
Abstract
Background
As a result of globalization and climate change, Dirofilaria immitis and Dirofilaria repens, the causative agents of dirofilariosis in Europe, continue to spread from endemic areas in the Mediterranean to northern and northeastern regions of Europe where autochthonous cases of dirofilarial infections have increasingly been observed in dogs and humans. Whilst D. repens was recently reported from mosquitoes in putatively non-endemic areas, D. immitis has never been demonstrated in mosquitoes from Europe outside the Mediterranean.
Methods
From 2011 to 2013, mosquitoes collected within the framework of a German national mosquito monitoring programme were screened for filarial nematodes using a newly designed filarioid-specific real-time PCR assay. Positive samples were further processed by conventional PCR amplification of the cytochrome c oxidase subunit I (COI) gene, amplicons were sequenced and sequences blasted against GenBank.
Results
Approximately 17,000 female mosquitoes were subjected to filarial screening. Out of 955 pools examined, nine tested positive for filariae. Two of the COI sequences indicated D. immitis, one D. repens and four Setaria tundra. Two sequences could not be assigned to a known species due to a lack of similar GenBank entries. Whilst D. immitis and the unknown parasites were detected in Culex pipiens/torrentium, D. repens was found in a single Anopheles daciae and all S. tundra were demonstrated in Aedes vexans. All positive mosquitoes were collected between mid-June and early September.
Conclusion
The finding of dirofilariae in German mosquitoes implies the possibility of a local natural transmission cycle. While the routes of introduction to Germany and the origin of the filariae cannot be determined retrospectively, potential culicid vectors and reservoir hosts must prospectively be identified and awareness among physicians, veterinarians and public health personnel be created. The health impact of S. tundra on the indigenous cervid fauna needs further investigation.
Similar content being viewed by others
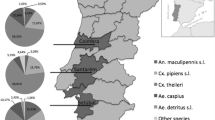

Background
The dirofilarial species D. immitis and D. repens are the causative agents of cardiopulmonary and subcutaneous dirofilariosis, respectively, in canines, felines and other carnivores in Europe [1]. In occasional infections of humans, the nematodes may evoke subcutaneous, subconjunctival and cardiovascular lesions with infections of the lung and pulmonary blood vessels. Intra- and retroocular infections as well as infections of deeper locations such as the peritoneal cavity, the omentum and the male sexual organs may occur [1, 2]. Also, rare cases of meningoencephalitis have been described [3]. Both worms are endemic in southern Europe where numbers of notified human cases of dirofilariasis have substantially increased recently [4, 5]. Contrary to previous assumptions, the majority of these had probably been caused by D. repens[6]. In addition, an ongoing north- and eastward spread of both species has been observed, attributed to increased travel and movement of infected animals, the expansion of vector-competent mosquito species, global warming and a change in human activities [7, 8]. Thus, autochthonous cases of D. repens infection in dogs were reported from Germany in 2004 [9] and from the Netherlands in 2008 [10], while several human autochthonous cases have been diagnosed in Poland since 2007 [11]. In 2007 and 2012, D. repens was again diagnosed in dogs in the German federal states of Baden-Wurttemberg and Brandenburg [12–14], suggesting that endemic circulation takes place. Moreover, autochthonous cases of D. immitis infection in dogs were reported from Hungary in 2009 [15], Slovakia in 2010 [16] and Poland in 2012 [17].
The primarily boreal filarial species S. tundra lives in the abdominal cavity of cervids. Setariae are commonly believed to be non-pathogenic in their natural hosts but severe disease outbreaks with associated peritonitis and perihepatitis caused by S. tundra have been reported [18]. In Scandinavian countries, reindeer is the main vertebrate host [19] whereas in Central Europe only roe and red deer have been found parasitized so far [20–22]. Human infections have not been described.
Dirofilariae are transmitted by culicid mosquitoes (Diptera, Culicidae) of various species, such as Cx. pipiens, Anopheles maculipennis s.l. and Aedes albopictus, which are probably the most important vectors in the Mediterranean [23–25]. Some of these, such as Cx. pipiens and An. maculipennis s.l., are widely distributed over Europe while others, such as Ae. albopictus, are expanding northwards from established distribution areas in the Mediterranean [26]. The main vectors of S. tundra are supposed to belong to the genus Aedes[27].
