
Abstract
Mitochondria play important roles in cellular energy metabolism, free radical generation, and apoptosis. Defects in mitochondrial function have long been suspected to contribute to the development and progression of cancer. In this review article, we aim to provide a brief summary of our current understanding of mitochondrial genetics and biology, review the mtDNA alterations reported in various types of cancer, and offer some perspective as to the emergence of mtDNA mutations, their functional consequences in cancer development, and therapeutic implications.
Similar content being viewed by others

Introduction
Mitochondrial defects have long been suspected to play an important role in the development and progression of cancer. Over 70 years ago, Warburg pioneered the research on mitochondrial respiration alterations in the context of cancer and proposed a mechanism to explain how they evolve during the carcinogenic process [1, 2]. In his series of landmark publications, he hypothesized that a key event in carcinogenesis involved the development of an "injury" to the respiratory machinery, resulting in compensatory increases in glycolytic ATP production. Eventually, malignant cells would satisfy their energy needs by producing a large portion of their ATP through glycolytic mechanisms rather than through oxidative phosphorylation. Due to the inherent inefficiency of glycolytic ATP generation, this represents a somewhat unique metabolic state of the malignant cells and would require high consumption of glucose to fulfill cellular energy requirements. This is in contrast to many normal cells, which use oxidative phosphorylation as the preferred means of ATP generation with high efficiency. The differences in energy metabolism between normal and cancer cells constitute a biochemical basis to speculate that therapeutic strategies might be developed to selectively kill cancer cells due to their inherently compromised respiratory state [1–3].
Since the initial publications by Warburg over half a century ago, a number of cancer-related mitochondrial defects have been identified and described in the literature. These defects include altered expression and activity of respiratory chain subunits and glycolytic enzymes, decreased oxidation of NADH-linked substrates, as well as mitochondrial DNA (mtDNA) mutations. While there are many reports of these phenomena, the mechanisms responsible for the initiation and evolution of mtDNA mutations, and their roles in the development of cancer, drug resistance, and disease progression still remain to be elucidated. This review article provides a brief summary of our current understanding of mitochondrial genetics and biological function, reviews the reported mtDNA alterations in human malignancies, and discusses possible mechanisms by which mtDNA mutations emerge in cancer cells and their clinical relevance.
Key aspects of mitochondrial biology and genetics
Mitochondria are semi-autonomous organelles that perform essential functions in cellular metabolism and the regulation of cell death. While the exact origin of mitochondria is still uncertain, it is widely believed that they arose from an endosymbiotic relationship between a glycolytic proto-eukaryotic cell and an oxidative bacterium [4–6]. Mitochondria possess a double-membrane structure and contain their own genome along with their own transcription, translation, and protein assembly machinery. As such, they are able to maintain genomic independence from the nucleus. However, as a consequence of proto-mitochondrial genes integrating into the nuclear genome throughout evolution, most mitochondrial proteins are encoded by nuclear DNA (nDNA) and imported into mitochondria. Although the replication of mitochondrial DNA (mtDNA) is not synchronized with nDNA replication, the overall number of mitochondria per cell remains fairly constant for specific cell types during proliferation, suggesting that the generation of mitochondria is largely influenced by extra-mitochondrial signal transduction events. This conclusion is further supported by the observation that mitochondrial biosynthesis continues even when mtDNA is deleted. Thus, the replication of mitochondria does not require the presence of mtDNA.
Energy and metabolism
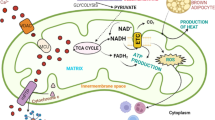
The most well-known and best-characterized function of mitochondria is the production of adenosine triphosphate (ATP) through oxidative phosphorylation. This process is accomplished by a series of protein complexes, collectively known as the respiratory chain, encoded by both nDNA and mtDNA. The complete respiratory chain contains at least 87 polypeptides, 13 of which are encoded by mtDNA. Hence, the majority of the respiratory chain components are nuclear-encoded and imported into mitochondria after their translation in the cytosol (Figure 1). Thus, oxidative phosphorylation is a unique biochemical process achieved by a well-coordinated effort of the protein products from two separate genomes (nuclear and mitochondrial) working in concert within the same cells. However, this process is not absolutely required to fulfill cellular energy requirements. Glycolysis can also generate ATP and provides a compensatory mechanism when oxidative phosphorylation becomes inefficient as a consequence of defects in the respiratory chain.

