
Abstract
Background
ACE2 is a novel homologue of angiotensin converting enzyme (ACE). ACE2 is highly expressed in human heart and animal data suggest that ACE2 is an essential regulator of cardiac function in vivo. Since overactivity of the renin-angiotensin system contributes to the progression of heart failure, this investigation assessed changes in gene expression of ACE2, ACE, AT1 receptor and renin in the human failing heart.
Methods
The sensitive technique of quantitative reverse transcriptase polymerase chain reaction was used to determine the level of mRNA expression of ACE and ACE2 in human ventricular myocardium from donors with non-diseased hearts (n = 9), idiopathic dilated cardiomyopathy (IDC, n = 11) and ischemic cardiomyopathy (ICM, n = 12). Following logarithmic transformation of the data, a one-way analysis of variance was performed for each target gene followed by a Dunnett's test to compare the two disease groups IDC and ICM versus control.
Results
As anticipated, ACE mRNA was found to be significantly increased in the failing heart with a 3.1 and 2.4-fold up-regulation found in IDC and ICM relative to non-diseased myocardium. Expression of ACE2 mRNA was also significantly up-regulated in IDC (2.4-fold increase) and ICM (1.8-fold increase) versus non-diseased myocardium. No change in angiotensin AT1 receptor mRNA expression was found in failing myocardium and renin mRNA was not detected.
Conclusions
These data suggest that ACE2 is up-regulated in human IDC and ICM and are consistent with the hypothesis that differential regulation of this enzyme may have important functional consequences in heart failure. This strengthens the hypothesis that ACE2 may be a relevant target for the treatment of heart failure and will hopefully spur further studies to clarify the functional effects in human myocardium of ACE2 derived peptides.
Similar content being viewed by others


Background
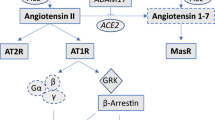
Angiotensin converting enzyme (ACE) catalyses the formation of angiotensin II (Ang II) and thereby plays a key role in cardiorenal regulation and blood pressure control. Overactivity of the renin-angiotensin system contributes to the pathophysiology of hypertension and heart failure, which is reflected in the use of ACE inhibitors in the treatment of these conditions. Interestingly, ACE2, a novel homolog of ACE, has recently been identified by two independent groups [1, 2]. ACE2 is a membrane-associated carboxypeptidase which is highly expressed in human heart, kidney, testis [1, 2] and gastrointestinal system [3]. ACE2 contains a single catalytic domain that is 42% identical to that of ACE. Importantly, ACE2 shows a markedly different substrate preference than ACE. Thus, of the key peptides of the renin-angiotensin system, ACE2 only cleaves Ang II with high catalytic efficiency [4], although ACE2 does have affinity for Ang I and can therefore compete with ACE for this substrate. In addition, the key product resulting from ACE2-induced hydrolysis of Ang II is Ang 1-7 [2, 4], whose functional effects (vasodilator, anti-proliferative) oppose those of Ang II [5, 6]. Taken together, these data are shaping the hypothesis that ACE2 may act to balance the activity of the renin-angiotensin system [7]. This is complemented by an important recent functional study [8], which demonstrated that ACE2 mRNA levels are reduced in several strains of hypertensive rats. Furthermore, Crackower et al. demonstrated that targeted disruption of ACE2 in mice results in a cardiac contractility defect, increased cardiac levels of Ang II and up-regulation of hypoxia-inducible genes in the heart [8].
Therefore, given the evidence that ACE2 may be an important regulator of the renin-angiotensin system and cardiac function, this study used the highly sensitive technique of real time quantitative reverse transcriptase PCR (QRT-PCR) to probe whether ACE2 and other components of the renin-angiotensin system are differentially regulated in the human failing heart.
Methods
Samples of human left ventricular free wall, which had been snap frozen and stored at -80°C, were selected based on the donor's clinical history and histological examination of the ventricular samples by a qualified pathologist. Tissues were obtained with approval from the appropriate ethics committees and in all cases with the full informed consent of the donor or their next of kin. The tissues were classified as Group 1, control, non diseased; Group 2, idiopathic dilated cardiomyopathy (IDC); Group 3, ischemic cardiomyopathy (ICM) (see Table 1 for donor information). All samples for groups 2 and 3 were obtained from hearts removed at transplantation. RNA was extracted using TriZol (Invitrogen Life Technologies) according to the manufacturer's protocols. Determining the optical density at 260 and 280 nm assessed the concentration and purity of the RNA, with only RNA yielding a ratio greater than 1:7 (260 : 280 nm) being used. All RNA solutions were diluted to 1 ug/ul in nuclease free water containing RNase inhibitor (N808-0119 Applied Biosystems). The quality of total RNA was further assessed by two methods. Firstly the RNA samples were run down denaturing agarose gels and only samples that had intact 18S and 28S ribosomal bands were used. Secondly, it was determined whether the samples were able to produce robust amplification of actin and transferrin receptor mRNA transcripts.