Within a German national mosquito monitoring programme launched in 2011, mosquito samples were screened for various pathogens such as viruses and filarial nematodes. We describe here the finding of at least four filarial nematode species in mosquitoes collected in Germany, including the first detection of D. immitis in Germany.
Methods
Adult mosquitoes were collected at numerous sites all over Germany between 2011 and 2013 using BG sentinel traps (Biogents, Germany) equipped with BG Lure™ and CO2 as attractants, or by hand. The mosquitoes were caught by trained non-specialists who kept them frozen until further processing. Upon transportation to the laboratory, the mosquitoes were identified morphologically [28, 29] or genetically, following RNA/DNA extraction as described below. Specifically, Maculipennis Group species (An. maculipennis s.l.) were identified by species-specific PCR [30], whereas mosquitoes neither identifiable morphologically nor by PCR were subjected to COI barcoding [31].
A total of approximately 17,000 female mosquitoes belonging to six genera (Aedes, Anopheles, Coquillettidia, Culex, Culiseta and Ochlerotatus) were pooled by species, collection site and date with up to 25 specimens per pool. Mosquitoes identified by COI barcoding or species-specific PCR represented pools consisting of one specimen only. Each pool was homogenized in the presence of stainless steel beads (diameter 3 mm) in a maximum of 750 μl minimum essential medium (MEM) containing 10 μg/ml gentamicin, 0.25 μg/ml amphotericin B, 100 U/ml penicillin and 100 μg/ml streptomycin by a TissueLyserII (Qiagen, Germany) for 3 min at 30 Hz. The homogenate was centrifuged for 1 min at 14,000 g, and the supernatant was used for simultaneous RNA/DNA extraction by means of the NucleoSpin 96 Virus Core Kit (Macherey-Nagel, Germany) according to the user manual.
For screening the mosquito pools for filarial nematodes (Filarioidea), a filarioid-specific real-time PCR assay was developed targeting a 90 bp fragment of the mitochondrial 16S rRNA gene with the newly designed primers PanFilaF (5’-TGTGCTGCGCTACATCGATG-3’) and PanFilaR (5’-AAACCGCTCTGTCTCACGAC-3’). The primers were constructed after alignment of partial and complete mitochondrial genome sequences of nine parasitic filarial nematode species (D. immitis, D. repens, S. tundra, Setaria digitata, Brugia malayi, Wuchereria bancrofti, Onchocerca flexuosa, Onchocerca volvulus), which are epidemiologically important in human and animal health, and, additionally, are appropriately represented in GenBank. Sequences were analysed with BioEdit Sequence Alignment Editor [32], and conserved DNA regions were identified for primer design taking into account standard rules of designing primers for real-time PCR assays [33]. Specificity of the primers was confirmed on D. immitis, D. repens, O. volvulus and W. bancrofti DNA. A more in-depth testing was not considered necessary as the PCR was meant for sample screening only, thus possibly allowing false negative but not false positive results. The real-time PCR was performed using the CFX96 Touch™ Real-Time PCR Detection System (BioRad, Germany) and ResoLight non-specific detection chemistry, followed by high-resolution melting-analysis. The reaction mixture (25 μl) contained 1 μl ResoLight dye (Roche Diagnostics, Germany), 10 μl of 2× QuantiTect Multiplex PCR Master Mix (Qiagen, Germany), 0.4 μM forward and reverse primer each, and 5 μl of extracted DNA. The thermoprofile consisted of an initial denaturation step at 95°C for 15 min, 35 cycles of 95°C for 45 sec, 58°C for 30 sec and 72°C for 45 sec, and a final extension step at 72°C for 5 min. All amplifications and detections were carried out in Multiplate™ Low-Profile 96-Well PCR Plates with optical Microseal 'B' Film (BioRad). After each annealing cycle, accumulation of PCR products was detected by monitoring the increase in fluorescence of double-stranded DNA-binding ResoLight at 518 nm. After the PCR, a dissociation curve was constructed for steps of 5°C in the range from 60°C to 95°C. All data were analyzed using the BioRad CFX-Manager software.