The mitochondrial electron transport chain complexes are encoded by two genetic systems: the mitochondrial DNA (mtDNA) and the nuclear DNA (nDNA). The mtDNA codes for 7 NADH dehydrogenase (ND) subunits for complex I, a cytochrome b for complex III, 3 cytochrome c oxidase (COX) subunits for complex IV, and 2 ATPase (ATPase6/8) for complex V. Complex II is solely encoded by nDNA. The solid arrows indicate the direction of electron flow in the respiration chain; dashed arrows indicate the flow of genetic information from mtDNA or nDNA to RNA to protein; the numbers indicate the actual number of protein subunits coded by each genetic system. Specific inhibitors of RNA transcription or protein translation with relative selectivity for each genetic system are indicated in blue.
It is important to note that as electrons are transported through the respiratory chain during mitochondrial respiration, some of the electrons may escape or leak from electron transport complexes and react with molecular oxygen to form superoxide radicals (O2-). This diversion or bifurcation of electron flow occurs mainly at complexes I and III [7, 8]. It is reasonable to assume that certain mtDNA mutations may cause an alteration of the electron transport components that compromises the normal electron flow. This can lead to an increase of bifurcation and generation of superoxide radicals, which are subsequently converted into other reactive oxygen species (ROS). In fact, mitochondrial DNA mutations [summarized in this review] and increased oxidative stress [for a review see [9]] have been observed in various types of cancer cells in several independent studies. However, the direct link between mtDNA mutations and the increase of ROS generation in cancer cells has not been demonstrated experimentally.
In addition to their critical role in the fulfillment of cellular energy (ATP) requirements, mitochondria are also essential in the processing of important metabolic intermediates for various pathways involved in the metabolism of carbohydrates, amino acids, and fatty acids. The tricarboxylic acid cycle in the mitochondrial matrix is a prime example of a biochemical pathway where many important metabolic intermediates converge [10, 11]. A major portion of the urea cycle is also compartmentalized within the mitochondria where metabolic processing of nitrogen-containing amino acid intermediates occurs [12]. Fatty acids are broken down into two-carbon units by a series of β-oxidations and further processed to acetyl CoA in the mitochondrial matrix. Thus, while the generation of ATP via oxidative phosphorylation represents an important, but not essential mitochondrial function, mitochondria are indispensable to eukaryotic cells due to their extensive involvement in other critical metabolic processes. This may explain why ρ0 cells, in which the mtDNA is deleted [13], continue to maintain mitochondrial mass without the mtDNA-encoded respiratory protein components.
Apoptosis and cell survival
During the past decade, the most significant research advance in mitochondrial biology may be the discovery that mitochondria play an important role in apoptosis, a fundamental biological process by which cells die in a well-controlled or programmed manner. Several nDNA-encoded pro-apoptotic proteins including cytochrome c, apoptosis inducing factor (AIF), endonuclease G, and smac/DIABLO normally reside in the mitochondria where they perform known or yet unidentified physiological functions. However, once these protein factors are released from mitochondria, they trigger a series of biochemical events leading to activation of apoptotic signaling cascades [for a review see [14]]. Perhaps the most well characterized apoptotic cascade is the activation of caspases (a class of proteases) by cytochrome c and apaf-1 in the presence of ATP or dATP [15]. In contrast, translocation of AIF from mitochondria to the nucleus appears to cause apoptosis in a caspase-independent manner [16]. Because apoptosis plays a critical role in cancer development and in the cellular response to anticancer agents, the significance of mtDNA mutations in cancer is obviously an important area of current investigation. However, the exact role of mtDNA mutations in the cellular apoptotic response to anticancer agents has not been defined. Several groups have used respiration-deficient ρ0 cells lacking mtDNA to evaluate the role of respiration in drug sensitivity, achieving various results. For instance, Singh et al reported that ρ0 cells were resistant to adriamycin and photodynamic therapy, whereas these cells were equally sensitive to alkylating agents and γ-radiation as compared with their parental cells with respiration-competent mitochondria [17]. A study by Cai et al demonstrated that cytochrome c release and caspase activation in response to staurosporine treatment were maintained in ρ0 cells. However, unlike the case with ρ+ cells, in which a dramatic oxidation of intracellular glutathione (GSH) occurred following cytochrome c release, the thiol-disulfide redox state in apoptotic ρ0 cells was relatively unchanged. Thus, mitochondrial signaling of caspase activation can be separated from its bioenergetic function [18]. In contrast, Joshi et al reported that a functioning mitochondrial respiratory chain was required for cellular sensitivity to BMD188, a novel prostate cancer chemotherapeutic [19]. The varying reports regarding the sensitivity of ρ0 cells to apoptosis induction are likely a reflection of the different mechanisms of action of the various stimuli in question.
