
Abstract
Background
This is the second in a series of three articles documenting the geographical distribution of 41 dominant vector species (DVS) of human malaria. The first paper addressed the DVS of the Americas and the third will consider those of the Asian Pacific Region. Here, the DVS of Africa, Europe and the Middle East are discussed. The continent of Africa experiences the bulk of the global malaria burden due in part to the presence of the An. gambiae complex. Anopheles gambiae is one of four DVS within the An. gambiae complex, the others being An. arabiensis and the coastal An. merus and An. melas. There are a further three, highly anthropophilic DVS in Africa, An. funestus, An. moucheti and An. nili. Conversely, across Europe and the Middle East, malaria transmission is low and frequently absent, despite the presence of six DVS. To help control malaria in Africa and the Middle East, or to identify the risk of its re-emergence in Europe, the contemporary distribution and bionomics of the relevant DVS are needed.
Results
A contemporary database of occurrence data, compiled from the formal literature and other relevant resources, resulted in the collation of information for seven DVS from 44 countries in Africa containing 4234 geo-referenced, independent sites. In Europe and the Middle East, six DVS were identified from 2784 geo-referenced sites across 49 countries. These occurrence data were combined with expert opinion ranges and a suite of environmental and climatic variables of relevance to anopheline ecology to produce predictive distribution maps using the Boosted Regression Tree (BRT) method.
Conclusions
The predicted geographic extent for the following DVS (or species/suspected species complex*) is provided for Africa: Anopheles (Cellia) arabiensis, An. (Cel.) funestus*, An. (Cel.) gambiae, An. (Cel.) melas, An. (Cel.) merus, An. (Cel.) moucheti and An. (Cel.) nili*, and in the European and Middle Eastern Region: An. (Anopheles) atroparvus, An. (Ano.) labranchiae, An. (Ano.) messeae, An. (Ano.) sacharovi, An. (Cel.) sergentii and An. (Cel.) superpictus*. These maps are presented alongside a bionomics summary for each species relevant to its control.
Similar content being viewed by others

Background
This paper is a second in a series of three contributions discussing the geographic distribution and bionomics of the dominant vector species (DVS) of human malaria [1, 2]. It deals specifically with the DVS of Africa, Europe and the Middle East.
Despite highly variable levels of transmission across Africa [3, 4], the global public heath impact of P. falciparum malaria is overwhelmingly felt on this continent [5, 6]. Africa contains areas with the highest entomological inoculation rates [3, 7] and prevalence levels [8] globally, and thus the highest morbidity and mortality [5]. This situation arises partly because Africa has the most effective and efficient DVS of human malaria [9, 10]: An. gambiae (sensu stricto - herein, referred to as 'An. gambiae'; it is not necessary to use 'sensu stricto' (or the abbreviation 's.s.') when there is no doubt that the biological species being referred to is the one that bears the name An. gambiae) [5, 10], with its sibling, An. arabiensis, also of major importance [11]. The DVS members of the An. gambiae complex also include the salt water tolerant, coastal species An. melas and An. merus [12] and these, whilst not being as efficient at transmitting malaria as An. gambiae or An. arabiensis, are often found in such high densities that they achieve DVS status [13–15]. Other members of the An. gambiae complex are either highly restricted in their distribution (e.g. An. bwambae, only currently known to occur in geothermal springs in western Uganda [11, 16]) or are zoophilic in behaviour and not considered vectors of human malaria (e.g. An. quadriannulatus and An. quadriannulatus B) [17]. In addition to the four DVS within the An. gambiae complex, large parts of Africa are also home to other DVS, including An. funestus, An. nili and An. moucheti, with An. funestus, in some cases, having a greater impact on malaria transmission even than An. gambiae [10, 11, 18]. The anthropophilic habits of these DVS are a major contributing factor to their public health impact, indeed An. funestus is considered to be one of the first species to have adapted to human hosts [19].
The vast majority of current malaria control efforts use interventions aimed at limiting human-vector contact [20, 21]. Foremost among these interventions has been the rapid scale-up of insecticide treated bednets (ITNs) [22], followed by the scale-up of indoor residual spraying (IRS) in Africa [23]. These interventions are often deployed without a detailed understanding of the distribution, species composition and behaviour of local vectors. This complicates impact monitoring [24], the appraisal of arguments for more holistic integrated vector control [25] and evaluation of the potential of novel vector control methods [26–28]. Distribution maps can also be applied to gauge the importance of emerging insecticide resistance among the DVS of Africa [29–37]. In contrast to Africa, the European and the Middle Eastern region contain areas with low to no malaria transmission [8]. Despite this, the existence of Anopheles species with the capacity to transmit malaria is often highlighted as providing the potential for the re-introduction of malaria [38–43].
A number of vector species modelling and mapping strategies have been applied on a country (e.g. [44–50]) and regional scale [51] and across the African continent [24, 52–55], with fewer attempts directed at the European and Middle Eastern species [56–58]. No previous mapping efforts formally incorporate expert opinion (EO) distributions and the methods used range in complexity, from simply plotting presence or abundance on a map [24, 44, 48, 57, 58], to the application of more sophisticated predictive models [45–47, 49, 50, 52–56]. This makes comparison between the maps difficult. Further difficulties also arise in the interpretation of existing maps as many previous studies include all historical occurrence records to compensate for poor data coverage. This can introduce taxonomic ambiguity; the An. gambiae complex, for example, was only fully categorised in 1998, with the addition of the provisionally designated An. quadriannulatus species B [12, 59] and, even now, the status of An. funestus is under question [60–63]. Moreover, the morphological similarity that hides members of a species complex adds a level of uncertainty to the identity of species data recorded before the advent of cytological or molecular identification techniques.
This current work attempts to overcome many of these problems. The same Boosted Regression Tree (BRT) methodology is applied to all DVS making comparison between predicted maps possible. Despite only using data collected after 31 December 1984 the assimilated DVS occurrence records together comprise the largest contemporary dataset for prediction, with this evidence base to be made available in the public domain. Significant efforts were also expended to update the EO maps for all species [1] and these were used to inform the predictions. The outcome of these efforts and that of a comprehensive bionomics review are presented here for the DVS of Africa, Europe and the Middle East.
Methods
The data assembly and mapping methods, climatic and environmental variable grid pre- and post-processing methods and the modelling protocol summarised here are described in detail in Sinka et al. [2]. The selection of the DVS is detailed in Hay et al. [1]. In brief, 13 DVS from a final list of 41 species and species complexes worldwide were considered, seven of which are found solely in Africa (Table 1) [1] with a further six distributed across Europe, the Middle East and in limited areas of northern Africa (Table 2).
Data assembly, data checks and expert opinion maps
Building on the existing Malaria Atlas Project (MAP [64]) library of parasite rate surveys, a systematic search of the published, peer-reviewed literature using online scientific bibliographic databases was performed and augmented with a range of other information previously described [2]. Literature searches were concluded on 31 October 2009 and all citations meeting our search criteria [2] were reviewed.
Occurrence data extracted from these sources (a detailed protocol is given in Hay et al. [1]) were subjected to a series of rigorous checks before being migrated from Excel into a web-based PostgreSQL database where a final series of checks were conducted (see Sinka et al. [2]).
Globally, the literature search resulted in 3857 publications or reports containing potential data to be reviewed. Of these publications, 2276 fulfilled the inclusion criteria, providing data for 147 countries. A total of 727 sources detailed surveys conducted across 46 countries in Africa with 45 sources found for 49 countries in Europe and the Middle East.
Using EO map overlays (Additional file 1: Expert opinion distribution maps for the seven DVS of Africa and the six DVS of the Europe and Middle Eastern region (Raster prediction files are available on request)), initially digitised from published, authoritative sources (Table 1, 2) and further refined by a Technical Advisory Group (TAG) of Anopheles experts (see acknowledgements), preliminary maps were produced displaying the occurrence data for each species. These maps were examined and points that fell outside the EO range were checked and either corrected or the EO maps adjusted to include all confirmed areas of occurrence.
Boosted Regression Trees, climatic/environmental variables and model protocol
The BRT method [65, 66] was chosen to generate the predictive maps of each DVS distribution. In a review comparing 16 species modelling methodologies, BRT consistently performed well [67] and benefits from being flexible (accommodating both categorical and continuous data), using freely available, reliable and well documented R code [68] and producing maps that are simple to interpret and include a ranked list of environmental or climatic predictors [2]. The method is described in full by Elith et al. [66] and its implementation for DVS mapping summarised by Sinka et al. [2]. The BRT also produces a number of evaluation statistics including Deviance, Correlation, Discrimination (Area Under the operating characteristic Curve: AUC) and Kappa (κ) which are used here as a guide to the predictive performance of each map.
The BRT model was provided with a suite of open access, environmental and climatic variable 5 × 5 km resolution grids, relevant to the ecology and bionomics of the DVS in the African, European and Middle Eastern regions. Each grid has undergone a series of processing steps to ensure all land and sea pixels exactly correspond, and, using nearest neighbour interpolation, to fill in any small gaps in the data due to, for example, cloud cover (see Sinka et al. [2]). Where the remotely sensed imagery was available as multi-temporal data, temporal Fourier analysis (TFA) was applied to ordinate the data, generating seven products for each temporal variable: the overall mean, maximum and minimum of the data cycles; the amplitude (maximum variation of the cycle around the mean) and the phase (the timing of the cycle) of the annual and bi-annual cycles [69]. The environmental/climatic variables applied to the BRT model included a digital elevation model (DEM) [70–72], precipitation and temperature [73, 74], land surface temperature (LST), middle infrared radiation (MIR) and the normalized difference vegetation index (NDVI) (Advanced Very High Resolution Radiometer (AVHRR) [75–78]), and 22 individual categories of land cover plus a further three grouped classes that encompassed flooded areas, forested areas and dry areas (Globcover [79]).
The AVHRR grids (LST, MIR and NDVI) were applied to all DVS except the European species An. messeae and An. atroparvus. These two species have the most northerly distribution of all the DVS, with An. messeae ranging up to 65° north. At these latitudes, the AVHRR satellite data can be problematic. Instead MODIS (MODerate Resolution Imaging Spectroradiometer) [70] data were used because it provides better coverage and fewer data gaps for these northern distributions. The MODIS grids include the Enhanced Vegetation Index (EVI) and LST [70].
Following the same protocol described in Sinka et al. [2], numerous model iterations were run to assess the 'optimal' mapping outputs, including assessing the buffer size surrounding the EO range from where pseudo-absences would be drawn, the number of pseudo-absences to apply to the model and the effects of including half weighted pseudo-presence data, allocated at random from within the EO boundary, alongside the occurrence data. As each of these categories required the use of different data inputs to the BRT, statistical comparison using the evaluation metrics was not strictly possible. Therefore the 'optimal' settings chosen are inherently subjective and based on visual examination and comparison of the various maps guided by, but not relying on, the evaluation statistics.
Bionomics
A full protocol describing the methodology used to extract species-specific bionomic data from the available literature (Table 3, 4) is given in the supplemental information accompanying Sinka et al. [2]. The bionomics summary of each species is included to accompany the predictive maps as the success of interventions and control methods, such as ITNs or IRS, in reducing malaria transmission is closely related to the behavioural characteristics of the local DVS. This review does not, however, include detailed information relating to insecticide resistance. This was a purposeful omission as it would not be possible to do full justice to this highly dynamic and important aspect of the DVS within the space confines of the current work. Moreover, insecticide resistance is being addressed in detail by other groups, including those at the Liverpool School of Tropical Medicine and the Innovative Vector Control Consortium (IVCC) [80]. Furthermore, there are a number of comprehensive reviews that have been recently produced that detail insecticide resistance amongst Afrotropical species which should be considered alongside this current work (e.g. [31, 35, 81, 82]).
Results
African DVS
A total of 4581 independent sites, of which 4234 were successfully geo-referenced, reported the presence of one or more African DVS, relating to 9300 (8646 geo-referenced) occurrences (i.e. including one or more temporal sample conducted at one independent site) (Table 5). The following results refer only to geo-referenced data, and of these 3951 sites were at a resolution (points and wide areas, <10 km2 and between 10 and 25 km2 respectively) suitable to be applied to the BRT model (from here on, for simplicity, referred to as points).
Data were recorded from a total of 46 countries, 44 of which reported points. The largest number of data were reported from Kenya, with a total of 757 sites (all area types), 686 points and 1599 occurrence data (all area types). In contrast, only one data point was reported from Togo (Kantindi) where An. gambiae was found [83] and studies from Mauritius only provided DVS location information, at a polygon level, for two sites. African DVS data were reported from Egypt, but only in the form of a polygon location that could not be successfully geo-referenced. Anopheles gambiae was reported from the largest number of countries (34) and from the highest number of point locations (1443), however occurrence data (from point locations only) were greater for both An. funestus and An. arabiensis (2692 and 2301, respectively) than for An. gambiae (2291). The least prevalent species was An. moucheti reported from only 66 point locations (Table 6) and Cameroon had the highest diversity of DVS with three sites (Nkoteng, Tibati and Mayo Mbocki) showing the presence of five DVS (An. arabiensis, An. funestus, An. gambiae, An. nili and An. moucheti) [84–86].
Adult resting collections were the most popular sampling method, with 424 studies collecting females resting inside houses compared to 178 studies that collected females biting indoors. Outdoor resting sampling was comparably rare with 56 studies collecting from outdoor shelters, 22 studies searching inside animal sheds and 21 studies where the details of the outdoor location sampled were not recorded. Outdoor landing catches were conducted in 132 studies and 181 studies collected larvae, relating to 675 point locations.
Molecular techniques examining nucleic acids, which have only been applied for identification on a regular basis since the 1990s [87], were well represented, with 338 studies reporting the use of Polymerase Chain Reaction (PCR) methods. Morphological methods were used in 363 studies, often in conjunction with PCR techniques. At the other end of the scale, salinity tolerance tests were only attempted in four studies and cross-mating experiments only in five.
European and Middle Eastern DVS
Across the European and Middle Eastern region, 49 countries reported the presence of one or more DVS from 2820 point locations (all locations: 2891), of which 2784 were successfully geo-referenced (all geo-referenced locations: 2848) (Table 7). Relatively few polygon data were reported (all: 71/2891, georeferenced only: 64/2848) and longitudinal studies were also rare, with only 18 studies reporting sampling on more than one occasion at the same site. A total of 3020 geo-referenced occurrence data across all area types, with 2946 from point locations, were compiled. Considering only the geo-referenced data, DVS presence was reported from the most sites in Italy (all sites: 423, point only: 409).
Anopheles atroparvus was the species reported most often across the region, found at 1051 geo-referenced locations, of which 1044 were available to be used in the analyses. Anopheles sergentii was only present at 35 point locations, and within 11 polygon areas, but these related to a total of 113 occurrence data (102 points, 11 polygons) (Table 8).
In the European and Middle Eastern region, larval collections were the most common sampling method, with 23 studies sampling at 86 sites. Sampling methods were unknown for a large proportion of the data (1553 sites), of which 1488 related to a single data source [56]. Possibly due to the zoophilic nature of the majority of the European and Middle Eastern species (see below), resting adult females were collected from animal sheds at 85 locations compared to only 31 where resting collections were conducted inside human dwellings. Human landing collections were conducted indoors in only two studies, relating to only three sites, with three studies collecting by outdoor human landing at only eight sites.
Identification methods, amongst those studies that reported them, mainly relied on morphological characteristics and were conducted on specimens from 175 locations. Only 10 studies reported using PCR identification techniques but due to a large number of unknown or unreported methods, this ranked as the second most popular method, and was applied to specimens collected from 67 sites.
Mapping trials
The results for each mapping trial are given in Additional file 2 (Additional file 2: Summary tables showing evaluation statistics for all mapping trials and final BRT environmental and climatic variable selections for the final, optimal predictive maps). Optimal mapping categories were evaluated visually and using the deviance and AUC statistics, with the caveat that these could only be used as a guide rather than a definitive indication of predictive performance.
The EO mapping test indicated that where random pseudo-presences were created within the EO range, and no real occurrence data were included, the model would predict a high probability of presence within the whole EO range and calculate a high deviance value for all species, indicating an overall poor predictive performance. This was the case for the African species and those from the European and Middle Eastern region, and consistent with the results for the nine DVS in the Americas [2]. Where the hybrid method was used that incorporated both real occurrence data plus 500 half-weighted pseudo-presence points randomly assigned within the EO range, the mapping performance was greatly improved. Maps created using only the real presence data produced a low deviance value, but visually, predictive performance was judged to be poor, possibly due to a paucity of data for some species. It was therefore considered that the hybrid maps performed better overall and are presented here.
The optimal buffer width for the African DVS was judged to be 1500 km, producing the lowest deviance value for five out of the seven species. For the European and Middle Eastern species maps, all buffer widths other than 1000 km had high deviance values for all species. The 1000 km buffer therefore was judged to perform better for all six species and applied consistently to all final maps.
For both the African and the European and Middle Eastern species, a ratio of 10:1 pseudo-absences to presence data (not taking into account the 500, half weighted pseudo-presence created in the hybrid maps) was judged to perform better overall, but for both regions, the number of pseudo-absences appeared to have little effect on the predictive maps.
Predictive maps
The BRT maps for all seven African DVS and for the six European and Middle Eastern species are given in Additional file 3 (Additional file 3: Predictive species distribution maps for the seven DVS of Africa and the six DVS of the Europe and Middle Eastern region). Spatial constraints prevent all species being discussed in detail here, however, Anopheles gambiae (Figure 1) is the iconic and possibly the most important vector of malaria [88], and therefore is discussed further below.

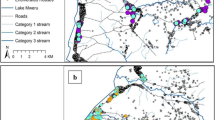
Map details: The predicted distribution of An. gambiae mapped using hybrid data (1443 occurrence data plus 500 pseudo-presences weighted at half that of the occurrence data and randomly selected from within the Expert Opinion (EO) range). Pseudo-absences (14430) were generated at a ratio of 10:1 absence to presence points, and were randomly selected from within the 1500 km buffer surrounding the EO (EO shown in the inset map). Predictions are not shown beyond the buffer boundary. The black dots show the 1443 occurrence records for An. gambiae. Map statistics: Deviance = 0.114, Correlation = 0.9195, Discrimination (AUC) = 0.989, Kappa = 0.9003. Environmental variables: 1. Prec (mean), 2. Prec (max), 3. DEM, 4. Prec (A2) 5. LST (min), (Please see Additional file 2 for abbreviations and definitions). Copyright: Licensed to the Malaria Atlas Project [64] under a Creative Commons Attribution 3.0 License. Citation: Sinka et al. (2010) The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis, Parasites & Vectors 2010, 3:117.
There have been a number of attempts to model the distribution of An. gambiae but the majority tend to focus on single countries and often just map presence points or abundance without further analysis (e.g. [44–49]). Continent-wide predictive maps for An. gambiae (plus other members of the An. gambiae complex) have also been attempted [1, 89], making use of satellite-derived environmental or climatic variables [24, 52–55] (Table 9). The methods range from simply overlaying presence and absence points over rainfall maps [24] to the application of more complex, spatial ecological niche models [53, 55].
Precipitation, in one form or another, is identified repeatedly in previous models (where these data are presented, Table 9) as an influential variable in predicting the range of An. gambiae. Within the top five contributing covariates from the suite applied to the BRT model, precipitation was identified three times, with mean precipitation as the highest contributor with a relative influence of over 37%. Maximum precipitation was placed second (19.42%) with the amplitude of the bi-annual cycle of precipitation ranked forth (8.85%). In common with the Maxent niche model presented by Moffett et al. [55], elevation (altitude) and minimum land surface temperature were also identified by the BRT model within the top five influencing climatic/environmental variables (relative influence of 12.36% and 5.68%, respectively).
Anopheles gambiae larvae are commonly found in temporary, shallow, small bodies of water, such as puddles in hoof prints, wheel ruts and small ground pools (see below), sites which are only present after rainfall. Hence the high influence of precipitation on the distribution of this species identified by the BRT model corresponds closely with the known bionomics of An. gambiae.
The predictive map of An. gambiae (Figure 1) loosely follows the boundary and distribution indicated by the EO map (Figure 1, inset) with one clear exception: the large gap in the range over southern Kenya and a large proportion of northern and central Tanzania. This gap may be driven by the presence of savannah-type vegetation [89] more commonly associated with An. arabiensis, or the increasing altitude of this region, and may be causal to the identification of elevation as an influencing factor to the distribution of An. gambiae. Similar gaps are also seen in the maps produced by Moffett et al. [55] and, to a slightly lesser extent, in the map of Levine et al. [53]. In Madagascar, Léong Pock Tsy et al. [44] identified altitude as a limiting factor for An. gambiae with numbers diminishing as altitude increased until, other than two specimens found at 1300 m, it was considered essentially absent over 1000 m. However, in the Kenyan highlands An. gambiae is commonly identified up to 2000 m [89–92] and specimens have been confirmed at sites up to 1800 m in Uganda [93]. Sampling across Africa, as stated by Coetzee [88], reflects the distribution of entomologists and not necessarily the distribution of the mosquitoes, and the area within this predicted gap, along with a great swath through central Africa, is clearly lacking in empirical occurrence data. Acknowledging these caveats, and similar ones in parts of the range of many of the DVS, it is obvious that samples from these poorly known areas would help improve substantially our predictive mapping.
