
Abstract
Surfactant Protein-A (SP-A) is the most prominent among four proteins in the pulmonary surfactant-system. SP-A is expressed by alveolar epithelial cells type II as well as by a portion of non small cell lung carcinomas (NSCLC).
The expression of SP-A is complexly regulated on the transcriptional and the chromosomal level. SP-A is a major player in the pulmonary cytokine-network and moreover has been described to act in the pulmonary host defense.
By the use of cell culture or animal models the functional properties have been repeatedly shown in many aspects, often bearing surprising properties which strongly indicate the physiological importance of SP-A. To date SP-A is recognized as a molecule essential for pulmonary development, structure and function. An upcoming number of reports deals with the role of SP-A for pulmonary pathology. This article gives an overview about the state of knowledge on SP-A focused in applications for human pulmonary disorders and points out the importance for pathology-orientated research approaches using immunohistochemistry or in situ hybridization as promising methods to further elucidate the role of this molecule in adult lung diseases.
Similar content being viewed by others
Review
Surfactant
The role of the surfactant system for the development of the human lung is known to be essential. Since it is synthesized by humans starting in the 28th week of pregnancy and reaching functional levels in the 34th week, surfactant-substitution-therapy is a fundamental part of the treatment of premature babies suffering from Infant Respiratory Distress Syndrome (IRDS)[1].
Pulmonary surfactant regulates dynamically the alveolar surface tension. The central role of the surfactant system for maintaining pulmonary function has been repeatedly shown by the use of cell culture or animal models [2].
Surfactant is a complex mixture of lipids, carbohydrates and four proteins (SP-A, SP-B, SP-C, SP-D). The initial descriptions of surfactant lead back to the 1950s, but little attention was given to the surfactant proteins until the 1980s [3]. The genes coding for these proteins are located on different chromosomes. SP-B and SP-C are similarly structured hydrophobic proteins participating in the adsorption of phospholipids at the alveolar border, which leads to rapid reduction of the surface tension. The hydrophilic proteins SP-A and SP-D are members of the collectins with C-type lectin domains. SP-D together with SP-A play a role in the pulmonary defense against Gram-negative bacteria [4].
SP-A: biochemical properties and genetic organization
SP-A is the major surfactant apoprotein exhibiting complex interactions and participation in processes fundamental for pulmonary structure and function with its expression restricted to alveolar epithelial cells type II. Moreover expression of SP-A was also described for a portion of NSCLC facilitating a diagnostic marker for these carcinomas [5, 6]. After characterization of the biochemical properties, a complex chromosomal organization of the genes coding for SP-A has been demonstrated [3]. The locus of the SP-A on the one hand consists of two functionally active genes and a pseudogene [7, 8]. The two active genes SP-A1 and SP-A2 on the other hand display several different alleles and splicing variants, moreover different oligomeric states have been described [3, 9]. During the development of the lung these two genes are regulated differentially, a process triggered by cAMP and glucocorticoids [10]. The SP-A1 and SP-A2 genes display a homology of 94% in their nucleotide sequences and even 96% homology in the amino acid sequences [11]. Fig. 1, as one example, shows the transcriptional activity of the SP-A1 and SP-A2 genes determined by RT-PCR in homogenates of biopsies from NSCLC and corresponding tumor-free samples. The importance of the differential transcription of SP-A1 and SP-A2 for maintaining pulmonary function has repeatedly been demonstrated [12]. Phylogenetic analyses revealed that an ancestor proto-SP-A gene was diverged into SP-A1 and a second gene which subsequently emerged to SP-A2 and the SP-A pseudogene [3]. The high level of homology between SP-A1 and SP-A2 up to date prevents a differential analysis of the two gene products in situ. The expression of SP-A is also complexly regulated on the transcriptional level. Moreover the protein-turnover and the release of SP-A into the serum represents a further point of regulation [12]. This sophisticated regulation of the genetic activity is recognized as a further hint for the functional importance of SP-A.

RT-PCR: Transcription of SP-A1 and SP-A2 in NSCLC tumors (T) and corresponding tumor-free tissues (TF) from the same cases in comparison to GAPDH.
SP-A in pulmonary diseases
In recent years the role of defects in the expression of SP-A in context with different pulmonary diseases has become an issue of scientific investigations. Initially numerous studies have been performed to elucidate the role of surfactant substitution in pediatrics [2].