In addition to alterations in sensitivity to anticancer agents, ρ0 cells derived from human solid tumor cell lines have been shown to lose their capacity for anchorage-independent growth and tumorigenicity when injected into nude mice [20]. Both of these properties were restored when the ρ0 cells were fused with cells containing normal mitochondria, indicating that mtDNA is required to maintain a cancer phenotype. Other studies have shown that while ρ0 cells lose their capacity for anchorage-independent growth, they maintain their tumorigenicity in nude mice [21]. The conflicting results observed in ρ0 cells may indicate a complex interaction between the nuclear and mitochondrial genomes in transformation. Lack of tumorigenicity in some ρ0 cells may be a reflection of insufficient changes in nuclear-encoded genes to maintain tumorigenicity, whereas the tumorigenicity of other ρ0 cells may be a consequence of additional transforming events. It should be emphasized that although ρ0 cells represent a valuable in vitro experimental model system to investigate the role of mitochondrial respiration in drug sensitivity, their clinical relevance is uncertain. While malignant cells are often characterized by mtDNA mutations and partial deletions, their mtDNA is unlikely to be completely deleted. As such, cells carrying different known mtDNA mutations represent a more appropriate model system for the investigation of the effects of mtDNA mutations on cellular sensitivity to anti-cancer agents.
Genetics and mutations
The human mitochondrial genome has been completely sequenced and each gene contained within it has been identified and characterized [22, 23]. The human mtDNA is a supercoiled, double-stranded circular molecule of 16,569 base pairs in size. As mentioned earlier, it codes for 13 of the 87 proteins required for oxidative phosphorylation as well as the 12S and 16S rRNAs and 22 tRNAs required for protein synthesis in the mitochondria (Figure 1). It is of interest to note that other than the aforementioned 13 respiratory chain components, no other genes for structural proteins are found in human mtDNA. It is believed that the early mitochondrial genome derived from symbiotic bacteria in proto-eukaryotic cells may have contained other genes, which may have been lost or integrated into the nuclear DNA during evolution. One interesting example is that the nuclear-coded mitochondrial enzyme superoxide dismutase (SOD2) is more homologous to the bacterial SOD than to the eukaryotic cytosolic isozyme (SOD1). In fact, mtDNA migrates into the nucleus at a much higher frequency than nuclear DNA integrates into mitochondria [24, 25]. The retention of genes coding only for the respiratory chain components in the mitochondrial genome is likely the consequence of a selection process rather than a random event, although the underlying mechanism remains unclear.
Each mitochondrion contains 2–10 copies of its genome. Although the number of mitochondria per cell varies with cell type, an individual cell typically contains a fairly constant copy number of mtDNA. During fertilization, the mitochondria are transmitted through the oocyte's cytoplasm. Thus, the mitochondrial genome is distinct from the nuclear genome in that it follows a strict maternal inheritance pattern [For a more in-depth review refer to [26–28]]. Within a cell, mtDNA replication is semi-autonomous and is not synchronized with the S phase of the cell cycle. However, the two genetic systems (nuclear and mitochondrial) appear to be closely coordinated by yet unknown mechanisms. The overall biogenesis of mitochondria keeps pace with cell proliferation and cell growth.
Although the mitochondrion contains the necessary machinery to express its genome, many of its protein components are encoded by nuclear DNA. It should be noted that mitochondrial RNA transcription can be preferentially inhibited by ethidium, whereas transcription of RNA from nDNA is preferentially inhibited by α-amanitin. Likewise, protein synthesis in mitochondria is sensitive to inhibition by tetracycline and chloramphenicol, whereas cytosolic protein synthesis is selectively inhibited by cycloheximide (Figure 1). This differential sensitivity to metabolic inhibitors further supports the hypothesis that mitochondria originated from bacteria. This also provides a basis for using these metabolic inhibitors as important tools to investigate the interaction between the two genetic systems.
Another feature that distinguishes the mitochondrial and nuclear genomes is their inherent susceptibilities to damage. It is known that mtDNA is far more vulnerable to mutations than nuclear DNA due to its lack of histone protection, limited repair capacity, and close proximity to the electron transport chain, which constantly generates superoxide radicals. Considering mtDNA lacks introns, most mutations occur in the coding sequences and are thus, likely to be of biological consequence [29]. However, there may be intracellular mechanisms that offset some of the effects of mtDNA mutations. Recombination of mtDNA molecules between fused mitochondria has been well-established in yeast and plant cells [30, 31]. In spite of this, this phenomenon had not been demonstrated in mammalian cells until recently, probably because the mtDNA sequence of individuals tends to be relatively uniform due to its maternal inheritance pattern. Ono et al isolated two types of respiration-deficient cell lines with pathogenic mtDNA mutations from patients with mitochondrial diseases. Generation of hybrids via fusion with normal cells was able to restore morphology and respiratory enzyme activity, indicating the exchange of genetic contents between the mitochondria. The authors proposed that this may represent a mechanism to defend against mitochondrial dysfunction caused by the accrual of mtDNA lesions with age [32]. It is likely that this process is also relevant in the context of cancer and other diseases and serves to provide a buffering capacity to maintain mitochondrial function in the presence of mtDNA mutations.
