
Abstract
Background
Control and eradication of intestinal infections caused by protozoa are important biomedical challenges worldwide. Prophylactic control of coccidiosis has been achieved with the use of anticoccidial drugs; however, the increase in anticoccidial resistance has raised concerns about the need for new alternatives for the control of coccidial infections. In fact, new strategies are needed to induce potent protective immune responses in neonatal individuals.
Methods
The effects of a dietary supplementation of mannan-oligosaccharide (yeast cell wall; YCW) on the local, humoral and cell-mediated immune responses, and intestinal replication of coccidia were evaluated in a neonatal animal model during natural exposure to Eimeria spp. A total of 840 one-day-old chicks were distributed among four dietary regimens: A) Control diet (no YCW) plus anticoccidial vaccine); B) Control diet plus coccidiostat; C) YCW diet plus anticoccidial vaccination; and D) YCW diet plus coccidiostat. Weight gain, feed consumption and immunological parameters were examined within the first seven weeks of life.
Results
Dietary supplementation of 0.05% of YCW increased local mucosal IgA secretions, humoral and cell-mediated immune responses, and reduced parasite excretion in feces.
Conclusion
Dietary supplementation of yeast cell wall in neonatal animals can enhance the immune response against coccidial infections. The present study reveals the potential of YCW as adjuvant for modulating mucosal immune responses.
Similar content being viewed by others
Background
Control and eradication of intestinal infections caused by protozoa are important biotechnological and medical challenges worldwide. The coccidial parasites Eimeria, Toxoplasma, Neospora and Cryptosporidium are the cause of serious diseases in humans and animals. Many of these intracellular protozoa affect the small intestine and produce devastating effects on immune compromised subjects [1–3]. These parasitic infections can cause a wide range of harm to the infected host; from diarrhea to abortion, death or chronic nutritional and cognitive sequels [4, 5]. Coccidial infections have been controlled, to a great extent, with the use of anticoccidial drugs; however, the increase in resistance to many of these products has raised concerns about the need for new anticoccidial alternatives [4]. A challenging factor is that protozoan parasites multiply inside of host cells; thus, cell mediated immune responses are required for protective immunity [4, 5]. Therefore, development of effective anticoccidial strategies requires substantial knowledge of the parasite biology and host-parasite interactions [4].
Progress on new anticoccidial strategies has been accomplished in livestock and poultry. In these animal species, protozoan intestinal infections are an important factor associated with poor performance and production efficiency [4, 6, 7]. Avian coccidiosis, caused by the apicomplexan protozoan Eimeria, is the major parasitic disease of poultry. For the world poultry industry, the cost of coccidiosis excess USD 3 billion annually [6].
Currently, prophylactic protection against coccidiosis in humans and animals can be accomplished with the use of anticoccidial vaccines. However, the use of live vaccines has been gradually adopted mainly due to economic reasons such as elevated cost of vaccination and adverse effects on early growth [4, 8–10]. Even though research and development of effective vaccine formulations are still in progress [11, 12], there is a need for new studies focused on the design of novel strategies to induce potent protective immune responses without adverse effects in neonate or immune-compromised individuals [4, 13, 14]. Here, a neonatal animal model was used to examine the effect of a dietary supplementation of mannan-oligosaccharide (in the form of yeast cell wall (YCW) on the local and humoral immune responses in neonatal chickens during natural exposure to coccidia parasite. This study revealed that dietary supplementation of 0.05% of YCW increases local, cellular and humoral immune responses, and reduces parasite excretion in feces.
Methods
Experimental design and animals
A total of 840 one-day-old Ross 308 chicks were distributed randomly in 28 pens bedded with fresh wood shavings (30 chickens per pen; 1:1 female/male ratio) in an experimental poultry house. The temperature was 32°C in each room on day 1 and was decreased weekly until the minimum temperature of 21°C was reached. Chickens were fed conventional starter (1 to 21 days old) and finisher (22 to 49 days) sorghum/soy based diets that met National Research Council requirements (Nutrient requirement of poultry, 1994). Fluorescent lighting was provided under the following schedule: day 0–2: 23 h light and 1 h dark, day 3–11: 18 h light and 6 h dark; day 12-end: 16 h light and 8 h dark. Chicks were allowed to consume feed and water ad libitum during the entire time of the experiment. The experiments were performed under poultry farm conditions, where animals are naturally exposed to different species of Eimeria (i.e. E. acervulina, E. maxima and E. tenella; [13]). Experimental procedures were approved by the Institutional Committee for Care and Use of Experimental Animals at the Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México.