Bionomics of the African DVS
Anopheles arabiensis
Anopheles arabiensis, when compared to An. gambiae, is described as a zoophilic, exophagic and exophilic species [94]. However, it is also known to have a wide range of feeding and resting patterns, depending on geographical location [11, 95, 96]. This behavioural plasticity allows An. arabiensis to adapt quickly to counter indoor IRS control, where suitable genotypes occur [97], showing behavioural 'avoidance' (deterrence from a sprayed surface) depending on the type of insecticide used [95, 98].
Anopheles arabiensis is considered a species of dry, savannah environments and sparse woodland [11, 24, 97, 99], yet it is known to occur in forested areas, but only where there is a history of recent land disturbance or clearance [24]. Its larval habitats are similar to those of An. gambiae (see below): generally small, temporary, sunlit, clear and shallow fresh water pools [100–103] (Table 10), although An. arabiensis is able to utilize a greater variety of locations than An. gambiae, including slow flowing, partially shaded streams [103–106] and a variety of large and small natural and man-made habitats (Tables 11, 12). It has been found in turbid waters [100, 107, 108] and, on occasion, in brackish habitats [109] (Harbach, unpub. obs.). It readily makes use of irrigated rice fields (Table 11), where larval densities are related to the height of the rice, peaking when the plants are still relatively short and then dropping off substantially as the rice plants mature [110–113]. Such density fluctuations are also reflected in the adult population, which also peak when rice stalks are small and decline as the plants mature [114–116]. These patterns may be due to a preference for sunlit areas of water with relatively limited emergent vegetation (Table 10), with densities decreasing as shade from the growing plants increases. Moreover, there is evidence that An. arabiensis may be attracted by the application of fertilisers or by the amount of dissolved oxygen within the paddy water [111–113, 117, 118]. However, with fertiliser application occurring at the start of plant cultivation, and dissolved oxygen content related to sunlight exposure (e.g. via increasing photosynthesis), the primary oviposition attractant in rice fields is uncertain.
The behavioural variability of An. arabiensis is clearly evident (Table 13), with similar numbers of studies reporting either anthropophilic or zoophilic behaviour. Bøgh et al. [119] stated: 'There is... great variation in the feeding preference depending on the local variation in host availability and composition of the local genotypes of the vector' [95, 96, 120]. Tirados et al. [121] suggested the existence of an east-west behavioural cline. They proposed that those populations found in western Africa display higher levels of anthropophily, and preferentially feed and rest indoors, whereas those in the east exhibit greater zoophily and rest outdoors. Overall, however, biting patterns tend to be exophagic [121–124], but such behaviour is often reported in comparison with highly endophagic species such as An. gambiae. For example, Fontenille et al. [125] reported An. arabiensis as 'more exophagic than An. gambiae and An. funestus' with 65.4% of vectors found biting outdoors identified as An. arabiensis, yet 59% of those found biting indoors were also identified as An. arabiensis.
Blood feeding times also vary in frequency but biting generally occurs during the night. Peak evening biting times can begin in the early evening (19:00) or early morning (03:00) [121, 123, 126–131] (Table 13). This species does, however, demonstrate a predisposition to exophilic (or partial exophilic) behaviour regardless of where it has blood fed or the source of its meal [121, 125, 130, 132–135], a behavioural trait considered to be related to polymorphic chromosomal inversions, to a greater or lesser extent, depending on location [97, 132, 136, 137].
Anopheles funestus
Anopheles funestus is a member of the Funestus Subgroup [138] (often mistakenly referred to as An. funestus complex), which includes: An. aruni, An. confusus, An. funestus, An. parensis and An. vaneedeni. The members of this subgroup exhibit important variation in their biology and behaviour, especially in regard to malaria vectorial capacity and are only morphologically distinguishable during certain stages in their development [10, 11, 18, 139]. Only An. funestus is regarded as an important vector of malaria in this subgroup [18].
A typical An. funestus larval habitat is a large, permanent or semi-permanent body of fresh water with emergent vegetation, such as swamps, large ponds and lake edges. Larvae have been found in shaded and sunlit environments (Table 10) and Gillies & de Meillon [10] concluded that An. funestus uses emergent vegetation as refuge against predation while the shading it casts, or the presence of shade from overhanging plants, is of lesser importance. In some areas, An. funestus larvae, as with An. arabiensis, are associated with rice cultivation (e.g. Madagascar, Mali) [140–144] (Table 11). Where they are found, their favoured environmental conditions are very different to those of An. arabiensis. Anopheles funestus replaces An. arabiensis in a successive temporal process during rice plant growth, exhibiting higher densities in older, maturing fields compared to the preceding open conditions preferred by An. arabiensis [115, 143, 144].
Anopheles funestus is considered to be highly anthropophilic [10, 86, 122, 145–151] (Table 13) (but see below), which led Charlwood et al. [19] to propose that An. funestus may have been the first anopheline species to specialise on biting humans, surmising that its preferred larval sites (permanent water bodies in savannah-like environments) are likely to have been areas where humans first settled. Behaviourally, its late-night biting patterns would also allow ready access to human blood without incurring undue density-dependant host avoidance. This late-night biting preference is clearly evident throughout its range, with all studies reviewed reporting a peak biting period occurring after 22:00, and most commonly between midnight and the early hours of the morning [123, 124, 128, 131, 145, 152–157] (Table 13). Endophilic resting behaviour is also commonly reported [84, 86, 114, 124, 125, 145, 146, 149, 152, 156, 158, 159], and combined with a relatively high longevity, makes it as good a vector, or better in some areas, as An. gambiae [10, 11, 18, 160]. These characteristics are also responsible for promoting the success of vector control using IRS and ITNs. However, this exposure has resulted in selection pressure and rapid development of insecticide resistance to pyrethroids, now well established in some populations and implicated as the primary reason for a major resurgence of epidemic malaria reported in Kwazulu-Natal, South Africa in the late 1990s [18, 161].
Compared to other DVS in Africa, An. funestus shows fairly consistent behaviour (generally anthropophilic and endophilic) throughout its range; however, it is a highly adaptable species, allowing it to occupy and maintain its wide distribution and utilise and conform to the many habitat types and climatic conditions contained therein. Behavioural differences between chromosomal forms have been identified, for example, Lochouarn et al. [162] reported anthropophilic behaviour in western Senegal and zoophilic behaviour in the east of the country, behaviours which correspond to chromosomal polymorphisms that also follow this east-west cline. Costantini et al. [60] identified two chromosomal forms in Burkina Faso associated with different resting and biting behaviour. This, coupled with a lack of heterokaryotypes in areas where the two forms co-exist, prompted these authors to suggest that the two forms were incipient species, and hence of the concept of an An. funestus complex. More recently, An. funestus populations from 12 countries have been divided into three molecular types: M, W, and MW, correlating to geographical locations, whereby M is essentially found in eastern Africa, W from western and central Africa, and MW from southern Africa [61]. Further investigations showed a more complicated situation with specimens from Malawi showing all three types, specimens from Tanzania showed the M- and MW-types, whereas specimens from Kenya showed M- and W-types. In addition, two more types were described, type Y from Malawi, and type Z from four localities of Angola, Malawi, Ghana and Zambia [62]. Finally, adding further to the complexity surrounding this species, recent studies in Malawi have revealed a new species of the subgroup, named An. funestus-like [63] that is identical to An. funestus but appears to have a different biology and role in malaria transmission, although this needs confirmation.
Anopheles gambiae
Anopheles gambiae is considered to be one of the most efficient vectors of malaria in the world and is one of the most well studied [88]. Like An. funestus, the variable ecological conditions present within the large geographical range of An. gambiae indicate a highly plastic species with corresponding chromosomal diversity currently separated into five chromosomal forms: Forest, Bamako, Savanna, Mopti and Bissau [163]. There is suggestion of reproductive isolation among the sympatric forms, and hence, of incipient speciation between them [163–165]. Independent of these chromosomal categories, two molecular forms, 'M' and 'S', have also been described [165], and are the forms more commonly referred to in the recent literature. These different forms exhibit ecological adaptations which further indicate possible speciation, for example the Mopti and M forms are associated with semi-permanent, often man-made, larval habitats such as rice fields or flooded areas, whereas the Savanna/Bamako and S forms are seen more commonly in temporary, rain-dependent sites such as ground puddles [166–171]. There appear to be no definitive studies that explicitly describe variability in adult biting or resting behaviour or role in malaria transmission between the two molecular forms.
Despite its wide range and variable ecology, a combination of traits allows An. gambiae to maintain its position as one of the most efficient vectors in sub-Saharan Africa. It is a relatively long-lived species (although not as long as An. funestus [160]) [172, 173], with a short larval development period and is often found in larval habitats associated with human activity (e.g. water in hoof prints, wheel ruts or areas of rice cultivation) (Tables 11, 12). It is considered to be highly anthropophilic, with 11 of 15 studies that examined biting behaviour (Table 13) reporting a marked preference for human hosts [131, 145, 149, 150, 157, 159, 174–177]. However, there are a number of studies that indicate An. gambiae is less discriminant and more opportunistic in its host selection and that host choice is, as with the majority of African DVS, highly influenced by location, host availability and the genetic make-up of the mosquito population. Moreover, many studies that report host preference using blood meal analysis are often conducted on resting, blood-fed specimens collected inside houses, thus introducing a potential study design or sampling bias favouring the likelihood that the blood meal will be from a human host [178]. Of the studies that report some level of zoophily, Diatta et al. [178] specifically examined the host preference of An. gambiae and An. arabiensis by comparing the number of females of each species captured either in a calf-baited or a human-baited net trap. There was no statistical difference between the host preferences of the two species, both expressing greater zoophily (e.g. 31% of An. gambiae were found in the human-baited trap and 69% in the calf-baited trap). Duchemin et al. [122] also reported zoophilic behaviour, yet highlighted this as unusual, suggesting that the high density of cattle in the sampling area may have influenced the propensity for zoophily in the population. Bøgh et al. [119] reported no specific preference for either human or animal hosts but that An. gambiae would feed readily on cattle.
As with An. arabiensis, An. gambiae larvae typically inhabit sunlit, shallow, temporary bodies of fresh water such as ground depressions, puddles, pools and hoof prints (although see above) [91, 101, 175, 179–183] (Table 10, 12). Gillies & de Meillon [10] suggested that this aspect of their bionomics allow members of the An. gambiae complex to avoid most predators, and the larvae are able to develop very quickly (~six days from egg to adult under optimal conditions and temperatures), possibly in response to the ephemeral nature of their larval habitats. Water in these larval sites can appear clear, turbid or polluted [101, 180, 184–186] (Table 10). Typically An. gambiae larval habitats are described as containing no (or very sparse) vegetation (Mbogo, unpub. obs.) due to their temporary nature. Gillies & de Meillon [10] summarised the great diversity of habitats utilised by An. gambiae, and as described before, different molecular or chromosomal forms are associated with either vegetated (e.g. rice fields) or temporary and non-vegetated (e.g. hoof prints) larval sites [101]. The studies reviewed here report An. gambiae from habitats containing floating and submerged algae, emergent grass, rice, or 'short plants' in roadside ditches and from sites devoid of any vegetation [91, 101, 109, 180, 181, 183] (Table 10).
Females of An. gambiae typically feed late at night, a characteristic shared with An. funestus that may increase their ability to effectively transmit malaria parasites (see above) [19, 123, 127, 145, 153, 154, 157, 175, 177, 185, 187–190] (Table 13). Anopheles gambiae is often described as an endophagic and endophilic species, both biting and resting indoors, however, the majority of studies listed herein (nine of 11), that compared indoor and outdoor human-landing catches reported no difference in the numbers of females collected at either location [123, 127, 145, 149, 153, 157, 175, 190, 191] and an equal number of studies recorded post-feeding exophilic resting [122, 131, 154, 175] as resting indoors [145, 149, 159, 178]. Bockarie et al. [175] linked differences in the exo-or endophilic behaviour of An. gambiae to their chromosomal forms, suggesting the Forest form (with no inversion) demonstrated stronger exophily in southern Sierra Leone whereas the Savannah form, with a 2La inversion, was mostly endophilic. Odiere et al. [192] used clay pots to sample outdoor resting females in western Kenya and found no clear preference for indoor or outdoor resting. They suggested that the designation of An. gambiae as a predominantly endophilic species may have been based on poor sampling comparisons. As with host preference, this species appears to exhibit greater phenotypic plasticity and opportunism in blood feeding and resting locations than commonly thought.
Anopheles melas
There is relatively little contemporary information about the behaviour of An. melas, perhaps because it is generally considered to be a vector of lesser importance, specifically where it occurs in sympatry with An. gambiae or An. arabiensis. Anopheles melas has a comparably lower sporozoite rate than either An. arabiensis or An. gambiae (e.g. 0.35% compared to 3.5% for An. gambiae in The Gambia) [13, 95, 193], yet in coastal areas where it can occur in very high densities it is still a problematic vector of malaria [13]. With the dearth of available contemporary data, those studies conducted prior to 1985 that closely examined the behaviour of this species have been included here.
Anopheles melas is commonly associated with brackish water and can utilise saline environments that other species, for example, An. gambiae, cannot tolerate [109, 171], yet does not appear to require brackish water for larval stage development [194–196]. It is generally restricted to coastal areas [194–197] but has been found up to 150 km inland along the Gambia River, where salt water can intrude great distances (up to 180 km) upriver [109, 171, 193]. Unlike other African DVS, the density fluctuations of An. melas are closely associated with tidal changes rather than seasons, for example, Gelfand [194] identified a peak in adult numbers 11 days after spring tides. The larvae of this species are associated with salt marsh grass (Paspalum spp.) and mangroves, but only trees of the genus Avicenna, which include white, grey and black mangrove, and not those from the genus Rhizophora ('true' or red mangrove spp.) [109, 194, 195, 197]. These positive and negative associations with mangroves are thought to be strongly influenced by the predominant soil type associated with the different tree genera. Anopheles melas preferentially oviposits on damp ground at low tide, rather than in open water, where the eggs are able to survive some degree of desiccation [196] until the tides rise again, and appears to prefer the poorly drained, peaty-like soil common to Avicenna forests compared to the sandy, gravelly or smooth, fibrous peat soils common to the Rhizophora stands [195, 198]. Giglioli [198] surmised, that this behaviour guarantees the larvae will have sufficient time to complete their larval development and pupate in the less saline, relatively permanent waters of the new tide before it begins to recede and the water either becomes too salty, or dries out completely.
Adult biting behaviour appears to be opportunistic. Anopheles melas has been described as both highly anthropophilic and a zoophilic species [193, 194, 197, 199, 200]. In a choice experiment, Muirhead-Thomson [197] varied the numbers of animal and human baits in traps to attempt to describe host preference. He found An. melas to be fairly indiscriminate: where there were more animal baits, An. melas would feed more often on animals, but still feed on humans. On the contrary, where there was an increase in the number of human hosts, a sharp decrease in the number of females feeding on animals occurred. Sampling bias towards anthropophily may be reported when blood fed females collected resting inside houses are tested for host blood type because An. melas generally appears to rest outdoors after feeding [193, 194, 197], although there has been limited success in locating and collecting from such natural outdoor resting sites. As previously described for An. gambiae, those females that bite and rest indoors are more likely to have fed on humans, and those biting or resting outdoors (or in animal sheds) are more likely to have bitten animals. Blood feeding activity appears to be fairly continuous throughout the night [194, 197, 200]. Gefland [194] observed continual biting from 19:00 to dawn, although Muirhead-Thomson [197] saw two peaks of biting activity: the first, and slightly smaller peak, between midnight and 02:00 and a second, larger peak, between 04:00 and dawn.
Anopheles merus
Anopheles merus has previously been considered as only a minor, or even an unimportant vector, potentially unable to sustain malaria transmission alone [95]. However, is has been identified as playing an 'unexpectedly important role' along the Tanzanian coast [14] and more recently in Mozambique [15]. It is also a species for which there is limited contemporary information. The differences in egg and larval morphology that distinguish An. melas from An. gambiae do not occur in An. merus and identification, before the advent of molecular techniques, was based on physiological characteristics involving larval salinity tolerance tests [201]. Originally, An. merus was referred to as a 'salt water An. gambiae' variant or subspecies. Indeed, Jepson et al. [202] had a number of specimens collected in the 1940s from saline, coastal swamps in Mauritius examined for distinguishing features, and found no obvious morphological distinguishing characters and stated 'All the specimens proved to be typical forms [of An. gambiae] and there was no evidence of the presence of An. gambiae var. melas'. They continued to regard 'An. gambiae' as a species with 'a considerable tolerance for pollution and salinity and is therefore to be found in domestic wastes and in crab holes and pools near the sea side, in addition to a host of natural breeding places such as marshes, rock pools and casual rainwater pools'. This Mauritian species was finally designated a subspecies of An. gambiae by Halcrow [203], who provisionally named it An. gambiae litoralis based on larvae found in '...water of high salinity in crab holes, depressions in coralline rocks, small tidal lagoons, pools close to tidal zone and [interior] salt pans, and are not associated with mangroves...' [203, 204]. Paterson [205] provided definitive proof of the specific status of An. merus and the validity of the name [206].
Halcrow's [203] description highlights a specific difference between An. merus and An. melas. Anopheles merus is rarely found in the mangrove forests on the east coast, however this may be due to the composition of the trees and soil type under of the stands of mangrove in this zone rather than inherent behavioural differences between the two species [10]. Anopheles merus is, instead, found in high numbers in shallow brackish pools and marsh or swamp areas along the coast. As a consequence, this species does not exhibit density changes in response to the tidal fluctuations as seen with An. melas, nor does it appear to tolerate the same high levels of salinity [201, 207]. Anopheles merus is also known to occur further inland, using salt pans and saline pools larval habitats [11, 208–211], and cross-mating experiments between inland and coastal populations have produced viable offspring indicating they are conspecific [212].
The biting behaviour of An. merus is similar to that of An. melas: generally opportunistic in host selection, depending on host availability [203, 213] and with a tendency to bite [207, 214] and rest outdoors [201, 206, 213, 214]. Gillies & de Meillon [10] suggested that An. merus shows a preference for animal hosts, referring to a laboratory test where, given a choice, females consistently fed on calf versus human bait. Two of the studies reviewed here reported anthropophily [150, 214], one indicated zoophily [203] and another concluded that no obvious preference was detected [213]. In the latter study, blood meal analysis was conducted on mosquitoes collected resting indoors (59.2% had fed on humans), and those collected resting outdoors (71.4% had fed on cattle and only 1.6% contained human blood) [213], highlighting the bias in drawing conclusions on host preference if only indoor or outdoor resting specimens are tested. Only one study, conducted on the Kenyan coast, examined the biting times of An. merus [214], which reported the number of bites gradually rising from early evening (18:00) peaking between midnight and 01:00 and then declining to 06:00 which corresponds to the accepted biting pattern for this species across its range (Bangs and Mbogo, unpub. obs.).
Anopheles moucheti
Anopheles moucheti is a species with two morphological forms: An. moucheti moucheti, and An. m. nigeriensis which are distinguishable by morphological features of the adult and larval stages [10]. Anopheles m. bervoetsi, previously considered a third morphological form, has recently been raised to full species status: An. bervoetsi by Antonio-Nkondjio et al. [215]. However, these authors do assert a level of caution in this new status as they point out that An. bervoetsi has only ever been reported from its type locality (Tsakalakuku, DRC) and has never been found in sympatry with An. moucheti. They do cite unpublished data that detected P. falciparum infection in An. bervoetsi specimens, and thus raises the possibility that this species could be transmitting malaria in central Africa [215]. The bionomic information detailed here is, in the most part, taken from sources that present data for 'An. moucheti'. Of these, the majority of studies have been conducted in Cameroon by Antonio-Nkondjio and colleagues or in Nigeria, so based on current knowledge the assumption is that these data refer to An. moucheti and not An. bervoetsi.
Despite its status as a DVS, An. moucheti is a poorly studied species. It is the only DVS with its range entirely restricted to forested areas [216], specifically where the canopy is broken allowing sunlight to penetrate to the ground, such as is found where large rivers flow through the forest [10]. Human activity, such as road building, settlements or cultivation, can therefore be beneficial to this species by breaking up the forest canopy, although larger areas of deforestation may decrease the density of An. moucheti and allow replacement by An. gambiae [217, 218]. Anopheles moucheti larvae are found at the edges of large, slow flowing or lentic rivers, often with turbid waters, and are associated with Pistia spp (water lettuce/water cabbage) [89, 217, 219]. Antonio-Nkondjio et al. [217] studied the larval habitats along the river networks of southern Cameroon and found the greatest numbers of An. moucheti larvae along the margins of rivers within deep, evergreen forest, substantially fewer in the degraded forest and none in the savannah areas. Where they were found, larvae were abundant near to areas of human habitation.