Because each cell contains many mitochondria with multiple copies of mtDNA, it is possible that wild-type and mutant mtDNA can co-exist in a state called heteroplasmy. During cell division, mitochondria are randomly distributed to daughter cells. Over time, the proportion of the mutant mtDNA within the cell can vary and may drift toward predominantly mutant or wild type to achieve homoplasmy. Thus, the biological impact of a given mutation may vary, depending on the proportion of mutant mtDNAs carried by the individual. This effect contributes to the various phenotypes observed amongst family members carrying the same pathogenic mtDNA mutation. Somatic mutations in mtDNA and their potential clinical implications will be further discussed below.
Mitochondrial DNA alterations in cancer
Mutations in mtDNA have been identified in various types of human cancer. Additional file 1 provides a summary of mutations found in the non-coding region and coding regions of the mtDNA in cancer cells (see Additional file 1).
Breast cancer
Comprehensive studies examining mtDNA mutations in breast cancer have been published recently. A study by Tan et al used a combination of temporal temperature gel electrophoresis and direct DNA sequencing to screen the complete mitochondrial genome for mutations in 19 sets of paired normal and tumor tissues from the same patients. Somatic mutations were identified in 14 of the 19 patients (74%). The bulk of the mutations (81.5%) were restricted to the D-loop region. However, mutations were also detected in the 16S rRNA, ND2, and ATPase 6 genes [33]. In another study, the investigators analyzed fine needle aspirates of primary breast tumors and matched lymphocytes of 18 patients. As observed in the aforementioned study, somatic mutations were detected in the majority of the patients (61%) and most of the mutations identified were in the D-loop region. The remaining mutations were found in the ND1, ND4, ND5, and Cytochrome b genes [34]. Earlier studies also revealed point mutations and deletions in breast tumors. The 4977 bp common deletion was found in malignant and normal breast tissue of a smaller number of patients with abnormalities of the breast [35]. In addition to mutations, elevated expression of cytochrome c oxidase II mRNA was also detected in breast tumors in a number of patients as compared to the normal tissue [36].
Ovarian cancer
Liu et al analyzed mtDNA from matched normal and malignant ovarian tissue samples obtained from 10 patients with ovarian carcinomas. A PCR-based method was used to sequence the complete mitochondrial genome in both sets of tissues. Comparison of the DNA sequences obtained from the tumor and normal tissues enabled the identification of somatic mtDNA mutations. This study revealed a high incidence (60%) of somatic mutations in the tumor tissues examined. Most of the identified mutations were T -> C or G -> A transitions. The somatic mutations were restricted to 4 regions of the mitochondrial genome: the D-loop, 12S rRNA, 16S rRNA, and cytochrome b. The authors speculated that these regions may represent mutational hotspots in ovarian cancer [37].
Colorectal cancer
The integrity of the mitochondrial genome has been examined in several colorectal cancer cell lines as well as in primary patient tissues. One study [38] examined normal and malignant colon cells from 10 individuals using a PCR-based method to sequence the entire mitochondrial genome. 7 of 10 patients had somatic mtDNA mutations. The mutations were found in the 12S rRNA, 16S rRNA, ND1, ND4L, ND5, Cytochrome b, COXI, COXII, and COXIII genes. The majority of the mutations were transitions with the exception of one insertion. Somatic mutations in the non-coding D-loop region have also been identified in colon cancer patients. Somatic A -> T and G -> C transitions and deletions were detected using a combination of heteroduplex analysis and single-stranded conformation polymorphism (SSCP) analysis [39]. Additionally, alterations in the expression of mitochondrial-encoded respiratory chain subunits have been detected in primary tumor samples from colorectal cancer patients as well as patients with pre-malignant familial polyposis coli (FPC) syndrome. A study utilizing tissues from 15 colorectal cancer patients found that ND2 mRNA levels were elevated in the malignant tissue as compared to normal tissue from the same individuals [40]. Elevated expression of ND1 mRNA and 16S rRNA was also detected in the polyps of FPC patients as compared to normal colon tissue [41]. Analysis of the mitochondrial genome in colorectal cancer cell lines also revealed abnormalities. Elevated expression of 16S rRNA, ND4, ND4L, cytochrome b, COXII, ATPase 6, and ATPase 8 was detected in the HT-29 cell line [42]. Another study examining the Caco-2 cell line detected mtDNA deletions [43]. Other investigators have evaluated a link between mitochondrial microsatellite instability and cancer. Analysis of 45 sporadic colorectal cancers using PCR-SSCP detected microsatellite alterations involving insertion or deletion of a cytidine in a polycytidine tract in the D-loop region in 44% of the examined cases. The authors also identified frame shift mutations in ND1 and ND5. Some of the detected alterations were heteroplasmic, while others where homoplasmic. No large-scale deletions were found [44, 45].