Basal diets were formulated without antibiotics and were identical across treatments with the following exceptions: treatments B and D included coccidiostat [125 ppm of nicarbazin (Helm, Naucalpan, Mexico) for starter diet and 65 ppm of salinomycin (Helm, Naucalpan, Mexico) for finisher diet]. Treatments C and D included 0.05% (w/w) of YCW (a concentrate of yeast hulls obtained via autolysis of Saccharomyces cerevisiae (Safmex, Toluca, Mexico). At arrival, chicks were randomly divided among treatments. The experimental design was a factorial arrangement of treatments, consisting of four dietary regimens: A) Control diet (no YCW) plus anticoccidial vaccine); B) Control diet (no YCW) plus coccidiostat; C) YCW diet plus anticoccidial vaccination; and D) YCW diet plus coccidiostat [7 replicates (pens) of each group, for a total of 210 animals per treatment]. Animals from the vaccinated groups were individually vaccinated at day 1 of age with a single oral dose of a commercial vaccine against coccidia (Advent cocci-vaccine; Viridus Animal Health LLC – Novus International Inc., St. Louis, MO) following the manufacturer's specifications. Animal weight gain and feed consumption were recorded at 21 and 49 days of age. At the end of the experimental period (49 days), 20 birds per treatment were euthanized by cervical dislocation. Carcass yield and breast skin pigmentation (yellow pigmentation after cold processing analyzed by reflectance colorimetric, Minolta-CR 400) were measured.
Assessment of the humoral immune response
The effect of different treatments on humoral immune response was examined by means of antibody production after a simultaneous vaccination against Newcastle disease virus (NDV). Ocular (eye drop 0.03 ml active virus, Nobilis ND Lasota, Intervet/Schering-Plough Animal Health; Mexico) and subcutaneous (0.25 ml inactivated virus, Intervet/Schering-Plough Animal Health) vaccines were administered at 10 days of age. Serum samples were taken at 7 and 14 days post-vaccination (35 samples per treatment), frozen at -20°C, and specific antibody titers against NDV were determined by hemagglutination inhibition assay [15].
Assessment of the mucosal immune response
The effect of different treatments on mucosal immune response was examined by means of production of secretory immunoglobulin A (IgA). Duodenum and tracheal samples (10 chickens per treatment) were collected at 21 days of age to measure production of secretory IgA. Ten centimeters of duodenum were taken and mucosal lavages were performed with 10 ml of cold and sterile phosphate-buffered saline solution (PBS). Tracheal samples were collected by tracheal swabs and placed (12 hrs) in tubes containing 10 ml of sterile PBS. All mucosal samples were clarified by centrifugation (1200 × g) and frozen at -20°C. Secretory IgA concentrations were measured by enzyme-linked immunosorbent assay (ELISA) test (chicken IgA ELISA Quantitation Kit, Bethyl Laboratories Inc. Montgomery, TX) following the manufacturer's protocol [16].
Assessment of the cell-mediated immune response
The effect of different treatments on cell-mediated immune response was examined by cutaneous basophilic hypersensitivity test [17, 18]. Analysis of cell-mediated immune response at 21 days was performed as descried elsewhere [17]. Briefly, an intradermic inoculation of phytohemagglutinin (PHA-A3; 0.1 mg/0.1 ml) was carried out in the inter-digital membrane of 3rd and 4rd phalanges of the right inferior extremity and sterile saline solution (0.1 ml) of the left foot (35 chickens per treatment). Twenty-four hours post-inoculation the thickness of the inter-digital membrane was measured in millimeters using a micrometer.
Assessment of the replication of coccidia in the intestinal tract
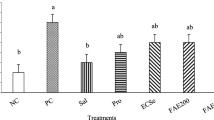
The effect of different treatments on the replication of coccidia in the intestinal tract was estimated by quantifying the number of oocysts excreted in feces. At 21 days of age, ten birds per treatment were placed in separate cages with clean sheets of paper on the bottom. Fecal samples were collected from the paper lining after an 8-hour period. Birds were returned to their respective pens at the end of the fecal collection period. The total number of oocysts was calculated following the method described previously [19].