Although the range of An. moucheti is relatively restricted within the equatorial forests, it derives its status as a DVS from its highly anthropophilic and endophilic behaviour (Table 13) [86, 145, 174, 219, 220]. Gillies & de Meillon [10] suggested such behaviour is unsurprising due to the lack of domestic animals found within forested environments. Anopheles moucheti is also described as highly endophagic, however this characteristic appears to be less than clear cut. For example, Antonio-Nkondjio et al. [220] found that in urbanised, forested environments (where An. moucheti was less abundant and replaced by An. gambiae) compared to rural localities (where An. moucheti was dominant), only 43% of females were found biting indoors, whereas in the rural areas 66% were found biting indoors. In a study conducted in a village only 2 km from Yaounde, Cameroon, Antonio-Nkondjio et al. [145] reported 51% biting indoors and described the sampled populations as 'mainly endophagic'. Overall, An. moucheti appears endophilic [86, 145] (Table 13). In a countrywide survey of Cameroon, of all females found resting, 1234 were located indoors, whereas only 12 were captured in outdoor shelters [86]. Only two studies examined the biting cycle of An. moucheti, with both reporting biting gradually increasing towards the second half of the night to dawn [145, 221]; Mattingly [221] reported peak biting activity in the early morning between 03:15 and 06:15.
Anopheles nili complex
The An. nili complex includes An. carnevalei, An. nili, An. ovengensis and An. somalicus [12]. As with An. moucheti, species of this complex have been generally overlooked in African vector studies despite being described as highly efficient vectors [6, 89, 222, 223]. Amongst members of the complex, An. nili is considered the most important vector, although An. carnevalei and An. ovengensis are implicated as secondary vectors of P. falciparum in Cameroon [86, 224]. Anopheles somalicus is considered zoo- and exophilic [6, 10]: it was not found to bite humans in Somalia [10] and no females were found in houses in Cameroon despite an abundance of larvae in the area [6].
Larvae of all members of the An. nili complex are found in vegetation at the edges of fast flowing streams and rivers [10, 89, 195, 217]. However, An. ovengensis and An. carnevalei appear to be restricted to areas of deep forest, whereas An. nili is more abundant along rivers in degraded forest and savannah [217]. A comprehensive survey of the river systems across Cameroon found An. nili larvae associated with sunlit sites whereas An. carnevalei larvae were more commonly found in shaded areas [217].
Anopheles nili is considered to be strongly anthropophilic [10, 86, 145, 148, 225–227], and will readily bite both indoors and out [145, 149, 226, 228] (Table 13). Carnevale & Zoulani [226] described biting patterns that exploited the behaviour of their human hosts, biting outdoors in the early evening when people are socialising, and then continuing to bite indoors once people move inside, with peak feeding occurring after midnight [145]. The resting habits of An. nili are also described as 'variable' [10]. Krafsur [227], in a lowland region of western Ethiopia, rarely found An. nili resting indoors despite the high densities found biting indoors, indicative of exophilic behaviour. Conversely, Antonio-Nkondjio et al. [86] examined populations across Cameroon and reported An. nili overwhelmingly resting indoors (466 females), with only one female captured in an outdoor shelter. In the same study they found no An. carnevalei females resting indoors or in outdoor shelters whereas all resting An. ovengensis captured were found indoors. Conversely, Awono-Ambene et al. [224] stated that An. ovengensis was rarely found resting indoors and concluded it had 'exophilic habits'.
Bionomics of the European and Middle Eastern DVS
Anopheles atroparvus
Anopheles atroparvus is a member of the Maculipennis Subgroup, which also includes An. (Ano.) daciae, An. (Ano.) labranchiae, An. (Ano.) maculipennis, An. (Ano.) martinius, An. (Ano.) melanoon, An. (Ano.) messeae, An. (Ano.) persiensis and An. (Ano.) sacharovi [12]. Of these, An. labranchiae, An. messeae and An. sacharovi are also designated as DVS (see below).
Anopheles atroparvus is described as a species with a preference for brackish larval habitats [229–232]. Hackett & Missiroli [231] summarised: 'In general it may be said that over its extensive range [An.] atroparvus is found in water of moderate salinity not exceeding 10 parts per 1000. It prefers relatively cool water, and its range does not overlap that of [An.] labranchiae, a warm water breeder'. However, the larval sites listed in the literature still include a number of predominantly fresh water habitats, for example canals, ditches, river margins, pools in river beds and rice fields [230], and Cambournac [233] defines An. atroparvus as a 'fresh water breeder'. Hackett [234] also stated that, in southern Europe, An. atroparvus 'inclines to breed in fresh water'. Of the few studies reporting primary data (Tables 14-17), larvae were identified in marshes and ditches/ground flood pools [235], pools in river beds, river margins and streams, rock pools, cement tanks, rice fields, wells and ground pools [229] and in small collections of water in used tyres [236] (Tables 15, 16).
Becker et al. [230] described sites to be 'usually sun exposed' and to contain 'a considerable amount of filamentous green algae and other floating submerged vegetation'. Pires et al. [229], in a study that sampled comprehensively across Portugal, reported An. atroparvus larvae to be found more frequently in sun-exposed habitats, although 'some shade was provided by grasses and aquatic vegetation'. They also reported filamentous algae present in 48 of 93 sites positive for An. atroparvus (Table 14).
Anopheles atroparvus is generally considered zoophilic [229, 230, 237], and described as 'very zoophilic' by Cambournac [233], who also stated that its hosts, in order of preference, are rabbit, horse, cow, pig and sheep, and suggested that a long association between rabbit and An. atroparvus (since approx. 1000 BC) may be responsible for this hierarchy of preference. Indeed, An. atroparvus has been implicated as an effective vector of the myxomatosis virus to domestic rabbits in the UK [238, 239]. Elsewhere, however, An. atroparvus is described as anthropophilic [89], which perhaps indicates the opportunistic nature of this species. Four studies identify An. atroparvus as zoophilic [229, 237, 240, 241] and one study, that did not distinguish a preference, reported the collection of An. atroparvus during night catches on horse bait, from indoor resting sites and during day- or night-time catches on humans [235] (Table 17). There is no clear evidence or information among any of the published studies, nor within the general literature, that identifies this species as preferentially biting indoors or outdoors. The opportunistic nature of its feeding habits and zoophilic proclivity in host choice, however, would suggest it is probably exophagic but that biting location could also depend upon the setting and accessibility of the host.
Anopheles atroparvus rests and hibernates in animal sheds and stables [229, 230, 235, 237, 238, 240, 241]. It hibernates as an adult female and is known to periodically feed, specifically if she has taken refuge in a relatively warm locality, but these meals do not result in egg production (i.e. gonotrophic disassociation) [230–232].
A number of investigators have discussed the inability of An. atroparvus to transmit tropical strains of P. falciparum, with most referring to studies conducted by Shute [242]. Unfortunately this reference could not be found, but in a study testing the susceptibility of Russian anopheline species to imported P. falciparum [40], no infection was detected in An. atroparvus females. Curtis & White [243] concluded (also referring to Shute [242]) that An. atroparvus is refractory to both Asian and African P. falciparum but competent in supporting a European strain, a conclusion reiterated by de Zulueta et al. [39] with Cambournac [233] stating that refractoriness of An. atroparvus to African and eastern strains of P. falciparum is an 'uncontroversial fact'. However, Capinha et al. [244] claimed the existence of local An. atroparvus in Portugal that could be infected with 'exotic strains of plasmodia', with reference to a comprehensive study by Souza [245]. However, on closer examination of these findings, even though Sousa did indeed infect An. atroparvus with P. falciparum, this was only after numerous attempts that resulted in formation of oocysts in only five out of 736 females. It would seem, therefore, that although An. atroparvus can be infected by tropical P. falciparum strains, it is very unlikely to happen under natural conditions and there is currently no conclusive evidence that such infection would result in salivary gland invasion by sporozoites.
Anopheles labranchiae
Despite similarity in larval site characteristics, An. labranchiae and An. atroparvus do not, or only have limited, overlap in their distributions [231]. This lack of sympatry may be simply a factor of temperature, with An. labranchiae making use of warmer waters than typical of An. atroparvus [230, 231]. However, when Capinha et al. [244] modelled the habitat suitability of An. atroparvus across Portugal, they concluded that the most suitable locations include drier areas with higher temperatures (i.e. conditions where An. labranchiae typically dominate), whereas wetter areas with milder temperatures, where An. atroparvus are mostly found, were unsuitable. They concluded that An. atroparvus is not found in many other 'suitable' Mediterranean areas due to competitive exclusion. Conversely, de Zulueta [246] suggested that the absence of An. atroparvus in Sardinia allowed the wide distribution of An. labranchiae on the island, where, despite a five-year eradication campaign instigated in 1946, An. labranchiae still occurs [247, 248].
Both species utilise brackish water marshes and lagoons along the coast [231], although in contrast to An. atroparvus, An. labranchiae will preferentially oviposit in fresh water [89, 247, 249–251]. Marchi & Munstermann [247], in a survey conducted across Sardinia, only identified An. labranchiae in fresh water sites, including rock holes, pits, ditches, drains or canals, streams/rivers, flooded ground pools and ponds, lakes or reservoirs. Despite an ability to tolerate some salinity, An. labranchiae larvae are not generally found at sites with significant levels of organic or mineral pollutants ([252], Mouchet, pers. com.). Larval sites are typically described as sunlit [89, 230, 249, 253], although in Sardinia Aitken [251] found larvae in 'almost every type of habitat except the very densely shaded', and Macdonald [250] also associated this species with habitats that have some level of shade. In general, An. labranchiae larvae are found in stagnant or slow moving waters [230, 249] and can make use of, and become very abundant in, rice fields [89, 253–256]. Indeed, Bettini et al. [254] described a survey in central Italy that identified high numbers of larvae in newly established (two years old) rice fields with correspondingly high numbers of adults found resting in animal shelters near these fields.
Female An. labranchiae can aggressively attack human hosts [230, 255], and are described as 'persistent' in their attempt to enter bedrooms during the night [230]. Nonetheless, this species is also described as zoophilic in some of the general literature, but overall, An. labranchiae appears opportunistic in its host choice, readily biting either humans or animals (Table 17) [89, 249, 250, 253, 255–257]. Romi et al. [256] found high percentages (86% and 90.7%) of females engorged with human blood resting inside houses whereas they also found that almost all specimens collected resting in animal shelters had fed on animals.
Anopheles labranchiae rests inside houses, animal shelters, and, to some degree, in natural shelters, depending on the location of its blood source [249, 253–259]. D'Alessandro et al. [249] described An. labranchiae as both endo- and exophilic, using whatever shelters are available. Females hibernate in stables/animal shelters and in natural sites such as crevices and tree cavities. Both incomplete (with occasional blood feeding but without ovipositioning) and complete (with fat bodies, without feeding and non-gonoactive) hibernation have been noted for this species [89, 230, 231, 249].
As with An. atroparvus, An. labranchiae has been found to be refractory to exotic strains of P. falciparum, with de Zulueta et al. [39] failing to infect An. labranchiae, albeit a small sample, with a Kenyan strain of P. falciparum. However, Toty et al. [58] reported historical evidence of naturally infected An. labranchiae, plus the results of a contemporary study conducted by the Centre de Production et d'Infection d'Anophèles (CEPIA) in Paris where 14% (13/99 specimens) of Corsican An. labranchiae were experimentally infected with the African NF54 laboratory-cultured strain of P. falciparum. This study also detected sporozoites in the salivary glands of three specimens, indicating that An. labranchiae is not only susceptible but also potentially able to transmit at least some strains of African P. falciparum [58]. However, this conclusion must only be considered alongside the knowledge that the NF54 P. falciparum strain is a highly attenuated, long-standing laboratory culture which may no longer reflect its origins (Bangs, unpub. obs.).
Anopheles messeae
Anopheles messeae is the third member of the Maculipennis Subgroup [12] to be designated as a DVS. It is the most widespread species of the subgroup [230], with a distribution extending from Ireland across Europe and Asia and into China and Russia [260]. A great deal of work on this species has been conducted in Russia and China. This review is therefore presented with the caveat that there may be details and data reported in the Chinese or Russian literature that are not included here due to access difficulties.
Di Luca et al. [261] identified a number of genetic polymorphisms within An. messeae and defined five separate haplotypes associated with different geographical areas across its distribution. However, they could not confirm whether these polymorphisms were indicative of altered behaviour at these different locations, although the large range of this species combined with such genetic variability would suggest that some area-specific biological or behavioural adaptations are likely to have occurred.
The larvae of An. messeae are typically found in shaded, clear, very slow flowing or stagnant, fresh water sites [230, 262–264] such as lake margins and marshes [263–265]. Despite only sampling resting females, Adamovic, in Serbia and Montenegro [266–268] and Adamovic & Paulus [269], in surveys of Slovenia and Croatia, continually associated the presence of adult An. messeae with stagnant, fresh water oxbow swamps and marshes within alluvial plains or valleys of large river systems and at sites near large lakes. Localities along rivers with saline or alkaline soil did not provide the same association [266, 268], however they did report the presence of An. messeae at sites near a marshy plain with brackish water [267]. Takken et al. [263] also inferred the presence of An. messeae in more brackish habitats, presenting a photograph in their paper of a drainage ditch labelled as containing brackish water and vegetation which 'supports Anopheles messeae'. However, they also indicated that engineering works in the Netherlands allowed the transition of brackish sites to fresh water, so whether or not they did find this species in brackish water is still unclear. Nonetheless, Takken et al. [263] did identify locations where An. messeae larvae were collected, including sites containing reeds, and those containing floating aquatic weeds and algae, relatively open ditches inside forests and clear water in small lakes within dunes.
Only one study could be found that sampled An. messeae females inside human habitations, animal shelters and in natural outdoor shelters [270]. All other studies only searched in animals shelters [240, 263, 266–269, 271–273]. Where comparisons were made, no An. messeae were found resting outdoors in urban areas (e.g. in vegetation surrounding buildings) but were found indoors such as in entryways, staircases and basements, although not in large numbers. In rural areas, An. messeae dominated the collections made from cattle sheds, with few specimens collected from natural outdoor sites (hollows, ground cavities, amongst vegetation surrounding marshes, ponds and streams) [270].
Takken et al. [263] argued that in the Netherlands An. messeae has never been considered as a malaria vector despite it being as susceptible to P. vivax infection as An. atroparvus [40]. They stated that its high degree of zoophily and outdoor feeding behaviour makes the likelihood of it being involved in local malaria transmission very remote. They supported their argument reporting that all resting An. messeae females collected in their study had fed on animals; however, all their samples were collected from animal shelters. Bates [265] also mentioned that in Albania, An. messeae (along with other members of the Maculipennis Group) is not considered a malaria vector using the same reasoning: '[An. maculipennis, An. messeae and An. melanoon (as An. subalpinus)] are generally supposed not to be malaria vectors because of their non-anthropophilous [sic] food habits'. Fyodorova et al. [270] found that 40% of the An. messeae females collected in urban areas contained human blood, with the remaining 60% having fed on cats (40%) and chickens (20%). However, in rural areas, no human blood meal was detected. Becker et al. [230] summed up the biting preferences of An. messeae somewhat ambiguously, stating that 'Blood-meals are taken from humans only when the density of An. messeae is very high and there is a shortage of livestock, but they also may attack humans in houses'. No studies were found that examined the feeding cycle of An. messeae.
Anopheles messeae, like An. atroparvus and An. labranchiae, hibernates as an adult female. However, unlike these other two species, An. messeae chooses hibernation sites in abandoned buildings, in the absence of animals [230, 271]. They enter full diapause, and do not feed during the winter, but instead, gain energy from fat reserves [271].
There is some evidence to suggest that, along with An. atroparvus, An. messeae may also be refractory (or essentially refractory) to tropical P. falciparum strains. In their study, testing the susceptibility of Russian anophelines to imported P. falciparum, Daškova & Rasnicyn [40] were unable to infect An. messeae. Indeed, the vector status of An. messeae has come into question, specifically since the discovery of a new species in 2004, formally named An. daciae, in Romania [273], which has since been recorded from south-western England [274]. Anopheles daciae can only be distinguished from An. messeae using egg morphology or by sequencing the internal transcribed spacer 2 (ITS2) of ribosomal DNA and the cytochrome oxidase 1 (COI) of mitochondrial DNA. It has been suggested that the presence of An. daciae, potentially sympatric across the full range of An. messeae, may be responsible for the high polymorphism previously reported for An. messeae [261, 274]. Combined with an ongoing debate about the capacity of An. messeae to transmit malaria (e.g. it is not considered a vector in northwestern Europe [275]), it is feasible that An. daciae, and not An. messeae, could be involved in malaria transmission, which will only be confirmed with further investigation into the epidemiological importance of each respective species (Harbach, unpub. obs.).
Anopheles sacharovi
Anopheles sacharovi is the final member of the Maculipennis Subgroup defined as a DVS and has been the target of a number of focussed, anti-vector campaigns across its range including Israel, Greece and Turkey [262, 276–279], yet this species still persists in all areas. Anopheles sacharovi is highly plastic in both adult behaviour and its choice of larval habitats. Zahar [262] states simply: '[An. sacharovi] breeds in all small water collections containing aquatic vegetation'. It makes use of fresh water habitats but is also described as more tolerant of salinity (up to 20%) than any other member of the Maculipennis Subgroup [230, 262]. It can survive in waters up to 38-40°C ([280] references within), and although it is generally considered to breed in stagnant waters, it can also cope with some, albeit weak, current [281, 282]. Throughout the literature there is general agreement that this species prefers sunlit sites with plenty of emergent and/or floating vegetation [89, 230, 262, 283–285]. A typical habitat would be an area of swamp or marsh [265, 279, 282], but larvae are also found at margins of rivers, streams and springs [281, 282], seepages [281], wadis [286], pools and ditches [265, 287]. It is associated with rice cultivation and other irrigated areas, specifically where irrigation channels are poorly constructed causing leakage, creating boggy areas or standing water [89, 230, 277, 279, 282, 284, 288, 289]. Despite its apparent adaptability, An. sacharovi cannot tolerate organic pollutants [262, 285]. Indeed, Saliternik [285] lists the organic pollution of streambed habitats, previously densely populated with An. sacharovi larvae, of greater impact than the wide-scale IRS application of DDT as causal to the near elimination of this species in Israel in the 1960s.
Anopheles sacharovi females feed opportunistically, despite being generally considered as anthropophilic [89, 230]. Only one study reviewed specifically tested host preference. Demirhan & Kasap [290], using baited feeding rooms, concluded that in the presence of other, equally available hosts (human, cow, sheep, chicken, horse and donkey), An. sacharovi preferentially fed on donkeys, and had a negative preference for humans. They also analysed the blood meals of engorged females from human habitations, animal shelters and abandoned or ruined buildings and reported the 'feeding preference' of females captured in the human dwellings to be cow, human, sheep, horse and chicken. Other studies reported similar results. Yaghoobi-Ershadi et al. [291] found high numbers of females collected from cow sheds or chicken coops had fed on animals (85.6 - 92.5%), whereas of those collected from bedrooms, only 38.5% had fed on humans, 38.5% on other animals and 23% on both. Boreham & Garrett-Jones [292] reported predominantly human blood in specimens collected from houses, predominantly animal blood (sheep or goat) from animal shelters, and those collected from outdoor pit shelters generally contained blood of mixed animal origin (sheep, goat, horse, dog or cow). Hadjinicolaou & Betzios [276] reported a high percentage of females containing human blood from human habitations, whereas females taken in pit shelters and animal sheds had mostly fed on domesticated animals. They concluded that An. sacharovi still exhibited significant levels of anthropophily despite a high ratio of animals to people (between 9:1 and 7.2:1) in the study area. Boreham & Garrett-Jones [292] suggested that An. sacharovi had increased tendencies towards zoophilic behaviour due to previous DDT spraying campaigns, but was reverting back to anthropophily.
Anopheles sacharovi, contrary to the accepted night-time biting habits of most anophelines, can 'in deeply shaded situations... attack viciously throughout the day' [289]. However, Djadid et al. [293], indicated that An. sacharovi (plus other members of the Maculipennis Subgroup) generally start biting in the early evening, peaking between 20:00 or 21:00 (refers to Djadid MSc thesis), with Alten et al. [294] noting higher densities of An. sacharovi between 20:00 and 22:00, although they did not specifically examine biting behaviour. Hadjinicolaou & Betzios [276] observed An. sacharovi to bite indoors and outdoors. Biting location is likely to be driven by host behaviour, for example, in the hotter parts of Turkey where both people and animals spend the night outdoors, biting would tend towards exophagy [289, 294].
Anopheles sacharovi is principally described as endophilic [89, 230, 284]. Its choice of resting location (and, arguably, for all species) is most likely driven by the need to find the most suitable microclimate for increased survival [289]. Demirhan & Kasap [290] observed An. sacharovi feeding on cows outside, and then entering houses or abandoned shelters to rest. Yaghoobi-Ershadi et al. [291] found An. sacharovi in cow sheds, chicken coops and bedrooms, but were unable to find any females resting outdoors. Boreham & Garrett-Jones [292] searched two artificial pit shelters and found 10 and 42 specimens compared to 377 and 333 from two cattle sheds and 260 in a village house. Abdel-Malek [282] failed to find An. sacharovi resting outdoors, but again, repeatedly found them resting in animal stables and human habitations. However, insecticide residual spraying in many areas has apparently affected endophilic behaviour [283, 295, 296], summed up by Gokberk [283]: 'Following the last ten years of DDT spraying, An. sacharovi recently began to show a tendency to be less domestic in habits'. Yet, there is evidence that once these IRS programmes ceased, An. sacharovi began to revert to more typical endophilic behavioural patterns [276].