Detection of mRNA was by QRT-PCR (ABI Prism 7700 Sequence Detection System; Applied Biosystems) as previously described [9]. Forward and reverse primers and fluorogenic probes (see Table 2) were designed using Primer Express software (Applied Biosystems). The primer probe set for each gene was designed so that it crosses two exons, thereby amplifying cDNA and not genomic DNA. In every PCR reaction, the level of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was also determined. Duplexing the assay with GAPDH further confirms sample integrity and excludes the possibility of the low expression of genes of interest being a result of sample degradation. In all samples used in the current study, GAPDH was greater than 1 × 104 copies, indicating high quality mRNA. The copy number of each target gene was expressed as absolute copies per 100 ng of total RNA.
For the statistical analysis a logarithmic transformation (log to the base 10) of the data was carried out because this data is log-normally distributed. A two-way analysis of covariance (using the Genstat statistical software) demonstrated that the GAPDH Ct values contribute to the analysis as covariates by reducing the background noise against which to measure the differences between the control and disease groups, thereby increasing sensitivity. However, the significance of the TargetGene × Group interaction term (p = 0.013) indicates that the group differences are not consistent across the target genes. Thus a separate one-way analysis of covariance of each target gene was preferable (using the Cellula software). As a post-hoc test to allow for multiple comparisons, Dunnett's test was carried out comparing both of the disease groups, IDC and ICM, versus the control group. Statistical significance was assigned at P < 0.05.
Results
Atrial natriuretic factor (ANF) mRNA was evaluated as a positive control and was confirmed to be up-regulated by approximately 13 and 8-fold in IDC and ICM, respectively, versus the control group (see Table 3; Figure 1). Also in accordance with previous reports, relative to non-diseased ventricle, ACE mRNA was increased approximately 3-fold and 2-fold in IDC and ICM, respectively. ACE2 mRNA was expressed in non-diseased ventricle (see Table 3, Figure 1) and expression was further enhanced in IDC (~2 fold increase) and ICM (~2-fold increase). Expression of the angiotensin AT1 receptor mRNA was not changed in either of the heart failure groups relative to control. Renin mRNA was not present (zero copies) in ventricle from either of the groups. The renin primer probe set was validated by demonstrating expression of renin mRNA (mean copy number = 1158) in human renal cortex from three donors.

Quantitative expression profile of ACE2; ACE; ANF and AT 1 in control; IDC and ICM human left ventricular myocardium. ACE2; ACE; ANF and AT1 mRNA copy number in human left ventricular myocardium classified as Control (non-diseased) (n = 9); IDC-idiopathic dilated cardiomyopathy (n = 11); ICM-ischemic cardiomyopathy (n = 12). Corresponding levels of the housekeeping gene GAPDH are also shown. Each bar represents the geometric mean mRNA copy number with 95 % CI on a log scale. * – P < 0.05 Vs. Control.
Discussion
First identified in 2000, ACE2 is known to be highly expressed in human heart and may act to oppose the activity of the renin-angiotensin system by converting Ang II to the vasodilator Ang I-7 and by competing with ACE for Ang I [1, 2, 4, 10]. In addition, recent animal data suggest that ACE2 can modulate cardiac contractility, suppress cardiac Ang II levels and down-regulate hypoxia-induced pro-apoptotic genes [8], pointing to a role for ACE2-derived peptides in the regulation of cardiac function. Given that overactivity of the renin-angiotensin system is implicated in the pathophysiology of chronic heart failure, this study explored whether cardiac ACE2 may be differentially regulated in this condition. The technique of QRT-PCR was applied as we have found this to be the most quantitative and sensitive technique for probing changes in the expression of specific genes in human tissues [11].
To validate the use of QRT-PCR to identify differentially regulated genes in heart failure, ANF was studied as a positive control. As previously reported [12], ANF mRNA was found to be abundantly expressed in left ventricular myocardium and to be up-regulated in idiopathic dilated cardiomyopathy. ANF mRNA expression was also 8-fold up-regulated in the ischemic cardiomyopathy group although the change (relative to control) narrowly missed being statistically significant (P = 0.058). Several components of the renin-angiotensin system were subsequently studied. In agreement with previous workers [13], ACE mRNA was found to be up-regulated in both forms of heart failure, in accordance with the hypothesis that local production of Ang II is enhanced in the failing heart. Interestingly, ACE2 appeared more highly expressed than ACE in the non-failing ventricle and, like ACE, the levels of ACE2 mRNA were found to be significantly increased in idiopathic and ischemic cardiomyopathy.