Gastric cancer
PCR-SSCP was utilized to examine nuclear microsatellite instability, mtDNA large deletions, and mtDNA mutations in one study of 32 gastric carcinomas. Surprisingly, nuclear microsatellite instability was not correlated with the presence of mtDNA mutations or deletions in the specimens examined. This observation suggests a possibility that the nucleus and mitochondria may use different repair mechanisms to repair DNA damage. The common deletion of mtDNA was detected in 54% of cases, and mtDNA mutations were identified in 44% of cases, suggesting that major mtDNA alterations tend to be associated with gastric cancer. Most of the mutations corresponded to insertions/deletions in the D-loop region or transitions in ND1, ND5, and COXI [46]. Earlier studies revealed the presence of mutations in the D-loop of both non-malignant and malignant gastric tumors [47, 48].
Hepatic cancer
An analysis of 19 tumors from patients with hepatocellular carcinoma (HCC) indicated that mutations in the D-loop were a frequent event and could be used as a molecular tool for the determination of clonality [49]. Somatic mtDNA mutations were identified in a separate study in both the malignant and non-malignant liver tissue of HCC patients. The frequency of mutations was correlated with the degree of malignancy [50]. An earlier study examining hepatic tumors found that mtDNA deletions were present in the cirrhotic liver tissue surrounding the tumors, but not in the tumors themselves [51]. Further studies are required to clearly determine if mtDNA mutations are correlated with malignant transformation.
Esophageal cancer
Two recent reports investigated the frequency of D-loop mutations in esophageal cancer. One group focused on adenocarcinomas of Barrett's esophagus. In that study, D-loop alterations were identified in 40% of the patients examined [52]. The other study showed that D-loop mutations were much less frequent in esophageal cancer, occurring in only 5% of specimens analyzed [53]. Clearly, analysis of mtDNA from more esophageal tumor samples is needed in order to determine the frequency of D-loop mutations and their relevance in this cancer type.
Pancreatic cancer
An investigation of 15 pancreatic cancer cell lines and xenografts by Jones et al revealed mtDNA alterations in nearly every case. The mutations were identified in several different genes including 12S rRNA, 16S rRNA, ND1, ND2, COXI, COXII, ATPase 6, COXIII, ND4, ND4L, ND5, ND6, Cyt b, as well as the non-coding D-loop region. The authors also found a 6-fold to 8-fold increase in the mtDNA mass in the tumor cells as compared to normal pancreatic cells. The very high frequency of mtDNA mutations and the increase of mtDNA contents in pancreatic cancer cells led the authors to speculate that mtDNA may serve as a diagnostic tool [54]. It is not clear if the increase of mtDNA mass reflects an increased replication of mtDNA in response to defective mtDNA function due to extensive mutations.
Renal cell carcinoma
A well-characterized alteration of mtDNA associated with renal cell carcinoma (RCC) is a 264-bp deletion of the ND1 gene. This deletion eliminates 264 nucleotides of the mitochondrial genome between nucleotide positions 3323 and 3588 [55]. A separate study reported the loss of mtDNA and mRNA coding for subunit the ND3 gene [56]. Alterations in the expression of both mitochondrial and nuclear-encoded protein components of the respiratory chain have also been observed in renal cell carcinoma. The earlier study revealed that the steady-state transcript levels of all mitochondrial-encoded subunits were strikingly lower than in the corresponding normal renal tissue. In contrast, the levels of nuclear-encoded respiratory chain subunits were higher than in the normal controls [57]. A more recent study that examined tumor samples from 25 RCC patients demonstrated that mitochondrial enzyme and DNA contents of all tumors differed significantly from normal tissue. However, the authors found that decreased content of respiratory chain subunits rather than alterations in mtDNA content correlated with tumor aggressiveness. In particular, nearly all tumors lost ATP synthase activity in Complex V [58]. Their data supports the hypothesis that decreased capacity for oxidative phosphorylation and increased glycolysis may favor tumor growth and invasiveness.