Statistical analysis
Results were statistically analyzed by ANOVA in a complete randomized design with 2 × 2 factorial arrangement and by the GLM procedure of the SAS software (SAS Institute, Cary, NC). Data of animal performance and immunological parameters were grouped and presented as mean ± SEM. P-values ≤ 0.05 were considered statistically significant. Two factors were examined and compared across the study, 1) diet: control sorghum/soy-based diet (Control) vs. YCW diet (YCW); and 2) anticoccidial program: anticoccidial vaccine (Vaccine) vs. coccidiostat drug (Coccidiostat).
Results
Animal growth
The effects of diet and anticoccidial program on animal performance at 21 and 49 days of age are described in Table 1 and 2, respectively. At 21 days of age (Table 1), YCW increased weight gain and feed consumption compared to Control (P < 0.01). However, no significant differences in feed conversion were observed between Control and YCW (P > 0.05). Also, it was found that Coccidiostat increased weight gain and feed consumption compared to Vaccine (P < 0.01), and that Vaccine increased feed conversion (P < 0.05). These results reveal the common adverse effects on growth of chicks caused by anticoccidial vaccination [8]. In the analysis at 49 days of age (Table 2), YCW increased weight gain and reduced feed conversion compared to Control (P < 0.05). There were no differences in percentage of carcass yield and skin pigmentation between Control and YCW (P > 0.05; data not shown). Also, there were no differences (P > 0.05) in weight gain, feed consumption, feed conversion (Table 2), percentage of carcass yield and skin pigmentation between Coccidiostat and Vaccine factors at day 49 of age (data not shown).
Assessment of humoral immune response
The effect of different treatments on the humoral immune response was examined by means of antibody production after a simultaneous vaccination against NDV. Ocular and subcutaneous vaccines were administered at 10 days of age and antibody production was measured at 7 and 14 days post-vaccination (Table 3). At 14 days of age, YCW induced higher NDV antibody titers compared to Control (P < 0.01). There were no differences in antibody titers between Vaccine and Coccidiostat (P > 0.05). The immune-regulatory effect was also observed at 21 days of age. YCW induced the higher NDV antibody titers compared to Control (P < 0.01). No significant effects were observed between Vaccine and Coccidiostat factors (P > 0.05). These results reveal that YCW enhances the humoral immune response against vaccine antigens in young animals.
Assessment of mucosal immune response
The effect of different treatments on mucosal immune response was examined by means of production of secretory IgA. Results of the IgA concentrations are presented in Table 3. In duodenal samples, YCW and Vaccine increased mucosal concentrations of intestinal IgA compared to Control and Coccidiostat, respectively (P < 0.01). In tracheal samples, YCW increased concentrations of mucosal IgA compared to Control (P < 0.05). No significant differences were observed between Vaccine and Coccidiostat.
Assessment of cell-mediated immune response
The effect of different treatments on cell-mediated immune response was examined by the cutaneous basophilic hypersensitivity test. This method reveals the status of T-cell response. Results of the cell-mediated immune response at 21 days are presented in Table 4. YCW increased cell-mediated immune response compared to Control (P < 0.05). No significant differences for this parameter were observed between Vaccine and Coccidiostat (P > 0.05). These results confirm the systemic immune-regulatory effects of YCW and reveal that this compound enhances cell-mediated immune responses in young animals.
Assessment of the replication of coccidia in the intestinal tract
The effect of the different treatments on the replication of coccidia in the intestinal tract was verified by quantifying the number of oocysts excreted in feces. Results of the oocyst count at 21 days of age are presented in Table 4. YCW reduced the number of oocysts excreted in feces compared to Control (P < 0.01). Also, as expected, Coccidiostat reduced oocyst excretion compared to Vaccine (P < 0.05). These results reveal that the immune-regulatory effects produced by YCW reduce the replication of coccidia in the intestinal tract of young animals.
Discussion
The development of new strategies for control of coccidiosis is essential for the poultry industry. Chickens are highly at risk for coccidial infections due to environmental conditions during production. High animal densities (> 25,000 chickens per building) on floor pens and warm surroundings are favorable for a high transmission, replication and accumulation of Eimeria spp. [13]. Moreover, the current practices for animal production create a strong selective pressure on coccidia parasites to develop anticoccidial drug resistance [20]. Together, these factors make neonatal chicken an excellent animal model for the study and design of new and effective strategies for the control of coccidiosis.