As one major function SP-A displays a protective role of the molecule in pulmonary host defense by interacting with various infectious agents such as bacteria, fungi and viruses. SP-A deficient knock-out mice – compared to wild type animals – are susceptible to infections with Pseudomonas aeruginosa [13] and the clearance of group B streptococcus is slower [14]. In accordance the defense of SP-A deficient mice against Respiratory Syncytial Virus (RSV) has been shown to be reduced and may be restored by exogenous SP-A administration [15–17].
By mediating the attachment of Mycobacterium tuberculosis to alveolar macrophages and promoting the phagocytosis of these bacteria, SP-A is important in the pathogenesis of tuberculosis [18–21]. SP-A also functionally interacts with staphylococci [22, 23], Haemophilus influenza Type A [24, 25], Pneumocystis carinii [26, 27], Influenza A Virus (less efficient than SP-D) [28], Candida tropicalis [29] and Aspergillus fumigatus [30].
Moreover, SP-A is involved in the complex pulmonary network of cytokines as a central player, for example interacting with TNF-alpha and several interleukins [31, 12, 14].
Therefore it is likely that defects in the expression of SP-A may be important in the course of non infectious pulmonary diseases of adult patients. In the case of idiopathic pulmonary fibrosis, for example, low levels of SP-A (measured by ELISA) have been reported in broncho alveolar lavages (BAL), but elevated levels were found in the sera [32–34]. Immunohistochemical examinations of the expression of SP-A in pulmonary fibrosis demonstrated evident defects by using specimens from different diseases displaying fibrotic changes in the lungs. In good agreement with the results in BAL reduced levels of SP-A have been observed in fibrotic lungs. This reduced SP-A-expression in fibrotic lungs may be caused by two reasons: a limited number of the SP-A producing type II pneumocytes and by a clearly reduced SP-A expression of the remaining cells [35].
Reduced levels of SP-A have been demonstrated also in other pulmonary diseases such as the Adult Respiratory Distress Syndrome (ARDS) and in pneumonia [36].
Keeping in mind that surfactant substitutes are widely available due to their application in pediatrics, a growing number of therapeutic possibilities may result from these findings.
In sarcoidosis elevated levels of SP-A have been described [37] using BAL; the same applies for the sera from patients with alveolar proteinosis [38].
Since SP-A represents a central molecule in pulmonary immunoregulation as well as in host-defense it is obvious that defects in the surfactant system may have functional influence in the course of these pulmonary disorders.
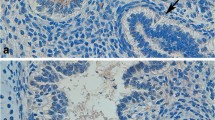
Another point of research concerning SP-A is the diagnostic value of this molecule, the expression of which is restricted to the lungs. It has been reported that SP-A levels in BAL or serum from patients suffering from idiopathic pulmonary fibrosis correlate with the progression of the disease and can be used to predict survival [34, 38]. In samples from airway secretions SP-A measurements are described to be useful also for the diagnostics of pulmonary edema where elevated levels have been found compared to healthy volunteers and ARDS patients [39]. By utilizing highly sensitive RT-PCR techniques the amplification of SP-A transcripts can be used for the detection of occult metastases in non small cell lung cancer patients [5, 40]. Comparative studies of different malignomas with pulmonary localization have shown the diagnostic properties of immunohistochemically determined SP-A [6, 41, 42]. In carcinomas of occult origin localized in the lungs the diagnosis has a crucial influence on the therapy. A positive detection of SP-A represents a clear hint for a primary location in the lung [43]. Fig. 2 as an example shows the immunohistochemical detection of SP-A in a moderately differentiated adenocarcinoma of the lung using the primary monoclonal antibody PE-10 and LSAB (AEC-substrate, × 100). The positive staining in the tumor cells (reddish) in this certain case helped to manifest the diagnosis as a primary carcinoma of the lung.

Immunohistochemical detection of SP-A using the monoclonal antibody PE-10 (LSAB, amonoethylcarbazole, 400×).
However, the choice of a suitable SP-A antibody is highly important since approaches using polyclonals display cross reactions with other tumors [44]. This procedure has already become a part of pathological routine diagnosis, and – along with other markers such as the Thyroid-transcription-factor-1 (TTF-1) – the detection of SP-A (by PE-10) is a useful part of the immunohistochemical panel in pulmonary pathology.
Immunohistochemical detection of SP-A even might be utilized for forensic purposes helping to distinguish between fatal drowning and postmortem immersion [45].