Prostate cancer
Until recently, there were no reports of mtDNA alterations in prostate cancer. Two recent studies suggest that mtDNA mutations and deletions may play a role in prostate carcinogenesis. The first study examined mtDNA deletions in 34 malignant prostate specimens. The entire mtDNA was amplified using long distance PCR. Electrophoresis was used to visualize alterations in the mtDNA size due to deletions. The authors found that the average number of deletions increased with advanced age [59]. In the other study, however, Jeronimo et al sequenced the D-loop region, 16S rRNA, and NADH subunits in matched premalignant prostate lesions, primary prostate tumors, and normal lymphocytes from 16 patients. Mutations were identified in 3/16 patients. In spite of the relatively infrequent incidence of mutations, the mutations were found to occur early in disease progression and were detectable in bodily fluids during the early stages of the disease [60].
Brain tumors
Malignant glioma is the best-characterized type of brain tumor with respect to mtDNA alterations. The most frequent observation involves changes in the copy number of mtDNA. A study that examined 45 glioma specimens found that mtDNA was highly amplified in 87% of the cases. The authors also examined a nuclear-encoded gene that is frequently amplified in human cancers as a reference. They found that the erb-b gene copy number increased in only 18% of the tumor specimens, indicating the mtDNA alterations may be much more frequent than those of nuclear-encoded genes in malignant gliomas [61]. A separate study of 15 tumor sections confirmed the high frequency of mtDNA copy number changes [62]. Interestingly, the authors also detected a transfer of mtDNA to the nucleus, and showed that the nuclear localization of the mtDNA segment was correlated with the increase of mtDNA copy numbers. It has been proposed that mtDNA integration into the nuclear genome may represent a mechanism of oncogene activation [63]. A later study of astrocytic tumors indicated that the heteroplasmy of a well-characterized polymorphic site in the D-loop region was lost in the tumor specimens, but not in the normal brain tissue references [64]. Clearly, mtDNA alterations are a frequent event in the development and progression of brain tumors and are worthy of further investigation.
Thyroid cancer
It has been known for more than 2 decades that thyroid tumors contain abnormally high numbers of mitochondria [65]. More recent studies have indicated that mtDNA alterations are present in thyroid cancer cells. An examination of 21 thyroid tumors using two-dimensional gene scanning revealed that somatic mutations were present in 23% of analyzed specimens. The majority of the mutations that were found occurred in the genes coding for subunits of complex I of the respiratory chain. Their findings indicate that somatic mtDNA mutations may be involved in thyroid tumorigenesis and that accumulation of certain mutations may play a role in tumor progression [66]. A more recent study of 79 thyroid tumors reported that the mtDNA common deletion was identified in 100% of Hürthle cell tumors (n = 43), 33.3% of adenomas (n = 15), and in 18.8% of non-Hürthle cell papillary carcinomas. A number of somatic point mutations in mtDNA were also identified in the tumor tissues, especially in Hürthle cell tumors. The authors concluded that mtDNA alterations in the genes coding for Complex I and Complex IV of the respiratory chain may contribute to the development of thyroid tumors. Germline polymorphisms of the ATPase 6 gene were correlated with the occurrence of the mtDNA common deletion, which appears to be a hallmark of Hürthle cell tumors [67].
Other solid tumors
The frequency and types of mtDNA alterations in other solid tumors have not been as well characterized compared to the aforementioned cancer types. There are a limited number of reports regarding mtDNA mutations in solid tumors of the bladder, head and neck, and lung [68, 69]. In general, there appear to be two features of mtDNA alterations in cancer irrespective of tumor type. First, the majority of mutations are base transitions from T to C and G to A. Second, while there is diversity in the individual genes in which mutations occur, the D-loop region seems to be the most frequent site of somatic mutations across most tumor types. A more thorough analysis is required in order to determine the functional significance of specific mtDNA alterations and their role in cancer development.
