
Abstract
Background
Surface charge and oxidative stress are often hypothesized to be important factors in cytotoxicity of nanoparticles. However, the role of these factors is not well understood. Hence, the aim of this study was to systematically investigate the role of surface charge, oxidative stress and possible involvement of mitochondria in the production of intracellular reactive oxygen species (ROS) upon exposure of rat macrophage NR8383 cells to silicon nanoparticles. For this aim highly monodisperse (size 1.6 ± 0.2 nm) and well-characterized Si core nanoparticles (Si NP) were used with a surface charge that depends on the specific covalently bound organic monolayers: positively charged Si NP-NH2, neutral Si NP-N3 and negatively charged Si NP-COOH.
Results
Positively charged Si NP-NH2 proved to be more cytotoxic in terms of reducing mitochondrial metabolic activity and effects on phagocytosis than neutral Si NP-N3, while negatively charged Si NP-COOH showed very little or no cytotoxicity. Si NP-NH2 produced the highest level of intracellular ROS, followed by Si NP-N3 and Si NP-COOH; the latter did not induce any intracellular ROS production. A similar trend in ROS production was observed in incubations with an isolated mitochondrial fraction from rat liver tissue in the presence of Si NP. Finally, vitamin E and vitamin C induced protection against the cytotoxicity of the Si NP-NH2 and Si NP-N3, corroborating the role of oxidative stress in the mechanism underlying the cytotoxicity of these Si NP.
Conclusion
Surface charge of Si-core nanoparticles plays an important role in determining their cytotoxicity. Production of intracellular ROS, with probable involvement of mitochondria, is an important mechanism for this cytotoxicity.
Similar content being viewed by others


Introduction
Silicon (Si) is conventionally regarded as a non-toxic semiconductor material and Si NP are proposed as an alternative for the highly toxic heavy metal quantum dots in biological applications such as in food industry and bioimaging [1]. However, once exposed to an aerobic atmosphere, Si NP readily get oxidized to silica (silicon dioxide; SiO2) [2], which is reported to result in cytotoxicity [3]. The cytotoxicity of silica nanoparticles has been reported to be size dependent [4]. Data on the actual toxicity of silicon NP (Si NP) are, however, scarce.
Recently, we developed a method for the gram-scale synthesis of Si NP [5], which can be coated with a covalently bound organic monolayer with different surface charges [6–8]. These nanoparticles have a silicon core of 1.6 ± 0.2 nm as determined by TEM [7, 8]. By attaching alkyl chains to the surface of the Si core with amine (NH2), azide (N3) and carboxylic acid (COOH) terminal moieties, Si NP with respectively positive (Si NP-NH2), neutral (Si NP-N3) and negative (Si NP-COOH) surface charges can be obtained. This coating prevents the oxidation of Si NP to SiO2. Also the influence of surface charges on the cytotoxicity remains largely unresolved, although there are several research articles pointing at a possible role of surface charge in cellular uptake and/or cytotoxicity of nanoparticles. Oskuee et al.[9], for example, reported a decrease in cytotoxicity with decreasing positive surface charge of polyethyleneimine NP-s. Sayin et al.[10] found that positively charged N-trimethyl chitosan NP-s were more cytotoxic than their negatively charged counterparts.
A brief overview of some recent articles pointing at the possible influence of surface charge of nanoparticles on their cellular uptake and/or cytotoxicity is given in Table 1, which summarizes the findings reported by different groups. The findings are based on different types of particles functionalized with different chemical groups. A consensus regarding the role of surface charge on cytotoxicity of nanoparticles is therefore hard to reach. Some research groups [11, 12] observed cytotoxic effects of positively charged nanoparticles. Mayer et al.[13] reported activation of the complement system and increased hemolysis in blood samples collected from healthy donors after being exposed to positively charged polystyrene nanoparticles. Some recent publications [14–20] reported different effects of surface charges on cytotoxicity, including a higher cytotoxicity of cationic nanoparticles as compared to anionic nanoparticles. Gupta et al.[21] recently observed a reduced cytotoxicity for nanoparticles with a positive surface charge. On the other hand, other research groups [22, 23] failed to observe any significant effect of surface charge of nanoparticles on their cytotoxicity.
Previously, we reported the synthesis and cytotoxic effects of differently charged Si NP towards human colonic adenocarcinoma derived Caco-2 cells [6]. Positively charged Si NP-NH2 exerted the highest toxicity, whereas the negatively charged Si NP-COOH were hardly toxic and the neutral Si NP-N3 showed intermediate toxicity towards the Caco-2 cells. Unfortunately, investigations regarding the mechanisms of cytotoxicity of nanoparticles are quite limited. Also, different research groups use different experimental models. Nevertheless, oxidative stress is proposed as one of the most important mechanisms for NP mediated toxicity [24–33]. In these studies a wide range of nanoparticles was tested in different cellular models. Still, a concerted study on the effect of surface charge on the production of intracellular ROS and subsequent oxidative stress and cytotoxicity is not available. Therefore, in the present study we systematically investigated the role of NP surface charge and oxidative stress in the rat alveolar macrophage NR8383 cell line. This cell line provides an adequate model system for studying the effect of NP on phagocytic cells, which can clarify possible effects on the innate immune response. In the current study the mechanism underlying the differential cytotoxicity of the various Si NP is investigated, with a focus on the possible role for the formation of intracellular ROS and oxidative stress. To this end the production of intracellular ROS in both Caco-2 and NR8383 cells exposed to increasing concentrations of the differently charged Si NP was investigated. Also the degree of protection provided by pre-incubation of the cells with the antioxidants vitamin E and vitamin C and the possible role of Si NP-induced mitochondrial activity in the production of the intracellular ROS was studied. The results indicate how the surface charge of the NP influences their capability to induce intracellular ROS formation and oxidative stress and point at a role of mitochondria in this NP-induced ROS production at the subcellular level.