The observation that ACE2 appears more highly expressed than ACE in human left ventricle should be interpreted cautiously as differences in primer probe set efficiencies can make difficult the quantitative comparison of expression levels of different targets. In this case, primer probe sets for ACE and ACE2 are believed to be of similar efficiency since they gave similar threshold values when used in a quantitative PCR with 10,000 genomic equivalents as the template. Therefore, the current data coupled to previous reports [3, 14] suggests that the relative expression levels of ACE:ACE2 mRNA is likely to vary significantly dependent on the species and tissue.
Unlike mRNA expression for ACE and ACE2, that of the angiotensin AT1 receptor was not found to be differentially expressed in failing myocardium. This contrasts with previous data [15, 16] indicating reduced AT1receptor expression in heart failure. Since the latter studies were carried out on right ventricular biopsies, it is possible that regional variation occurs in ventricular AT1 receptor expression in the failing heart. Finally, in contrast to all other genes studied, no expression (zero copies) of renin mRNA was detected in either non-failing or failing left ventricular myocardium. The question of whether renin may be produced locally in the myocardium has been an issue of controversy over the years but has not to our knowledge previously been addressed using QRT-PCR. However, even with this sensitive detection methodology, no ventricular expression (zero copies) was detected. This was not a methodology failure as the primer probe set used for renin transcript amplification worked as expected in human renal cortex. Therefore, these data are in agreement with previous findings [17], suggesting that the renin gene is not expressed in cardiac tissues.
Therefore, the key novel result from the current work is that ACE2 mRNA expression is increased in the human failing heart. This finding sits well with the emerging hypothesis that ACE2 may counterbalance the effects of stimulation of the renin-angiotensin system by reducing Ang II levels, and generating peptide products such as Ang1-7, which may benefit cardiac function. Indeed, recent data suggest that administration of Ang 1-7 can ameliorate the development of ischemia-induced heart failure in rats [18, 19]. Since the completion of the current study, it has been discovered that the G protein coupled receptor, Mas, is a high affinity, functional receptor for Ang 1-7 [20]. The question of whether Mas, like ACE2, is also differentially regulated in heart failure will be an interesting subject for future studies to address. Whether up regulation of ACE2 has important functional consequences in the human failing heart remains to be determined, however, the emerging data raise the intriguing possibility that a novel therapeutic approach to heart failure may be achieved through enhancement of ACE2 activity.
It is worth noting some limitations of the current work. First, only changes in mRNA have been addressed. While in the case of ACE and ANF it has been shown by other workers that in the failing heart increased mRNA translates into increased protein, this remains to be proven for ACE2. In addition, it will be of interest to determine which cell types in the failing heart increase the expression of ACE2 mRNA and protein. Secondly, while the experimental groups were matched for sex and age as closely as possible, the donors in both the heart failure groups were receiving various pharmacological therapies. Thus, we must acknowledge the possibility that these therapies could have influenced the levels of expression of ACE2 mRNA. Clarification of this question will likely require studies in animal models of heart failure where background can be strictly controlled. Finally, only end stage heart failure has been examined in this study, thereby giving no temporal data regarding the expression of ACE2 during the development of failure.
Conclusion
This study demonstrates that increased expression of ACE2 mRNA occurs in the human failing myocardium. This strengthens the hypothesis that ACE2 may be a relevant target for the treatment of heart failure and will hopefully spur further studies to clarify the functional effects in human myocardium of ACE2 derived peptides.
References
Donoghue M, Hsieh F, Baronas E, Godbout K, Gosselin M, Stagliano N, Donovan M, Woolf B, Robison K, Jeyaseelan R, Breitbart RE, Acton S: A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9. Circ Res. 2000, 87: E1-9.
Tipnis SR, Hooper NM, Hyde R, Karran E, Christie G, Turner AJ: A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J Biol Chem. 2000, 275: 33238-33243. 10.1074/jbc.M002615200.
Harmer D, Gilbert M, Borman R, Clark KL: Quantitative mRNA expression profiling of ACE 2, a novel homologue of angiotensin converting enzyme. FEBS Lett. 2002, 532: 107-110. 10.1016/S0014-5793(02)03640-2.
Vickers C, Hales P, Kaushik V, Dick L, Gavin J, Tang J, Godbout K, Parsons T, Baronas E, Hsieh F, Acton S, Patane M, Nichols A, Tummino P: Hydrolysis of biological peptides by human angiotensin-converting enzyme-related carboxypeptidase. J Biol Chem. 2002, 277: 14838-14843. 10.1074/jbc.M200581200.
Ferrario CM, Chappell MC, Tallant EA, Brosnihan KB, Diz DI: Counterregulatory actions of angiotensin-(1–7). Hypertension. 1997, 30: 535-541.
Ferrario CM: Angiotension-(1–7) and antihypertensive mechanisms. J Nephrol. 1998, 11: 278-283.