Hematologic malignancies
Alterations in mtDNA in hematologic malignancies have been reported by several groups. The first hematologic mtDNA abnormalities were discovered by Clayton et al in 1967 using cesium chloride-ethidium density cetrifugation and electron microscopy to examine the structure of mtDNA in leukemic leukocytes. The authors identified alternate mtDNA structures comprised of circular dimers, catenated dimers, and catenated trimers. While these alternate mtDNA structures can also exist in normal cells, the authors found an unusually high percentage of them in leukemic cells as compared to normal controls [70]. In a follow-up study, the authors examined the leukocytes of 14 patients with acute and chronic granulocytic leukemias. Circular dimers were found in all 14 patients, but not in leukocytes from 3 healthy donors. It was also noted that the percentage of circular dimers decreased in some leukemic patients following chemotherapeutic treatment, suggesting that the severity of leukemia may be related to the presence of circular dimers [71]. The decrease of circular mtDNA dimers after chemotherapy also suggests a possibility that the leukemia cells containing this mtDNA abnormality may be more vulnerable to elimination by therapeutic agents.
Analysis of mtDNA from acute myelogenous leukemia (AML) cells revealed that the origins of abnormal mtDNA structures could be traced back to the bone marrow [72]. A separate study in leukemia cells from patients with acute lymphoblastic leukemia (ALL) identified mtDNA point mutations in 11 of the 30 patients [73]. Mutations of mtDNA have also been identified in patients with myelodysplastic syndrome (MDS). Gattermann et al described mutations in cytochrome b, cytochrome c oxidases I and II and ATPase 8 in MDS cells [74, 75]. The more recent study further confirmed the increased mutations in the mitochondrially-encoded COX I and COX II genes [76]. Interestingly, it has also been shown that increased mtDNA content and alterations in the expression of the mtDNA-encoded electron transport subunits may occur in the absence of mtDNA mutations. For instance, Jia et al showed that CEM/VLB100 cells expressing MDR-1 contained significantly higher mtDNA content than the parental CEM cells which lack MDR-1. In spite of this increase in mtDNA content, the authors did not find a corresponding increase in mitochondrial protein or lipid levels. However, it was noted that ATP generation was enhanced and proliferation was significantly decreased in the MDR-1 expressing cells as compared to parental CEM cells [77]. Another study examining doxorubicin-resistant, MDR-1 positive K562 cells found decreased expression of COX I and II and increased expression of COXIII. These alterations in COX expression levels were not related to mtDNA mutations or changes in transcription rates [78]. MtDNA amplification in the absence of mtDNA mutations has been reported in patients with acute myeloid leukemia [79]. Finally, while mtDNA alterations in human lymphomas have not been reported, studies in murine lymphoma showed that the expression of ND5 was increased in a metastatic variant of RAW 117-H10 large cell lymphoma. It is hypothesized that the increased expression of ND5 may play a role in tumor metastasis [80, 81].
Research perspectives and implications
Mitochondria possess their own genome which codes for 13 of the 87 proteins required for oxidative phosphorylation as well as the 12S and 16S rRNAs and 22 tRNAs necessary for protein synthesis in the mitochondria [26–28]. Mutations and alterations in mitochondrial DNA have been shown to play an important role in certain diseases such as Leber's hereditary optic neuropathy, maternally inherited diabetes melittus, and Leigh's syndrome [82]. While these diseases of the mtDNA are due to germline mutations, somatic mutations have been observed in other diseases, especially in cancer. Given the substantial role of mitochondria in ATP metabolism, in generation of free radicals, and in regulation of apoptosis, mutations in mtDNA are likely to affect cellular energy capacities, increase oxidative stress, cause ROS-mediated damage to DNA, and alter the cellular response to apoptosis induction by anticancer agents as indicated in this review and others [83, 84]. Thus, the role of mtDNA mutations in cancer development, genetic instability and disease progression, and the development of drug resistance warrants a comprehensive investigation in the future. Furthermore, the high frequency of mtDNA alterations in cancer and their presence in the early stages of disease could possibly be exploited as clinical markers for early cancer detection [85].
The existence of multiple mitochondria with many copies of mtDNA in a cell presents a unique situation and a challenge for investigating mtDNA mutations and their functional consequences. Analysis of germline homoplasmic mutations (a mutation present in all copies of mtDNA in cells from an individual) is relatively simple and can be achieved using straightforward laboratory techniques. However, it is possible that the wild-type and mutant mtDNA may co-exist in a state known as heteroplasmy. This is particularly important in somatic mutations relevant to cancer. Detection of heteroplasmic mtDNA mutations and their evolution during cancer development requires special techniques and careful consideration.