Results
Figure 1 shows the cytotoxic effect of increasing concentrations of various Si NP on NR8383 cells as detected by the reduction in the mitochondrial metabolic activity (MTT assay) of these cells after 24 hours exposure. Si NP-NH2 showed the highest cytotoxicity (EC50 value = 12 ng/ml) followed by Si NP-N3 which were cytotoxic at relatively higher concentrations (EC50 value = 270 ng/ml). Si NP-COOH failed to show any cytotoxicity up to concentrations of 3000 ng/ml (Table 2).

Effect of 24 hours exposure of NR8383 cells to increasing concentrations of Si NP-NH 2 (black triangle), Si NP-N 3 (black square) and Si NP-COOH (black dot) on mitochondrial metabolic activity (measured by the MTT assay). Error bars show standard error of mean (n = 3). The asterisk (*) sign signifies P < 0.05.
The effects of increasing concentrations of the differently charged Si NP on the phagocytic index of NR8383 cells upon 24 hours of exposure are shown in figure 2. Exposure to Si NP-NH2 and Si NP-N3 resulted in a concentration-dependent decrease in the phagocytic index. The EC50 value for Si NP-NH2 was 60 ng/ml and for Si NP-N3 it was 320 ng/ml (Table 2). Exposure to Si NP-COOH did not result in a decrease in phagocytosis index, but surprisingly, even resulted in a steady increase in the phagocytosis index up to 130 % of the control values at a concentration of 3000 ng/ml.

Effect of 24 hours exposure of NR8383 cells to increasing concentrations of Si NP-NH 2 (black triangle), Si NP-N 3 (black square) and Si NP-COOH (black dot) on phagocytosis (measured as the phagocytic index). Error bars show standard error of mean (n = 3). The asterisk (*) sign signifies P < 0.05.
In figure 3A the intracellular ROS production (measured by DCFH-DA assay) in NR8383 cells after 24 hours exposure to various Si NP is shown. Exposure of NR8383 cells to Si NP-NH2 resulted in a dose-dependent increase in intracellular ROS production (EC50 value = 22 ng/ml). Upon exposure to Si NP-N3 also an increase in intracellular ROS production was observed (EC50 value = 190 ng/ml), although the rate of intracellular ROS production was lower than that observed upon exposure of the NR8383 cells to Si NP-NH2 (Table 2). Upon exposure of the NR8383 cells to Si NP-COOH no increase in intracellular ROS production was observed.

Induction of intracellular ROS production (measured using the DCFH-DA assay) in NR8383 (A) and Caco-2 (B) cells after 24 hours exposure to increasing concentrations of Si NP-NH 2 (black triangle), Si NP-N 3 (black square) and Si NP-COOH (black dot). Error bars show standard error of mean (n = 3). The asterisk (*) sign signifies P < 0.05.
In figure 3B the intracellular ROS production (measured by DCFH-DA assay) in Caco-2 cells after 24 hours exposure to various Si NP is shown. Upon exposure to Si NP-NH2 intracellular ROS production started at low concentrations and then grew rapidly reaching a plateau at concentrations above 46 ng/ml (EC50 value = 18 ng/ml; Table 2). For Caco-2 cells exposed to Si NP-N3, intracellular ROS production increased significantly at concentrations above 50 ng/ml and increased up to higher concentrations (EC50 value = 310 ng/ml; Table 2). For Caco-2 cells exposed to Si NP-COOH, no increase in intracellular ROS production was observed.
In figure 4 ROS production (measured by DCFH-DA assay) upon 90 minutes incubation of an isolated rat liver mitochondrial fraction with different Si NP is shown. Incubation with Si NP-NH2 resulted in a concentration-dependent increase in ROS production, which started at concentrations as low as 6 ng/ml. The calculated EC50 value for Si NP-NH2 induced mitochondrial ROS production was 80 ng/ml (Table 2). Incubation of the mitochondrial fraction with Si NP-N3 resulted in a concentration-dependent increase in ROS formation starting at 90 ng/ml, with an EC50 value of 1050 ng/ml (Table 2). No increase in ROS production was found for the Si NP-COOH even for concentrations as high as 3000 ng/ml.

Induction of ROS production (measured using the DCFH-DA assay) upon 90 minutes incubations of an isolated rat liver mitochondrial fraction with increasing concentrations of Si NP-NH 2 (black triangle), Si NP-N 3 (black square) and Si NP-COOH (black dot). Error bars show standard error of mean (n = 3). The asterisk (*) sign signifies P < 0.05.
In figure 5A the effects are depicted of the antioxidants vitamin E and C on the cytotoxicity in NR8383 cells upon 24 hours exposure to Si NP-NH2. Preincubation of the NR8383 cells with vitamin E resulted in a significant reduction in the Si NP-NH2-induced cytotoxicity with EC50 values of 12 ng/ml and 60 ng/ml (p < 0.05) for NR8383 cells not preincubated with antioxidants (control) and cells preincubated with 100 μM vitamin E, respectively (Table 2). Although a higher EC50 value of 32 ng/ml was found for NR8383 cells preincubated with 1 mM of vitamin C in comparison to NR8383 cells not preincubated with antioxidants (control) the magnitude of reduction in cytotoxicity for 1 mM vitamin C was less than preincubation of NR8383 cells with 100 μM vitamin E (EC50 value = 60 ng/ml; Table 2).

Protective effect of preincubation of NR8383 cells with vitamin E (open square) and vitamin C (open circle) against cytotoxicity (measured using the MTT assay) detected after 24 hours exposure to serial dilutions of (A) Si NP-NH 2 (black triangle) and (B) Si NP-N 3 (black square). Error bars show standard error of mean (n = 3). The asterisk (*) sign signifies P < 0.05 (compared to data without antioxidants).
The effects of vitamins E and C on the cytotoxic effects of Si NP-N3 in NR8383 cells are shown in figure 5B. For cells exposed to Si NP-N3, vitamin C offered a significant protection, whereas preincubation of the cells with vitamin E did not result in protection against the cytotoxicity of the NP. The corresponding EC50 values for Si NP-N3 were 270 ng/ml, 310 ng/ml and 510 ng/ml (p < 0.05), respectively, for NR8383 cells not preincubated with antioxidants (control), cells preincubated with 100 μM vitamin E, and cells preincubated with 1 mM vitamin C.