Samples yielding a signal in the real-time PCR were processed by a second conventional PCR amplifying about 650 bp of the filarioid COI gene [34]. After agarose gel electrophoresis, PCR products were excised from the gels and recovered by the QIAquick Gel Extraction Kit (Qiagen, Germany). They were cycled bidirectionally using the BigDye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems, Germany), and sequencing products were purified by SigmaSpin Sequencing Reaction Clean-Up Columns (Sigma Aldrich, Germany) before loading onto a 3130 Genetic Analyser (Applied Biosystems). For species identification, consensus sequences of positive samples were compared with sequences available in GenBank.
Results
16,878 female mosquitoes representing 16 species or species complexes in six genera were processed, with Cx. pipiens/torrentium (73%), Anopheles plumbeus (11%) and Aedes vexans (8%) being the most frequent mosquito taxa examined (Table 1). In total, 955 pools were screened using the pan-filarioid real-time PCR. Nine pools (0.94%) testing positive were confirmed to contain filarioid DNA by the COI PCR assay. Of the sequences obtained, two showed 100% and 99% identity to D. immitis (Table 2) while presenting two nucleotide differences in direct comparison. One sequence displayed 99% identity to D. repens and four sequences exhibited 99% identity to S. tundra, with the S. tundra sequences being variable among each other at six positions. Two positive samples, displaying 91% homology in direct comparison, could not be assigned to a species due to insufficient identities to GenBank entries (89 and 92% maximum; Table 2). The D. immitis-positive pools as well as those with unknown filarioid DNA sequences were composed of Cx. pipiens/torrentium specimens, the D. repens-positive pool was equivalent to a single An. daciae female, and the S. tundra-positive pools contained Ae. vexans mosquitoes (Table 2). All filariae-carrying culicids had been collected between mid-June and early September in 2011, 2012 and 2013 in four German federal states (Table 2, Figure 1).

Geographic origin of the mosquitoes tested positive (red dots: D. immitis , green dot: D. repens , blue dots: S. tundra , yellow dots: filariae of unknown species).
The COI DNA sequences of the identified worms found in the mosquitoes have been deposited in GenBank under accession numbers KF692100-KF692106.
Discussion
As dirofilariosis is a vector-borne disease, its epidemiology is highly susceptible to climatic and environmental conditions. In the recent past, it has become an emerging problem in numerous countries of the world, including many European ones [1, 7]. The two causative agents in Europe, D. immitis and D. repens, have been found outside their traditional distribution ranges in the Mediterranean with increasing frequency. In particular, in Central and eastern European states, such as Austria, Hungary, Slovakia and Poland, autochthonous cases have been diagnosed in dogs and humans [11, 16, 35–38].
With D. immitis and D. repens, two mosquito-borne zoonotic filarial nematode species endemic to southern Europe were detected in mosquitoes collected in Germany. Hence, this is the first report from Germany of D. immitis apparently acquired locally.
Notably, the various filarial species were demonstrated in their putative vectors which included An. daciae, a recently recognized member of the Maculipennis Group [39], a complex of several closely related isomorphic Anopheles species, in addition to two previously described potential vectors of the filariae, Ae. vexans and Cx. pipiens (due to the pooling of the mosquitoes, a molecular differentiation between the morphologically indistinguishable females of Cx. pipiens and Cx. torrentium was not carried out in this study). The detection of the filariae in these mosquito species is not surprising as Cancrini et al.[23, 24] found D. repens in Cx. pipiens and Maculipennis Group specimens in Italy while Bocková et al. [40] only recently reported D. repens in a pool of Ae. vexans mosquitoes from Slovakia. Dirofilaria immitis has been described from Cx. pipiens in Spain [41], and from Ae. vexans and Cx. pipiens in Turkey [42].