Conclusion
It is evident that SP-A is a molecule which already proves to be an interesting subject for medical research. However, the studies concerning the possible role of surfactant-defects in pulmonary diseases of adults have been performed mainly in different cell culture or animal models hardly analyzing adult human lung tissue. For these reasons SP-A is a promising target for histomorphological approaches using pathological specimens which exactly represent the scenarios of various diseases with all the different cell types involved which are difficult to simulate in models. With the modern tools of molecular pathology, the genetic activities of genes can be analyzed in situ, which provides evidence of the cellular activities in the context of a human native tissue. One example is shown in Fig. 3: a lung section hybridized with a digoxigenin-labeled SP-A probe to analyze the transcriptional activity in situ; the reddish signals of the transcripts are visible in the cytoplasm of type II pneumocytes. When analyzing the expression of SP-A in histological sections in context with other molecules of the pulmonary cytokine network one can expect further clues for the scenarios taking place in the course of interstitial lung diseases.

In situ hybridization targeting using a 663 bp digoxigenated DNA-probe complementary to SP-A mRNA. Detection was achieved by Anti-digoxigenin antibody conjugated to alkaline phosphatase with NBT/BCIP as a chromogen (400×).
Taken together, SP-A is a complexly regulated molecule with surprising properties and essential importance for pulmonary development, structure and function which is getting more and more into focus concerning various diseases of the adult lung. Thus, as an outlook, it will become an issue of pulmonary pathology which might provide promising perspectives for applications in research, diagnosis and therapy.
References
Gibson AT: Surfactant and the neonatal lung. Br J Hosp Med. 1997, 58: 381-384.
Kattwinkel J: 2Surfactant. Evolving issues. Clin Pedinatol. 1998, 25 (1): 17-32.
Floros J, Karinch AM: Human SP-A: then and now. Am J Physiol. 1995, 268: L162-L165.
Hoppe HJ, Reid KBM: Collectins – soluble proteins containing collagenous regions and lectin domains – and their roles in innate immunity. Prot Sci. 1994, 3: 1143-1158. 10.1002/pro.5560030801.
Betz C, Papadopoulos Th, Buchwald J, Dämmrich J, Müller-Hermelink HK: Surfactant protein gene expresssion in metastatic and micrometastatic pulmonary adenocarcinomas and other non-small cell lung carcinomas: detection by reverse transcriptase-polymerase chain reaction. Cancer Res. 1995, 55: 4283-4286.
Dempo K, Satoh M, Tsuji S, Mori M, Kuroki Y, Akino T: Immunohistochemical studies on the expression of pulmonary surfactant apoproteins in human lung carcinomas using monoclonal antibodies. Pathol Res Pract. 1987, 182: 669-675.
White RT, Damm D, Miller J, Spratt K, Schilling J, Hawgood S, Benson B, Cordell B: Isolation and characterization of the human pulmonary surfactant apoprotein gene. Nature. 1985, 317: 361-363. 10.1038/317361a0.
Korfhagen TR, Glasser SW, Bruno MD, McMahan MJ, Whitsett JA: A portion of the human surfactant protein A (SP-A) gene locus consists of a pseudogene. Am J Respir Cell Mol Biol. 1991, 4: 463-469.
Hickling TP, Malhorta R, Sim RB: Human lung surfactant protein A exsits in several different oligomeric states: oligomer size distribution varies between patient groups. Molecular Medicine. 1998, 4: 266-275.
McCormick SM, Mendelson CR: Human SP-A1 and SP-A2 genes are differentially regulated during development and by cAMP and glucocorticoids. Am J Physiol. 1994, 266: L367-L374. (Lung Cell Mol Physiol 10)
McCormick SM, Boggaram V, Mendelson CR: Characterization of mRNA transcripts and organization of human SP-A1 and SP-A2 genes. Am J Physiol. 1994, 266: L354-366. (Lung Cell Mol Physiol 10)
Korfhagen TR, Glasser SW, Stripp BR: Regulation of gene expression in the lung. Curr Opin Pediatr. 1994, 6: 255-261. 10.1097/00008480-199406000-00005.
LeVine AM, Kurak KE, Bruno MD, Stark JM, Whitsett JA, Korfhagen TR: Surfactant protein-A-deficient mice are susceptible to Pseudomonas aeruginosa infection. Am J Respir Cell Mol Biol. 1998, 19: 700-708.
LeVine AM, Kurak KE, Wright JR, Watford WT, Bruno MD, Ross GF, Whitsett JA, Korfhagen TR: Surfactant protein-A binds group B streptococcus enhancing phagocytosis and clearance from lungs of surfactant protein-A-deficient mice. Am J Respir Cell Mol Biol. 1999, 20: 279-286.
