
Abstract
Head
Building synthetic gene networks with highly transient dynamics requires rapid protein degradation. We show that the degradation conferred by two commonly used ssrA tags is highly temperature dependent. Synthetic gene networks are being used increasingly in real-world applications where they may be subjected to variable conditions, and be required to display precise, quantitative dynamics, which will be more susceptible to environmental changes than the general qualitative dynamics focussed on so far.
Similar content being viewed by others


Introduction
Rapid protein degradation is vital for the creation of synthetic gene networks with highly transient dynamics. The standard means of achieving rapid protein degradation in bacterial synthetic gene networks has been through the use of ssrA tags. These are short (usually 11 amino acid) peptide tags added cotranslationally to the C-terminus of a nascent protein, that are recognised by the AAA+ ClpXP and ClpAP host proteases[1] and target the tagged protein for degradation[2, 3]. ssrA tags are part of a larger family of degradation tags, or ‘degrons’, which can be present in both the N- and C- terminus of proteins[4]. The ssrA tags are generally referred to by the final three amino acids, for instance the wild-type tag ends in the amino acid sequence LAA and is known as the LAA-ssrA tag. ssrA tags have been used frequently in synthetic oscillators[5–8].
Synthetic biology is increasingly moving towards real-world applications[9] in which synthetic gene networks will not always be present at favourable laboratory conditions. Networks designed under one set of conditions (growth rate, nutrient supply, temperature etc) will not necessarily function as designed under another set of conditions. Networks must therefore be designed to function correctly, or at least predictably, in sub-optimal and changeable environments. A robustness to temperature variation will need to form part of this, as demonstrated by the observation of a substantial effect of temperature on a synthetic gene oscillator[6]. It has recently been shown that the bacterial chemotaxis machinery is highly temperature compensated[10]. Therefore a relevant question is whether the bacterial degradation machinery also exhibits a significant level of temperature compensation. This is important not only for synthetic biologists investigating the effects of temperature on gene network performance, but also for those studying the degradation machinery itself.
Here we examine the degradation of LacI, a protein frequently used in bacterial synthetic gene networks[5, 6, 8, 11–13], tagged with either the LAA or LVA ssrA tag. We present results that demonstrate a marked temperature dependence in the degradation rate conferred by both tags.
Results and discussion
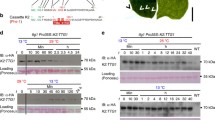
Figure1 shows the amount of LacI, tagged with either an LAA (LacI-LAA) or LVA (LacI-LVA) ssrA tag, or left untagged, from E. coli cells grown at 37 or 25°C, before the addition of chloramphenicol (used to stop translational elongation) (0 minutes) and at 10, 20, 30, 45 and 60 minutes post addition of chloramphenicol.

Degradation of ssrA-tagged and untagged LacI at 25°C and 37°C. This figure shows representative western blots and quantification of the levels of LacI at 0 minutes (prior to the addition of chloramphenicol) and 10-60 minutes after addition of chloramphenicol in E. coli M182 grown at 25°C (squares) and 37°C (diamonds). Sample loading was normalised by culture OD. LacI levels are shown relative to the amount present at t=0, are the mean of three independent experiments and are shown with standard error.
LacI carrying an LAA or LVA tag was degraded approximately 3-5 fold faster in cells grown at 37°C than in cells grown at 25°C. The apparent half-life of LacI-LAA increased from ∼8 minutes at 37°C to ∼38 minutes at 25°C and the apparent half-life of LacI-LVA increased from ∼13 minutes to ∼41 minutes. The stability of untagged LacI was unaffected by temperature, indicating that the observations were dependent on the presence of the ssrA tags.
The result presented here demonstrates that degradation conferred by both LAA and LVA ssrA tags is substantially affected by temperature, and shows that little or no mechanism for temperature compensation exists in this system. Degradation of some tagged cellular proteins has been shown to be temperature dependent[14], but to the best of our knowledge the effect of tempe- rature on the stability of proteins tagged with ssrA tags commonly used within synthetic biology has not been previously investigated. This temperature dependence has been demonstrated for LacI, and there is no reason to assume that it will not extend to other proteins.
This result is potentially important for the future construction of synthetic gene networks in bacteria. Future networks will be increasingly designed for real-world applications where they may need to function over a range of temperatures and conditions. This result highlights a limitation of the current means of tuning protein degradation within bacterial synthetic gene networks and suggests that new, perhaps synthetic, protein degradation pathways that are robust to environmental (and physiological) variations need to be developed.
Materials and methods
E. coli strain M182[15] was transformed with one of three plasmids containing lacI tagged with either an LAA (amino acid sequence: AANDENYALAA. DNA sequence: GCTGCTAACGACGAAAACTACGCTCTGGCTGCT) or LVA (amino acid sequence: AANDENYALVA. DNA sequence: GCTGCAAACGACGAAAACTACGCTTTA- GTAGCT) ssrA tag, or left untagged, referred to as pOP_L-LAA, pOP_L-LVA and pOP_L-NoTg respectively. The plasmids were constructed by cloning lacI and its two tagged variants into an expression cassette comprising the Prrn promoter and the U0 RBS region[16], upstream of the lacI coding region, and the terminator BBa_B0015[17] downstream. The plasmid backbone was pSB6A1[18]. Transformed cells were grown overnight on LB agar with ampicillin (100μ g/ml). Colonies were grown in 5 ml of EZ rich media (Teknova, CA) overnight. 0.5 ml of culture was added to 25 ml EZ rich media, and grown at both 25 and 37°C with shaking (250 rpm) to OD600 of 0.4-0.6. A 1 ml sample was taken, centrifuged at 11,700x g and supernatant removed. The pellet was frozen in liquid nitrogen. Chloramphenicol was added to the cultures to a final concentration of 136 μ g/ml. 1 ml samples were taken at subsequent time points of 10, 20, 30, 45 and 60 minutes. The same procedure for the initial time point sample was followed. Samples were stored at -80°C.
