induction [6–10]. In addition to cagE, multiple virB4 homologues exist within the H. pylori genome, with both sequenced strains J99 and 26695 possessing three additional virB4 homologues [11, 12]. Recently Lu et al. demonstrated that the partial virB4 homologues JHP0917 and JHP0918, which are present in the plasticity zone of J99 but not 26695, form one continuous gene in clinical isolates the duodenal ulcer promoting gene (dupA). and that this gene was a functional virB4 homologous ATPase [13]. dupA, like cagE, is associated with IL-8 induction [13] and thus it is possible that this may be part of an as yet unidentified TFSS. In the original study examining the role of dupA in more severe H. pylori related disease, dupA possessing isolates were found to be associated with the development of DU in patients from Columbia, Japan and Korea [13]. In contrast no association was found with gastric cancer suggesting that dupA, and the TFSS it is associated with, may be a major determinant in DU-development. In contrast, two recent studies, one in Brazil [14] and a second in which subjects from Belgium, China, South Africa and the USA were examined [15] did not find an association between dupA and DU development. Given that H. pylori pathogenicity-associated factors in isolates from different populations have been shown to possess distinct genotypes [16–18], it is important that the association between a newly identified putative pathogenicity-associated factor, such as dupA and specific disease outcomes, be assessed in clinical isolates from a range of different regions and ethnic groups [19].
In the present study we determined the prevalence of dupA across a range of countries and ethnic groups including Australia, Sweden and the three primary ethnic groups resident within Malaysia, namely Chinese, Indians and Malays and examined the relationship between dupA and DU and GC in the Swedish and ethnic Chinese populations. We also sequenced dupA PCR products from isolates from each ethnic and disease group and determined that they possessed the C/T insertion required for a continuous dupA gene, and additionally that there was a only a small degree of sequence diversity between isolates. Finally, we determined that the induction of IL-8 secretion by AGS cells was not associated with the presence of dupA in the clinical isolates investigated in vitro.
Methods
Patients, H. pylori strains and extraction of genomic DNA
Malaysian & Singaporean
As part of a larger study examining the role of bacterial, host and environmental factors in GC development, gastric biopsies were obtained from consecutively enrolled unrelated patients undergoing routine endoscopic examination of gastrointestinal symptoms at the University Hospital, Kuala Lumpur, Malaysia and the Changi Hospital, Singapore (between mid-2004 and 2007). Based on endoscopic and histological examination, patients were diagnosed with either distal GC or functional dyspepsia (FD). Gastric biopsies were stored at -70°C in brain heart infusion (BHI) broth containing 20% glycerol and transported to Australia on dry ice. Gastric biopsies were smeared onto campylobacter selective medium (CSA), incubated and identified using morphology, microscopy and biochemical tests as previously described [20, 21]. Genomic DNA from one single colony isolate per patient was extracted using the Gentra Puregene DNA extraction kit (Gentra, Minneapolis, Minnesota, USA). In the present study, 142 single colony isolates from 52 Chinese, 42 Indian and 14 Malay FD patients, 22 Chinese GC patients, and 16 Chinese DU patients were included.
Swedish
In a previous case-control study which recruited patients from eight hospitals in Sweden biopsies were collected, cultured for H. pylori and bacterial DNA extracted as previously described [22]. In the present study 52 single colony isolates from Swedish patients diagnosed with FD (20), DU (11) and GC (21) were included.
Australian
In a previous study which recruited patients from Sydney, Australia, biopsies were collected, cultured for H. pylori and bacterial DNA extracted as previously described [23]. In the present study, 45 single colony isolates from Caucasian Australian patients with FD were included.