Kerr MH, Paton JY: Surfactant protein levels in severe respiratory syncytial virus infection. Am J Respir Crit Care Med. 1999, 159: 1115-1118.
LeVine AM, Gwozdz J, Stark J, Bruno M, Whitsett J, Korfhagen T: Surfactant protein-A enhances respiratory syncytial virus clearance in vivo. J Clin Invest. 1999, 103: 1015-1021. 10.1172/JCI5849.
Van Schaik SM, Vargas I, Welliver RC, Enhorning G: Surfactant dysfunction develops BAB/c mice infected with respiratory syncytial virus. Pediatr Res. 1997, 42: 169-173. 10.1203/00006450-199708000-00007.
Downing JF, Pasula R, Wright JR, Twigg HL, Martin WJ: Surfactant protein a promotes attachment of Mycobacterium tuberculosis to alveolar macrophages during infection with human immunodeficiency virus. Proc Natl Acad Sci USA. 1995, 92: 4848-4852. 10.1073/pnas.92.11.4848.
Gaynor CD, McCormack FX, Voelker DR, McGowan SE, Schlesinger LS: Pulmonary surfactant protein A mediates enhanced phagocytosis of Mycobacterium tuberculosis by a direct interaction with human macrophages. J Immunol. 1995, 155: 5343-5351.
Martin WJ, Downing JF, Williams MD, Pasula R, Twigg HL, Wright JR: Role of surfactant protein A in the pathogenesis of tuberculosis in subjects with human immunodeficiency virus infection. Proc Assoc Am Physicians. 1995, 107: 340-345.
Pasula R, Downing JF, Wright JR, Kachel DL, Davis TE, Martin WJ: Surfactant protein A (SP-A) mediates attachment of Mycobacterium tuberculosis to murine alveolar macrophages. Am J Respir Cell Mol Biol. 1997, 17: 209-217.
Manz-Keinke H, Plattner H, Schlepper-Schafer J: Lung surfactant protein A (SP-A) enhances serum-independent phagocytosis of bacteria by alveolar macrophages. Eur J Cell Biol. 1992, 57: 95-100.
Van Iwaarden F, Welmers B, Verhoef J, Haagsman HP, van Golde LM: Pulmonary surfactant protein A enhances the host-defense mechanism of rat alveolar macrophages. Am J Respir Cell Mol Biol. 1990, 2: 91-98.
McNeely TB, Coonrod JD: Aggregation and opsonization of type A but not type B Hemophilus influenzae by surfactant protein A. Am J Respir Cell Mol Biol. 1994, 11: 114-122.
Tino MJ, Wright JR: Surfactant protein A stimulates phagocytosis of specific pulmonary pathogens by alveolar macrophages. Am J Physiol. 1996, 270: L677-L688.
Koziel H, Phelps DS, Fishman JA, Armstrong MY, Richards FF, Rose RM: Surfactant protein-A reduces binding and phagocytosis of pneumocystis carinii by human alveolar macrophages in vitro. Am J Respir Cell Mol Biol. 1998, 18: 834-843.
Zimmerman PE, Voelker DR, McCormack FX, Paulsrud JR, Martin WJ: 120-kD surface glycoprotein of Pneumocystis carinii is a ligand for surfactant protein A. J Clin Invest. 1992, 89: 143-9. 10.1172/JCI115554.
Hartshorn KL, White MR, Shepherd V, Reid K, Jensenius JC, Crouch EC: Mechanisms of anti-influenza activity of surfactant proteins A and D: comparison with serum collectins. Am J Physiol. 1997, 273: L1156-1166.
Pison U, Max M, Neuendank A, Weissbach S, Pietschmann S: Host defence capacities of pulmonary surfactant: evidence for 'non-surfactant' functions of the surfactant system. Eur J Clin Invest. 1994, 24: 586-99. 10.1111/j.1365-2362.1994.tb01110.x.
Madan T, Eggleton P, Kishore U, Strong P, Aggrawal SS: Binding of pulmonary surfactant proteins A and D to Aspergillus fumigatus conidia enhances phagocytosis and killing by human neutrophils and alveolar macrophages. Infect Immun. 1997, 65: 3171-3179.
Arias-Diaz J, Vara E, Garcia C, Gomez M, Balibrea JL: Tumor necrosis factor-alpha inhibits synthesis of surfactant by isolated human type II pneumocytes. Eur J Surg. 1993, 159: 541-549.