As with other European or Middle East DVS that occur in warmer climates, hibernation is incomplete, with intermittent feeding during winter, but without oviposition [89, 297], often making use of the same localities chosen for resting in the summer months [288].
Anopheles sergentii
There is some confusion as to the taxonomic status of An. sergentii. It has previously been considered to have two geographical forms: An. sergentii sergentii and An. sergentii macmahoni, but in accordance with the published literature, the Walter Reed Biosystematics Unit (WRBU) online catalogue of the Culicidae [298] and the Mosquito Taxonomic Inventory [299], An. macmahoni is currently considered a subspecies of An. sergentii. This subspecies has never been found biting humans and is of no known medical importance [10, 300].
Anopheles sergentii is known as the 'oasis vector' or the 'desert malaria vector' due to its distribution within oases across the Saharan belt in northern Africa into the Middle East, and its ability to cope with the extreme climate across this region [301, 302]. It may be able to survive in such harsh conditions due to its adaptability. It makes use of a range of larval habitats, including streams, seepages, canals, irrigation channels, springs, rice fields [103, 230, 262, 286, 300, 303–308], and most other non-polluted, shallow sites that contain fresh water with a slow current, slight shade and emergent vegetation or algae (Table 14) [89, 103, 230, 262, 302, 306, 308]. However, larvae have also been found in moderately brackish habitats, areas of stagnant water, light to moderately polluted locations or in sites in full sunlight [303–305, 307]. In general, the presence of vegetation or algae seems to be the only characteristic common to all larval habitats of this species [103, 230, 302, 303, 305–308].
Farid [302] described An. sergentii as '...an indiscriminate biter of both humans and animals, both indoors and out.', however, no study could be found that specifically tested host preference. Six studies have reported blood meal analyses of resting mosquitoes, taken from both human and animal shelters (Table 17). Of these, five described An. sergentii as principally or even highly zoophilic [103, 259, 306, 308, 309], with Kenawy et al. [308] stating the key factor that limits oasis malaria transmission in Egypt is the zoophilic feeding behaviour of An. sergentii. Faraj et al. [259], in Morocco, also described 'a marked preference for zoophily' in this species.
Kenawy et al. [309] related human biting to local animal stabling practices. They found that in villages where animals were housed in rooms within human habitations, a lower proportion of the An. sergentii females collected resting in the houses contained human blood. In an earlier study, however, the proportion of females with human blood was higher in those taken from houses containing animal rooms (86%), although the absolute numbers (calculated here from percentages reported in the paper) of 18/21 versus 26/54 (48%) from houses with no animal rooms also indicate that An. sergentii were diverted away from human hosts and towards the animals when in close proximity of one another. In this latter study, the animal rooms within the houses had the highest number of resting mosquitoes (209), of which 13% (equivalent to 27 mosquitoes) had fed on humans compared to those in isolated animal sheds (126) with only 7% (equivalent to nine mosquitoes) containing human blood.
No studies were found that specifically tested biting location, yet Abdoon & Alshahrani [310] reported high numbers biting outdoors and concluded An. sergentii was both exophagic and exophilic after finding few females resting inside houses. However, with no comparable indoor biting data and no sampling of resting mosquitoes from animal shelters, this conclusion may not indicate a true blood feeding preference. Indeed, Saliternik [285] described An. sergentii as feeding and resting both indoors and outdoors, but referred to 'exophilic habits'. Barkai & Saliternik [304] suggested that an 'exophilic strain' of An. sergentii had developed because of indoor spraying with DDT in Israel, finding fewer adults at indoor resting places where they had been common in the past, despite the local abundance of larvae.
Anopheles sergentii can overwinter as both adult females or larvae [230, 302], although no details regarding hibernation, blood feeding and oviposition could be found.
Anopheles superpictus
Preliminary data on An. superpictus populations sampled across Iran recently identified three genotypes (designated X, Y and Z) and raised the possibility of An. superpictus as a species complex [311]. These data have yet to be confirmed, but the wide distribution of this species across a number of diverse climatic regions (Mediterranean across to central and southwestern Asia) and the existence of eight junior synonyms, suggests the realistic possibility of An. superpictus being a complex of species and therefore warrants further investigation (Harbach, unpub. obs.).
In the published literature, An. superpictus larvae are continually associated with gravel or pebble river and stream beds in shallow, slow-flowing clear water in full sunlight [230, 250, 265, 282, 285, 289, 304, 312–314]. Typical, natural sites are small pools within or next to drying river beds, conditions which are closely related to seasonal fluctuations in precipitation [230, 265, 289, 314, 315]. At such sites, larval abundance increases only in late summer when pools are created as the river levels decline and, once water levels rise with the increasing rain during the onset of winter, these locations again become unsuitable as aquatic habitats [230, 289, 315].
Such natural limiting conditions could restrict both the distribution, abundance and period of adult activity of this species, however An. superpictus has easily adapted to human-influenced habitats, making use of irrigation channels and storage tanks and pools formed from their leakage, rice fields, ditches, borrow pits and hoof prints, amongst others [230, 282, 313, 315, 316]. Anopheles superpictus larvae have also been found in brackish water habitats [314] and in stagnant water [304, 313]. Jetten & Takken [275] state that it can occur in polluted sites, although here, no primary data were found to confirm this statement, which is also contradicted by other observations. For example, Berberian [314], stated that 'A. superpictus is never found in polluted or filthy water...' and the decline of An. superpictus (and An. sergentii) in Israel has been closely associated with sewage pollution of many of the natural streams it previously inhabited [285]. Anopheles superpictus survives at relatively high altitudes, up to 2800 m [264], replacing An. sacharovi [316] that may dominate at lower altitudes.
No publication could be found that reports any definitive host preference for An. superpictus, but it is generally given to be a zoophilic species that also readily feeds on humans [230]. Tshinaev [315] reported from Latyshaev [317] (reference unavailable) that, in Uzbekistan 'individuals that who sleep out during the summer on the flat roofs of houses and on towers are not attacked by this mosquito'. Conversely, Ramsdale & Haas [289] stated that An. superpictus in Turkey has a 'marked preference for animals but feeds on man in their absence...', but still described An. superpictus as an 'unusually dangerous mosquito' for those people who spend nights out in the open, away from villages and towns.
Again, no primary data were found describing biting location. However, An. superpictus appears to be opportunistic in its feeding habits and will enter houses to feed [250], but is generally regarded as exophagic [318, 319].
Discussion
The BRT model has been applied to contemporary data on the occurrence of 13 DVS in Africa, Europe and the Middle East using the most comprehensive database of DVS occurrence currently available. These maps and the underlying database will be made available in the public domain. We stress that the predictive maps produced will not be perfect representations of the true geographical distributions of these species but nevertheless, they represent a substantive step in improving our knowledge of the range of those DVS studied. One particular issue for any environmental-niche based mapping technique is predictions in areas that, although environmentally suitable, may not contain the vector for other biogeographical reasons. The model predicts, for example, the presence of An. arabiensis throughout Madagascar. Anopheles arabiensis is a species commonly associated with dry, savannah-type habitats and is considered absent, or at least, is rarely encountered in the humid climate of the eastern coast of Madagascar (Manguin, unpub. obs.). Conversely, An. nili has never been recorded in Madagascar (Manguin, unpub. obs.) but, as identified in the predicted distribution, there are areas where conditions are suitable for An. nili to become established if it were to be introduced.
Biases in collection location, variation in sampling methodologies, limited data for some species or an absence of data over large areas of suspected occurrence all contribute to uncertainty in the final predictions. Yet despite these limitations, the maps represent the first attempt to model DVS distributions across Africa, Europe and the Middle East using a combination of extensive occurrence data combined with contemporary EO distributions. All this information should be triangulated when evaluating the utility of the maps which are best considered as the beginning of an on-going process of understanding, describing and better predicting the range of these DVS. This process may be further complicated by the ever evolving revision of taxonomic status of a number of the African DVS that may lead to further stratification of the occurrence data and revisions of the predictions. This is particularly important where newly identified forms are associated with varying bionomics relevant to their control; the molecular and chromosomal forms of An. gambiae are but one example.
Bionomics
The behavioural plasticity, large geographic ranges, and changing taxonomic categorisation, in particular with the African DVS, present challenges when summarising the bionomics of individual species. Moreover, conclusions drawn about behavioural characteristics based on biased sampling may mask the true variability in a population and behavioural adaptation to human influences, such as insecticide use or environmental disturbance, can also influence local variation in species bionomics. The bionomics data are again viewed as a significant compendium but with the caveat that expert, local knowledge should always complement the information provided.
Future work
This is the second in a series of three publications describing the distribution and relevant bionomics of the global DVS of malaria. The first publication [2] detailed the DVS of the Americas and the final publication will examine the DVS of the Asian Pacific region (Sinka et al: The dominant Anopheles vectors of human malaria in the Asia Pacific region: occurrence data, distribution maps and bionomic précis, unpublished). Together, these three publications are intended to provide a baseline set of data and maps and summarise the current knowledge of the bionomics of the 41 DVS identified as the primary vectors of P. falciparum and P. vivax malaria.
Conclusions
The maps and data presented here, and those relating to the DVS of the Americas [2], and the Asian Pacific region, will be available on the MAP website [64] in accordance with the open access principles of the MAP (please contact authors for details). These data and maps are provided as a dataset to be improved and built upon. Undoubtedly, the process of species distribution mapping will improve, environmental and climatic spatial data will become available at higher resolutions, and more refined understanding of the ecology that limits a given DVS distribution attained. The single most important factor, however, will be more spatially comprehensive occurrence data and this exercise has been additionally valuable in identifying the paucity of information in large areas in Africa, Europe and the Middle East. An increasing willingness to share data between research groups and national malaria control programmes has been instrumental in this initiative and is critical to its sustained future.
Abbreviations
- AUC:
-
Area Under the operating characteristic Curve
- AVHRR:
-
Advanced Very High Resolution Radiometer
- BRT:
-
Boosted Regression Trees
- COI:
-
(mitochondrial) Cytochrome Oxidase 1
- DEM:
-
Digital Elevation Model
- DVS:
-
Dominant Vector Species
- EO:
-
Expert Opinion
- EVI:
-
Enhanced Vegetation Index
- GIS:
-
Geographic Information System
- IRS:
-
Insecticide Residual Spraying
- ITNs:
-
Insecticide Treated Bednets
- ITS2:
-
Internal Transcribed Spacer 2
- IVCC:
-
Innovative Vector Control Consortium
- LST:
-
Land Surface Temperature
- MAP:
-
Malaria Atlas Project
- MIR:
-
Middle Infrared Radiation
- MODIS:
-
MODerate Resolution Imaging Spectroradiometer
- NDVI:
-
Normalized Difference Vegetation Index
- PCR:
-
Polymerase Chain Reaction
- TAG:
-
Technical Advisory Group
- TFA:
-
Temporal Fourier Analysis
- WRBU:
-
Walter Reed Biosystematics Unit.
References
Hay SI, Sinka ME, Okara RM, Kabaria CW, Mbithi PM, Tago CT, Benz D, Gething PW, Howes RE, Patil AP, Temperley WH, Bangs MJ, Chareonviriyaphap T, Elyazar IRF, Harbach RE, Hemingway J, Manguin S, Mbogo CM, Rubio-Palis Y, Godfray HCJ: Developing global maps of the dominant Anopheles vectors of human malaria. PLoS Med. 2010, 7: e1000209-10.1371/journal.pmed.1000209.
Sinka ME, Rubio-Palis Y, Manguin S, Patil AP, Temperley WH, Gething PW, Van Boeckel T, Kabaria CW, Harbach RE, Hay SI: The dominant Anopheles vectors of human malaria in the Americas: occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010, 3: 72-10.1186/1756-3305-3-72.
Hay SI, Guerra CA, Tatem AJ, Atkinson PM, Snow RW: Urbanization, malaria transmission and disease burden in Africa. Nat Rev Microbiol. 2005, 3: 81-90. 10.1038/nrmicro1069.
Hay SI, Rogers DJ, Toomer JF, Snow RW: Annual Plasmodium falciparum entomological inoculation rates (EIR) across Africa: literature survey, internet access and review. Trans R Soc Trop Med Hyg. 2000, 94: 113-127. 10.1016/S0035-9203(00)90246-3.
Guerra CA, Gikandi PW, Tatem AJ, Noor AM, Smith DL, Hay SI, Snow RW: The limits and intensity of Plasmodium falciparum transmission: implications for malaria control and elimination worldwide. PLoS Med. 2008, 5: e38-10.1371/journal.pmed.0050038.
Fontenille D, Simard F: Unravelling complexities in human malaria transmission dynamics in Africa through a comprehensive knowledge of vector populations. Comp Immunol Microbiol Infect Dis. 2004, 27: 357-375. 10.1016/j.cimid.2004.03.005.
Hay SI, Okiro EA, Gething PW, Patil AP, Tatem AJ, Guerra CA, Snow RW: Estimating the global clinical burden of Plasmodium falciparum malaria in 2007. PLoS Med. 2010, 7: e1000290-10.1371/journal.pmed.1000290.
Hay SI, Guerra CA, Gething PW, Patil AP, Tatem AJ, Noor AM, Kabaria CW, Manh BH, Elyazar IR, Brooker S, Smith DL, Moyeed RA, Snow RW: A world malaria map: Plasmodium falciparum endemicity in 2007. PLoS Med. 2009, 6: e1000048-
Coluzzi M: The clay feet of the malaria giant and its African roots: hypotheses and inferences about origin, spread and control of Plasmodium falciparum. Parassitologia. 1999, 41: 277-283.
Gillies MT, de Meillon B: The Anophelinae of Africa South of the Sahara (Ethiopian zoogeographical region). 1968, Johannesburg: The South African Institute for Medical Research, 2
Gillies MT, Coetzee M: A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). 1987, Johannesburg: The South African Institute for Medical Research
Harbach RE: The classification of genus Anopheles (Diptera: Culicidae): a working hypothesis of phylogenetic relationships. Bull Entomol Res. 2004, 94: 537-553. 10.1079/BER2004321.
Bryan JH: Anopheles gambiae and A. melas at Brefet, The Gambia, and their role in malaria transmission. Ann Trop Med Parasitol. 1983, 77: 1-12.
Temu EA, Minjas JN, Coetzee M, Hunt RH, Shift CJ: The role of four anopheline species (Diptera: Culicidae) in malaria transmission in coastal Tanzania. Trans R Soc Trop Med Hyg. 1998, 92: 152-158. 10.1016/S0035-9203(98)90724-6.
Cuamba N, Mendis C: The role of Anopheles merus in malaria transmission in an area of southern Mozambique. J Vector Borne Dis. 2009, 46: 157-159.
White GB: Anopheles bwambae sp.n., a malaria vector in the Semliki Valley, Uganda, and its relationships with other sibling species of the An. gambiae complex (Diptera: Culicidae). Syst Entomol. 1985, 10: 501-522. 10.1111/j.1365-3113.1985.tb00155.x.
Coluzzi M: Heterogeneities of the malaria vectorial system in tropical Africa and their significance in malaria epidemiology and control. Bull World Health Organ. 1984, 62 (Suppl): 107-113.
Coetzee M, Fontenille D: Advances in the study of Anopheles funestus, a major vector of malaria in Africa. Insect Biochem Mol Biol. 2004, 34: 599-605. 10.1016/j.ibmb.2004.03.012.
Charlwood JD, Smith T, Kihonda J, Heiz B, Billingsley PF, Takken W: Density-independent feeding success of malaria vectors (Diptera: Culicidae) in Tanzania. Bull Entomol Res. 1995, 85: 29-35. 10.1017/S0007485300051981.
WHO (World Health Organization): World Malaria Report 2009. 2009
Enayati A, Hemingway J: Malaria management: past, present, and future. Annu Rev Entomol. 2010, 55: 569-591. 10.1146/annurev-ento-112408-085423.
Noor AM, Mutheu JJ, Tatem AJ, Hay SI, Snow RW: Insecticide-treated net coverage in Africa: mapping progress in 2000-07. Lancet. 2009, 373: 58-67. 10.1016/S0140-6736(08)61596-2.
Pluess B, Tanser FC, Lengeler C, Sharp BL: Indoor residual spraying for preventing malaria. Cochrane Database Syst Rev. 2010, 4: CD006657-
Coetzee M, Craig M, le Sueur D: Distribution of African malaria mosquitoes belonging to the Anopheles gambiae complex. Parasitol Today. 2000, 16: 74-77. 10.1016/S0169-4758(99)01563-X.
Utzinger J, Tanner M, Kammen DM, Killeen GF, Singer BH: Integrated programme is key to malaria control. Nature. 2002, 419: 431-10.1038/419431a.
Alphey L, Beard CB, Billingsley P, Coetzee M, Crisanti A, Curtis C, Eggleston P, Godfray C, Hemingway J, Jacobs-Lorena M, James AA, Kafatos FC, Mukwaya LG, Paton M, Powell JR, Schneider W, Scott TW, Sina B, Sinden R, Sinkins S, Spielman A, Toure Y, Collins FH: Malaria control with genetically manipulated insect vectors. Science. 2002, 298: 119-121. 10.1126/science.1078278.
Sinkins SP, Godfray HC: Use of Wolbachia to drive nuclear transgenes through insect populations. Proc Biol Sci. 2004, 271: 1421-1426. 10.1098/rspb.2004.2740.
Deredec A, Burt A, Godfray HC: The population genetics of using homing endonuclease genes in vector and pest management. Genetics. 2008, 179: 2013-2026. 10.1534/genetics.108.089037.
Awolola TS, Oduola AO, Oyewole IO, Obansa JB, Amajoh CN, Koekemoer LL, Coetzee M: Dynamics of knockdown pyrethroid insecticide resistance alleles in a field population of Anopheles gambiae s.s. in southwestern Nigeria. J Vector Borne Dis. 2007, 44: 181-188.
Elissa N, Mouchet J, Riviere F, Meunier JY, Yao K: Resistance of Anopheles gambiae s.s. to pyrethroids in Côte d'Ivoire. Ann Soc Belg Med Trop. 1993, 73: 291-294.
Baleta A: Insecticide resistance threatens malaria control in Africa. Lancet. 2009, 374: 1581-1582. 10.1016/S0140-6736(09)61933-4.
Vulule JM, Beach RF, Atieli FK, McAllister JC, Brogdon WG, Roberts JM, Mwangi RW, Hawley WA: Elevated oxidase and esterase levels associated with permethrin tolerance in Anopheles gambiae from Kenyan villages using permethrin-impregnated nets. Med Vet Entomol. 1999, 13: 239-244. 10.1046/j.1365-2915.1999.00177.x.
Akogbéto M, Yakoubou S: Résistance des vecteurs du paludisme vis-à-vis des pyréthrinoïdes utilisés pour l'imprégnation des moustiquaires au Bénin, Afrique de l'Ouest. Bull Soc Pathol Exot. 1999, 92: 123-130.
Pinto J, Lynd A, Elissa N, Donnelly MJ, Costa C, Gentile G, Caccone A, do Rosario VE: Co-occurrence of East and West African kdr mutations suggests high levels of resistance to pyrethroid insecticides in Anopheles gambiae from Libreville, Gabon. Med Vet Entomol. 2006, 20: 27-32. 10.1111/j.1365-2915.2006.00611.x.
Chandre F, Darriet F, Manga L, Akogbeto M, Faye O, Mouchet J, Guillet P: Status of pyrethroid resistance in Anopheles gambiae sensu lato. Bull World Health Organ. 1999, 77: 230-234.
Etang J, Manga L, Chandre F, Guillet P, Fondjo E, Mimpfoundi R, Toto JC, Fontenille D: Insecticide susceptibility status of Anopheles gambiae s.l. (Diptera : Culicidae) in the Republic of Cameroon. J Med Entomol. 2003, 40: 491-497. 10.1603/0022-2585-40.4.491.
Ranson H, Abdallah H, Badolo A, Guelbeogo WM, Kerah-Hinzoumbe C, Yangalbe-Kalnone E, Sagnon N, Simard F, Coetzee M: Insecticide resistance in Anopheles gambiae: data from the first year of a multi-country study highlight the extent of the problem. Malar J. 2009, 8: 299-10.1186/1475-2875-8-299.
Reiter P: Global warming and malaria: knowing the horse before hitching the cart. Malar J. 2008, 7 (Suppl 1): S3-10.1186/1475-2875-7-S1-S3.
de Zulueta J, Ramsdale CD, Coluzzi M: Receptivity to malaria in Europe. Bull World Health Organ. 1975, 52: 109-111.
Daškova NG, Rasnicyn SP: Review of data on susceptibility of mosquitoes in the USSR to imported strains of malaria parasites. Bull World Health Organ. 1982, 60: 893-897.
EDEN project (Emerging Diseases in a changing European eNvironment. [http://www.eden-fp6project.net/emerging_diseases/malaria]
Linard C, Poncon N, Fontenille D, Lambin EF: Risk of malaria reemergence in southern France: testing scenarios with a multiagent simulation model. Ecohealth. 2009, 6: 135-147. 10.1007/s10393-009-0236-y.