A current strategy for the control of avian coccidiosis is the use of live anticoccidial vaccines; however, this approach has been gradually accepted because of high cost of the vaccine and some adverse effects on early animal growth observed after vaccination [8]. In the present study, the vaccinated group showed a negative effect on chicken growth rates during the first 3-weeks of age; nonetheless, by 7-weeks of age, this group had a compensatory growth and the animal performance was similar among all groups by the end of the experiment. In addition, the data of the present study revealed that live-attenuated Eimeria spp. strains contained in the anticoccidial vaccine were effective to replace intestinal replication and spreading of wild Eimeria spp. strains. This notion is supported by the fact that vaccinated birds excreted high numbers of oocysts in feces with no effects on animal productivity. Although the beneficial effects of anticoccidial vaccines in poultry are well documented, the high costs associated with this prophylactic approach limit its widespread use in animal production [8]. To this end, the design of novel and not expensive alternatives for the control of coccidiosis is an important task to maintain the productivity of the poultry industry.
The present study revealed that dietary supplementation of 0.05% of YCW increases local mucosal IgA secretions, cellular and humoral immune responses, and reduces parasite excretion in feces of neonatal animals infected with coccidia. These results provide clear evidence of the potential of this dietary strategy as an anticoccidial alternative. Specifically, this study reveals the potential of YCW dietary supplementation to enhance cell mediated immune responses, an important factor for the protection against coccidial infections [4, 5, 21].
In the present study, dietary supplementation of YCW improved weight gain and feed conversion, and reduces parasite excretion in feces of animals naturally infected with coccidia. Only a few studies have examined the effect of YCW on coccidial infections. Chickens supplemented with yeast-culture residues in the diet and raised on coccidia-infected litters [22]; and chickens fed with YCW and experimentally infected with Eimeria spp. [23] showed improved growth performance compared to control treatments. Results of all three reports consistently indicate that dietary supplementation of YCW or derivates could be an important alternative for reducing performance losses provoked by coccidial infection.
The effects of diets supplemented with YCW on systemic and local immune responses have been demonstrated in many animal models including cows [24], pigs [25, 26], dogs [27, 28], rats [29], chickens [30, 31], rainbow trout [32], etc. Likewise, it has been shown that YCW supplemented diets boost immune responses against vaccine antigens; for example, in cows, YCW enhanced humoral and cellular immune responses against rotavirus vaccine [24]. In piglets, YCW improved humoral and cellular immune responses against Aujeszky's disease vaccination [25]. Interestingly, in humans, oral supplementation of a yeast-based product significantly reduced the incidence and duration of influenza outbreaks [33]. In the present study, dietary supplementation of YCW increased the production of antibodies against NDV and IgA concentrations in the intestinal and tracheal mucosa. Moreover, YCW supplementation reduced the number of Eimeria spp. oocysts excreted in feces. A similar effect on parasite oocysts excretion was reported elsewhere [34]. Although the specific mechanisms of action by intestinal IgA on coccidial infections are still unknown, it is thought that secretory IgA attaches to the parasite surface and prevents its binding to the intestinal epithelium. Also IgA reduces the development of sporozoites or merozoites and prevents invasion within host cells [35, 36]. Interestingly, the present study also revealed that the YCM treatment increased IgA secretions in both the intestinal and tracheal tissues while the vaccine treatment increased this antibody only in the intestinal samples. These observations indicate that YCM treatment stimulate IgA secretions systemically while the vaccine treatment stimulate secretions only in tissues exposed to the parasite. These results, taken together, provide new and clear evidence that YCW has an immune-regulatory effect at the local and systemic level.
It can be hypothesized that the increase in humoral responses induced by YCW supplementation are responsible for the reduction in coccidia replication in the intestinal mucosa [37]. However, recent evidence revealed that humoral immunity plays only a minor role in protection against this disease [4, 35]. Cell-mediated immune responses are thought to be the most important factor for protection against coccidiosis. CD4+ and CD8+ T-cells populations limit coccidia replication in the intestinal tract [4, 35, 38]. The effects of T-cells on Eimeria spp. are mediated by the cytokines that these cells release. Interferon-gamma and tumor necrosis factor-alpha limit oocyst production in either primary or secondary infections (for details see reviews [4, 35, 38]). The immune-regulatory effects of YCW on cell-mediated responses have been confirmed in several human and animal studies [39–42]. Therefore, it is possible to hypothesize that the reduction in Eimeria spp. replication in the intestine was caused by the YCW-induction of T-cell responses. Even though T-cells phenotypes were not directly examined in the present study, cutaneous basophilic hypersensitivity test is an excellent and well accepted method for the determination of T-cell response [17, 18, 43, 44]. To our knowledge this study provides the first evidence of the effect of YCW on regulation of T-cell responsiveness, which could be important for immunity against coccidial infections. Further studies will elucidate T-cell repertoire and cytokines responses stimulated by YCW dietary supplementation.