Primer design and detection of dupA in single colony isolates
Primer pair dupA1274F and dupA1674R (table 1) were designed to amplify a 399–400 bp fragment corresponding to nucleotides 1016315–1016714 in J99 (accession no. AE001439). The PCR primers amplified over the region of dupA containing the 1 bp insertion/deletion detected by Lu et al. 2005 [13], that is the forward primer bound to a sequence from JHP0917 and the reverse primer bound to a sequence from JHP0918, suggesting that PCR positive isolates have both JHP0917 and JHP0918. PCR was performed in a 25 μL reaction containing 0.825 U Taq DNA polymerase (Fisher Biotech, Subiaco, Australia), 45 nmol magnesium-chloride, 10 pmol of each primer and 10 ng of genomic DNA. All PCR runs included a negative control (no DNA) and a positive control (J99). PCR was performed as follows: initial denaturation for 5 minutes at 95°C, followed by 35 cycles of 95°C for 20 seconds, 55°C for 20 seconds and 72°C for 40 seconds, and a final at 72°C for 7 minutes. PCR products were visualized by agarose gel electrophoresis.
Validation of primers and investigation of dupA by DNA sequencing
29 single colony isolates (4 Chinese, 2 Indian, 4 Malay, 3 Australian and 3 Swedish FD isolates, 4 Chinese and 3 Swedish DU isolates, and 3 Chinese and 3 Swedish GC isolates) were sequenced to validate the specificity of the primers dupA F and dupA R and to ascertain whether JHP0917-0918 formed a single continuous gene in these isolates. Sequencing was performed using the BigDye™ Terminator version 3.1 (Applied Biosystems, Foster City, California, USA). Sequencing analysis was performed on an ABI3730 Capillary DNA sequencer (GENterprise, Mainz, Germany). Sequences were verified using Blast [24] available from Biomanager by ANGIS http://www.angis.org.au. Sequence alignments of the 29 strains, with the original dupA gene sequence from Gene Bank (accession no. AB196363) [13] and two additional dupA- like sequences (accession no. EF076755 and EF076756) available in the GenBank database http://www.ncbi.nlm.nih.gov/Genbank/index.html were created using ClustalW accurate [25] also available from BioManager.
Development of a dupA multiplex PCR
To confirm that isolates negative for the dupA PCR truly lacked the dupA gene, rather than being the result of sequence diversity over the primer annealing site, a multiplex PCR was designed involving the use of 2 forward (dupA 113F and dupA 1274F) and 3 reverse (dupA 1083R, dupA 1674R and dupA 1830R) primers which produce 5 differently sized PCR products if all primers bind (Figure 1). Primer sequences are presented in table 1.

PCR amplification using the dupA multiplex PCR. (A)Genomic DNA from H. pylori strains J99 (positive control), 26695 (negative control) and FD527 (clinical isolate) were used to PCR amplify dupA using the forward primers dupA113 F and dupA 1274F, and the reverse primers dupA 1083R, dupA 1674R and dupA 1830R. M, size markers (in base pairs). (B) Schematic representation of dupA showing the annealing positions of the forward primers dupA113 F and dupA 1274F, and the reverse primers dupA 1083R, dupA 1674R and dupA 1830R, and the expected sizes of the amplified PCR products.
The PCR was performed in a 25 uL reaction as described above, with an initial denaturation for 5 minutes at 95°C, followed by 35 cycles of 95°C for 30 seconds, 50°C for 30 seconds and 65°C for 2 minutes, and a final at 65°C for 7 minutes.
Infection assay and IL-8 induction in vitro
AGS cells (ATCC CRL-1739) were routinely maintained in RMPI1640 supplemented with 10% FCS and glutamine (All from Gibco Invitrogen, Paisley, UK), and were cultured at 37°C in 5%CO2/95% air. Cells were seeded in 24-well tissue culture plates and infected with approximately 3 × 107 bacterial cells. 22 H. pylori isolates from Malaysian patients (8 dupA- positive and 14 dupA-negative) were available for in vitro analysis. Two H. pylori control strains (67:20 (cag PAI negative) and 67:21 (cag PAI positive) [26] were also available for in vitro analysis. H. pylori isolates were cultured overnight in brucella broth (Sigma) supplemented with 5% FCS (Gibco Invitrogen). Bacteria were added to AGS cells in duplicate, and were co-incubated for 6 hours before cell culture supernatant was collected and stored at -20°C prior to use. At least three independent experiments were performed; results represent the average for these experiments. The concentration of IL-8 in the supernatant was analysed by ELISA, using the BD OptEIA: Human IL-8 ELISA set (BD, San Diego, CA, USA) according to the manufacturers instructions.