Discussion
The surface charge of nanoparticles plays an important role in their interaction with cells and cytotoxicity. In this paper the role of surface charge on cytotoxicity is studied and mechanistic factors (like oxidative stress) causing this cytotoxicity are probed. The main conclusions are that positively charged Si NP-NH2 are more cytotoxic towards NR8383 cells both in terms of reducing mitochondrial metabolic activity and phagocytosis than neutral Si NP-N3, while negatively charged Si NP-COOH showed very little or no cytotoxicity. Si NP-NH2 produced most intracellular ROS followed by Si NP-N3, with no ROS produced by Si NP-COOH. Part of this ROS production probably originates from mitochondria, as a similar trend in ROS production was observed in NR8383 cells and in incubations of isolated rat liver mitochondrial fractions with Si NP. The Si NP-NH2 and Si NP-COOH are slightly more hydrophilic than Si NP-N3. However, since the difference in cytotoxicity and ROS production is much larger between Si NP-NH2 and Si NP-COOH than between Si NP-NH2 or Si NP-COOH and Si NP-N3 we conclude that surface charge is much more important than differences in hydrophilicity or hydrophobicity.
The role of ROS was also reflected in the protection against cytotoxicity by Si NP upon pretreatment of the NR8383 cells with vitamins E and C. Interestingly, pretreatment of the NR8383 cells with vitamin E showed more protection towards Si NP-NH2 than vitamin C, while vitamin C showed more protection towards Si NP-N3. The mechanism behind this difference may be related to the nature of the ROS formed and/or the location(s) and mechanism(s) of this ROS formation, although the results of the present study reveal that in both cases mitochondrial ROS production is likely involved. Pan et al.[34] previously reported different protective effects offered by different antioxidants (N-acetylcysteine, glutathione, triphenylphosphine monosulphonate and vitamin C) while studying the cytotoxic effects of gold nanoparticles on HeLa cells.
The relatively higher cytotoxicity of Si NP-NH2 as compared to Si NP-N3 or Si NP-COOH is in agreement with our previous study, where similar effects of these Si NP were observed on human colonic adenocarcinoma-derived Caco-2 cells [6]. Shiohara et al.[35] recently reported the synthesis and cytotoxicity of Si NP with different functionalized surfaces containing amine, epoxide, diene and diol terminated alkyl chains. These authors also reported a higher toxicity of amine-terminated Si NP as compared to the other Si NP. However, the functional groups of their Si NP were mostly different from our Si NP. Also, their Si NP were substantially larger (~4 nm in size), more polydisperse and prepared by a different synthetic technique. Goodman et al.[36] tested the cytotoxicity of cationic and anionic monolayer-coated gold nanoparticles on Cos-1 cells and found that cationic particles were toxic, whereas the anionic particles were not toxic at all, which matches our observation as well. Table 1 presents an overview of additional literature studies on nanoparticles, showing the role of surface charge on cellular interaction, including cellular uptake and cytotoxicity. It seems that the findings reported in these studies can be summarized to an overall match to our results since mostly positive charged nanoparticles are cytotoxic, although the cytological and experimental models used and endpoints studied in these reported studies are quite different from the ones we applied.
Our data also showed that positively charged Si NP-NH2 and the neutral Si NP-N3 decreased the phagocytic activity in NR8383 cells, but surprisingly reveal that the negatively charged Si NP-COOH increased the phagocytic index significantly. Although the precise mechanism of this phenomenon is not fully understood (see below), several indications have been reported in the literature that suggest that this is in fact not an unexpected phenomenon. One study reported comparable results for etoposide-incorporated tripalmitin nanoparticles with positive and negative surface charges [37]. That study showed that in rats, after 24 hours of intravenous administration, the concentration of positively charged particles in blood was 1.64 times that of negatively charged ones. Sengupta et al.[38] showed that etoposide-incorporated cationic liposomes attain a longer half-life in blood than anionic liposomes containing etoposide, due to a decreased phagocytic activity of macrophages for cationic particles that is not seen in the case of anionic particles. A similar charge-dependent increase (for negatively charged liposomes) or decrease (for positively charged liposomes) of phagocytosis of liposomes containing stearylamine was reported by Levchenko et al.[39]. Comparable results were reported by Aoki et al.[40] while investigating the disposition kinetics of liposomes with encapsulated methyl-2-amino-6-palmitoyl-D-glycoside. They found that the surface charge of liposomes is an important factor for phagocytosis of liposomes by the reticule-endothelial system: positively charged liposomes decreased phagocytosis, while negatively charged ones did not affect phagocytosis. It should be noted that all studies quoted above [37–40] were bioavailability studies and only potentially non-toxic nanoparticles were tested irrespective of their surface charge. Furthermore, in most of these studies no proper cytotoxicity tests were performed simultaneously with the bioavailability experiments.
As the Si NP-COOH proved to exert very little or no cytotoxicity at the tested concentrations, the induction of phagocytosis by negatively charged Si NP-COOH may be compared with similar findings reported in the above-mentioned studies. However, these findings should not be compared with our results on positively charged Si NP-NH2 which are cytotoxic within the tested concentration range. Hence, the decrease in phagocytic capability of NR8383 cells after exposure to positively charged Si NP-NH2 is the result of the positive surface charge and NP cytotoxicity but does not reflect an absence or induction in cellular uptake. A possible explanation for the induction of phagocytosis by negatively charged nanoparticles is given by Hernandez-Caselles et al.[41], who reported better binding of opsonin serum proteins with negatively charged nanoparticles than with positively charged nanoparticles. As a result, negatively charged nanoparticles get more covered with opsonin proteins, which stimulates phagocytosis by macrophages. It should be noted that the cell culture medium F12-K used in our experiments had fetal calf serum, which contained opsonin proteins (like immunoglobulins) in it as well.