Conclusion
The present data show that dietary supplementation of YCW in neonatal animals can enhance the immune responses against coccidial infections. Moreover, the present study provides a new dietary strategy for the alternative control of coccidiosis and more importantly, confirms the potential of YCW as adjuvant for modulating mucosal immune responses [45]. Despite the difficulties of designing effective approaches for the control of coccidiosis, this study provides a promising dietary strategy with the potential to enhance the health of immune-compromised subjects and children in developing countries.
Abbreviations
- ANOVA:
-
analysis of variance
- ELISA:
-
enzyme-linked immunosorbent assay
- IgA:
-
secretory immunoglobulin A
- NDV:
-
Newcastle disease virus
- NRC:
-
National Research Council
- PBS:
-
phosphate-buffered saline solution
- PHA-A3:
-
phytohemagglutinin
- SEM:
-
standard error of the mean
- YCW:
-
yeas cell wall.
References
Brandonisio O: Waterborne transmission of Giardia and Cryptosporidium. Parassitologia. 2006, 48: 91-94.
Farthing MJ: Treatment options for the eradication of intestinal protozoa. Nat Clin Pract Gastroenterol Hepatol. 2006, 3: 436-445. 10.1038/ncpgasthep0557.
Schito ML, Barta JR, Chobotar B: Comparison of four murine Eimeria species in immunocompetent and immunodeficient mice. J Parasitol. 1996, 82: 255-262. 10.2307/3284157.
Innes EA, Vermeulen AN: Vaccination as a control strategy against the coccidial parasites Eimeria, Toxoplasma and Neospora. Parasitology. 2006, 133 (Suppl): S145-168. 10.1017/S0031182006001855.
Pantenburg B, Dann SM, Wang HC, Robinson P, Castellanos-Gonzalez A, Lewis DE, White AC: Intestinal immune response to human Cryptosporidium sp. infection. Infect Immun. 2008, 76: 23-29. 10.1128/IAI.00960-07.
Dalloul RA, Lillehoj HS: Poultry coccidiosis: recent advancements in control measures and vaccine development. Expert Rev Vaccines. 2006, 5: 143-163. 10.1586/14760584.5.1.143.
Daugschies A, Najdrowski M: Eimeriosis in cattle: current understanding. J Vet Med B Infect Dis Vet Public Health. 2005, 52: 417-427.
Williams RB: Anticoccidial vaccines for broiler chickens: pathways to success. Avian Pathol. 2002, 31: 317-353. 10.1080/03079450220148988.
Lavine MD, Arrizabalaga G: Invasion and egress by the obligate intracellular parasite Toxoplasma gondii: potential targets for the development of new antiparasitic drugs. Curr Pharm Des. 2007, 13: 641-651. 10.2174/138161207780162854.
Bhopale GM: Development of a vaccine for toxoplasmosis: current status. Microbes Infect. 2003, 5: 457-462. 10.1016/S1286-4579(03)00048-0.
Blake DP, Shirley MW, Smith AL: Genetic identification of antigens protective against coccidia. Parasite Immunol. 2006, 28: 305-314. 10.1111/j.1365-3024.2006.00831.x.
Shirley MW, Blake D, White SE, Sheriff R, Smith AL: Integrating genetics and genomics to identify new leads for the control of Eimeria spp. Parasitology. 2004, 128 (Suppl 1): S33-42.
Shirley MW, Smith AL, Tomley FM: The biology of avian Eimeria with an emphasis on their control by vaccination. Adv Parasitol. 2005, 60: 285-330. 10.1016/S0065-308X(05)60005-X.
Lillehoj HS, Lillehoj EP: Avian coccidiosis. A review of acquired intestinal immunity and vaccination strategies. Avian Dis. 2000, 44: 408-425. 10.2307/1592556.