The finding of two unknown filarial species in Cx. pipiens/torrentium mosquito pools suggests avian bloodmeal sources of the mosquitoes due to the feeding preferences within this group of culicids, and therefore avian nematode species, one of them possibly being Cardiofilaria pavlovskyi, as discussed by Cjaka et al. [43].
The route through which the dirofilariae found their way to Germany or the sources of filarial ingestion by the mosquitoes, respectively, must remain speculative. The D. immitis-positive mosquito/es from Baden-Wurttemberg was/were collected at the same site where Ae. albopictus had repeatedly been trapped previously [26]. This site is characterized by its close proximity to a railway transshipment station where cargo from trucks coming in from southern Europe is transferred to trains. Hence, it is conceivable that, as with Ae. albopictus, the filarioid-positive Cx. pipiens/torrentium mosquito/es was/were introduced from southern Europe by vehicle transport. By contrast, the finding of D. immitis in a pool of mosquitoes from Brandenburg must be attributed most probably to a local uptake by the feeding mosquito/es. A possible source might have been a dog imported from, or with a travel history to, southern Europe. The detection of D. repens in An. daciae is noteworthy not only because nothing is known about the vector potential of this mosquito species but also because it was collected in the same area where D. repens had been isolated from dogs in 2007 [12]. Possibly, a local transmission cycle has established in that area.
The third mosquito-borne filarial nematode described, S. tundra, seems to be more common in Germany than generally assumed, as it had been detected microscopically or by PCR on several occasions in the past [22, 43]. Detailed studies regarding its abundance, distribution and even pathology in areas south of Scandinavia, however, are lacking.
As surprising as the dirofilarial findings are, Genchi and colleagues [5, 44] considered both Baden-Wurttemberg and Brandenburg as climatically suitable for dirofilarial development in mosquitoes and assigned to these regions a risk of stable endemicity.
Simón et al.[1] calculated a transmission period of 3–4 months in Central Europe for both D. repens and D. immitis, taking into account the extrinsic incubation periods of the worms. At a mean temperature of 18°C, for example, these will last about 29 days, at 22°C still 16–20 days. Setaria tundra needs an average of 14–16 days at 21°C to reach the metacyclic infectious L3 stage in the mosquito [45]. Considering the lifespan of a mosquito of a few weeks at its best, these long developmental periods may presently limit the rate of dispersal of the filariae. However, as hot summer periods are predicted to become more frequent and longer as a result of climate change, mosquito-borne filarioses will probably become a growing problem to veterinary and human health in Central and eastern Europe in the future.
Conclusion
With progressing globalization and climate change, the risk of the introduction of zoonotic D. immitis and D. repens from endemic areas in southern Europe to previously infection-free areas in northern Europe and their subsequent establishment increases. Vector-competent mosquitoes are probably already present there, and the climatic conditions are regionally and seasonally adequate for the filariae to finish their development in infected mosquitoes. The possibility of dirofilarial infections in dogs and other carnivores as well as in humans should therefore be considered with regard to differential diagnosis in unclear cases of appropriate symptomatology.
No information exists on a possible spread and an increase in prevalence of S. tundra in Central Europe as respective studies are missing. Although this worm does not appear to significantly affect the health of indigenous cervids at present, further research on the epidemiology of setariosis in Central Europe is desirable.
Abbreviations
- COI:
-
Cytochrome c oxidase subunit I.
References
Simón F, Siles-Lucas M, Morchón R, González-Miguel J, Mellado I, Carretón E, Montoya-Alonso JA: Human and animal dirofilariasis: the emergence of a zoonotic mosaic. Clin Microbiol Rev. 2012, 25: 507-544. 10.1128/CMR.00012-12.
McCall JW, Genchi C, Kramer LH, Guerrero J, Venco L: Heartworm disease in animals and humans. Adv Parasitol. 2008, 66: 193-285.
Poppert S, Hodapp M, Krueger A, Hegasy G, Niesen WD, Kern WV, Tannich E: Dirofilaria repens infection and concomitant meningoencephalitis. Emerg Infect Dis. 2009, 15: 1844-1846. 10.3201/eid1511.090936.