In order to get more insight in the mechanism behind the cytotoxicity of Si NP towards NR8383 and Caco-2 cells we assessed the production of intracellular ROS. From our results it is evident that positively charged Si NP-NH2 were more efficient in producing intracellular ROS than the neutral Si NP-N3, which produced ROS only at much higher concentrations, and than negatively charged Si NP-COOH that did not produce any intracellular ROS at all. Xia et al.[42] did an extensive study on murine macrophage RAW 264.7 cells with a broad range of nanoparticles (including metal oxides like titanium dioxide, fullerols, carbon black and amine terminated polystyrene nanoparticles/PS-NH2) and found that only carbon black and cationic PS-NH2 were able to generate intracellular ROS production. ROS production by cationic PS-NH2 is in line with our study where Si NP-NH2 were also able to induce production of intracellular ROS. Stone et al.[43] also mentioned oxidative stress as an important mechanism for cytological injury caused by nanoparticles. Previously, Foucaud et al.[44] showed that carbon black ultrafine particles are capable of producing ROS in biologically relevant medium (like cell culture medium). Additionally, there are several other reports in which oxidative stress has been identified as an important mechanism for cytotoxicity of nanoparticles [45–47].
Subsequently, we investigated the capability of mitochondria to act as a source for nanoparticle-induced ROS production, since mitochondria have previously been recognized as a probable subcellular target of nanoparticles [48]. The results of our study reveal that mitochondria might indeed interact and contribute to the intracellular ROS production induced by NP. Karataş et al.[49] also reported that the mitochondrial membrane may be a target for NP interaction. They used an isolated mitochondrial fraction from A549 lung cancer cells incubated with negatively charged 13 nm gold nanoparticles, and studied the interaction with surface-enhanced Raman scattering (SERS). Using AFM and TEM they also demonstrated interaction of the gold nanoparticles with the mitochondrial membrane. There is one more report [34] in which 1.4 nm and 15 nm gold nanoparticles were tested on HeLa cells. The 1.4 nm gold nanoparticles were coated with triphenylphosphine sulphonate groups and their cytotoxic effect on the mitochondrial membrane potential (ΔΨ) was compared with bigger 15 nm gold nanoparticles. However, that study focused more on the size effect of gold nanoparticles on the mitochondrial membrane potential, and did not report the effect of surface charge of nanoparticles in interaction of the NP with the mitochondrial membrane. To the best of our knowledge, the present study is the first one to show production of ROS directly from mitochondria when exposed to NP. The observation that incubation of the mitochondrial fraction with positively charged Si NP-NH2 produced more ROS than incubation with neutral Si NP-N3 and negatively charged Si NP-COOH is in agreement with findings reported by Xia et al.[42]. With electron microscopy they saw that positively charged NH2-PS nanospheres completely disrupted the morphology of mitochondria after 16 hours of exposure, while negatively charged COOH-PS nanospheres did not cause any harm to the mitochondria. They also reported a dissipation in mitochondrial membrane potential (ΔΨ) with NH2-PS but not for COOH-PS nanospheres. These data suggest that the interaction of cationic NP with mitochondrial membranes exerts significant toxic effects.
Conclusion
The present study demonstrates the importance of surface charge in the cytotoxicity of nanoparticles. Dose-dependent and charge-dependent production of intracellular ROS further emphasizes the role of oxidative stress as an important mechanism for cytotoxicity. Mitochondria are likely target organelles for nanoparticles in the production of intracellular ROS. The negative Si NP-COOH show potential for further biological applications, whilst positive Si NP-NH2 should be applied with care.
Methods
Silicon nanoparticles
Si NP-NH2 and Si NP-N3 were prepared and characterized by methods described by Rosso-Vasic et al.[8] and Si NP-COOH were prepared by the method described by Ruizendaal et al.[6]. All three NP were of comparable sizes with a mean core diameter of 1.6 ± 0.2 nm as determined by TEM [7, 8] and all three Si NP are modified by about 25 surface groups [5]. Both Si NP-NH2 and Si NP-COOH were dissolved in ultrapure sterile water, while the stock solution of Si NP-N3 was dissolved in pure DMSO. The required final concentration ranges of all three Si NP were prepared by serial dilution with medium and amounted to 0.1 - 370 ng/ml for the Si NP-NH2; 0.1 - 2200 ng/ml for the Si NP-N3, and 0.1 - 3000 ng/ml for the Si NP-COOH which were the final tested concentration ranges for each of the assays described in this paper unless otherwise mentioned. No aggregation of these particles was observed with time in medium with DLS for at least 2 weeks and the solutions were vortexed at least for 1 minute before use. The solutions were filtered through 0.2 μm cellulose filters before cytotoxicity assays to maintain sterility. The amount of DMSO in the wells containing Si NP-N3 was calculated to be < 1% (v/v), at which concentration no cytotoxic effect of the DMSO was observed.
Cell Lines and tissue fractions
NR8383 cells
Rat alveolar macrophage cells were obtained from ATCC (Manassas, VA). The cells were cultured in 150 cm2 cell culture flasks with 25 ml F12-K culture medium (Gibco 21127) supplemented with 10% (v/v) heat-inactivated FCS in a humidified atmosphere containing 5% CO2 at 37°C.
Caco-2 cells
The human colonic adenocarcinoma cells were obtained from ATCC (Manassas, VA). The cell line was cultured in a humidified atmosphere containing 5% CO2 at 37°C in DMEM medium supplemented with 10% (v/v) heat-inactivated FCS, 1% (v/v) NEAA and 0.1% (v/v) gentamicin.