Saini SS, Sodhi SS, Maiti NK, Sharma SN: Immune response of chicks to oral vaccination against Newcastle disease and fowl pox. Comp Immunol Microbiol Infect Dis. 1990, 13: 1-6. 10.1016/0147-9571(90)90002-B.
Schuijffel DF, van Empel PC, Pennings AM, van Putten JP, Nuijten PJ: Passive immunization of immune-suppressed animals: chicken antibodies protect against Ornithobacterium rhinotracheale infection. Vaccine. 2005, 23: 3404-3411. 10.1016/j.vaccine.2005.01.095.
Corrier DE, DeLoach JR: Evaluation of cell-mediated, cutaneous basophil hypersensitivity in young chickens by an interdigital skin test. Poult Sci. 1990, 69: 403-408.
Lessard M, Hutchings D, Cave NA: Cell-mediated and humoral immune responses in broiler chickens maintained on diets containing different levels of vitamin A. Poult Sci. 1997, 76: 1368-1378.
Dalloul RA, Lillehoj HS, Shellem TA, Doerr JA: Effect of vitamin A deficiency on host intestinal immune response to Eimeria acervulina in broiler chickens. Poult Sci. 2002, 81: 1509-1515.
Peek HW, Landman WJ: Higher incidence of Eimeria spp. field isolates sensitive for diclazuril and monensin associated with the use of live coccidiosis vaccination with paracox-5 in broiler farms. Avian Dis. 2006, 50: 434-439. 10.1637/7486-121205R.1.
Dzierszinski FS, Hunter CA: Advances in the use of genetically engineered parasites to study immunity to Toxoplasma gondii. Parasite Immunol. 2008, 30: 235-244. 10.1111/j.1365-3024.2007.01016.x.
Stanley VG, Gray C, Daley M, Krueger WF, Sefton AE: An alternative to antibiotic-based drugs in feed for enhancing performance of broilers grown on Eimeria spp.-infected litter. Poult Sci. 2004, 83: 39-44.
Nollet L, Huyghebaert G, Spring P: Effect of dietary mannan oligosaccharide (Bio-Mos) on live performance of broiler chickens given an anticoccidial vaccine (Paracox) followed by a mild coccidial challenge. J Appl Poult Res. 2007, 16: 397-403.
Franklin ST, Newman MC, Newman KE, Meek KI: Immune parameters of dry cows fed mannan oligosaccharide and subsequent transfer of immunity to calves. J Dairy Sci. 2005, 88: 766-775.
Nochta I, Tuboly T, Halas V, Babinszky L: Effect of different levels of mannan-oligosaccharide supplementation on some immunological variables in weaned piglets. J Anim Physiol Anim Nutr (Berl). 2008,
Davis ME, Maxwell CV, Erf GF, Brown DC, Wistuba TJ: Dietary supplementation with phosphorylated mannans improves growth response and modulates immune function of weanling pigs. J Anim Sci. 2004, 82: 1882-1891.
Middelbos IS, Godoy MR, Fastinger ND, Fahey GC: A dose-response evaluation of spray-dried yeast cell wall supplementation of diets fed to adult dogs: effects on nutrient digestibility, immune indices, and fecal microbial populations. J Anim Sci. 2007, 85: 3022-3032. 10.2527/jas.2007-0079.
Grieshop CM, Flickinger EA, Bruce KJ, Patil AR, Czarnecki-Maulden GL, Fahey GC: Gastrointestinal and immunological responses of senior dogs to chicory and mannan-oligosaccharides. Arch Anim Nutr. 2004, 58: 483-493. 10.1080/00039420400019977.
Lim BO, Yamada K, Nonaka M, Kuramoto Y, Hung P, Sugano M: Dietary fibers modulate indices of intestinal immune function in rats. J Nutr. 1997, 127: 663-667.
Shashidhara RG, Devegowda G: Effect of dietary mannan oligosaccharide on broiler breeder production traits and immunity. Poult Sci. 2003, 82: 1319-1325.
Zou XT, Qiao XJ, Xu ZR: Effect of beta-mannanase (Hemicell) on growth performance and immunity of broilers. Poult Sci. 2006, 85: 2176-2179.
Staykov Y, Spring P, Denev S, Sweetman J: Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss). Aquacul Int. 2007, 15: 153-161. 10.1007/s10499-007-9096-z.