Simón F, López-Belmonte J, Marcos-Atxutegi C, Morchón R, Martín-Pacho JR: What is happening outside North America regarding human dirofilariasis?. Vet Parasitol. 2005, 133: 181-189. 10.1016/j.vetpar.2005.03.033.
Genchi C, Kramer LH, Rivasi F: Dirofilarial infections in Europe. Vector-borne Zoon Dis. 2011, 11: 1307-1317. 10.1089/vbz.2010.0247.
Pampiglione S, Rivasi F, Gustinelli A: Dirofilarial human cases in the Old World, attributed to Dirofilaria immitis: a critical analysis. Histopathology. 2009, 54: 192-204. 10.1111/j.1365-2559.2008.03197_a.x.
Genchi C, Mortarino M, Rinaldi L, Cringoli G, Traldi G, Genchi M: Changing climate and changing vector-borne disease distribution: the example of Dirofilaria in Europe. Vet Parasitol. 2011, 176: 295-299. 10.1016/j.vetpar.2011.01.012.
Morchón R, Carretón E, González-Miguel J, Mellado-Hernández I: Heartworm disease (Dirofilaria immitis) and their vectors in Europe - new distribution trends. Front Physiol. 2012, 3: 196-
Hermosilla C, Pantchev N, Dyachenko V, Gutmann M, Bauer C: First autochthonous case of canine ocular Dirofilaria repens infection in Germany. Vet Rec. 2006, 158: 134-135. 10.1136/vr.158.4.134.
Overgaauw P, van Dijk E: Autochthonous case of Dirofilaria repens in a dog in the Netherlands. Vet Rec. 2009, 164: 158-10.1136/vr.164.5.158.
Cielecka D, Żarnowska-Prymek H, Masny A, Salamatin R, Wesołowska M, Gołąb E: Human dirofilariosis in Poland: the first cases of autochthonous infections with Dirofilaria repens. Ann Agric Environ Med. 2012, 19: 445-450.
Pantchev N, Norden N, Lorentzen L, Rossi M, Rossi U, Brand B, Dyachenko V: Current surveys on the prevalence and distribution of Dirofilaria spp. in dogs in Germany. Parasitol Res. 2009, 105: S63-S74. 10.1007/s00436-009-1497-7.
Sassnau R, Dyachenko V, Pantchev N, Stöckel F, Dittmar K, Daugschies A: Dirofilaria repens infestation in a sled dog kennel in the federal state of Brandenburg (Germany). Diagnosis and therapy of canine cutaneous dirofilariosis. Tierärztl Prax. 2009, 37 (K): 95-101. [Article in German]
Sassnau R, Kohn M, Demeler J, Kohn B, Müller E, Krücken J, von Samson-Himmelstjerna G: Is Dirofilaria repens endemic in the Havelland district in Brandenburg, Germany?. Vector-borne Zoon Dis. 2013, 13: 888-891. 10.1089/vbz.2012.1293.
Jacso O, Mandoki M, Majoros G, Petsch M, Mortarino M, Genchi C, Fok E: First autochthonous Dirofilaria immitis (Leidy, 1856) infection in a dog in Hungary. Helminthologia. 2009, 46: 159-161. 10.2478/s11687-009-0030-y.
Miterpáková M, Antolová D, Hurníková Z, Dubinský P, Pavlacka A, Németh J: Dirofilaria infections in working dogs in Slovakia. J Helminthol. 2010, 84: 173-176. 10.1017/S0022149X09990496.
Świątalska A, Demiaszkiewicz AW: First autochthonous case of Dirofilaria immitis invasion in dog in Poland. Žycie Weterynaryjne. 2012, 87: 685-686. [Article in Polish]
Laaksonen S, Kuusela J, Nikander S, Nylund M, Oksanen A: Outbreak of parasitic peritonitis in reindeer in Finland. Vet Rec. 2007, 160: 835-841. 10.1136/vr.160.24.835.