Honda Y, Kuroki Y, Shijubo N, Fujishima T, Takahashi H, Hosoda K, Akino T, Abe S: Aberrant appearance of lung surfactant protein A in sera of patients with idiopathic pulmonary fibrosis and its clinical significance. Respiration. 1995, 62 (2): 64-69. 10.1159/000196393.
Kuroki Y, Tsutahara S, Shijubo N, Takahashi H, Shiratori M, Hattori A, Honda Y, Abe S, Akino T: Elevated levels of lung surfactant protein A in sera from patients with idiopathic pulmonary fibrosis and pulmonary alveolar proteinosis. Am Rev Respir Dis. 1993, 147: 723-729.
McCormack FX, King TE, Bucher BL, Nielsen L, Mason RJ: Surfactant protein A predicts survival in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 1995, 152 (2): 751-759.
Deutschbein ME, Goldmann T, Galle J, Vollmer E: Patterns of Surfactant Aporotein-A (SP-A) Expression in Pulmonary Fibrosis. E J Pathol. 1999, 5: 99-101.
Gunther A, Siebert C, Schmidt R, Ziegler S, Grimminger F, Yabut M, Temmesfeld B, Walmrath D, Morr H, Seeger W: Surfactant alterations in severe pneumonia, acute respiratory distress syndrome, and cardiogenic lung edema. Am J Respir Crit Care Med. 1996, 153: 176-184.
Hamm H, Luhrs J, Guzman y Rotaeche J, Costabel U, Fabel H, Bartsch W: Elevated surfactant protein A in bronchoalveolar lavage fluids from sarcoidosis and hypersensitivity pneumonitis patients. Chest. 1994, 106 (6): 1766-1770. 10.1378/chest.106.6.1766.
Kinder BW, Brown KK, McCormack FX, Ix JH, Kervitsky A, Schwarz MI, King TE: Serum Surfactant Protein-A Is a Strong Predictor of Early Mortality in Idiopathic Pulmonary Fibrosis. Chest. 2009,
Shimura S, Masuda T, Takishima T, Shirato K: Surfactant Protein-A concentration in airway secretions for the detection of pulmonary oedema. Eur Respir J. 1996, 9: 2525-2530. 10.1183/09031936.96.09122525.
Papadopoulos T: Detection of surfactant protein gene expression by reverse transcriptase-polymerase chain reaction (RT-PCR) identifies metastases and occult tumor spread of pulmonary adenocarcinomas. Verh Dtsch Ges Pathol. 1996, 80: LVII-LXI.
Sugiyama K, Kawai T, Nakanishi K, Suzuki M: Histochemical reactivities of lectins and surfactant apoprotein in pulmonary adenocarcinomas and their metastases. Mod Pathol. 1992, 5: 273-276.
Nicholson AG, McCormick CJ, Shimosato Y, Butcher DN, Sheppard MN: The value of PE-10, a monoclonal antibody against pulmonary surfactant, in distinguishing primary and metastatic lung tumors. Histopathol. 1995, 25: 57-60. 10.1111/j.1365-2559.1995.tb00291.x.
Goldmann T, Vollmer E: Immunohistochemical detection of SP-A is of diagnostik support in a patient suffering froma metastatic adenocarcinoma of the lungs. E J Pathol. 1999, 5: 993-998.
Bejarano PA, Baughman RP, Biddinger PW, Miller MA, Fenoglio-Preiser C, Al-Kafaji B, Di Lauro R, Whitsett JA: Surfactant Proteins and Thyroid Transcription Factor-1 in Pulmonary and Breast Carcinomas. Mod Pathol. 1996, 9: 445-452.
Stemberga V, Stifter S, Cuculiæ D, Coklo M, Bosnar A: Immunohistochemical surfactant protein-A expression: Fatal drowning vs. postmortem immersion. Med Hypotheses. 2009, 72: 413-415. 10.1016/j.mehy.2008.11.024. Epub 2009
Acknowledgements
The authors thank J. Tiebach and M. Lammers for excellent technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
TG, DK and EV conceived the study. HS, MA, DSL and FS took part in the conception and writing of the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Goldmann, T., Kähler, D., Schultz, H. et al. On the significance of Surfactant Protein-A within the human lungs. Diagn Pathol 4, 8 (2009). https://doi.org/10.1186/1746-1596-4-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1746-1596-4-8