Moyad MA: Brewer's/baker's yeast (Saccharomyces cerevisiae) and preventive medicine: Part II. Urol Nurs. 2008, 28: 73-75.
Elmusharafa MA, Peekb HW, Nolletc L, Beynena AC: The effect of an in-feed mannanoligosaccharide preparation (MOS) on a coccidiosis infection in broilers. Anim Feed Sci Tech. 2007, 134: 347-354. 10.1016/j.anifeedsci.2006.11.022.
Yun CH, Lillehoj HS, Lillehoj EP: Intestinal immune responses to coccidiosis. Dev Comp Immunol. 2000, 24: 303-324. 10.1016/S0145-305X(99)00080-4.
Girard F, Fort G, Yvore P, Quere P: Kinetics of specific immunoglobulin A, M and G production in the duodenal and caecal mucosa of chickens infected with Eimeria acervulina or Eimeria tenella. Int J Parasitol. 1997, 27: 803-809. 10.1016/S0020-7519(97)00044-1.
Jeurissen SH, Janse EM, Vermeulen AN, Vervelde L: Eimeria tenella infections in chickens: aspects of host-parasite: interaction. Vet Immunol Immunopathol. 1996, 54: 231-238. 10.1016/S0165-2427(96)05689-9.
Ovington KS, Alleva LM, Kerr EA: Cytokines and immunological control of Eimeria spp. Int J Parasitol. 1995, 25: 1331-1351. 10.1016/0020-7519(95)00069-E.
Kogan G, Pajtinka M, Babincova M, Miadokova E, Rauko P, Slamenova D, Korolenko TA: Yeast cell wall polysaccharides as antioxidants and antimutagens: can they fight cancer?. Neoplasma. 2008, 55: 387-393.
Kimura K, Goff JP, Schmerr MJ, Stabel JR, Inumaru S, Yokomizo Y: Activation of immune cells in bovine mammary gland secretions by zymosan-treated bovine serum. J Dairy Sci. 2008, 91: 1852-1864. 10.3168/jds.2007-0895.
Sauerwein H, Schmitz S, Hiss S: Effects of a dietary application of a yeast cell wall extract on innate and acquired immunity, on oxidative status and growth performance in weanling piglets and on the ileal epithelium in fattened pigs. J Anim Physiol Anim Nutr (Berl). 2007, 91: 369-380. 10.1111/j.1439-0396.2006.00663.x.
Hunter KW, Gault RA, Berner MD: Preparation of microparticulate beta-glucan from Saccharomyces cerevisiae for use in immune potentiation. Lett Appl Microbiol. 2002, 35: 267-271. 10.1046/j.1472-765X.2002.01201.x.
Sundaresan NR, Ahmed KA, Saxena VK, Sastry KV, Saxena M, Pramod AB, Nath M, Singh KB, Rasool TJ, DevRoy AK, Singh RV: Differential expression of inducible nitric oxide synthase and cytokine mRNA in chicken lines divergent for cutaneous hypersensitivity response. Vet Immunol Immunopathol. 2005, 108: 373-385. 10.1016/j.vetimm.2005.06.011.
Ahmed KA, Saxena VK, Ara A, Singh KB, Sundaresan NR, Saxena M, Rasool TJ: Immune response to Newcastle disease virus in chicken lines divergently selected for cutaneous hypersensitivity. Int J Immunogenet. 2007, 34: 445-455. 10.1111/j.1744-313X.2007.00722.x.
Cox E, Verdonck F, Vanrompay D, Goddeeris B: Adjuvants modulating mucosal immune responses or directing systemic responses towards the mucosa. Vet Res. 2006, 37: 511-539. 10.1051/vetres:2006014.
Acknowledgements
The authors express their especial gratitude to Dr Miguel Angel Baladez for his help and comments on this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
GGV, ACC, CLP, EAG, and GMN designed the study. GGV and ACC performed the experiments. GGV, EAG, and GMN analyzed the data and prepared the manuscript. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Gómez-Verduzco, G., Cortes-Cuevas, A., López-Coello, C. et al. Dietary supplementation of mannan-oligosaccharide enhances neonatal immune responses in chickens during natural exposure to Eimeria spp. Acta Vet Scand 51, 11 (2009). https://doi.org/10.1186/1751-0147-51-11
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1751-0147-51-11