Statistical Analysis
Fisher's exact test was used to calculate statistical significance; two-sided P values < 0.05 were considered significant.
Sequences have been deposited in GenBank under the following accession numbers: EU253504–EU253532.
Results
Prevalence of dupA
Following PCR analysis, dupA was identified in 15/52 (28.9%) Chinese, 5/14 (35.7%) Malay, 3/42 (7.1%) Indian, 17/45 (37.8%) Australian and 13/20 (65.0%) Swedish FD clinical isolates (table 2). The prevalence of dupA in Swedish FD isolates was higher (P = 0.005, P = 0.162, P = 0.025) and the prevalence in Indian FD isolates was lower (P = 0.025, P = 0.0517, P = 0.094) as compared with the Chinese, Malay and Australian FD isolates investigated. There was no significant difference in the prevalence of dupA between isolates from Chinese, Malay and Australian FD patients (P = 0.052 to P = 1.000). Among isolates from Chinese or Swedish DU patients there was no significant difference between the two ethnicities, with 10/16 (62.5%) and 7/11 (63.6%) of the isolates possessing dupA respectively (P = 1.000). Similarly, there was no significant difference between isolates from Chinese and Swedish GC patients with 12/22 (54.6%) and 13/21 (61.9%) isolates possessing dupA respectively (P = 0.759). Among isolates from Chinese patients, the prevalence of isolates possessing dupA was significantly higher in isolates from DU and GC patients than in isolates from FD control patients (P = 0.015, P = 0.032), suggesting an association with the development of disease in this population. There was no significant difference in dupA prevalence between the isolates from Chinese DU and Chinese GC patients (P = 0.744). Conversely, there was no significant difference in the prevalence of dupA between isolates from Swedish FD, DU and GC patients (P = 1.000).
Sequence Analysis
Blast analysis of sequences from 29 clinical isolates revealed significant sequence similarity to the dupA gene sequence deposited in GenBank, accession no. AB196363[13] and the sequence of a dupA-like protein with GenBank accession no. EF076755. Of the 4 Chinese, 2 Indian, 4 Malay FD, 3 Swedish FD and 3 Australian FD isolates, 4 Chinese and 3 Swedish DU isolates, and 3 Chinese and 3 Swedish GC isolates, all isolates were observed to possess a 1 bp C or T insertion after position 1385 relative to gene JHP0917 in J99 suggesting the presence of 1 continuous open-reading frame.
Sequence alignments from the 29 clinical isolates with 3 comparable sequences deposited in GenBank (AB196363, EF076755, EF076756) revealed that dupA possessed a pair-wise sequence variation of only 1.90/100 bp (maximum variation: 14.71/100 bp, minimum variation 0.46/100 bp, median variation 0.92/100 bp, standard deviation 3.16/100 bp). Two isolates examined (the Chinese FD isolate FD553 (GenBank sequence: EU253520) and Iranian isolate EF076756), demonstrated a sequence variations of 14.71/100 bp and 12.64/100 bp respectively, being more than 2 standard variations above the mean. In the case of FD553 the high degree of sequence variation was the direct result of a 60 bp insertion at base pair 1465 relative to AB196363. The remaining isolates were within 1 standard deviation of the mean.
Multiplex PCR
The use of the multiplex PCR revealed no additional isolates positive for dupA, when compared to the use of dupA1274F and dupA1674R alone, suggesting that the use of the single primer pair is adequate to determine the prevalence of dupA in both western and east Asian populations. An example of the multiplex PCR is presented in figure 1; all five bands are clearly distinguishable in clinical isolate FD527.
In vitro IL-8 production
The uninoculated control secreted 13 pg/ml, with the negative control strain (67:20) secreting 80 pg/ml and the positive control strain (67:21) secreting 1368 pg/ml (figure 2A). Individual IL-8 secretion levels varied between 126 pg/ml and 1855 pg/ml for dupA-positive isolates, and 123 pg/ml and 1633 pg/ml for dupA-negative isolates. However, no significant difference (p > 0.5) was observed in the average level of IL-8 secreted by AGS cells between clinical isolates possessing dupA (1330 pg/mL) and those lacking dupA (1378 pg/mL) (figure 2B).