Isolation of the mitochondrial fraction from rat liver tissue
A Wistar rat of body weight 200 - 250 g was sacrificed by decapitation following anesthesia with isoflurane. The liver was excised at 4°C and homogenized in ice-cold saline using a Potter homogenizer with 20 - 25 strokes. The homogenized tissue was then centrifuged at 500 g for 10 minutes at 4°C. The supernatant was then removed and the pellet after being weighed was resuspended in ice-cold saline and centrifuged at 10.000 g for 10 minutes at 4°C. The supernatant was removed again and the pellet containing the isolated mitochondrial fraction was suspended in phosphate buffered saline (PBS) at 3 mg pellet/ml concentration with the addition of 0.4 mM glutamate and 0.4 mM NAD+ as substrates for mitochondrial oxidative phosphorylation. The mitochondrial fraction was then kept in an incubator at 37°C with 5% CO2 atmosphere for 4 hours to ensure start of mitochondrial respiration.
MTT assay
Cytotoxicity of Si NP
An NR8383 cell suspension was collected and centrifuged at 140 g for 5 minutes before resuspending the cell pellet in medium followed by counting and adjusting the cellular concentration to 2 × 105 cells/ml. The cells were then seeded in a 96-well plate (50 μl/well) and the plate was kept in a 5% CO2 incubator at 37°C for 24 hours. Next day 50 μl of serial dilutions of Si NP were added to the cells to obtain the required final concentrations of Si NP and then incubated for 24 hours. After 24 hours 5 μl of MTT solution in PBS (5 mg/ml) was added to each well and the plate was incubated for another 4 hours. Then 100 μl of pure DMSO was added to each well to dissolve the formazan crystals. Now the absorption of each well was measured at 562 nm in a 96-well plate reader and the background absorption at 612 nm [50] was subtracted. Mitochondrial metabolic activity for each concentration of Si NP was expressed as % of corresponding negative control reading. Medium without Si NP and medium with Triton-X (0.1%) were used as negative and positive controls respectively. Control experiments were done to exclude a possible reaction between MTT salt and Si NP. Additional control experiments were performed with stoichiometrically equivalent amounts of the coating materials (allylamine in case of Si NP-NH2, 10-undecenyl azide in case of Si NP-N3 and 1-butenoic acid in case of Si NP-COOH) to exclude cytotoxicity arising from the coating material molecules possibly remaining in the stock solutions of Si NPs as impurities. However, at these concentrations no cytotoxic effects were found for these compounds.
Protection by cellular preincubation with vitamin E
NR8383 cells were plated as described above (1 × 104 cells/well; 50 μl/well) in F12-K medium containing 100 μM vitamin E. After 24 hours 50 μl of serial dilutions of Si NP were added to the wells to obtain the required final concentrations of Si NP (the final concentration of vitamin E upon addition of the Si NP was reduced to 50 μM/well). Upon incubation for another 24 hours MTT reagent was added and the MTT assay was performed as described above. Control experiments were run with vitamin E only or Si NP only; both showed no activity in the MTT test.
Protection by cellular preincubation with vitamin C
NR8383 cells were plated in a 96-well plate (1 × 104 cells/well; 50 μl/well) in F12-K medium and after 22 hours vitamin C was added to reach a concentration of 1 mM vitamin C. After 2 more hours of incubation 50 μl of serial dilutions of Si NP were added to the wells to obtain the required final concentrations of Si NP (this reduced the final concentration of vitamin C to 500 μM). Upon incubation for another 24 hours MTT reagent was added and the MTT assay was performed as described above. Control experiments were run with vitamin C only or Si NP only; both showed no activity in the MTT test.
Phagocytic Index measurement
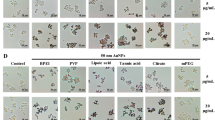
A NR8383 cell suspension (8 × 106 cells/ml) was plated in a 96-well plate (50 μl/well) in F12-K medium, followed by addition of 50 μl/well of serial dilutions of Si NP to obtain the required final concentrations of Si NP. Plain F12-K medium without Si NP and medium containing 100 μM CuSO4 were used as negative and positive control respectively. After 24 hours the cells were exposed to yellow green fluorescent latex beads (1 μm size) at a ratio of beads to cells in each well of 50:1. After 4 hours of incubation counting samples were taken from the wells and viewed first under a fluorescent microscope to visualize the fluorescent beads, followed by bright field view to visualize the cells [see additional file 1]. Also samples were taken out of each well to assess the cell viability by Trypan Blue exclusion test. The phagocytic index was determined by calculating the average number of fluorescent beads phagocytosed per viable cell and expressed as % of the negative control. Medium without Si NP served as negative control.
DCFH-DA assay
NR8383 cells
The cell suspension was adjusted to 2 × 105 cells/ml and seeded in a 96-well plate (50 μl/well) in F12-K medium. 50 μl/well of serial dilutions of Si NP in F12-K medium were added to obtain the required final concentrations of Si NP. A final concentration of 10 mM H2O2 was used as positive control and F12-K medium without nanoparticles as negative control. After 6 hours of exposure to the Si NP, 5 μl of a 20 mM solution of DCFH-DA were added to each well and the plates were incubated for another 18 hours in a 5% CO2 atmosphere at 37°C. The fluorescence was then measured on a fluorometer at 485 nm excitation and 538 nm emission wavelengths. The fluorescence induction factor for each concentration of Si NP was calculated by dividing the reading of each well by the average reading of the negative control and expressed as %. Control experiments were performed by incubating the Si NP at their test concentrations with DCFH-DA in the absence of cells to check the possibility of a positive fluorescence reading caused by reaction with Si NP alone.
Caco-2 cells
The cells were suspended in DMEM medium to a concentration of 1 × 105 cells/ml after trypisinization and were plated in a 96-well plate (100 μl/well). After 24 hours the cells were exposed to 100 μl/well of final concentrations of Si NP. Following another 6 hours of Si NP exposure, 5 μl of a 20 mM solution of DCFH-DA were added. The plate was further incubated for 18 hours before measurement of the fluorescence was carried out as described above. Control experiments were performed by incubating the Si NP at their test concentrations with DCFH-DA in the absence of cells to check the possibility of a positive fluorescence reading caused by reaction with Si NP alone.