Sainz-Elipe S, Latorre JM, Escosa R, Masia M, Fuentes MV, Mas-Coma S, Bargues MD: Malaria resurgence risk in southern Europe: climate assessment in an historically endemic area of rice fields at the Mediterranean shore of Spain. Malar J. 2010, 9: 221-10.1186/1475-2875-9-221.
Léong Pock Tsy JM, Duchemin JB, Marrama L, Rabarison P, Le Goff G, Rajaonarivelo V, Robert V: Distribution of the species of the Anopheles gambiae complex and first evidence of Anopheles merus as a malaria vector in Madagascar. Malar J. 2003, 2: 33-10.1186/1475-2875-2-33.
Ayala D, Costantini C, Ose K, Kamdem GC, Antonio-Nkondjio C, Agbor JP, Awono-Ambene P, Fontenille D, Simard F: Habitat suitability and ecological niche profile of major malaria vectors in Cameroon. Malar J. 2009, 8: 307-10.1186/1475-2875-8-307.
Sogoba N, Vounatsou P, Bagayoko MM, Doumbia S, Dolo G, Gosoniu L, Traore SF, Toure YT, Smith T: The spatial distribution of Anopheles gambiae sensu stricto and An. arabiensis (Diptera: Culicidae) in Mali. Geospat Health. 2007, 2: 213-222.
de Souza D, Kelly-Hope L, Lawson B, Wilson M, Boakye D: Environmental factors associated with the distribution of Anopheles gambiae s.s in Ghana; an important vector of lymphatic filariasis and malaria. PLoS One. 2010, 5: e9927-10.1371/journal.pone.0009927.
Onyabe DY, Conn JE: The distribution of two major malaria vectors, Anopheles gambiae and Anopheles arabiensis, in Nigeria. Mem Inst Oswaldo Cruz. 2001, 96: 1081-1084. 10.1590/S0074-02762001000800009.
Kulkarni MA, Desrochers RE, Kerr JT: High resolution niche models of malaria vectors in northern Tanzania: a new capacity to predict malaria risk?. PLoS One. 2010, 5: e9396-10.1371/journal.pone.0009396.
Okara RM, Sinka ME, Minakawa N, Mbogo CM, Hay SI, Snow RW: Distribution of the main malaria vectors in Kenya. Malar J. 9: 69-10.1186/1475-2875-9-69.
Bayoh MN, Thomas CJ, Lindsay SW: Mapping distributions of chromosomal forms of Anopheles gambiae in West Africa using climate data. Med Vet Entomol. 2001, 15: 267-274. 10.1046/j.0269-283x.2001.00298.x.
Lindsay SW, Parson L, Thomas CJ: Mapping the ranges and relative abundance of the two principal African malaria vectors, Anopheles gambiae sensu stricto and An. arabiensis, using climate data. Proc Biol Sci. 1998, 265: 847-854. 10.1098/rspb.1998.0369.
Levine RS, Peterson AT, Benedict MQ: Geographic and ecologic distributions of the Anopheles gambiae complex predicted using a genetic algorithm. Am J Trop Med Hyg. 2004, 70: 105-109.
Rogers DJ, Randolph SE, Snow RW, Hay SI: Satellite imagery in the study and forecast of malaria. Nature. 2002, 415: 710-715. 10.1038/415710a.
Moffett A, Shackelford N, Sarkar S: Malaria in Africa: vector species' niche models and relative risk maps. PLoS One. 2007, 2: e824-10.1371/journal.pone.0000824.
Kuhn KG, Campbell-Lendrum DH, Davies CR: A continental risk map for malaria mosquito (Diptera: Culicidae) vectors in Europe. J Med Entomol. 2002, 39: 621-630. 10.1603/0022-2585-39.4.621.
Snow K: Distribution of Anopheles mosquitoes in the British Isles. Euro Mosq Bull. 1998, 1: 9-13.
Toty C, Barré H, Le Goff G, Larget-Thiéry I, Rahola N, Couret D, Fontenille D: Malaria risk in Corsica, former hot spot of malaria in France. Malar J. 2010, 9: 231-10.1186/1475-2875-9-231.
Hunt RH, Coetzee M, Fettene M: The Anopheles gambiae complex: a new species from Ethiopia. Trans R Soc Trop Med Hyg. 1998, 92: 231-235. 10.1016/S0035-9203(98)90761-1.
Costantini C, Sagnon N, Ilboudo-Sanogo E, Coluzzi M, Boccolini D: Chromosomal and bionomic heterogeneities suggest incipient speciation in Anopheles funestus from Burkina Faso. Parassitologia. 1999, 41: 595-611.
Garros C, Koekemoer LL, Kamau L, Awolola TS, Van Bortel W, Coetzee M, Coosemans M, Manguin S: Restriction fragment length polymorphism method for the identification of major African and Asian malaria vectors within the Anopheles funestus and An. minimus groups. Am J Trop Med Hyg. 2004, 70: 260-265.
Koekemoer LL, Kamau L, Garros C, Manguin S, Hunt RH, Coetzee M: Impact of the Rift Valley on restriction fragment length polymorphism typing of the major African malaria vector Anopheles funestus (Diptera: Culicidae). J Med Entomol. 2006, 43: 1178-1184. 10.1603/0022-2585(2006)43[1178:IOTRVO]2.0.CO;2.
Spillings BL, Brooke BD, Koekemoer LL, Chiphwanya J, Coetzee M, Hunt RH: A new species concealed by Anopheles funestus Giles, a major malaria vector in Africa. Am J Trop Med Hyg. 2009, 81: 510-515.
MAP. [http://www.map.ox.ac.uk/]
De'ath G: Boosted trees for ecological modeling and prediction. Ecology. 2007, 88: 243-251. 10.1890/0012-9658(2007)88[243:BTFEMA]2.0.CO;2.
Elith J, Leathwick JR, Hastie T: A working guide to boosted regression trees. J Anim Ecol. 2008, 77: 802-813. 10.1111/j.1365-2656.2008.01390.x.
Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberon J, Williams S, Wisz MS, Zimmermann NE: Novel methods improve prediction of species' distributions from occurrence data. Ecography. 2006, 29: 129-151. 10.1111/j.2006.0906-7590.04596.x.
R: A language and environment for statistical computing. R Foundation for Statistical Computing. [http://www.R-project.org]
Scharlemann JP, Benz D, Hay SI, Purse BV, Tatem AJ, Wint GR, Rogers DJ: Global data for ecology and epidemiology: a novel algorithm for temporal Fourier processing MODIS data. PLoS One. 2008, 3: e1408-10.1371/journal.pone.0001408.
MODIS (NASA). [http://modis.gsfc.nasa.gov/about]
NASA Jet Propulsion Laboratory. [http://www2.jpl.nasa.gov/srtm/]
United States Geological Survey. [http://dds.cr.usgs.gov/srtm/version2_1/SRTM30/srtm30_documentation.pdf]
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A: Very high resolution interpolated climate surfaces for global land areas. Int J Climatol. 2005, 25: 1965-1978. 10.1002/joc.1276.
Worldclim Database. [http://www.worldclim.org]
Hay SI: An overview of remote sensing and geodesy for epidemiology and public health application. Adv Parasitol. 2000, 47: 1-35. full_text.
Myneni RB, Hall FG, Sellers PJ, Marshak AL: The interpretation of spectral vegetation indexes. IEEE T Geosci Remote. 1995, 33: 481-486. 10.1109/36.377948.
Li ZL, Becker F: Feasibility of land surface temperature and emissivity determination from AVHRR data. Rem Sens Environ. 1993, 43: 67-85. 10.1016/0034-4257(93)90065-6.
Boyd DS, Petitcolin F: Remote sensing of the terrestrial environment using middle infrared radiation (3.0-5.0 μm). Int J Remote Sens. 2004, 25: 3343-3368. 10.1080/01431160310001654356.
The Globcover Project. .http://postel.mediasfrance.org, [http://www.esa.int/due/ionia/globcover]
IVCC. [http://www.ivcc.com/]
Pinto J, Lynd A, Vicente JL, Santolamazza F, Randle NP, Gentile G, Moreno M, Simard F, Charlwood JD, do Rosario VE, Caccone A, Della Torre A, Donnelly MJ: Multiple origins of knockdown resistance mutations in the Afrotropical mosquito vector Anopheles gambiae. PLoS One. 2007, 2: e1243-10.1371/journal.pone.0001243.
Brooke BD, Koekemoer LL: Major effect genes or loose confederations? The development of insecticide resistance in the malaria vector Anopheles gambiae. Parasit Vectors. 2010, 3: 74-10.1186/1756-3305-3-74.
Pombi M, Caputo B, Simard F, Di Deco MA, Coluzzi M, Della Torre A, Costantini C, Besansky NJ, Petrarca V: Chromosomal plasticity and evolutionary potential in the malaria vector Anopheles gambiae sensu stricto: insights from three decades of rare paracentric inversions. BMC Evol Biol. 2008, 8: 309-10.1186/1471-2148-8-309.
Cohuet A, Simard F, Wondji CS, Antonio-Nkondjio C, Awono-Ambene P, Fontenille D: High malaria transmission intensity due to Anopheles funestus (Diptera: Culicidae) in a village of savannah-forest transition area in Cameroon. J Med Entomol. 2004, 41: 901-905. 10.1603/0022-2585-41.5.901.
Antonio-Nkondjio C, Atangana J, Ndo C, Awono-Ambene P, Fondjo E, Fontenille D, Simard F: Malaria transmission and rice cultivation in Lagdo, northern Cameroon. Trans R Soc Trop Med Hyg. 2008
Antonio-Nkondjio C, Kerah CH, Simard F, Awono-Ambene P, Chouaibou M, Tchuinkam T, Fontenille D: Complexity of the malaria vectorial system in Cameroon: contribution of secondary vectors to malaria transmission. J Med Entomol. 2006, 43: 1215-1221. 10.1603/0022-2585(2006)43[1215:COTMVS]2.0.CO;2.
Walton C, Sharpe RG, Pritchard SJ, Thelwell NJ, Butlin RK: Molecular identification of mosquito species. Biol J Linn Soc. 1999, 68: 241-256. 10.1111/j.1095-8312.1999.tb01168.x.
Coetzee M: Distribution of the African malaria vectors of the Anopheles gambiae complex. Am J Trop Med Hyg. 2004, 70: 103-104.
Manguin S, Carnevale P, Mouchet J, Coosemans M, Julvez J, Richard-Lenoble D, Sircoulon J: Biodiversity of malaria in the world. 2008, Montrouge, France: John Libbey Eurotext
Omlin FX, Carlson JC, Ogbunugafor CB, Hassanali A: Anopheles gambiae exploits the treehole ecosystem in western Kenya: a new urban malaria risk?. Am J Trop Med Hyg. 2007, 77: 264-269.
Minakawa N, Sonye G, Mogi M, Yan G: Habitat characteristics of Anopheles gambiae s.s. larvae in a Kenyan highland. Med Vet Entomol. 2004, 18: 301-305. 10.1111/j.0269-283X.2004.00503.x.
Himeidan YE, Zhou G, Yakob L, Afrane Y, Munga S, Atieli H, El-Rayah el A, Githeko AK, Yan G: Habitat stability and occurrences of malaria vector larvae in western Kenya highlands. Malar J. 2009, 8: 234-10.1186/1475-2875-8-234.
Mouchet J, Manguin S, Sircoulon J, Laventure S, Faye O, Onapa AW, Carnevale P, Julvez J, Fontenille D: Evolution of malaria in Africa for the past 40 years: impact of climatic and human factors. J Am Mosq Control Assoc. 1998, 14: 121-130.
White GB: The Anopheles gambiae complex and malaria transmission around Kisumu, Kenya. Trans R Soc Trop Med Hyg. 1972, 66: 572-581. 10.1016/0035-9203(72)90302-1.
White GB: Anopheles gambiae complex and disease transmission in Africa. Trans R Soc Trop Med Hyg. 1974, 68: 278-301. 10.1016/0035-9203(74)90035-2.
Sharp BL, Lesueur D: Behavioral variation of Anopheles arabiensis (Diptera: Culicidae) populations in Natal, South Africa. Bull Entomol Res. 1991, 81: 107-110. 10.1017/S000748530005330X.
Coluzzi M, Sabatini A, Petrarca V, Di Deco MA: Chromosomal differentiation and adaptation to human environments in the Anopheles gambiae complex. Trans R Soc Trop Med Hyg. 1979, 73: 483-497. 10.1016/0035-9203(79)90036-1.
Ameneshewa B, Service MW: Resting habits of Anopheles arabiensis in the Awash River valley of Ethiopia. Ann Trop Med Parasitol. 1996, 90: 515-521.
Service MW: Anopheles gambiae: Africa's principal malaria vector, 1902-1984. Bull Entomol Soc Am. 1985, Autumn: 8-12.
Gimnig JE, Ombok M, Kamau L, Hawley WA: Characteristics of larval anopheline (Diptera: Culicidae) habitats in Western Kenya. J Med Entomol. 2001, 38: 282-288. 10.1603/0022-2585-38.2.282.
Edillo FE, Toure YT, Lanzaro GC, Dolo G, Taylor CE: Spatial and habitat distribution of Anopheles gambiae and Anopheles arabiensis (Diptera: Culicidae) in Banambani village, Mali. J Med Entomol. 2002, 39: 70-77. 10.1603/0022-2585-39.1.70.
Himeidan YE, Rayah Eel A: Role of some environmental factors on the breeding activity of Anopheles arabiensis in New Halfa town, eastern Sudan. East Mediterr Health J. 2008, 14: 252-259.
Abdullah MA, Merdan AI: Distribution and ecology of the mosquito fauna in the southwestern Saudi Arabia. J Egypt Soc Parasitol. 1995, 25: 815-837.
Kamau L, Munyekenye GO, Vulule JM, Lehmann T: Evaluating genetic differentiation of Anopheles arabiensis in relation to larval habitats in Kenya. Infect Genet Evol. 2007, 7: 293-297. 10.1016/j.meegid.2006.11.006.
Shililu J, Mbogo C, Ghebremeskel T, Githure J, Novak R: Mosquito larval habitats in a semi-arid ecosystem in Eritrea: impact of larval habitat management on Anopheles arabiensis populations. Am J Trop Med Hyg. 2007, 76: 103-110.
Shililu J, Ghebremeskel T, Seulu F, Mengistu S, Fekadu H, Zerom M, Ghebregziabiher A, Sintasath D, Bretas G, Mbogo C, Githure J, Brantly E, Novak R, Beier JC: Larval habitat diversity and ecology of anopheline larvae in Eritrea. J Med Entomol. 2003, 40: 921-929. 10.1603/0022-2585-40.6.921.
Ye-Ebiyo Y, Pollack RJ, Spielman A: Enhanced development in nature of larval Anopheles arabiensis mosquitoes feeding on maize pollen. Am J Trop Med Hyg. 2000, 63: 90-93.
Charlwood JD, Edoh D: Polymerase chain reaction used to describe larval habitat use by Anopheles gambiae complex (Diptera: Culicidae) in the environs of Ifakara, Tanzania. J Med Entomol. 1996, 33: 202-204.
Bøgh C, Clarke SE, Jawara M, Thomas CJ, Lindsay SW: Localized breeding of the Anopheles gambiae complex (Diptera: Culicidae) along the River Gambia, West Africa. Bull Entomol Res. 2003, 93: 279-287. 10.1079/BER2003239.
Mutero CM, Blank H, Konradsen F, van der Hoek W: Water management for controlling the breeding of Anopheles mosquitoes in rice irrigation schemes in Kenya. Acta Trop. 2000, 76: 253-263. 10.1016/S0001-706X(00)00109-1.
Mwangangi JM, Muturi EJ, Shililu J, Muriu SM, Jacob B, Kabiru EW, Mbogo CM, Githure J, Novak R: Survival of immature Anopheles arabiensis (Diptera: Culicidae) in aquatic habitats in Mwea rice irrigation scheme, central Kenya. Malar J. 2006, 5: 114-10.1186/1475-2875-5-114.
Mwangangi J, Shililu J, Muturi E, Gu WD, Mbogo C, Kabiru E, Jacob B, Githure J, Novak R: Dynamics of immature stages of Anopheles arabiensis and other mosquito species (Diptera: Culicidae) in relation to rice cropping in a rice agro-ecosystem in Kenya. J Vector Ecol. 2006, 31: 245-251. 10.3376/1081-1710(2006)31[245:DOISOA]2.0.CO;2.
Mwangangi JM, Muturi EJ, Shililu JI, Muriu S, Jacob B, Kabiru EW, Mbogo CM, Githure JI, Novak RJ: Environmental covariates of Anopheles arabiensis in a rice agroecosystem in Mwea, Central Kenya. J Am Mosq Control Assoc. 2007, 23: 371-377. 10.2987/5605.1.
Githeko AK, Service MW, Mbogo CM, Atieli FK: Resting behaviour, ecology and genetics of malaria vectors in large scale agricultural areas of Western Kenya. Parassitologia. 1996, 38: 481-489.
Chandler JA, Highton RB, Hill MN: Mosquitoes of the Kano Plain, Kenya. I. Results of indoor collections in irrigated and nonirrigated areas using human bait and light traps. J Med Entomol. 1975, 12: 504-510.
Chandler JA, Highton RB: Breeding of Anopheles gambiae Giles (Diptera: Culicidae) in rice fields in the Kisumu area of Kenya. J Med Entomol. 1976, 13: 211-215.
Mutero CM, Ng'ang'a PN, Wekoyela P, Githure J, Konradsen F: Ammonium sulphate fertiliser increases larval populations of Anopheles arabiensis and culicine mosquitoes in rice fields. Acta Trop. 2004, 89: 187-192. 10.1016/j.actatropica.2003.08.006.
Muturi EJ, Mwangangi J, Shililu J, Jacob BG, Mbogo C, Githure J, Novak RJ: Environmental factors associated with the distribution of Anopheles arabiensis and Culex quinquefasciatus in a rice agro-ecosystem in Mwea, Kenya. J Vector Ecol. 2008, 33: 56-63. 10.3376/1081-1710(2008)33[56:EFAWTD]2.0.CO;2.
Bøgh C, Clarke SE, Pinder M, Sanyang F, Lindsay SW: Effect of passive zooprophylaxis on malaria transmission in The Gambia. J Med Entomol. 2001, 38: 822-828. 10.1603/0022-2585-38.6.822.
White GB, Rosen P: Comparative studies on sibling species of Anopheles gambiae Giles complex (Dipt: Culicidae). II. Ecology of Species A and B in savanna around Kaduna, Nigeria, during transition from wet to dry season. Bull Entomol Res. 1973, 62: 613-625. 10.1017/S0007485300005502.
Tirados I, Costantini C, Gibson G, Torr SJ: Blood-feeding behaviour of the malarial mosquito Anopheles arabiensis: implications for vector control. Med Vet Entomol. 2006, 20: 425-437. 10.1111/j.1365-2915.2006.652.x.
Duchemin JB, Tsy JM, Rabarison P, Roux J, Coluzzi M, Costantini C: Zoophily of Anopheles arabiensis and An. gambiae in Madagascar demonstrated by odour-baited entry traps. Med Vet Entomol. 2001, 15: 50-57. 10.1046/j.1365-2915.2001.00276.x.
Oyewole IO, Awolola TS: Impact of urbanisation on bionomics and distribution of malaria vectors in Lagos, southwestern Nigeria. J Vector Borne Dis. 2006, 43: 173-178.
Oyewole IO, Awolola TS, Ibidapo CA, Oduola AO, Okwa OO, Obansa JA: Behaviour and population dynamics of the major anopheline vectors in a malaria endemic area in southern Nigeria. J Vector Borne Dis. 2007, 44: 56-64.
Fontenille D, Lochouarn L, Diagne N, Sokhna C, Lemasson JJ, Diatta M, Konate L, Faye F, Rogier C, Trape JF: High annual and seasonal variations in malaria transmission by anophelines and vector species composition in Dielmo, a holoendemic area in Senegal. Am J Trop Med Hyg. 1997, 56: 247-253.
Yohannes M, Haile M, Ghebreyesus TA, Witten KH, Getachew A, Byass P, Lindsay SW: Can source reduction of mosquito larval habitat reduce malaria transmission in Tigray, Ethiopia?. Trop Med Int Health. 2005, 10: 1274-1285. 10.1111/j.1365-3156.2005.01512.x.
Lemasson JJ, Fontenille D, Lochouarn L, Dia I, Simard F, Ba K, Diop A, Diatta M, Molez JF: Comparison of behavior and vector efficiency of Anopheles gambiae and An. arabiensis (Diptera: Culicidae) in Barkedji, a Sahelian area of Senegal. J Med Entomol. 1997, 34: 396-403.
Robert V, Le Goff G, Andrianaivolambo L, Randimby FM, Domarle O, Randrianarivelojosia M, Raharimanga V, Raveloson A, Ravaonjanahary C, Ariey F: Moderate transmission but high prevalence of malaria in Madagascar. Int J Parasitol. 2006, 36: 1273-1281. 10.1016/j.ijpara.2006.06.005.