IL-8 induction in AGS cells by clinical isolates in vitro. (A) Induction of IL-8 secretion in AGS cells incubated with indicated strains for 6 hours. The error bars show the standard deviations of each independent experiment. (B) Average induction of IL-8 secretion in AGS cells incubated with dupA positive (blue) or negative (red) strains. The error bars show the standard deviations for each group of strains.
Discussion
The current investigation of the prevalence of dupA in clinical isolates from Australia and Sweden as well as from the three major ethnic groups (Chinese, Indians and Malays) resident in Malaysia and Singapore clearly shows that there is significant variability in the prevalence of dupA between not only geographical locations, but also between ethnic groups resident in the same country. In the present study the prevalence of dupA in H. pylori isolates collected from FD patients varied significantly between nationalities/ethnicities, with isolates from Swedish patients being significantly more likely to possess dupA (65.0%) than isolates from Australia (26.3%) or ethnic Chinese (28.9%), Indians (7.1%) or Malays (35.7%) resident in Malaysia or Singapore. Interestingly isolates from ethnic Indian FD patients were significantly less likely to possess dupA (7.1%) than isolates from any of the other groups. This difference in dupA prevalence between countries and ethnic groups is supported by several studies [15, 27] including the initial Lu et al. 2005 [13] which identified that dupA was more prevalent in isolates from Columbian gastritis patients (39%) than in isolates from Japanese (14%) and Koreans (7%) gastritis patients. Furthermore, there is no significant difference in dupA prevalence reported previously for isolates from Chinese patients (25% and 35.3% respectively) [15, 28] and for isolates from ethnic Chinese Malaysians and Singaporeans reported in the present study (28.9%), despite different primer pairs being used, suggesting that the primer pair designed in this study is appropriate. Conversely, the prevalence reported in a north Indian population was considerably higher than that found in the ethnic Indian Malaysians in the present study (16/70 as compared with 3/42) [29]. One possible explanation is that the Indian Malaysians are likely to be predominantly ethnic Tamil [29], originating from Southern India, rather than the north Indian Indeed, two studies in Brazil reported significantly different prevalences for dupA (92.3% vs. 62%) [14, 30].
We also demonstrated that the association between dupA and severe gastroduodenal disease was inconsistent between the Chinese and Swedish populations. In isolates from ethnic Chinese patients the prevalence of dupA was significantly higher in patients diagnosed with DU (62.5%) or GC (54.6%) as compared with those diagnosed with FD. This is in contrast to the observations of Lu et al. 2005 [13] and Zhang et al. 2008 [28] who reported a negative association with GC, but supports the observations by Argent et al. [15]. In contrast, in the Swedish population there was no significant difference in the prevalence of dupA in isolates from patients diagnosed with DU, GC or FD, similar to findings reported from Brazil [14, 30]. Our observations are consistent with the reported variation in other H. pylori virulence factors such as the cagA and vacA genes [31], and further emphasise the fact that before a new virulence factor is associated with a specific disease outcome, studies must be undertaken in a range of geographic locations as well in different ethnic groups.
Lu et al. have reported that, with the exception of J99, JHP0917–JHP0918 forms a continuous open reading frame, dupA, due to the presence of a 1 bp C/T insertion in clinical isolates [13]. In the present study, all clinical isolates sequenced possessed a continuous dupA gene, signified by the presence of the C/T insertion, a finding that is consistent with previous reports in other populations [13, 14, 32]. Our results also indicate that dupA is highly conserved over the region sequenced with an average partial sequence variation of only 1.9%. This indicates a high degree of conservation comparable to housekeeping genes such as atpA, ureI, efp, and ppa [33]. We must acknowledge that as only 29 clinical isolates were sequenced and the sequences used for comparison represent less than 400 bp, the pairwise sequence variation for the entire gene and in a larger population may be higher than that described here. Interestingly in a recent study Douraghi et al. compared 3 regions of the dupA gene – JHP0917 (289 bp), JHP0918 (259 bp) and the junction region over JHP0917 and JHP0918 i.e. 'dupA' (216 bp) in 6 Iranian strains with that of 10 Brazilian and 3 Indian strains whose sequences had been deposited in GenBank [32]. The reported sequence similarities in that study ranged from 86.1%–100% for JHP0917, 88–98.8% for JHP0918 and 93.4–99.5% for 'dupA, a finding that would support the high degree of conservation of this gene observed in our study. Surprisingly in the Douraghi et al. study none of the strains were compared for all 3 regions.