Effect on isolated mitochondrial fraction
The isolated mitochondrial fraction (3 mg pellet/ml in PBS) was plated in a 96-well plate (50 μl/well) and serial dilutions of Si NP and 5 μl of DCFH-DA probe were added. The plate was incubated for 90 minutes at 37°C in a humidified 5% CO2 atmosphere. The plate was then measured at 485 nm excitation and 538 nm emission wavelength. Medium without Si NP and with 75 μM DNP in DMSO were used as negative and positive controls respectively. Results were expressed as % of negative control.
Statistical analysis
Data were analyzed with Origin Pro (version 8.0) graphing software. For statistical analysis a student's t-test was performed and data with P < 0.05 (compared to negative control, except for Figure 5) are marked with an asterisk (*) sign. Each data point represents the average from three independent experiments (n = 3) and is presented as the arithmetic mean ± SEM.
Abbreviations
- AFM:
-
atomic force microscopy
- ATCC:
-
American type culture collection
- ATP:
-
adenosine triphosphate
- BrdU:
-
5-bromo-2-deoxyuridine
- C3a:
-
C5a: complement component 3 and 5
- CdSe:
-
cadmium selenide
- CO2:
-
carbon dioxide
- CuSO4:
-
copper sulphate
- DCFH-DA:
-
2',7'-dichlorofluorescein-diacetate
- DIC:
-
disseminated intravascular coagulation
- DLS:
-
dynamic light scattering
- DMEM:
-
Dulbecco's modified eagle medium
- DMSO:
-
dimethyl sulfoxide
- DNA:
-
deoxyribonucleic acid
- DNP:
-
2,4-dinitrophenol
- EC50:
-
half maximal effective concentration
- ELISA:
-
enzyme-linked immunosorbant assay
- ETC:
-
electron transport chain
- FCS:
-
fetal calf serum
- F12-K:
-
Ham's F12- medium with Kaighn's modification
- FTIR:
-
Fourier transform infrared spectroscopy
- GPCR:
-
G-protein coupled receptor
- H2O2:
-
hydrogen peroxide
- LDH:
-
lactate dehydrogenase
- MTT:
-
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- ml:
-
milliliter
- mM:
-
millimolar
- μl:
-
microliter
- μM:
-
micromolar
- NAD:
-
nicotinamide dinucleotide
- NEAA:
-
non-essential amino acids
- ng:
-
nanogram
- NMR:
-
nuclear magnetic resonance
- PBS:
-
phosphate buffered saline
- PEG:
-
polyethylene glycol
- PLGA:
-
poly (D,L-lactide-co-glycolide)
- RBC:
-
red blood corpuscles
- ROS:
-
reactive oxygen species
- RPM:
-
rotations per minute
- SEM:
-
standard error of mean
- SFM:
-
scanning force microscopy
- Si:
-
silicon
- Si NP:
-
silicon nanoparticles
- Si NP-COOH:
-
carboxylic acid terminated silicon nanoparticles
- Si NP-NH2:
-
amine terminated silicon nanoparticles
- Si NP-N3:
-
azide terminated silicon nanoparticles
- TEER:
-
trans epithelial electric resistance
- TEM:
-
transmission electron microscopy
- TIRF:
-
total internal reflection fluorescence (microscopy)
- WBC:
-
white blood corpuscles
References
Derfus AM, Chan WCW, Bhatia SN: Probing the cytotoxicity of semiconductor quantum dots. Nano Lett 2004, 4: 11–18. 10.1021/nl0347334
Klauser F, Stijepovic R, Endstrasser N, Jaksch S, Memmel N, Scheier P: Oxidation study of silicon nanoparticle thin films on HOPG. Surf Sci 2009, 603: 2999–3004. 10.1016/j.susc.2009.08.007
Park J, Gu L, Maltzahn GV, Ruoslahti E, Bhatia SN, Sailor MJ: Biodegradable luminescent porous silicon nanoparticles for in vivo applications. Nat Mater 2009, 8(4):331–6. 10.1038/nmat2398
Burns AA, Vider J, Ow H, Herz E, Penate-Medina O, Baumgart M, Larson SM, Wiesner U, Bradbury M: Fluorescent silica nanoparticles with efficient urinary excretion for nanomedicine. Nano Lett 2009, 9(1):442–448. 10.1021/nl803405h
Rosso-Vasic M, Sprujit E, van Lagen B, De Cola L, Zuilhof H: Alkyl-functionalized oxide-free silicon nanoparticles: synthesis and optical properties. Small 2008, 4(10):1835–1841. 10.1002/smll.200800066
Ruizendaal L, Bhattacharjee S, Pournazari K, Rosso-Vasic M, de Haan LHJ, Alink GM, Marcelis ATM, Zuilhof H: Synthesis and cytotoxicity of silicon nanoparticles with covalently attached organic monolayers. Nanotoxicology 2009, 3(4):339–347. 10.3109/17435390903288896
Rosso-Vasic M, De Cola L, Zuilhof H: Efficient energy transfer between silicon nanoparticles and a Ru-polypyridine complex. J Phys Chem C 2009, 113: 2235–2240. 10.1021/jp804623w
Rosso-Vasic M, Spruijt E, Popovi Z, Overgaag K, van Lagen B, Grandidier B, Vanmaekelbergh D, Domínguez-Gutiérrez D, De Cola L, Zuilhof H: Amine-terminated silicon nanoparticles: synthesis, optical properties and their use in bioimaging. J Mater Chem 2009, 19: 5926–5933. 10.1039/b902671a
Oskuee RK, Dehshahri A, Shier WT, Ramezani M: Alkylcarboxylate grafting to polyethylenimine: a simple approach to producing a DNA nanocarrier with low toxicity. J Gene Med 2009, 11(10):921–932. 10.1002/jgm.1374
Sayin B, Somavarapu S, Li XW, Thanou M, Sesardic D, Alpar HO, Senel S: Mono-N-carboxymethyl chitosan (MCC) and N-trimethyl chitosan (TMC) nanoparticles for non-invasive vaccine delivery. Int J Pharm 2008, 363(1–2):139–148. 10.1016/j.ijpharm.2008.06.029
Nafee N, Schneider M, Schaefer UF, Lehr CM: Relevance of the colloidal stability of chitosan/PLGA nanoparticles on their cytotoxicity profile. Int J Pharm 2009, 381(2):130–139. 10.1016/j.ijpharm.2009.04.049
Hauck TS, Ghazani AA, Chan WC: Assessing the effect of surface chemistry on gold nanorod uptake, toxicity, and gene expression in mammalian cells. Small 2008, 4(1):153–159. 10.1002/smll.200700217
Mayer A, Vadon M, Rinner B, Novak A, Wintersteiger R, Fröhlich E: The role of nanoparticle size in hemocompatibility. Toxicology 2009, 258(2–3):139–147. 10.1016/j.tox.2009.01.015
Sadiq IM, Chowdhury B, Chandrasekaran N, Mukherjee A: Antimicrobial sensitivity of Escherichia coli to alumina nanoparticles. Nanomedicine 2009, 5(3):282–286.