Boehm M, Nabel EG: Angiotensin-converting enzyme 2-a new cardiac regulator. N Engl J Med. 2002, 347: 1795-1797. 10.1056/NEJMcibr022472.
Crackower MA, Sarao R, Oudit GY, Yagil C, Kozieradzki I, Scanga SE, Oliveira-dos-Santos AJ, da Costa J, Zhang L, Pei Y, Scholey J, Ferrario CM, Manoukian AS, Chappell MC, Backx PH, Yagil Y, Penninger JM: Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature. 2002, 417: 822-828. 10.1038/nature00786.
Bowen WP, Carey JE, Miah A, McMurray HF, Munday PW, James RS, Coleman RA, Brown AM: Measurement of cytochrome P450 gene induction in human hepatocytes using quantitative real-time reverse transcriptase-polymerase chain reaction. Drug Metab Dispos. 2000, 28: 781-788.
Oudit GY, Crackower MA, Backx PH, Penninger JM: The Role of ACE2 in Cardiovascular Physiology. Trends Cardiovasc Med. 2003, 13: 93-101. 10.1016/S1050-1738(02)00233-5.
Coleman RA, Clark KL: Target validation using human tissue: from gene expression to function. Targets. 2003, 2: 58-64. 10.1016/S1477-3627(03)02298-0.
Saito Y, Nakao K, Arai H, Nishimura K, Okumura K, Obata K, Takemura G, Fujiwara H, Sugawara A, Yamada T, et al: Augmented expression of atrial natriuretic polypeptide gene in ventricle of human failing heart. J Clin Invest. 1989, 83: 298-305.
Studer R, Reinecke H, Muller B, Holtz J, Just H, Drexler H: Increased angiotensin-I converting enzyme gene expression in the failing human heart. Quantification by competitive RNA polymerase chain reaction. J Clin Invest. 1994, 94: 301-310.
Tikellis C, Johnston CI, Forbes JM, Burns WC, Burrell LM, Risvanis J, Cooper ME: Characterization of renal angiotensin-converting enzyme 2 in diabetic nephropathy. Hypertension. 2003, 41: 392-397. 10.1161/01.HYP.0000060689.38912.CB.
Haywood GA, Gullestad L, Katsuya T, Hutchinson HG, Pratt RE, Horiuchi M, Fowler MB: AT1 and AT2 angiotensin receptor gene expression in human heart failure. Circulation. 1997, 95: 1201-1206.
Regitz-Zagrosek V, Fielitz J, Dreysse R, Hildebrandt AG, Fleck E: Angiotensin receptor type 1 mRNA in human right ventricular endomyocardial biopsies: downregulation in heart failure. Cardiovasc Res. 1997, 35: 99-105. 10.1016/S0008-6363(97)00089-8.
von Lutterotti N, Catanzaro DF, Sealey JE, Laragh JH: Renin is not synthesized by cardiac and extrarenal vascular tissues. A review of experimental evidence. Circulation. 1994, 89: 458-470.
Ferrario CM: Does angiotensin-(1–7) contribute to cardiac adaptation and preservation of endothelial function in heart failure?. Circulation. 2002, 105: 1523-1525. 10.1161/01.CIR.0000013787.10609.DC.
Loot AE, Roks AJ, Henning RH, Tio RA, Suurmeijer AJ, Boomsma F, van Gilst WH: Angiotensin-(1–7) attenuates the development of heart failure after myocardial infarction in rats. Circulation. 2002, 105: 1548-1550. 10.1161/01.CIR.0000013847.07035.B9.
Santos RA, Simoes e Silva AC, Maric C, Silva DM, Machado RP, de Buhr I, Heringer-Walther S, Pinheiro SV, Lopes MT, Bader M, Mendes EP, Lemos VS, Campagnole-Santos MJ, Schultheiss HP, Speth R, Walther T: Angiotensin-(1–7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc Natl Acad Sci USA. 2003, 100: 8258-8263. 10.1073/pnas.1432869100.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1741-7015/2/19/prepub
Acknowledgements
Thank you to PRISM Training & Consultancy Ltd; http://www.prismtc.co.uk(Ian Macpherson and Paul Nelson) for their advice and assistance in statistical analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
None declared.
Authors' contributions
AG and KC participated in the hypothesis generation, experimental design, statistical analysis and writing of the manuscript. MG participated in collection of the study material, collation of donors' clinical history and scientific discussion. JA participated in RNA extraction and setting up and performance of QRT-PCR. All authors have read and approved the final version of the manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Goulter, A.B., Goddard, M.J., Allen, J.C. et al. ACE2 gene expression is up-regulated in the human failing heart. BMC Med 2, 19 (2004). https://doi.org/10.1186/1741-7015-2-19
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1741-7015-2-19