The high degree of sequence conservation reported and the finding that dupA is present in strains from different continents and in populations with different genetic backgrounds, suggests that dupA may confer a fitness advantage of some sort, which has resulted in the conservation of the gene and the sequence, throughout human migration. An alternate explanation, since dupA is located in the plasticity zone, is that the acquisition was a relatively recent phenomenon. Although there is, as yet, no specific evidence for either scenario, we believe the former explanation is more likely. Whole gene sequence analysis of dupA over an increased sample size is necessary to adequately investigate these points.
Although the initial study by Lu et al reported that dupA was associated with IL-8 induction in vitro in both knockout studies and in clinical isolates [13], in the present study we observed no significant difference in the level of IL-8 induction between strains with or without dupA. It is possible however that the strains used in the present study lacked the other essential components of the dupA type IV secretion system, however, as none of these components have yet been identified it is impossible to assess whether this is responsible for the lack of association with IL-8 induction in the present study. More comprehensive studies with strains lacking genes known to contribute to IL-8 induction such as cag PAI components and with oipA off would be beneficial in elucidating the contribution of dupA to IL-8 induction.
Conclusion
Although based on our findings and those of others, it is unlikely that dupA is itself a determinant or indicator of a specific clinical outcome across all populations; it may be shown to be an important virulence factor if a role as part of a novel TFSS is identified.
References
Pritchard DM, Crabtree JE: Helicobacter pylori and gastric cancer. Curr Opin Gastroenterol. 2006, 22: 620-625. 10.1097/01.mog.0000245539.50765.f6.
Odenbreit S, Puls J, Sedlmaier B, Gerland E, Fischer W, Haas R: Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science. 2000, 287: 1497-1500. 10.1126/science.287.5457.1497.
Backert S, Ziska E, Brinkmann V, Zimny-Arndt U, Fauconnier A, Jungblut PR, Naumann M, Meyer TF: Translocation of the Helicobacter pylori CagA protein in gastric epithelial cells by a type IV secretion apparatus. Cell Microbiol. 2000, 2: 155-164. 10.1046/j.1462-5822.2000.00043.x.
Backert S, Selbach M: Role of type IV secretion in Helicobacter pylori pathogenesis. Cell Microbiol. 2008, 10: 1573-1581. 10.1111/j.1462-5822.2008.01156.x.
Viala J, Chaput C, Boneca IG, Cardona A, Girardin SE, Moran AP, Athman R, Mémet S, Huerre MR, Coyle AJ, et al: Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat Immunol. 2004, 5: 1166-1174. 10.1038/ni1131.
Maeda S, Ogura K, Yoshida H, Kanai F, Ikenoue T, Kato N, Shiratori Y, Omata M: Major virulence factors, VacA and CagA, are commonly positive in Helicobacter pylori isolates in Japan. Gut. 1998, 42: 338-343.
Owen RJ, Sharp S, Lawson AJ, Durrani Z, Rijpkema S, Kidd M: Investigation of the biological relevance of Helicbacter pylori cagE locus diversity, presence of CagA motifs and vacuolating cytotoxin genotype on IL-8 induction in gastric epithelial cells. FEMS Immunol Med Microbiol. 2003, 36: 135-140. 10.1016/S0928-8244(03)00022-1.
Tummuru MK, Sharma SA, Blaser MJ: Helicobacter pylori picB, a homologue of the Bordetella pertussis toxin secretion protein, is required for induction of IL-8 in gastric epithelial cells. Mol Microbiol. 1995, 18: 867-876. 10.1111/j.1365-2958.1995.18050867.x.