Xu F, Yuan Y, Shan X, Liu C, Tao X, Sheng Y, Zhou H: Long-circulation of hemoglobin-loaded polymeric nanoparticles as oxygen carriers with modulated surface charges. Int J Pharm 2009, 377(1–2):199–206. 10.1016/j.ijpharm.2009.05.015
Pathak A, Kumar P, Chuttani K, Jain S, Mishra AK, Vyas SP, Gupta KC: Gene expression, biodistribution, and pharmacoscintigraphic evaluation of chondroitin sulfate-PEI nanoconstructs mediated tumor gene therapy. ACS Nano 2009, 3(6):1493–1505. 10.1021/nn900044f
Zhang LW, Monteiro-Riviere NA: Mechanisms of quantum dot nanoparticle cellular uptake. Toxicol Sci 2009, 110(1):138–155. 10.1093/toxsci/kfp087
Nam HY, Kwon SM, Chung H, Lee SY, Kwon SH, Jeon H, Kim Y, Park JH, Kim J, Her S, Oh YK, Kwon IC, Kim K, Jeong SY: Cellular uptake mechanism and intracellular fate of hydrophobically modified glycol chitosan nanoparticles. J Control Release 2009, 135(3):259–267. 10.1016/j.jconrel.2009.01.018
Kim BY, Jiang W, Oreopoulos J, Yip CM, Rutka JT, Chan WC: Biodegradable quantum dot nanocomposites enable live cell labeling and imaging of cytoplasmic targets. Nano Lett 2008, 8(11):3887–3892. 10.1021/nl802311t
Orr G, Panther DJ, Phillips JL, Tarasevich BJ, Dohnalkova A, Hu D, Teeguarden JG, Pounds JG: Submicrometer and nanoscale inorganic particles exploit the actin machinery to be propelled along microvilli-like structures into alveolar cells. ACS Nano 2007, 1(5):463–475. 10.1021/nn700149r
Gupta K, Singh VP, Kurupati RK, Mann A, Ganguli M, Gupta YK, Singh Y, Saleem K, Pasha S, Maiti S: Nanoparticles of cationic chimeric peptide and sodium polyacrylate exhibit striking antinociception activity at lower dose. J Control Release 2009, 134(1):47–54. 10.1016/j.jconrel.2008.10.008
Geys J, De Vos R, Nemery B, Hoet PH: In vitro translocation of quantum dots and influence of oxidative stress. Am J Physiol Lung Cell Mol Physiol 2009, 297(5):L903–911. 10.1152/ajplung.00029.2009
Corsi F, De Palma C, Colombo M, Allevi R, Nebuloni M, Ronchi S, Rizzi G, Tosoni A, Trabucchi E, Clementi E, Prosperi D: Towards ideal magnetofluorescent nanoparticles for bimodal detection of breast-cancer cells. Small 2009, 5(22):2555–2564. 10.1002/smll.200900881
Shvedova AA, Castranova V, Kisin ER, Schwegler-Berry D, Murray AR, Gandelsman VZ, Maynard A, Baron PJ: Exposure to carbon nanotube material: assessment of nanotube cytotoxicity using human keratinocyte cells. J Toxicol Environ Health A 2003, 66(20):1909–1926. 10.1080/713853956
Xia T, Korge P, Weiss JN, Li N, Venkatesen MI, Sioutas C, Nel A: Quinones and aromatic chemical compounds in particulate matter induce mitochondrial dysfunction: implications for ultrafine particle toxicity. Environ Health Perspect 2004, 112(14):1347–1358. 10.1289/ehp.7167
Green M, Howman E: Semiconductor quantum dots and free radical induced DNA nicking. Chem Commun 2005 (1):121–123.