Taye A, Hadis M, Adugna N, Tilahun D, Wirtz RA: Biting behavior and Plasmodium infection rates of Anopheles arabiensis from Sille, Ethiopia. Acta Trop. 2006, 97: 50-54. 10.1016/j.actatropica.2005.08.002.
Shililu J, Ghebremeskel T, Seulu F, Mengistu S, Fekadu H, Zerom M, Asmelash GE, Sintasath D, Mbogo C, Githure J, Brantly E, Beier JC, Novak RJ: Seasonal abundance, vector behavior, and malaria parasite transmission in Eritrea. J Am Mosq Control Assoc. 2004, 20: 155-164.
Githeko AK, Adungo NI, Karanja DM, Hawley WA, Vulule JM, Seroney IK, Ofulla AV, Atieli FK, Ondijo SO, Genga IO, Odada PK, Situbi PA, Oloo JA: Some observations on the biting behavior of Anopheles gambiae s.s., Anopheles arabiensis, and Anopheles funestus and their implications for malaria control. Exp Parasitol. 1996, 82: 306-315. 10.1006/expr.1996.0038.
Mnzava AE, Rwegoshora RT, Wilkes TJ, Tanner M, Curtis CF: Anopheles arabiensis and An. gambiae chromosomal inversion polymorphism, feeding and resting behaviour in relation to insecticide house-spraying in Tanzania. Med Vet Entomol. 1995, 9: 316-324. 10.1111/j.1365-2915.1995.tb00140.x.
Mosha FW, Njau RJ, Alfred J: Efficacy of Esbiothrin mosquito coils at community level in northern Tanzania. Med Vet Entomol. 1992, 6: 44-46. 10.1111/j.1365-2915.1992.tb00033.x.
Fontenille D, Lepers JP, Campbell GH, Rakotoarivony I, Coluzzi M, Coulanges P: Les vecteurs du paludisme et leur rôle dans la transmission, a Manarintsoa sur les Hauts Plateaux de Madagascar de 1988 a 1990. Arch Inst Pasteur Madagascar. 1990, 57: 335-368.
Mahande A, Mosha F, Mahande J, Kweka E: Feeding and resting behaviour of malaria vector, Anopheles arabiensis with reference to zooprophylaxis. Malar J. 2007, 6: 100-10.1186/1475-2875-6-100.
Ralisoa Randrianasolo BO, Coluzzi M: Genetical investigations on zoophilic and exophilic Anopheles arabiensis from Antananarivo area (Madagascar). Parassitologia. 1987, 29: 93-97.
Petrarca V, Nugud AD, Ahmed MA, Haridi AM, Di Deco MA, Coluzzi M: Cytogenetics of the Anopheles gambiae complex in Sudan, with special reference to An. arabiensis: relationships with East and West African populations. Med Vet Entomol. 2000, 14: 149-164. 10.1046/j.1365-2915.2000.00231.x.
Garros C, Harbach RE, Manguin S: Morphological assessment and molecular phylogenetics of the Funestus and Minimus groups of Anopheles (Cellia). J Med Entomol. 2005, 42: 522-536. 10.1603/0022-2585(2005)042[0522:MAAMPO]2.0.CO;2.
Cohuet A, Simard F, Toto JC, Kengne P, Coetzee M, Fontenille D: Species identification within the Anopheles funestus group of malaria vectors in Cameroon and evidence for a new species. Am J Trop Med Hyg. 2003, 69: 200-205.
Laventure S, Mouchet J, Blanchy S, Marrama L, Rabarison P, Andrianaivolambo L, Rajaonarivelo E, Rakotoarivony I, Roux J: Le riz source de vie et de mort sur les plateaux de Madagascar. Santé. 1996, 6: 79-86.
Marrama L, Rajaonarivelo E, Laventure S, Rabarison P: Anopheles funestus et la riziculture sur les plateaux de Madagascar. Santé. 1995, 5: 415-419.
Klinkenberg E, Takken W, Huibers F, Toure YT: The phenology of malaria mosquitoes in irrigated rice fields in Mali. Acta Trop. 2003, 85: 71-82. 10.1016/S0001-706X(02)00254-1.
Sogoba N, Doumbia S, Vounatsou P, Bagayoko MM, Dolo G, Traore SF, Maiga HM, Toure YT, Smith T: Malaria transmission dynamics in Niono, Mali: the effect of the irrigation systems. Acta Trop. 2007, 101: 232-240. 10.1016/j.actatropica.2007.02.005.
Carnevale P, Guillet P, Robert V, Fontenille D, Doannio J, Coosemans M, Mouchet J: Diversity of malaria in rice growing areas of the Afrotropical region. Parassitologia. 1999, 41: 273-276.
Antonio-Nkondjio C, Awono-Ambene P, Toto JC, Meunier JY, Zebaze-Kemleu S, Nyambam R, Wondji CS, Tchuinkam T, Fontenille D: High malaria transmission intensity in a village close to Yaounde, the capital city of Cameroon. J Med Entomol. 2002, 39: 350-355. 10.1603/0022-2585-39.2.350.
Awolola TS, Oyewole IO, Koekemoer LL, Coetzee M: Identification of three members of the Anopheles funestus (Diptera: Culicidae) group and their role in malaria transmission in two ecological zones in Nigeria. Trans R Soc Trop Med Hyg. 2005, 99: 525-531. 10.1016/j.trstmh.2004.12.003.
Dossou-Yovo J, Doannio JM, Diarrassouba S: Préférences trophiques des vecteurs du paludisme dans la ville de Bouake et dans les villages environnants de Côte d'Ivoire. Bull Soc Pathol Exot. 1998, 91: 257-258.
Costantini C, Diallo M: Preliminary lack of evidence for simian odour preferences of savanna populations of Anopheles gambiae and other malaria vectors. Parassitologia. 2001, 43: 179-182.
Dabire KR, Diabate A, Pare-Toe L, Rouamba J, Ouari A, Fontenille D, Baldet T: Year to year and seasonal variations in vector bionomics and malaria transmission in a humid savannah village in west Burkina Faso. J Vector Ecol. 2008, 33: 70-75. 10.3376/1081-1710(2008)33[70:YTYASV]2.0.CO;2.
Mwangangi JM, Mbogo CM, Nzovu JG, Githure JI, Yan G, Beier JC: Blood-meal analysis for anopheline mosquitoes sampled along the Kenyan coast. J Am Mosq Control Assoc. 2003, 19: 371-375.
Temu EA, Minjas JN, Tuno N, Kawada H, Takagi M: Identification of four members of the Anopheles funestus (Diptera: Culicidae) group and their role in Plasmodium falciparum transmission in Bagamoyo coastal Tanzania. Acta Trop. 2007, 102: 119-125. 10.1016/j.actatropica.2007.04.009.
Aniedu I: Biting activity and resting habits of malaria vectors in Baringo District, Kenya. Anz Schädlingskd Pfl Umwelt. 1993, 66: 72-76. 10.1007/BF01903074.
Appawu M, Owusu-Agyei S, Dadzie S, Asoala V, Anto F, Koram K, Rogers W, Nkrumah F, Hoffman SL, Fryauff DJ: Malaria transmission dynamics at a site in northern Ghana proposed for testing malaria vaccines. Trop Med Int Health. 2004, 9: 164-170. 10.1046/j.1365-3156.2003.01162.x.
Cano J, Berzosa PJ, Roche J, Rubio JM, Moyano E, Guerra-Neira A, Brochero H, Mico M, Edu M, Benito A: Malaria vectors in the Bioko Island (Equatorial Guinea): estimation of vector dynamics and transmission intensities. J Med Entomol. 2004, 41: 158-161. 10.1603/0022-2585-41.2.158.
Mathenge EM, Gimnig JE, Kolczak M, Ombok M, Irungu LW, Hawley WA: Effect of permethrin-impregnated nets on exiting behavior, blood feeding success, and time of feeding of malaria mosquitoes (Diptera: Culicidae) in western Kenya. J Med Entomol. 2001, 38: 531-536. 10.1603/0022-2585-38.4.531.
Rajaonarivelo V, Le Goff G, Cot M, Brutus L: Les anophèles et la transmission du paludisme à Ambohimena, village de la marge occidentale des Hautes-Terres malgaches. Parasite. 2004, 11: 75-82.
Wanji S, Tanke T, Atanga SN, Ajonina C, Nicholas T, Fontenille D: Anopheles species of the Mount Cameroon region: biting habits, feeding behaviour and entomological inoculation rates. Trop Med Int Health. 2003, 8: 643-649. 10.1046/j.1365-3156.2003.01070.x.
Bøgh C, Pedersen EM, Mukoko DA, Ouma JH: Permethrin-impregnated bednet effects on resting and feeding behaviour of lymphatic filariasis vector mosquitoes in Kenya. Med Vet Entomol. 1998, 12: 52-59. 10.1046/j.1365-2915.1998.00091.x.
Githeko AK, Service MW, Mbogo CM, Atieli FK, Juma FO: Origin of blood meals in indoor and outdoor resting malaria vectors in western Kenya. Acta Trop. 1994, 58: 307-316. 10.1016/0001-706X(94)90024-8.
Okoye PN, Brooke BD, Hunt RH, Coetzee M: Relative developmental and reproductive fitness associated with pyrethroid resistance in the major southern African malaria vector, Anopheles funestus. Bull Entomol Res. 2007, 97: 599-605. 10.1017/S0007485307005317.
Hargreaves K, Koekemoer LL, Brooke BD, Hunt RH, Mthembu J, Coetzee M: Anopheles funestus resistant to pyrethroid insecticides in South Africa. Med Vet Entomol. 2000, 14: 181-189. 10.1046/j.1365-2915.2000.00234.x.
Lochouarn L, Dia I, Boccolini D, Coluzzi M, Fontenille D: Bionomical and cytogenetic heterogeneities of Anopheles funestus in Senegal. Trans R Soc Trop Med Hyg. 1998, 92: 607-612. 10.1016/S0035-9203(98)90782-9.
Coluzzi M, Petrarca V, Dideco MA: Chromosomal inversion intergradation and incipient speciation in Anopheles gambiae. B Zool. 1985, 52: 45-63.
Fanello C, Petrarca V, della Torre A, Santolamazza F, Dolo G, Coulibaly M, Alloueche A, Curtis CF, Toure YT, Coluzzi M: The pyrethroid knock-down resistance gene in the Anopheles gambiae complex in Mali and further indication of incipient speciation within An. gambiae s.s. Insect Mol Biol. 2003, 12: 241-245. 10.1046/j.1365-2583.2003.00407.x.
Della Torre A, Fanello C, Akogbeto M, Dossou-yovo J, Favia G, Petrarca V, Coluzzi M: Molecular evidence of incipient speciation within Anopheles gambiae s.s. in West Africa. Insect Mol Biol. 2001, 10: 9-18. 10.1046/j.1365-2583.2001.00235.x.
Della Torre A, Tu ZJ, Petrarca V: On the distribution and genetic differentiation of Anopheles gambiae s.s. molecular forms. Insect Biochem Molec. 2005, 35: 755-769. 10.1016/j.ibmb.2005.02.006.
Diabate A, Dabire RK, Kim EH, Dalton R, Millogo N, Baldet T, Simard F, Gimnig JE, Hawley WA, Lehmann T: Larval development of the molecular forms of Anopheles gambiae (Diptera: Culicidae) in different habitats: a transplantation experiment. J Med Entomol. 2005, 42: 548-553. 10.1603/0022-2585(2005)042[0548:LDOTMF]2.0.CO;2.
Diabate A, Dabire RK, Heidenberger K, Crawford J, Lamp WO, Culler LE, Lehmann T: Evidence for divergent selection between the molecular forms of Anopheles gambiae: role of predation. BMC Evol Biol. 2008, 8: 5-10.1186/1471-2148-8-5.
Della Torre A, Costantini C, Besansky NJ, Caccone A, Petrarca V, Powell JR, Coluzzi M: Speciation within Anopheles gambiae-the glass is half full. Science. 2002, 298: 115-117. 10.1126/science.1078170.
Costantini C, Ayala D, Guelbeogo WM, Pombi M, Some CY, Bassole IH, Ose K, Fotsing JM, Sagnon N, Fontenille D, Besansky NJ, Simard F: Living at the edge: biogeographic patterns of habitat segregation conform to speciation by niche expansion in Anopheles gambiae. BMC Ecol. 2009, 9: 16-10.1186/1472-6785-9-16.
Caputo B, Nwakanma D, Jawara M, Adiamoh M, Dia I, Konate L, Petrarca V, Conway DJ, Della Torre A: Anopheles gambiae complex along The Gambia river, with particular reference to the molecular forms of An. gambiae s.s. Malar J. 2008, 7: 182-10.1186/1475-2875-7-182.
Olayemi IK, Ande AT: Life table analysis of Anopheles gambiae (Diptera: Culicidae) in relation to malaria transmission. J Vector Borne Dis. 2009, 46: 295-298.
Olayemi IK, Ande AT: Survivorship of Anopheles gambiae in relation to malaria transmission in Ilorin, Nigeria. Online J Health Allied Scs. 2008, 7:
Awolola TS, Okwa O, Hunt RH, Ogunrinade AF, Coetzee M: Dynamics of the malaria-vector populations in coastal Lagos, south-western Nigeria. Ann Trop Med Parasitol. 2002, 96: 75-82. 10.1179/000349802125000538.
Bockarie MJ, Service MW, Touré YT, Traoré S, Barnish G, Greenwood BM: The ecology and behaviour of the forest form of Anopheles gambiae s.s. Parassitologia. 1993, 35: 5-8.
Costantini C, Gibson G, Sagnon N, Della Torre A, Brady J, Coluzzi M: Mosquito responses to carbon dioxide in a west African Sudan savanna village. Med Vet Entomol. 1996, 10: 220-227. 10.1111/j.1365-2915.1996.tb00734.x.
Magesa SM, Wilkes TJ, Mnzava AEP, Njunwa KJ, Myamba J, Kivuyo MDP, Hill N, Lines JD, Curtis CF: Trial of pyrethroid impregnated bednets in an area of Tanzania holoendemic for malaria. Part 2. Effects on the malaria vector population. Acta Trop. 1991, 49: 97-108. 10.1016/0001-706X(91)90057-Q.
Diatta M, Spiegel A, Lochouarn L, Fontenille D: Similar feeding preferences of Anopheles gambiae and A. arabiensis in Senegal. Trans R Soc Trop Med Hyg. 1998, 92: 270-272. 10.1016/S0035-9203(98)91005-7.
Betson M, Jawara M, Awolola TS: Status of insecticide susceptibility in Anopheles gambiae s.l. from malaria surveillance sites in The Gambia. Malar J. 2009, 8: 187-10.1186/1475-2875-8-187.
Blackwell A, Johnson SN: Electrophysiological investigation of larval water and potential oviposition chemo-attractants for Anopheles gambiae s.s. Ann Trop Med Parasitol. 2000, 94: 389-398.
Koenraadt CJ, Githeko AK, Takken W: The effects of rainfall and evapotranspiration on the temporal dynamics of Anopheles gambiae s.s. and Anopheles arabiensis in a Kenyan village. Acta Trop. 2004, 90: 141-153. 10.1016/j.actatropica.2003.11.007.
Mutuku FM, Bayoh MN, Gimnig JE, Vulule JM, Kamau L, Walker ED, Kabiru E, Hawley WA: Pupal habitat productivity of Anopheles gambiae complex mosquitoes in a rural village in western Kenya. Am J Trop Med Hyg. 2006, 74: 54-61.
Mwangangi JM, Mbogo CM, Muturi EJ, Nzovu JG, Githure JI, Yan G, Minakawa N, Novak R, Beier JC: Spatial distribution and habitat characterisation of Anopheles larvae along the Kenyan coast. J Vector Borne Dis. 2007, 44: 44-51.
Awolola TS, Oduola AO, Obansa JB, Chukwurar NJ, Unyimadu JP: Anopheles gambiae s.s. breeding in polluted water bodies in urban Lagos, southwestern Nigeria. J Vector Borne Dis. 2007, 44: 241-244.
Klinkenberg E, McCall PJ, Wilson MD, Amerasinghe FP, Donnelly MJ: Impact of urban agriculture on malaria vectors in Accra, Ghana. Malar J. 2008, 7: 151-10.1186/1475-2875-7-151.
Kristan M, Fleischmann H, della Torrey A, Stich A, Curtis CF: Pyrethroid resistance/susceptibility and differential urban/rural distribution of Anopheles arabiensis and An. gambiae s.s. malaria vectors in Nigeria and Ghana. Med Vet Entomol. 2003, 17: 326-332. 10.1046/j.1365-2915.2003.00449.x.
Charlwood JD, Pinto J, Sousa CA, Ferreira C, Gil V, Do Rosario VE: Mating does not affect the biting behaviour of Anopheles gambiae from the islands of Sao Tome and Principe, West Africa. Ann Trop Med Parasitol. 2003, 97: 751-756. 10.1179/000349803225002345.
Doannio JM, Dossou-Yovo J, Diarrassouba S, Rakotondraibe ME, Chauvancy G, Chandre F, Riviere F, Carnevale P: La dynamique de la transmission du paludisme à Kafine, un village rizicole en zone de savane humide de Côte d'Ivoire. Bull Soc Pathol Exot. 2002, 95: 11-16.
Dossou-Yovo J, Diarrassouba S, Doannio J, Darriet F, Carnevale P: Le cycle d'agressivité d'Anopheles gambiae s.s. à l'intérieur des maisons et la transmission du paludisme dans la région de Bouake (Côte d'Ivoire). Interêt de l'utilisation de la moustiquaire imprégnée. Bull Soc Pathol Exot. 1999, 92: 198-200.
Girod R, Orlandi-Pradines E, Rogier C, Pages F: Malaria transmission and insecticide resistance of Anopheles gambiae (Diptera: Culicidae) in the French military camp of Port-Bouet, Abidjan (Côte d'Ivoire): implications for vector control. J Med Entomol. 2006, 43: 1082-1087. 10.1603/0022-2585(2006)43[1082:MTAIRO]2.0.CO;2.
Fontenille D, Lochouarn L, Diatta M, Sokhna C, Dia I, Diagne N, Lemasson JJ, Ba K, Tall A, Rogier C, Trape JF: Four years' entomological study of the transmission of seasonal malaria in Senegal and the bionomics of Anopheles gambiae and A. arabiensis. Trans R Soc Trop Med Hyg. 1997, 91: 647-652. 10.1016/S0035-9203(97)90506-X.
Odiere M, Bayoh MN, Gimnig J, Vulule J, Irungu L, Walker E: Sampling outdoor, resting Anopheles gambiae and other mosquitoes (Diptera: Culicidae) in western Kenya with clay pots. J Med Entomol. 2007, 44: 14-22. 10.1603/0022-2585(2007)44[14:SORAGA]2.0.CO;2.
Bryan JH, Petrarca V, Di Deco MA, Coluzzi M: Adult behaviour of members of the Anopheles gambiae complex in the Gambia with special reference to An. melas and its chromosomal variants. Parassitologia. 1987, 29: 221-249.
Gelfand HM: Anopheles gambiae Giles and Anopheles melas Theobald in a coastal area of Liberia, West Africa. Trans R Soc Trop Med Hyg. 1955, 49: 508-527. 10.1016/0035-9203(55)90023-7.
Muirhead-Thomson RC: Studies on the breeding places and control of Anopheles gambiae and A. gambiae var. melas in coastal districts of Sierra Leone. Bull Entomol Res. 1946, 36: 185-252. 10.1017/S0007485300024020.
Giglioli ME: Tides, salinity and the breeding of Anopheles melas (Theobald, 1903) during the dry season in the Gambia. Riv Malariol. 1964, 43: 245-263.
Muirhead-Thomson RC: Studies on Anopheles gambiae and A. melas in and around Lagos. Bull Entomol Res. 1948, 38: 527-558. 10.1017/S0007485300023221.
Giglioli ME: Oviposition by Anopheles melas and its effect on egg survival during the dry season in the Gambia, West Africa. Ann Entomol Soc Am. 1965, 58: 885-891.
Diop A, Molez JF, Konate L, Fontenille D, Gaye O, Diouf M, Diagne M, Faye O: Rôle d'Anopheles melas Theobald (1903) dans la transmission du paludisme dans la mangrove du Saloum (Senegal). Parasite. 2002, 9: 239-246.
Tuno N, Kjaerandsen J, Badu K, Kruppa T: Blood-feeding behavior of Anopheles gambiae and Anopheles melas in Ghana, western Africa. J Med Entomol. 2010, 47: 28-31. 10.1603/033.047.0104.
Muirhead-Thomson RC: Studies on salt-water and fresh-water Anopheles gambiae on the East African coast. Bull Entomol Res. 1951, 41: 487-502. 10.1017/S0007485300027772.
Jepson WF, Moutia A, Courtois C: The malaria problem in Mauritius; the bionomics of Mauritian anophelines. Bull Entomol Res. 1947, 38: 177-208. 10.1017/S0007485300030273.
Halcrow JG: A new sub-species of Anopheles gambiae Giles from Mauritius. East Afr Med J. 1957, 34: 133-135.