Laaksonen S, Oksanen A: Status and review of the vector-borne nematode Setaria tundra in Finnish cervids. Alces J. 2009, 45: 81-84.
Böhm LK, Supperer R: Untersuchungen über Setarien (Nematoda) bei heimischen Wiederkäuern und deren Beziehung zur „epizootischen cerebrospinalen Nematodiasis“ (Setariosis). Z Parasitenk. 1955, 17: 165-174.
Kutzer E, Hinaidy HK: Die Parasiten der wildlebenden Wiederkäuer Österreichs. Z Parasitenk. 1969, 32: 354-368.
Rehbein S, Lutz W, Visser M, Winter R: Beiträge zur Kenntnis der Parasitenfauna des Wildes in Nordrhein-Westfalen. 1. Der Endoparasitenbefall des Rehwildes. Z Jagdwiss. 2000, 4: 248-269.
Cancrini G, Frangipane Di Regalbono A, Ricci I, Tessarin C, Gabrielli S, Pietrobelli M: Aedes albopictus is a natural vector of Dirofilaria immitis in Italy. Vet Parasitol. 2003, 118: 195-202. 10.1016/j.vetpar.2003.10.011.
Cancrini G, Magi M, Gabrielli S, Arispici M, Tolari F, Dell’Omodarme M, Prati MC: Natural vectors of dirofilariasis in rural and urban areas of the Tuscan region, central Italy. J Med Entomol. 2006, 43: 574-579. 10.1603/0022-2585(2006)43[574:NVODIR]2.0.CO;2.
Cancrini G, Scaramozzino P, Gabrielli S, Di Paolo M, Toma L, Romi R: Aedes albopictus and Culex pipiens implicated as natural vectors of Dirofilaria repens in central Italy. J Med Entomol. 2007, 44: 1064-1066. 10.1603/0022-2585(2007)44[1064:AAACPI]2.0.CO;2.
Kampen H, Kronefeld M, Zielke D, Werner D: Further specimens of the Asian tiger mosquito Aedes albopictus (Diptera, Culicidae) trapped in southwest Germany. Parasitol Res. 2013, 112: 905-907. 10.1007/s00436-012-3128-y.
Laaksonen S, Solismaa M, Kortet R, Kuusela J, Oksanen A: Vectors and transmission dynamics for Setaria tundra (Filarioidea; Onchocercidae), a parasite of reindeer in Finland. Parasit Vectors. 2009, 2: 3-10.1186/1756-3305-2-3.
Schaffner F, Angel G, Geoffroy B, Hervy JP, Rhaiem A, Brunhes J: The Mosquitoes of Europe (CD-ROM). 2001, Montpellier, France: IRD Éditions & EID Méditerrannée
Becker N, Petrić D, Zgomba M, Boase C, Madon M, Dahl C, Kaiser A: Mosquitoes and their control. 2010, Heidelberg: Springer, 2
Proft J, Maier WA, Kampen H: Identification of six sibling species of the Anopheles maculipennis complex (Diptera: Culicidae) by a polymerase chain reaction assay. Parasitol Res. 1999, 85: 837-843. 10.1007/s004360050642.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R: DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994, 3: 294-299.
Hall TA: BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser. 1999, 41: 95-98.
Dorak T: Real-time PCR. 2006, New York, USA: Taylor & Francis Group
Casiraghi M, Anderson TJC, Bandi C, Bazzocchi C, Genchi C: A phylogenetic analysis of filarial nematodes: comparison with the phylogeny of Wolbachia endosymbionts. Parasitology. 2001, 122: 93-103. 10.1017/S0031182000007149.
Duscher G, Feiler A, Wille-Piazzai W, Bakonyi T, Leschnik M, Miterpáková M, Kolodziejek J, Nowotny N, Joachim A: Detection of Dirofilaria in Austrian dogs. Berl Munch Tierarztl Wochenschr. 2009, 122: 199-203. [Article in German]
Auer H, Susani M: The first autochthonous case of subcutaneous dirofilariosis in Austria. Wien Klin Wochenschr. 2008, 120: 104-106. 10.1007/s00508-008-1031-4. [Article in German]
Szénási Z, Kovács AH, Pampiglione S, Fioravanti ML, Kucsera I, Tánczos B, Tiszlavicz L: Human dirofilariosis in Hungary: an emerging zoonosis in central Europe. Wien Klin Wochenschr. 2008, 120: 96-102. 10.1007/s00508-008-0928-2.