Hussain SM, Hess KL, Gearhart JM, Geiss KT, Schlager JJ: In vitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicol in Vitro 2005, 19(7):975–983. 10.1016/j.tiv.2005.06.034
Sayes CM, Gobin AM, Ausman KD, Mendez J, West JL, Colvin VL: Nano-C60 cytotoxicity is due to lipid peroxidation. Biomaterials 2005, 26(36):7587–7595. 10.1016/j.biomaterials.2005.05.027
Foster KA, Galeffi F, Gerich FJ, Turner DA, Muller M: Optical and pharmacological tools to investigate the role of mitochondria during oxidative stress and neurodegeneratation. Prog Neurobiol 2006, 79(3):136–171. 10.1016/j.pneurobio.2006.07.001
Lin W, Huang YW, Zhou XD, Ma Y: Toxicity of cerium oxide nanoparticles in human lung cancer cells. Int J Toxicol 2006, 25(6):451–457. 10.1080/10915810600959543
Limbach LK, Wick P, Manser P, Grass RN, Bruinink A, Stark WJ: Exposure of engineered nanoparticles to human lung epithelial cells: influence of chemical composition and catalytic activity on oxidative stress. Environ Sci Technol 2007, 41(11):4158–4163. 10.1021/es062629t
Monteiller C, Tran L, MacNee W, Faux S, Jones A, Miller B, Donaldson K: The pro-inflammatory effects of low-toxicity low-solubility particles, nanoparticles and fine particles, on epithelial cells in vitro: the role of surface area. Occup Environ Med 2007, 64(9):609–615. 10.1136/oem.2005.024802
Schubert D, Dargusch R, Raitano J, Chan SW: Cerium and yttrium oxide nanoparticles are neuroprotective. Biochem Biophys Res Commun 2006, 342(1):86–91. 10.1016/j.bbrc.2006.01.129
Pan Y, Leifert A, Ruau D, Neuss S, Bornemann J, Schmid G, Brandau W, Simon U, Jahnen-Dechent W: Gold nanoparticles of diameter 1.4 nm trigger necrosis by oxidative stress and mitochondrial damage. Small 2009, 5(18):2067–2076. 10.1002/smll.200900466
Shiohara A, Hanada S, Prabakar S, Fujioka K, Lim TH, Yamamoto K, Northcote PT, Tilley RD: Chemical reactions on surface molecules attached to silicon quantum dots. J Am Chem Soc 2010, 132(1):248–253. 10.1021/ja906501v
Goodman CM, McCusker CD, Yilmaz T, Rotello VM: Toxicity of gold nanoparticles functionalized with cationic and anionic side chains. Bioconjugate Chem 2004, 15(4):897–900. 10.1021/bc049951i
Reddy LH, Sharma RK, Chuttani K, Mishra AK, Murthy RR: Etoposide-incorporated tripalmitin nanoparticles with different surface charge: formulation, characterization, radiolabelling, and biodistribution studies. AAPS J 2004, 6(3):e23. 10.1208/aapsj060323
Sengupta S, Tyagi P, Velpandian T, Gupta YK, Gupta SK: Etoposide encapsulated in positively charged liposomes: pharmacokinetic studies in mice and formulation stability studies. Pharmacol Res 2000, 42(5):459–464. 10.1006/phrs.2000.0714
Levchenko TS, Rammohan R, Lukyanov AN, Whiteman KR, Torchilin VP: Liposome clearance in mice: the effect of a separate and combined presence of surface charge and polymer coating. Int J Pharm 2002, 240(1–2):95–102. 10.1016/S0378-5173(02)00129-1
Aoki H, Sun C, Fuji K, Miyajima K: Disposition kinetics of liposomes modified with synthetic aminoglycolipids in rats. Int J Pharm 1995, 115: 183–191. 10.1016/0378-5173(94)00252-Z
Hernandez-Caselles T, Villalain J, Gomez-Fernandez JC: Influence of liposome charge and composition on their interaction with human blood serum proteins. Mol Cell Biochem 1993, 120: 119–126. 10.1007/BF00926084
Xia T, Kovochich M, Brant J, Hotze M, Sempf J, Oberley T, Sioutas C, Yeh JI, Wiesner MR, Nel AE: Comparison of the abilities of ambient and manufactured nanoparticles to induce cellular toxicity according to an oxidative stress paradigm. Nano Lett 2006, 6(8):1794–1807. 10.1021/nl061025k
Stone V, Donaldson K: Nanotoxicology: signs of stress. Nat Nanotechnol 2006, 1(1):23–24. 10.1038/nnano.2006.69
Foucaud L, Wilson MR, Brown DM, Stone V: Measurement of reactive species production by nanoparticles prepared in biologically relevant media. Toxicol Lett 2007, 174(1–3):1–9. 10.1016/j.toxlet.2007.08.001
Wilson MR, Lightbody JH, Donaldson K, Sales J, Stone V: Interactions between ultrafine particles and transition metals in vivo and in vitro. Toxicol Appl Pharmacol 2002, 184(3):172–179. 10.1006/taap.2002.9501
Donaldson K, Aitken R, Tran L, Stone V, Duffin R, Forrest G, Alexander A: Carbon nanotubes: a review of their properties in relation to pulmonary toxicology and workplace safety. Toxicol Sci 2006, 92(1):5–22. 10.1093/toxsci/kfj130
Brown DM, Donaldson K, Borm PJ, Schins RP, Dehnhardt M, Gilmour P, Jimenez LA, Stone V: Calcium and ROS-mediated activation of transcription factors and TNF-alpha cytokine gene expression in macrophages exposed to ultrafine particles. Am J Physiol Lung Cell Mol Physiol 2004, 286(2):L344-L353. 10.1152/ajplung.00139.2003
Li N, Sioutas C, Cho A, Schmitz D, Misra C, Sempf J, Wang M, Oberley T, Froines J, Nel A: Ultrafine particulate pollutants induce oxidative stress and mitochondrial damage. Environ Health Perspect 2003, 111(4):455–460. 10.1289/ehp.6000
Karataş ÖF, Sezgin E, Aydin Ö, Çulha M: Interaction of gold nanoparticles with mitochondria. Colloids Surf B Biointerfaces 2009, 71(2):315–318. 10.1016/j.colsurfb.2009.02.020
Supino R: Methods in Molecular Biology. In Vitro Toxicity Testing Protocols. Volume 43. Edited by: S. O'Hare and C. K. Atterwill. Humana Press, Totowa, NJ; Ch. 16
Acknowledgements
The authors would like to thank the Wageningen UR strategic research program Bionanotechnology and Graduate School VLAG for generous funding and colleagues of Wageningen University animal housing (CKP) facility for providing biotechnical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
SB, LHJDH, GMA, IMCMR conceived and designed the experiments. SB, NME (supervised by LHJDH) and XJ (supervised by SB) performed the experiments. ATMM and HZ supervised the synthesis of Si NP. SB analyzed the data and wrote the manuscript. ATMM, HZ, GMA and IMCMR supervised the project and corrected the manuscript. All authors read, corrected and approved the final manuscript.
Electronic supplementary material
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Bhattacharjee, S., de Haan, L.H., Evers, N.M. et al. Role of surface charge and oxidative stress in cytotoxicity of organic monolayer-coated silicon nanoparticles towards macrophage NR8383 cells. Part Fibre Toxicol 7, 25 (2010). https://doi.org/10.1186/1743-8977-7-25
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1743-8977-7-25