Bruce-Chwatt LJ: Correspondence of the editor: Anopheles gambiae complex in Mauritius. Trans R Soc Trop Med Hyg. 1974, 68: 497-498. 10.1016/0035-9203(74)90076-5.
Paterson HE: Status of East African salt water breeding variant of Anopheles gambiae Giles. Nature. 1962, 195: 469-470. 10.1038/195469a0.
Paterson HE: "Saltwater Anopheles gambiae" on Mauritius. Bull World Health Organ. 1964, 31: 635-644.
Mosha FW, Mutero CM: The influence of salinity on larval development and population dynamics of Anopheles merus Donitz (Diptera: Culicidae). Bull Entomol Res. 1982, 72: 119-128. 10.1017/S0007485300050331.
Govere J, Durrheim DN, Coetzee M, Hunt RH, la Grange JJ: Captures of mosquitoes of the Anopheles gambiae complex (Diptera : Culicidae) in the Lowveld Region of Mpumalanga Province, South Africa. Afr Entomol. 2000, 8: 91-99.
Masendu HT, Hunt RH, Koekemoer LL, Brooke BD, Govere J, Coetzee M: Spatial and temporal distributions and insecticide susceptibility of malaria vectors in Zimbabwe. Afr Entomol. 2005, 13: 25-34.
Kloke RG: New distribution record of Anopheles merus Donitz (Diptera: Culicidae) in Zambia. Afr Entomol. 1997, 5: 361-362.
Paterson HE, Paterson JS: Records of the breeding of 'Saltwater Anopheles gambiae' at inland localities in Southern Africa. Nature. 1964, 201: 524-525. 10.1038/201524a0.
Coetzee M, Cross H: Mating experiments with two populations of Anopheles merus Donitz (Diptera: Culicidae). J Entomol Soc S Afr. 1983, 46: 257-259.
Iyengar R: The bionomics of salt-water Anopheles gambiae in East Africa. Bull World Health Organ. 1962, 27: 223-229.
Mutero CM, Mosha FW, Subra R: Biting activity and resting behaviour of Anopheles merus Donitz (Diptera: Culicidae) on the Kenya Coast. Ann Trop Med Parasitol. 1984, 78: 43-47.
Antonio-Nkondjio C, Ndo C, Kengne P, Mukwaya L, Awono-Ambene P, Fontenille D, Simard F: Population structure of the malaria vector Anopheles moucheti in the equatorial forest region of Africa. Malar J. 2008, 7: 120-10.1186/1475-2875-7-120.
Antonio-Nkondjio C, Ndo C, Awono-Ambene P, Ngassam P, Fontenille D, Simard F: Population genetic structure of the malaria vector Anopheles moucheti in south Cameroon forest region. Acta Trop. 2007, 101: 61-68. 10.1016/j.actatropica.2006.12.004.
Antonio-Nkondjio C, Ndo C, Costantini C, Awono-Ambene P, Fontenille D, Simard F: Distribution and larval habitat characterization of Anopheles moucheti, Anopheles nili, and other malaria vectors in river networks of southern Cameroon. Acta Trop. 2009, 112: 270-276. 10.1016/j.actatropica.2009.08.009.
Manga L, Toto JC, Carnevale P: Malaria vectors and transmission in an area deforested for a new international airport in southern Cameroon. Ann Soc Belg Med Trop. 1995, 75: 43-49.
Mouchet J, Gariou J: Anopheles moucheti au Cameroun. Cahiers ORSTOM, Entomol Med Parasitol. 1966, 4: 71-81.
Antonio-Nkondjio C, Simard F, Awono-Ambene P, Ngassam P, Toto JC, Tchuinkam T, Fontenille D: Malaria vectors and urbanization in the equatorial forest region of south Cameroon. Trans R Soc Trop Med Hyg. 2005, 99: 347-354. 10.1016/j.trstmh.2004.07.003.
Mattingly PF: Studies on West African forest mosquitoes. Part II. The less commonly occurring species. Bull Entomol Res. 1949, 40: 387-402. 10.1017/S0007485300022860.
Awono-Ambene HP, Simard F, Antonio-Nkondjio C, Cohuet A, Kengne P, Fontenille D: Multilocus enzyme electrophoresis supports speciation within the Anopheles nili group of malaria vectors in Cameroon. Am J Trop Med Hyg. 2006, 75: 656-658.
Mouchet J, Carnevale P, Coosemans M, Julvez J, Manguin S, Richard-Lenoble D, Sircoulon J: Biodiversité du paludisme dans le monde. 2004, Montrouge, France: John Libbey Eurotext
Awono-Ambene HP, Kengne P, Simard F, Antonio-Nkondjio C, Fontenille D: Description and bionomics of Anopheles (Cellia) ovengensis (Diptera: Culicidae), a new malaria vector species of the Anopheles nili group from south Cameroon. J Med Entomol. 2004, 41: 561-568. 10.1603/0022-2585-41.4.561.
Dia I, Diop T, Rakotoarivony I, Kengne P, Fontenille D: Bionomics of Anopheles gambiae Giles, An. arabiensis Patton, An. funestus Giles and An. nili (Theobald) (Diptera: Culicidae) and transmission of Plasmodium falciparum in a Sudano-Guinean zone (Ngari, Senegal). J Med Entomol. 2003, 40: 279-283. 10.1603/0022-2585-40.3.279.
Carnevale P, Zoulani A: Agressivité d'Anopheles nili (Theobald), 1904 à l'intérieur et à l'extérieur des maisons. Cahiers ORSTOM, Entomol Med Parasitol. 1975, 13: 69-73.
Krafsur ES: Anopheles nili as a vector of malaria in a lowland region of Ethiopia. Bull World Health Organ. 1970, 42: 1-8.
Coene J: Malaria in urban and rural Kinshasa: the entomological input. Med Vet Entomol. 1993, 7: 127-137. 10.1111/j.1365-2915.1993.tb00665.x.
Pires CA, Ribeiro H, Capela RA, da Cunha Ramos H: Research on the mosquitoes of Portugal (Diptera: Culicidae) VI. The mosquitoes of Alentejo. An do Inst de Hig e Med Trop. 1982, 8: 79-102.
Becker N, Petric D, Boase C, Lane J, Zgomba M, Dahl C, Kaiser A: Mosquitoes and their control. 2003, New York: Kluwer Academic/Plenum Publishers
Hackett LW, Missiroli A: The varieties of Anopheles maculipennis and their relation to the distribution of malaria in Europe. Riv Malariol. 1935, 14: 1-67.
Hackett LW: The present status of our knowledge of the sub-species of Anopheles maculipennis. Trans R Soc Trop Med Hyg. 1934, 28: 109-140. 10.1016/S0035-9203(34)90135-8.
Cambournac FJ: Contribution to the history of malaria epidemiology and control in Portugal and some other places. Parassitologia. 1994, 36: 215-222.
Hackett LW: Recent additions to our knowledge of 'Anopheles maculipennis' races. Bul Hlth Org, League of Nations. 1937, 6: 1-16.
Romi R, Pontuale G, MG CI, Fiorentini G, Marchi A, Nicoletti L, Cocchi M, Tamburro A: Potential vectors of West Nile virus following an equine disease outbreak in Italy. Med Vet Entomol. 2004, 18: 14-19. 10.1111/j.1365-2915.2004.0478.x.
Roiz D, Eritja R, Escosa R, Lucientes J, Marques E, Melero-Alcibar R, Ruiz S, Molina R: A survey of mosquitoes breeding in used tires in Spain for the detection of imported potential vector species. J Vector Ecol. 2007, 32: 10-15. 10.3376/1081-1710(2007)32[10:ASOMBI]2.0.CO;2.
Poncon N, Toty C, L'Ambert G, Le Goff G, Brengues C, Schaffner F, Fontenille D: Biology and dynamics of potential malaria vectors in Southern France. Malar J. 2007, 6: 18-10.1186/1475-2875-6-18.
Muirhead-Thomson RC: Field studies of the role of Anopheles atroparvus in the transmission of myxomatosis in England. J Hyg (Lond). 1956, 54: 472-477. 10.1017/S0022172400044752.
Andrewes CH, Muirhead-Thomson RC, Stevenson JP: Laboratory studies of Anopheles atroparvus in relation to myxomatosis. J Hyg (Lond). 1956, 54: 478-486. 10.1017/S0022172400044764.
Zamburlini R, Cargnus E: Anofelismo residuo nel litorale altoadriatico a 50 anni dalla scomparsa della malaria. Parassitologia. 1998, 40: 431-437.
Ribeiro H, Ramos HC, Capela RA, Pires CA: Research on the mosquitoes of Portugal (Diptera, Culicidae). XI. The mosquitoes of Beiras. Garcia De Orta Ser Zool. 1989, 16: 137-161.
Shute PG: Failure to infect English specimens of Anopheles maculipennis var. atroparvus with certain strains of Plasmodium falciparum of tropical origin. J Trop Med Hyg. 1940, 43: 175-178.
Curtis CF, White GB: Plasmodium falciparum transmission in England: entomological and epidemiological data relative to cases in 1983. J Trop Med Hyg. 1984, 87: 101-114.
Capinha C, Gomes E, Reis E, Rocha J, Sousa CA, do Rosario VE, Almeida AP: Present habitat suitability for Anopheles atroparvus (Diptera: Culicidae) and its coincidence with former malaria areas in mainland Portugal. Geospat Health. 2009, 3: 177-187.
Sousa CA: Malaria vectorial capacity and competence of Anopheles atroparvus Van Thiel, 1927 (Diptera: Culicidae): Implications for the potential re-emergence of malaria in Portugal. PhD. 2008, Universidade Nova de Lisboa, Instituto de Hygiene e Medicina Tropical
de Zulueta J: Forty years of malaria eradication in Sardinia. A new appraisal of a great enterprise. Parassitologia. 1990, 32: 231-236.
Marchi A, Munstermann LE: The mosquitoes of Sardinia: species records 35 years after the malaria eradication campaign. Med Vet Entomol. 1987, 1: 89-96. 10.1111/j.1365-2915.1987.tb00327.x.
Trapido H, Aitken TH: Study of a residual population of Anopheles L. labranchiae Falleroni in the Geremeas Valley, Sardinia. Am J Trop Med Hyg. 1953, 2: 658-676.
D'Alessandro G, Bruno Smiraglia C, Lavagnino A: Further studies on the biology of Anopheles labranchiae labranchiae Falleroni in Sicily. 1971, WHO, 1-11.
Macdonald WW: Anophelines of Libya and their control. Gariounis Med J. 1982, 5: 72-74.
Aitken THG: The Culicidae of Sardinia and Corsica (Diptera). Bull Entomol Res. 1954, 45: 437-494. 10.1017/S0007485300029564.
Coluzzi M, Coluzzi A: Rischio di una ripresa della trasmissione della malaria in Calabria e problemi connessi al controllo dei vettori. Riv Parassitologia. 1980, 41: 117-125.
Service MW: Appendix II. Characteristics of some major Anopheles vectors of human malaria. Bruce-Chwatt's Essential Malariology. Edited by: Gilles HM, Warrell DA. 1993, London: Edward Arnold, 305-310. Third
Bettini S, Gradoni L, Cocchi M, Tamburro A: Rice culture and Anopheles labranchiae in central Italy. 1978, WHO, 1-6.
Di Luca M, Boccolini D, Severini F, Toma L, Barbieri FM, Massa A, Romi R: A 2-year entomological study of potential malaria vectors in central Italy. Vector Borne Zoonotic Dis. 2009, 9: 703-711. 10.1089/vbz.2008.0129.
Romi R, Pierdominici G, Severini C, Tamburro A, Cocchi M, Menichetti D, Pili E, Marchi A: Status of malaria vectors in Italy. J Med Entomol. 1997, 34: 263-271.
Lavagnino A: Anopheles labranchiae in Sicily: decrease of susceptibility to insecticides. Trans R Soc Trop Med Hyg. 1983, 77: 883-10.1016/0035-9203(83)90326-7.
Cefalù M, Oddo F, Saccà G: Vita extra-domestica di Anopheles labranchiae in Sicilia. Osservazioni in un'area di sospensione dei trattamenti con DDT. Parassitologia. 1961, 3: 23-50.
Faraj C, Adlaoui E, Ouahabi S, Rhajaoui M, Fontenille D, Lyagoubi M: Entomological investigations in the region of the last malaria focus in Morocco. Acta Trop. 2009, 109: 70-73. 10.1016/j.actatropica.2008.09.021.
White GB: Malaria. Geographical distribution of arthropod-borne diseases and their principal vectors. 1989, Geneva: World Health Organization, Division of Vector Biology and Control, 7-22.
Di Luca M, Boccolini D, Marinuccil M, Romi R: Intrapopulation polymorphism in Anopheles messeae (An. maculipennis complex) inferred by molecular analysis. J Med Entomol. 2004, 41: 582-586. 10.1603/0022-2585-41.4.582.
Zahar AR: Vector bionomics in the epidemiology and control of malaria. Part II. The WHO European region and the WHO Eastern Mediterranean region. Volume II: Applied field studies. Section III: Vector bionomics, malaria epidemiology and control by geographical areas. (A): The Mediterranean Basin. 1990, WHO, 226p-
Takken W, Geene R, Adam W, Jetten TH, van der Velden JA: Distribution and dynamics of larval populations of Anopheles messeae and A. atroparvus in the delta of the rivers Rhine and Meuse, The Netherlands. Ambio. 2002, 31: 212-218.
Bublikova LI: Anophelines (Diptera, Nematocera) in the Chu valley (Kirgiz Republic). Entomol Rev. 1998, 78: 398-402.
Bates M: Field studies of the anopheline mosquitoes of Albania. Proc Ent Soc Wash. 1941, 43: 37-58.
Adamovic Z: Anopheles messeae Falleroni in Serbia and Macedonia (Diptera: Culicidae). Acta Vet (Beograd). 1980, 30: 217-223.
Adamovic Z: Vertical distribution of the anopheline mosquitoes in Montenegro, Yugoslavia. Acta Vet (Beograd). 1984, 34: 287-293.
Adamovic ZR: Distribution and relative abundance of the anophelines (Diptera: Culicidae) in Kacer and Takovo, Serbia. Bull Serbian Acad Sci Arts Sec Sci Math Nat Sci. 1982, 82: 97-104.
Adamovic Z, Paulus R: Distribution and abundance of anophelines (Diptera: Culicidae) in the Sava Valley from Ljubljana to Zagreb, Yugoslavia. Acta Vet (Beograd). 1988, 38: 31-36.
Fyodorova MV, Savage HM, Lopatina JV, Bulgakova TA, Ivanitsky AV, Platonova OV, Platonov AE: Evaluation of potential West Nile virus vectors in Volgograd region, Russia, 2003 (Diptera: Culicidae): species composition, bloodmeal host utilization, and virus infection rates of mosquitoes. J Med Entomol. 2006, 43: 552-563. 10.1603/0022-2585(2006)43[552:EOPWNV]2.0.CO;2.
Jaenson TGT, Ameneshewa B: Prehibernation diet and reproductive condition of female Anopheles messeae in Sweden. Med Vet Entomol. 1991, 5: 243-252. 10.1111/j.1365-2915.1991.tb00547.x.
Linton YM, Samanidou-Voyadjoglou A, Harbach RE: Ribosomal ITS2 sequence data for Anopheles maculipennis and An. messeae in northern Greece, with a critical assessment of previously published sequences. Insect Mol Biol. 2002, 11: 379-383. 10.1046/j.1365-2583.2002.00338.x.
Nicolescu G, Linton YM, Vladimirescu A, Howard TM, Harbach RE: Mosquitoes of the Anopheles maculipennis group (Diptera: Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence. Bull Entomol Res. 2004, 94: 525-535. 10.1079/BER2004330.
Linton YM, Lee AS, Curtis C: Discovery of a third member of the Maculipennis Group in SW England. Euro Mosq Bull. 2005, 19: 5-9.
Jetten TH, Takken W: Anophelism without malaria in Europe. A review of the ecology and distribution of the genus Anopheles in Europe. 1994, Wageningen: Wageningen Agricultural University Papers, 69-
Hadjinicolaou J, Betzios B: Resurgence of Anopheles sacharovi following malaria eradication. Bull World Health Organ. 1973, 48: 699-703.
Hadjinicolaou J, Betzios B: Gambusia fish as a means of biological control of Anopheles sacharovi in Greece. 1973, WHO, 1-7.
Kitron U, Spielman A: Suppression of transmission of malaria through source reduction: anti-anopheline measures applied in Israel, the United States, and Italy. Rev Infect Dis. 1989, 11: 391-406.
Gratz NG: The effect of water development programmes on malaria vectors in Turkey. 1987, City: FAO
Sedaghat MM, Linton YM, Nicolescu G, Smith L, Koliopoulos G, Zounos AK, Oshaghi MA, Vatandoost H, Harbach RE: Morphological and molecular characterization of Anopheles (Anopheles) sacharovi Favre, a primary vector of malaria in the Middle East. Syst Entomol. 2003, 28: 241-256. 10.1046/j.1365-3113.2003.00211.x.
Pener H, Kitron U: Spatial and temporal changes in the distribution of Anopheles sacharovi in Israel. Isr J Med Sci. 1985, 21: 850-852.
Abdel-Malek AA: The anopheline mosquitoes of northern Syria. Bull Soc Entomol Egypte. 1958, 42: 519-535.
Gökberk C: Anopheles sacharovi (Favre 1903) in Turkey. Mosq News. 1961, 21: 101-102.
Etherington D, Sellick G: Notes on the bionomics of Anopheles sacharovi in Persia and Iraq. Bull Entomol Res. 1946, 37: 191-195. 10.1017/S0007485300022148.
Saliternik Z: Historical notes on the bionomics and distribution of malaria vectors in Israel. 1974, WHO, 1-11.
Pener H, Orshan L, Kitron U, Shalom U: The unexpected presence of four malaria vectors in southern Israel. Isr J Med Sci. 1994, 30: 287-288.
Kampen H, Proft J, Etti S, Maltezos E, Pagonaki M, Maier WA, Seitz HM: Individual cases of autochthonous malaria in Evros Province, northern Greece: entomological aspects. Parasitol Res. 2003, 89: 252-258.
Kasap H, Akbaba M, Kasap M, Lüleyap Ü, Alpaslan N, Alptekin D: A study of malaria and Anopheles sacharovi in Tuzla, Adana, Turkey. Acta Parasit Turcica. 1995, 19: 535-540.
Ramsdale CD, Haas E: Some aspects of epidemiology of resurgent malaria in Turkey. Trans R Soc Trop Med Hyg. 1978, 72: 570-580. 10.1016/0035-9203(78)90005-6.
Demirhan O, Kasap M: Bloodfeeding behavior of Anopheles sacharovi in Turkey. J Am Mosq Control Assoc. 1995, 11: 11-14.
Yaghoobi-Ershadi MR, Namazi J, Piazak N: Bionomics of Anopheles sacharovi in Ardebil province, northwestern Iran during a larval control program. Acta Trop. 2001, 78: 207-215. 10.1016/S0001-706X(01)00080-8.
Boreham PF, Garrett-Jones C: Prevalence of mixed blood meals and double feeding in a malaria vector (Anopheles sacharovi Favre). Bull World Health Organ. 1973, 48: 605-614.
Djadid ND, Gholizadeh S, Tafsiri E, Romi R, Gordeev M, Zakeri S: Molecular identification of Palearctic members of Anopheles maculipennis in northern Iran. Malar J. 2007, 6: 6-10.1186/1475-2875-6-6.
Alten B, Caglar SS, Simsek FM, Kaynas S: Effect of insecticide-treated bednets for malaria control in Southeast Anatolia-Turkey. J Vector Ecol. 2003, 28: 97-107.
Kasap H, Kasap M, Akbaba M, Alptekin D, Demirhan O, Luleyap U, Pazarbasi A, Akdur R, Wade J: Residual efficacy of pirimiphos methyl (Actellic) on Anopheles sacharovi in Cukurova, Turkey. J Am Mosq Control Assoc. 1992, 8: 47-51.
Abul-hab J, al-Kassal SM: Impact of anti-malaria spraying on the occurrence of Anopheles (Diptera: Culicidae) in Iraq. Bull Endem Dis. 1986, 27: 37-51.
Soliman AA: A study of the dispersal of Anopheles sacharovi Favr. from a natural breeding source in the Rouge-Valley (Aleppo-Syria U. A. R.). The Fourth Arab Science Congress; February; Cairo. 1961, 526-530.
Walter Reed Biosystematics Unit (WRBU), Mosquito Catalog. [http://www.mosquitocatalog.org/default.aspx?pgID=8]
Mosquito Taxonomic Inventory. [http://mosquito-taxonomic-inventory.info/]
Ramsdale CD, de Zulueta J: Anophelism in the Algerian Sahara and some implications of the construction of a trans-Saharan highway. J Trop Med Hyg. 1983, 86: 51-58.
Beier JC, Beier MS, el Kordy EA, el Said S, Kenawy MA: Colonization of the oasis malaria vector, Anopheles sergentii, in Egypt. J Am Mosq Control Assoc. 1986, 2: 104-105.
Farid MA: The implications of Anopheles sergentii for malaria eradication programmes east of the Mediterranean. Bull World Health Organ. 1958, 15: 821-828.