To assay LacI abundance, SDS loading buffer was added to samples in an amount relative to the OD of the sample. Samples were heated for 3 minutes at 95°C, before being analysed by SDS-PAGE and western blotting, using a mouse monoclonal antibody against LacI (Abcam) and a goat anti-mouse IgG-HRP conjugated secondary antibody (Santa Cruz Biotechnology). Blots were quantified using ImageJ software.
References
Gottesman S: Proteolysis in bacterial regulatory circuits. Annu Rev cell Dev Biol 2003, 19: 565-587. 10.1146/annurev.cellbio.19.110701.153228
Oh BK, Chauhan AK, Isono K, Apirion D: Location of a gene (ssrA) for a small, stable RNA (10Sa RNA) in the Escherichia coli chromosome. J Bacteriol 1990,172(8):4708-4709.
McGinness K, Baker T, Sauer R: Engineering controllable protein degradation. Mol Cell 2006, 22: 701-707. 10.1016/j.molcel.2006.04.027
Flynn JM, Neher SB, Kim YI, Sauer RT, Baker TA: Proteomic discovery of cellular substrates of the ClpXP protease reveals five classes of ClpX-recognition signals. Mol Cell 2003,11(3):671-683. 10.1016/S1097-2765(03)00060-1
Mondragón-Palomino O, Danino T, Selimkhanov J, Tsimring L, Hasty J: Entrainment of a population of synthetic genetic oscillators. Science 2011,333(6047):1315-1319. 10.1126/science.1205369
Stricker J, Cookson S, Bennett MR, Mather WH, Tsimring LS, Hasty J: A fast, robust and tunable synthetic gene oscillator. Nature 2008,456(7221):516-U39. 10.1038/nature07389
Fung E, Wong WW, Suen JK, Bulter T, Lee Sg, Liao JC: A synthetic gene-metabolic oscillator. Nature 2005,435(7038):118-122. 10.1038/nature03508
Elowitz MB, Leibler S: A synthetic oscillatory network of transcriptional regulators. Nature 2000,403(6767):335-338. 10.1038/35002125
Khalil AS, Collins JJ: Synthetic biology: applications come of age. Nat Rev Genet 2010,11(5):367-379.
Oleksiuk O, Jakovljevic V, Vladimirov N, Carvalho R, Paster E, Ryu WS, Meir Y, Wingreen NS, Kollmann M, Sourjik V: Thermal Robustness of Signaling in Bacterial Chemotaxis. Cell 2011,145(2):312-321. 10.1016/j.cell.2011.03.013
Gardner TS, Cantor CR, Collins JJ: Construction of a genetic toggle switch in Escherichia coli. Nature 2000,403(6767):339-342. 10.1038/35002131
Lou C, Liu X, Ni M, Huang Y, Huang Q, Huang L, Jiang L, Lu D, Wang M, Liu C, Chen D, Chen C, Chen X, Yang L, Ma H, Chen J, Ouyang Q: Synthesizing a novel genetic sequential logic circuit: a push-on push-off switch. Mol Syst Biol 2010, 6: 350.
Basu S, Gerchman Y, Collins CH, Arnold FH, Weiss R: A synthetic multicellular system for programmed pattern formation. Nature 2005,434(7037):1130-1134. 10.1038/nature03461
Herman C, Thévenet D, Bouloc P, Walker GC, D’Ari R: Degradation of carboxy-terminal-tagged cytoplasmic proteins by the Escherichia coli protease HflB (FtsH). Genes & development 1998,12(9):1348-1355. 10.1101/gad.12.9.1348
Casadaban M, Cohen S: Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J Mol Biol 1980,138(2):179-207. 10.1016/0022-2836(80)90283-1
Neupert J, Karcher D, Bock R: Design of simple synthetic RNA thermometers for temperature-controlled gene expression in Escherichia coli. Nucleic Acids Res 2008,36(19):e124. 10.1093/nar/gkn545
BBa_B0015 registry information (MIT parts registry) [. http://partsregistry.org/Part:BBa_B0015] [].
pSB6A1 registry information (MIT parts registry) [. http://partsregistry.org/Part:pSB6A1] [].
Acknowledgements
Funding was provided by the Bristol Centre for Complexity Sciences through EPSRC-GB Grant No. EP/E501214/1.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors’ contributions
OP, NJS, CSG and MdB conceived the experiments. OP carried out the experiments. OP and NJS wrote the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Purcell, O., Grierson, C.S., Bernardo, M.d. et al. Temperature dependence of ssrA-tag mediated protein degradation. J Biol Eng 6, 10 (2012). https://doi.org/10.1186/1754-1611-6-10
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1754-1611-6-10