Demiaszkiewicz AW, Polańczyk G, Pyziel AM, Kuligowska I, Lachowicz J: The first foci of dirofilariosis of dogs evoked by Dirofilaria repens Railliet et Henry, 1911 in central Poland. Wiad Parazytol. 2009, 55: 367-370. [Article in Polish]
Kronefeld M, Dittmann M, Zielke D, Werner D, Kampen H: Molecular confirmation of the occurrence in Germany of Anopheles daciae (Diptera, Culicidae). Parasit Vectors. 2012, 5: 250-10.1186/1756-3305-5-250.
Bocková E, Rudolf I, Kočišová A, Betášová L, Venclíková K, Mendel J, Hubálek Z: Dirofilaria repens microfilariae in Aedes vexans mosquitoes in Slovakia. Parasitol Res. 2013, 112: 3465-3470. 10.1007/s00436-013-3526-9.
Morchón R, Bargues MD, Latorre JM, Melero-Alcíbar R, Pou-Barreto C, Mas-Coma S, Simón F: Haplotype H1 of Culex pipiens implicated as a natural vector of Dirofilaria immitis in an endemic area of Western Spain. Vector-borne Zoon Dis. 2007, 7: 653-658. 10.1089/vbz.2007.0124.
Yildirim A, Inci A, Duzlu O, Biskin Z, Ica A, Sahin I: Aedes vexans and Culex pipiens as the potential vectors of Dirofilaria immitis in Central Turkey. Vet Parasitol. 2011, 178: 143-147. 10.1016/j.vetpar.2010.12.023.
Czajka C, Becker N, Poppert S, Jöst H, Schmidt-Chanasit J, Krüger A: Molecular detection of Setaria tundra (Nematoda: Filarioidea) and an unidentified filarial species in mosquitoes in Germany. Parasit Vectors. 2012, 5: 14-10.1186/1756-3305-5-14.
Sassnau R, Genchi C: Qualitative risk assessment for the endemisation of Dirofilaria repens in the state of Brandenburg (Germany) based on temperature-dependent vector competence. Parasitol Res. 2013, 112: 2647-2652. 10.1007/s00436-013-3431-2.
Laaksonen S: Setaria tundra, an emerging parasite of reindeer, and an outbreak it caused in Finland in 2003–2006. 2010, Finland: Ph.D. thesis, University of Helsinki
Acknowledgments
This work was financially supported by the German Federal Ministry of Food, Agriculture and Consumer Protection (BMELV) through the Federal Office for Agriculture and Food (BLE), grant number 2810HS022, and by the Robert Koch Institute, grant number 1362/1-982. We are grateful to Anna Albers, Institute of Medical Microbiology, Immunology and Parasitology, University of Bonn, Germany, for providing filarial control DNA, to Petra Kranz, Friedrich-Loeffler-Institut, for preparing the figure and to Adrian Pont, Oxford University Museum for Natural History, UK, for critically reading the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MK, HK and DW designed the study, collected and identified the mosquitoes and contributed to the data analysis and writing of the manuscript. MK established the detection techniques and screened the mosquitoes in the laboratory. HK and DW supervised the field and laboratory work. RS organized mosquito trapping in BB and added to preparing the manuscript. All authors read and approved the final version of the manuscript.
Mandy Kronefeld, Helge Kampen and Doreen Werner contributed equally to this work.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Kronefeld, M., Kampen, H., Sassnau, R. et al. Molecular detection of Dirofilaria immitis, Dirofilaria repens and Setaria tundra in mosquitoes from Germany. Parasites Vectors 7, 30 (2014). https://doi.org/10.1186/1756-3305-7-30
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1756-3305-7-30