Gad A, Kenawy MA, El-Said S, Merdan AI: Field studies on anopheline mosquito larvae in Egypt (Diptera: Culicidae). I. Different types and characteristics of the breeding places in relation to the abundance of anopheline species in Egypt. J Egypt Public Health Assoc. 1982, 57: 541-562.
Barkai A, Saliternik Z: Anopheline mosquitos found breeding in Israel in 1963-1965 during last stage of the malaria eradication project. Bull Entomol Res. 1968, 58: 353-366. 10.1017/S0007485300056881.
Harbach RE, Harrison BA, Gad AM, Kenawy MA, El-Said S: Records and notes on mosquitoes (Diptera: Culicidae) collected in Egypt. Mosq Syst. 1988, 20: 317-342.
Barber MA, Rice JB: A survey of malaria in Egypt. Am J Trop Med. 1937, s1-17: 413-436.
Gad AM, el Said S, Hassan AN: Ecology of Anopheles (Cellia) sergentii Theobald in the eastern desert, Red Sea Governorate, Egypt. J Egypt Soc Parasitol. 1984, 14: 1-6.
Kenawy MA, Beier JC, el Said S: First record of malaria and associated Anopheles in El Gara Oasis, Egypt. J Am Mosq Control Assoc. 1986, 2: 101-103.
Kenawy MA, Beier JC, Asiago CM, el Said S: Factors affecting the human-feeding behavior of anopheline mosquitoes in Egyptian oases. J Am Mosq Control Assoc. 1990, 6: 446-451.
Abdoon AM, Alshahrani AM: Prevalence and distribution of anopheline mosquitoes in malaria endemic areas of Asir region, Saudi Arabia. East Mediterr Health J. 2003, 9: 240-247.
Oshaghi MA, Yaghobi-Ershadi MR, Shemshad K, Pedram M, Amani H: The Anopheles superpictus complex: introduction of a new malaria vector complex in Iran. Bull Soc Pathol Exot. 2008, 101: 429-434.
Sabatini A, Coluzzi M, Boccolini D: Field studies on inversion polymorphism in Anopheles superpictus from southern Italy. Parassitologia. 1989, 31: 69-87.
Mulligan HW: Malaria in Iraq and Persia. Protozoology. 1967, 2: 113-146.
Berberian DA: The species of anopheline mosquitoes found in Syria and Lebanon. Their habits, distribution and eradication. J of Palestine Arab Med Assoc. 1946, 1: 120-145.
Tshinaev PP: On the ecology and biology of the malarial mosquito Anopheles superpictus Grassi (Diptera: Culicidae) in Uzbekistan. Entomol Rev. 1963, 42: 169-175.
Chinayev PP: Range and epidemiological importance of the malarial mosquitoes Anopheles maculipennis sacharovi Fv. superpictus Gr. and A. pulcherrimus Theob. (Diptera: Culicidae) in Soviet central. Entomol Rev. 1965, 4: 164-169.
Latyshev NL: The biology of Anopheles superpictus Grassi in Tadzhikistan. Animal Parasites and parasitic diseases of Tadzhikistan. 1929, 41-59.
Severini C, Menegon M, Di Luca M, Abdullaev I, Majori G, Razakov SA, Gradoni L: Risk of Plasmodium vivax malaria reintroduction in Uzbekistan: genetic characterization of parasites and status of potential malaria vectors in the Surkhandarya region. Trans R Soc Trop Med Hyg. 2004, 98: 585-592. 10.1016/j.trstmh.2004.01.003.
Oshaghi MA, Shemshad K, Yaghobi-Ershadi MR, Pedram M, Vatandoost H, Abaie MR, Akbarzadeh K, Mohtarami F: Genetic structure of the malaria vector Anopheles superpictus in Iran using mitochondrial cytochrome oxidase (COI and COII) and morphologic markers: a new species complex?. Acta Trop. 2007, 101: 241-248. 10.1016/j.actatropica.2007.02.006.
Macdonald G: Local features of malaria. The epidemiology and control of malaria. 1957, London: Oxford University Press, 63-99.
Service MW: The Anopheles vector. Bruce-Chwatt's Essential Malariology. Edited by: Gilles HM, Warrell DA. 1993, London: Edward Arnold, 96-123. Third
Kiszewski A, Mellinger A, Spielman A, Malaney P, Sachs SE, Sachs J: A global index representing the stability of malaria transmission. Am J Trop Med Hyg. 2004, 70: 486-498.
Waka M, Hopkins RJ, Akinpelu O, Curtis C: Transmission of malaria in the Tesseney area of Eritrea: parasite prevalence in children, and vector density, host preferences, and sporozoite rate. J Vector Ecol. 2005, 30: 27-32.
Ameneshewa B: The behavior and biology of Anopheles arabiensis in relation to epidemiology and control of malaria in Ethiopia. PhD. 1995, University of Liverpool
Beier JC, Copeland R, Oyaro C, Masinya A, Odago WO, Oduor S, Koech DK, Roberts CR: Anopheles gambiae complex egg-stage survival in dry soil from larval development sites in western Kenya. J Am Mosq Control Assoc. 1990, 6: 105-109.
Al-Maktari MT, Bassiouny HK: Bionomics of anopheline vectors in Zabid District, Al-Hodeidah Governorate, Republic of Yemen. East Mediterr Health J. 1999, 5: 698-705.
Braack LE, Coetzee M, Hunt RH, Biggs H, Cornel A, Gericke A: Biting pattern and host-seeking behavior of Anopheles arabiensis (Diptera: Culicidae) in northeastern South Africa. J Med Entomol. 1994, 31: 333-339.
Adugna N, Petros B: Determination of the human blood index of some anopheline mosquitos by using ELISA. Ethiop Med J. 1996, 34: 1-10.
Robert V, Awono-Ambene HP, Thioulouse J: Ecology of larval mosquitoes, with special reference to Anopheles arabiensis (Diptera: Culcidae) in market-garden wells in urban Dakar, Senegal. J Med Entomol. 1998, 35: 948-955.
Romi R, Ravoniharimelina B, Ramiakajato M, Majori G: Field trials of Bacillus thuringiensis H-14 and Bacillus sphaericus (strain 2362) formulations against Anopheles arabiensis in the central highlands of Madagascar. J Am Mosq Control Assoc. 1993, 9: 325-329.
Minakawa N, Seda P, Yan G: Influence of host and larval habitat distribution on the abundance of African malaria vectors in western Kenya. Am J Trop Med Hyg. 2002, 67: 32-38.
Minakawa N, Mutero CM, Githure JI, Beier JC, Yan G: Spatial distribution and habitat characterization of anopheline mosquito larvae in Western Kenya. Am J Trop Med Hyg. 1999, 61: 1010-1016.
Petrarca V, Beier JC, Onyango F, Koros J, Asiago C, Koech DK, Roberts CR: Species composition of the Anopheles gambiae complex (Diptera: Culicidae) at two sites in western Kenya. J Med Entomol. 1991, 28: 307-313.
Coosemans M, Petrarca V, Barutwanayo M, Coluzzi M: Species of the Anopheles gambiae complex and chromosomal polymorphism in a rice-growing area of the Rusizi Valley (Republic of Burundi). Parassitologia. 1989, 31: 113-122.
Awono-Ambene HP, Robert V: Survival and emergence of immature Anopheles arabiensis mosquitoes in market-gardener wells in Dakar, Senegal. Parasite. 1999, 6: 179-184.
Edillo FE, Tripet F, Toure YT, Lanzaro GC, Dolo G, Taylor CE: Water quality and immatures of the M and S forms of Anopheles gambiae s.s. and An. arabiensis in a Malian village. Malar J. 2006, 5: 35-10.1186/1475-2875-5-35.
Jacob BG, Muturi EJ, Funes JE, Shililu JI, Githure JI, Kakoma II, Novak RJ: A grid-based infrastructure for ecological forecasting of rice land Anopheles arabiensis aquatic larval habitats. Malar J. 2006, 5: 91-10.1186/1475-2875-5-91.
Mwangangi JM, Mbogo CM, Muturi EJ, Nzovua JG, Kabiru EW, Githure JI, Novak RJ, Beier JC: Influence of biological and physicochemical characteristics of larval habitats on the body size of Anopheles gambiae mosquitoes (Diptera: Culicidae) along the Kenyan coast. J Vector Borne Dis. 2007, 44: 122-127.
Muturi EJ, Mwangangi J, Shililu J, Muriu S, Jacob B, Kabiru E, Gu W, Mbogo C, Githure J, Novak R: Mosquito species succession and physicochemical factors affecting their abundance in rice fields in Mwea, Kenya. J Med Entomol. 2007, 44: 336-344. 10.1603/0022-2585(2007)44[336:MSSAPF]2.0.CO;2.
Jacob BG, Muturi E, Halbig P, Mwangangi J, Wanjogu RK, Mpanga E, Funes J, Shililu J, Githure J, Regens JL, Novak RJ: Environmental abundance of Anopheles (Diptera: Culicidae) larval habitats on land cover change sites in Karima Village, Mwea Rice Scheme, Kenya. Am J Trop Med Hyg. 2007, 76: 73-80.
Fettene M, Hunt RH, Coetzee M, Tessema F: Behaviour of Anopheles arabiensis and An. quadriannulatus sp. B mosquitoes and malaria transmission in southwestern Ethiopia. Afr Entomol. 2004, 12: 83-87.
Karanja DMS, Githeko AK, Vulule JM: Small-scale field-evaluation of the monomolecular surface-film 'Arosurf Msf' against Anopheles arabiensis Patton. Acta Trop. 1994, 56: 365-369. 10.1016/0001-706X(94)90107-4.
Shililu JI, Tewolde GM, Brantly E, Githure JI, Mbogo CM, Beier JC, Fusco R, Novak RJ: Efficacy of Bacillus thuringiensis israelensis, Bacillus sphaericus and temephos for managing Anopheles larvae in Eritrea. J Am Mosq Control Assoc. 2003, 19: 251-258.
Chouaibou M, Etang J, Brevault T, Nwane P, Hinzoumbe CK, Mimpfoundi R, Simard F: Dynamics of insecticide resistance in the malaria vector Anopheles gambiae s.l. from an area of extensive cotton cultivation in Northern Cameroon. Trop Med Int Health. 2008, 13: 476-86. 10.1111/j.1365-3156.2008.02025.x.
Abdalla H, Matambo TS, Koekemoer LL, Mnzava AP, Hunt RH, Coetzee M: Insecticide susceptibility and vector status of natural populations of Anopheles arabiensis from Sudan. Trans R Soc Trop Med Hyg. 2008, 102: 263-271. 10.1016/j.trstmh.2007.10.008.
Tuno N, Githeko AK, Nakayama T, Minakawa N, Takagi M, Yan GY: The association between the phytoplankton, Rhopalosolen species (Chlorophyta: Chlorophyceae), and Anopheles gambiae sensu lato (Diptera: Culicidae) larval abundance in western Kenya. Ecol Res. 2006, 21: 476-482. 10.1007/s11284-005-0131-0.
Awono-Ambene HP, Diawara L, Robert V: Comparison of direct and membrane feeding methods to infect Anopheles arabiensis with Plasmodium falciparum. Am J Trop Med Hyg. 2001, 64: 32-34.
Jawara M, Pinder M, Drakeley CJ, Nwakanma DC, Jallow E, Bogh C, Lindsay SW, Conway DJ: Dry season ecology of Anopheles gambiae complex mosquitoes in The Gambia. Malar J. 2008, 7: 156-10.1186/1475-2875-7-156.
Molina R, Benito A, Roche J, Blanca F, Amela C, Sanchez A, Alvar J: Baseline entomological data for a pilot malaria control program in Equatorial Guinea. J Med Entomol. 1993, 30: 622-624.
Munga S, Yakob L, Mushinzimana E, Zhou G, Ouna T, Minakawa N, Githeko A, Yan G: Land use and land cover changes and spatiotemporal dynamics of anopheline larval habitats during a four-year period in a highland community of Africa. Am J Trop Med Hyg. 2009, 81: 1079-1084. 10.4269/ajtmh.2009.09-0156.
Muriu SM, Muturi EJ, Shililu JI, Mbogo CM, Mwangangi JM, Jacob BG, Irungu LW, Mukabana RW, Githure JI, Novak RJ: Host choice and multiple blood feeding behaviour of malaria vectors and other anophelines in Mwea rice scheme, Kenya. Malar J. 2008, 7: 43-10.1186/1475-2875-7-43.
Muturi EJ, Shililu J, Jacob B, Gu W, Githure J, Novak R: Mosquito species diversity and abundance in relation to land use in a riceland agroecosystem in Mwea, Kenya. J Vector Ecol. 2006, 31: 129-137. 10.3376/1081-1710(2006)31[129:MSDAAI]2.0.CO;2.
Nigatu W, Petros B, Lulu M, Adugna N, Wirtz R: Species composition, feeding and resting behaviour of the common anthropophilic anopheline mosquitoes in relation to malaria transmission in Gambella, south west Ethiopia. Insect Sci Appl. 1994, 15: 371-377.
Midega JT, Mbogo CM, Mwambi H, Wilson MD, Ojwang G, Mwangangi JM, Nzovu JG, Githure JI, Yan G, Beier JC: Estimating dispersal and survival of Anopheles gambiae and Anopheles funestus along the Kenyan coast by using mark-release-recapture methods. J Med Entomol. 2007, 44: 923-929. 10.1603/0022-2585(2007)44[923:EDASOA]2.0.CO;2.
Minakawa N, Munga S, Atieli F, Mushinzimana E, Zhou G, Githeko AK, Yan G: Spatial distribution of anopheline larval habitats in Western Kenyan highlands: effects of land cover types and topography. Am J Trop Med Hyg. 2005, 73: 157-165.
Minakawa N, Sonye G, Dida GO, Futami K, Kaneko S: Recent reduction in the water level of Lake Victoria has created more habitats for Anopheles funestus. Malar J. 2008, 7: 119-10.1186/1475-2875-7-119.
Manoukis NC, Toure MB, Sissoko I, Doumbia S, Traore SF, Diuk-Wasser MA, Taylor CE: Is vector body size the key to reduced malaria transmission in the irrigated region of Niono, Mali?. J Med Entomol. 2006, 43: 820-827. 10.1603/0022-2585(2006)43[820:IVBSTK]2.0.CO;2.
Howard AF, Zhou G, Omlin FX: Malaria mosquito control using edible fish in western Kenya: preliminary findings of a controlled study. BMC Public Health. 2007, 7: 199-10.1186/1471-2458-7-199.
Ijumba JN, Mosha FW, Lindsay SW: Malaria transmission risk variations derived from different agricultural practices in an irrigated area of northern Tanzania. Med Vet Entomol. 2002, 16: 28-38. 10.1046/j.0269-283x.2002.00337.x.
Koudou BG, Tano Y, Doumbia M, Nsanzabana C, Cisse G, Girardin O, Dao D, N'Goran EK, Vounatsou P, Bordmann G, Keiser J, Tanner M, Utzinger J: Malaria transmission dynamics in central Côte d'Ivoire: the influence of changing patterns of irrigated rice agriculture. Med Vet Entomol. 2005, 19: 27-37. 10.1111/j.0269-283X.2005.00530.x.
Kulkarni MA, Kweka E, Nyale E, Lyatuu E, Mosha FW, Chandramohan D, Rau ME, Drakeley C: Entomological evaluation of malaria vectors at different altitudes in Hai district, northeastern Tanzania. J Med Entomol. 2006, 43: 580-588. 10.1603/0022-2585(2006)43[580:EEOMVA]2.0.CO;2.
Dia I, Lochouarn L, Diatta M, Sokhna CS, Fontenille D: Préférences trophiques des femelles endophiles d'Anopheles funestus au Sénégal. Bull Soc Pathol Exot. 2001, 94: 210-213.
Adja AM, N'Goran KE, Kengne P, Koudou GB, Toure M, Koffi AA, Tia E, Fontenille D, Chandre F: Transmission vectorielle du paludisme en savane arborée à Ganse en Côte d'Ivoire. Med Trop (Mars). 2006, 66: 449-455.
Ramphul U, Boase T, Bass C, Okedi LM, Donnelly MJ, Muller P: Insecticide resistance and its association with target-site mutations in natural populations of Anopheles gambiae from eastern Uganda. Trans R Soc Trop Med Hyg. 2009, 103: 1121-6. 10.1016/j.trstmh.2009.02.014.
Kaufmann C, Briegel H: Flight performance of the malaria vectors Anopheles gambiae and Anopheles atroparvus. J Vector Ecol. 2004, 29: 140-153.
Carnevale P, Boreham PFL: Etudes des préférences trophiques d' Anopheles nili (Theo.), 1904. Cahiers ORSTOM, Entomol Med Parasitol. 1978, 16: 17-22.
Dia I, Diallo D, Duchemin JB, Ba Y, Konate L, Costantini C, Diallo M: Comparisons of human-landing catches and odor-baited entry traps for sampling malaria vectors in Senegal. J Med Entomol. 2005, 42: 104-109. 10.1603/0022-2585(2005)042[0104:COHCAO]2.0.CO;2.
Patsoula E, Samanidou-Voyadjoglou A, Spanakos G, Kremastinou J, Nasioulas G, Vakalis NC: Molecular characterization of the Anopheles maculipennis complex during surveillance for the 2004 Olympic Games in Athens. Med Vet Entomol. 2007, 21: 36-43. 10.1111/j.1365-2915.2007.00669.x.
Beier JC, Zimmerman JH, Kenawy MA, el Said S, Abbassy MM: Host-feeding patterns of the mosquito community (Diptera: Culicidae) in two Faiyum Governorate villages, Egypt. J Med Entomol. 1987, 24: 28-34.
Kenawy M, Zimmerman JH, Beier JC, el Said S, Abbassy MM: Host-feeding patterns of Anopheles sergentii and An. multicolor (Diptera: Culicidae) in Siwa and El Gara oases, Egypt. J Med Entomol. 1986, 23: 576-577.
Matthys B, Sherkanov T, Karimov SS, Khabirov Z, Mostowlansky T, Utzinger J, Wyss K: History of malaria control in Tajikistan and rapid malaria appraisal in an agro-ecological setting. Malar J. 2008, 7: 217-10.1186/1475-2875-7-217.
Rowland M, Mohammed N, Rehman H, Hewitt S, Mendis C, Ahmad M, Kamal M, Wirtz R: Anopheline vectors and malaria transmission in eastern Afghanistan. Trans R Soc Trop Med Hyg. 2002, 96: 620-626. 10.1016/S0035-9203(02)90331-7.
Amr ZS, al-Khalili Y, Arbaji A: Larval mosquitoes collected from northern Jordan and the Jordan Valley. J Am Mosq Control Assoc. 1997, 13: 375-378.
Acknowledgements
We wish to thank Rosalind Howes, Edward Haynes, Philip Mbithi, Owen Yang, Carolynn Tago, and Elisabeth Thiveyrat for primary data abstraction. We also thank the Technical Advisory Group for extended support over the duration of the project (in addition to co-authors Michael Bangs, Sylvie Manguin, Maureen Coetzee, Ralph Harbach, Janet Hemingway and Charles M. Mbogo, these include, Theeraphap Chareonviriyaphap and Yasmin Rubio-Palis). MES is funded by a project grant from the Wellcome Trust (#083534) to SIH. SIH is funded by a Senior Research Fellowship from the Wellcome Trust (#079091) which also supports CWK and PWG. APP and WHT are funded by a Wellcome Trust Principal Research Fellowship (#079080) to Professor Robert Snow. This work forms part of the output of the Malaria Atlas Project (MAP, http://www.map.ox.ac.uk), principally funded by the Wellcome Trust, U.K.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
SIH conceived the study and managed its design and implementation. MES wrote the first draft of the manuscript and assembled the occurrence data with assistance from CWK and RMO, CWK also digitised and edited all the expert opinion maps. WHT designed and maintained the databases and implemented the map figures. APP implemented the BRT scripts for predictive mapping. PWG processed the environmental and climatic data grids, with assistance from TVB. All TAG members (MJB, SM, MC, CMM and JH) provided data and advice in updating the EO range maps. Experiments were devised by SIH and MES and implemented by MES. All authors participated in the interpretation of results and in the writing and editing of the manuscript. MJB, MC, SM, HCJG, CMM and REH advised on bionomics and nomenclature issues, and provided additional comments and input to the manuscript.
Electronic supplementary material
13071_2010_245_MOESM1_ESM.ZIP
Additional file 1: Expert opinion distribution maps for the seven DVS of Africa and the six DVS of the Europe and Middle Eastern region. (ZIP 4 MB)
13071_2010_245_MOESM2_ESM.DOC
Additional file 2: Summary tables showing evaluation statistics for all mapping trials and final Boosted Regression Tree environmental and climatic variable selections for the final, optimal predictive maps. (DOC 398 KB)
13071_2010_245_MOESM3_ESM.PDF
Additional file 3: Predictive species distribution maps for the seven DVS of Africa and the six DVS of the Europe and Middle Eastern region. (PDF 3 MB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Sinka, M.E., Bangs, M.J., Manguin, S. et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasites Vectors 3, 117 (2010). https://doi.org/10.1186/1756-3305-3-117
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1756-3305-3-117