Day AS, Jones NL, Lynett JT, Jennings HA, Fallone CA, Beech R, Sherman PM: cagE is a virulence factor associated with Helicobacter pylori-induced duodenal ulceration in children. J Infect Dis. 2000, 181: 1370-1375. 10.1086/315394.
Censini S, Lange C, Xiang Z, Crabtree JE, Ghiara P, Borodovsky M, Rappuoli R, Covacci A: cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease associated virulence factors. Proc Natl Acad Sci USA. 1996, 93: 14648-14653. 10.1073/pnas.93.25.14648.
Alm RA, Ling LS, Moir DT, King BL, Brown ED, Doig PC, Smith DR, Noonan B, Guild BC, deJonge BL, et al: Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature. 1999, 397: 176-180. 10.1038/16495.
Tomb J-F, White O, Kerlavage AR, Clayton RA, Sutton GG, Fleischmann RD, Ketchum KA, Klenk HP, Gill S, Dougherty BA, et al: The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature. 1997, 388: 539-10.1038/41483.
Lu H, Hsu P-I, Graham D, Yamaoka Y: Duodenal ulcer promoting gene of Helicobacter pylori. Gastroenterol. 2005, 128: 833-848. 10.1053/j.gastro.2005.01.009.
Gomes LI, Rocha GA, Rocha AM, Soares TF, Oliveira CA, Bittencourt PF, Queiroz DM: Lack of association between Helicobacter pylori infection with dupA- positive strains and gastroduodenal diseases in Brazilian patients. Int J Med Microbiol. 2008, 298: 223-230. 10.1016/j.ijmm.2007.05.006.
Argent RH, Burette A, Miendje Deyi VY, Atherton JC: The presence of dupA in Helicobacter pylori is not significantly associated with duodenal ulcer in Belgium, South Africa, China or North America. Clin Infect Dis. 2007, 45: 1204-1206. 10.1086/522177.
Miehlke S, Kibler K, Kim JC, Figura N, Small SM, Graham DY, Go MF: Allelic variation in the cagA gene of Helicobacter pylori obtained from Korea compared to the United States. Am J Gastroenterol. 1996, 91: 1322-1325.
Cao P, Lee KJ, Blaser MJ, Cover TL: Analysis of hopQ alleles in East Asian and Western strains of Helicobacter pylori. FEMS Microbiol Lett. 2005, 251: 37-43. 10.1016/j.femsle.2005.07.023.
Tan HJ, Rizal AM, Rosmadi MY, Goh KL: Distribution of Helicobacter pylori cagA, cagE and vacA in different ethnic groups in Kuala Lumpur, Malaysia. J Gastroenterol Hepatol. 2005, 20: 589-594. 10.1111/j.1440-1746.2005.03783.x.
Lu H, Yamaoka Y, Graham DY: Helicobacter pylori virulence factors: facts and fantasies. Curr Opin Gastroenterol. 2005, 21: 653-659. 10.1097/01.mog.0000181711.04529.d5.
Lee A, O'Rourke J, Ungria MCD, Robertson B, Daskalopoulos G, Dixon MF: Erratum: a standardized mouse model of Helicobacter pylori infection – introducing the Sydney strain. Gastroenterol. 1997, 113: 732-
Lee A, O'Rourke J, Ungria MCD, Robertson B, Daskalopoulos G, Dixon MF: A standardized mouse model of Helicobacter pylori infection – introducing the Sydney strain. Gastroenterol. 1997, 112: 1386-1397. 10.1016/S0016-5085(97)70155-0.
Enroth H, Kraaz W, Engstrand L, Nyrén O, Rohan T: Helicobacter pylori strain types and risk of gastric cancer: a case-control study. Cancer Epidemiol Biomarkers Prev. 2000, 9: 981-985.
Borody T, Clancy R, Warren EF, Surace R, Brusentsev S, Mitchell H: Antibiotic sensitivities of Helicobacter pylori vary at different gastric mucosal sites. Helicobacter pylori: Basic mechanisms to clinical cure 2002. Edited by: Hunt R, Tytgat G. 2003, Dordrecht: Kluwer Academic Publishers, 373-382.
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ: Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucl Acids Res. 1997, 25: 3389-3402. 10.1093/nar/25.17.3389.
Thompson JD, Higgins DG, Gibson TJ: CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl Acids Res. 1994, 22: 4673-4680. 10.1093/nar/22.22.4673.
Bjorkholm B, Lundin A, Sillen A, Guillemin K, Salama N, Rubio C, Gordon JI, Falk P, Engstrand L: Comparison of genetic divergence and fitness between two subclones of Helicobacter pylori. Infect Immun. 2001, 69: 7832-7838. 10.1128/IAI.69.12.7832-7838.2001.
Hussein N, Mohammadi M, Talebkhan Y, Doraghi M, Letley D, Muhammad M, Argent R, Atherton J: Differences in virulence markers between Helicobacter pylori strains from Iraq and those from Iran: potential importance of regional differences in H. pylori-associated disease. J Clin Microbiol. 2008, 46: 1774-1779. 10.1128/JCM.01737-07.
Zhang Z, Zheng Q, Chen X, Xiao S, Liu W, Lu H: The Helicobacter pylori duodenal ulcer promoting gene, dupA, in China. BMC Gastroenterol. 2008, 8: 49-10.1186/1471-230X-8-49.
Arachchi HS, Kalra V, Lal B, Bhatia V, Baba CS, Chakravarthy S, Rohatgi S, Sarma PM, Mishra V, Das B, Ahuja V: Prevalence of duodenal ulcer promoting gene (dupA) of Helicobacter pylori in patients with duodenal ulcer in north Indian population. Helicobacter. 2007, 12: 591-597.
Pacheco AR, Proença-Módena JL, Sales AI, Fukuhara Y, Silveira WDd, Pimenta-Módena JL, Oliveira RBd, Brocchi M: Involvement of the Helicobacter pylori plasticity region and cag pathogenicity island genes in the development of gastroduodenal diseases. Eur J Clin Microbiol Infect Dis. 2008, 27: 1053-1059. 10.1007/s10096-008-0549-8.
Ahmed N, Sechi LA: Helicobacter pylori and gastroduodenal pathology: new threats from an old friend. Ann Clin Microbiol Antimicrob. 2005, 4: 1-10.1186/1476-0711-4-1.
Douraghi M, Mohammadi M, Oghalaie A, Abdirad A, Mohagheghi M, Hosseini M, Zeraati H, Ghasemi A, Esmaieli M, Mohajerani N: dupA as a risk determinant in Helicobacter pylori infection. J Med Microbiol. 2008, 57: 554-562. 10.1099/jmm.0.47776-0.
Falush D, Wirth T, Linz B, Pritchard JK, Stephens M, Kidd M, Blaser MJ, Graham DY, Vacher S, Perez-Perez GI, et al: Traces of human migrations in Helicobacter pylori populations. Science. 2003, 299: 1582-1585. 10.1126/science.1080857.
Acknowledgements
This work was supported by The Cancer Council of New South Wales, Australia (REF 66/04). Sönke Andres was supported by a grant from the Marie Curie Early Stage Research Training Fellowship of the European Community's Sixth Framework Program called IMO-train.
We would like to thank Alfred Tay for providing the calculations on the pairwise sequence variation for the Falush et al. [33] study. We would also like to thank Quanjiang Dong for his help with primer design.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
HMAS designed the study, prepared the Malaysian isolates for analysis, designed and carried out the molecular genetic studies, carried out the sequence alignment and analysis, performed the statistical analysis, helped design and participated in the in vitro studies and drafted the manuscript. SA helped design and participated in the in vitro studies. NK participated in the molecular genetic studies. LEr prepared the Swedish isolates for analysis. HM was awarded grant to support the study, participated in the design of the study and helped to draft the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Schmidt, HM.A., Andres, S., Kaakoush, N.O. et al. The prevalence of the duodenal ulcer promoting gene (dupA) in Helicobacter pylori isolates varies by ethnic group and is not universally associated with disease development: a case-control study. Gut Pathog 1, 5 (2009). https://doi.org/10.1186/1757-4749-1-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1757-4749-1-5