Both homoplasmic and heteroplasmic mutations have been observed in cancer cells. The mechanisms by which homoplasmy arises from heteroplasmic mutations in cancer cells still remain to be defined. A recent study by Coller et al using extensive computer modeling suggests that if a mtDNA mutation occurs in a tumor progenitor cell, homoplasmy can be achieved entirely by chance through unbiased mtDNA replication and sorting during cell division without selection for physiological advantage [86]. However, considering the fact that mtDNA lacks sizeable introns, mutations in the coding sequences have a high probability of changing the amino acid composition of the encoded proteins. Thus, many of the mtDNA mutations are likely to have physiological consequences and would confer a cellular advantage or disadvantage in growth and survival. Figure 2 schematically illustrates some possible mechanisms by which heteroplasmic mutations may emerge and evolve with various outcomes. Somatic mtDNA mutations can be induced by various DNA-damaging agents, including endogenous mitochondrial ROS and/or certain anticancer drugs and radiation (step 1). The lack of histone protection, limited repair ability, and close proximity to the electron transport chain make mtDNA far more susceptible to DNA-damaging agents. Since such induced mutation does not involve all copies of mtDNA, the initial mutation is heteroplasmic with the mutated mtDNA being the minority. If a mutation leads to a disadvantage in cell growth or survival, such mutation is deemed to diminish (step 2). Conversely, if a mutation confers cell growth/survival advantage or facilitates mtDNA replication, such a mutation is likely to survive the selection and emerge as a heteroplasmic mutation (steps 3 and 4). Depending on the duration and the degree of growth/survival advantage, the cells carrying the "advantageous" mtDNA mutation may eventually become dominant and evolve to establish a homoplasmic mutant state (step 5). A growth/survival advantage may also be acquired through a second event such as additional DNA alterations caused by ROS (step 6). It is possible that mtDNA mutations may cause a functional change of the affected protein (subunit of the electron transport complexes) in such a way that it compromises the efficiency of the electron transport chain, leading to electron flow bifurcation and increased generation of ROS (step 7). A moderate increase of ROS has been found to stimulate cellular proliferation and mitochondrial biogenesis [87, 88]. The increase in endogenous ROS thus provides a constant stimulus for cell proliferation, and most importantly, may cause further damage to both mtDNA and nDNA, leading to cancer development, genetic instability, and disease progression (step 8). Further investigations are needed to test this hypothesis. Future studies using both experimental model systems in the laboratory and primary cancer specimens from patients in a sequential manner are likely to provide critical information.

Schematic illustration of somatic mtDNA mutation. Mitochondrial DNA mutations can be induced by endogenous or exogenous DNA-damaging agents such as ROS, chemical agents, and radiation. The numbered arrows indicate possible outcomes of mtDNA mutations including heteroplasmic and homoplasmic states. See text for detail.
Cancer cells are generally more active than normal cells in metabolic ROS generation and are constantly under oxidative stress [for a review see [9]]. It is conceivable that certain mtDNA mutations may be caused by endogenous ROS in cancer cells. Mutations in mtDNA could in turn cause further increases of ROS production by the mechanism discussed above, leading to more mutations and additional oxidative stress. Studies in our laboratory have demonstrated that primary leukemia cells from patients with chronic lymphocytic leukemia contain significantly higher levels of cellular ROS compared to normal lymphocytes [89]. The ROS levels are especially high in CLL cells from patients who have had prior chemotherapy. Chemotherapeutic agents have been shown to accumulate in mitochondria and damage mtDNA [90, 91]. It is possible that some of the mtDNA mutations identified in cancer cells are the result of treatment with chemotherapeutic agents. Such somatic mutations would likely emerge as heteroplasmic mutations and might eventually become homoplasmic with time due to clonal expansion if they confer replication and survival advantages. Analysis of mtDNA mutations in these leukemia samples and evaluation of their correlation with cellular ROS contents are important to test this possibility.
Given the critical role of mitochondria in apoptosis, it is conceivable that mutations in mtDNA in cancer cells could significantly affect the cellular apoptotic response to anticancer agents. As discussed above, experiments with ρ0 cells lacking mtDNA to evaluate the role of respiration in drug sensitivity produced various results. This may reflect complex interactions between ρ0 cells and anticancer agents with different mechanisms of action. Evaluating changes in drug sensitivity in cancer cells bearing mtDNA mutations would be even more complicated, since different mtDNA mutations are likely to have varying effects on the apoptotic response. However, the clonal selection/expansion hypothesis would predict that the mutations recovered from cancer cells that survive chemotherapy are likely to be associated with resistance to the particular drugs used in prior therapy. Furthermore, certain mtDNA mutations in cancer cells are likely to cause malfunctions of the respiratory chain and increase free radical generation. This biochemical change provides an unique opportunity to selectively kill this population of cancer cells by using agents that inhibit free radical elimination and cause further ROS accumulation, leading to lethal damage in the cancer cells [92]. Taken together, it is evident that mtDNA mutations are clinically relevant and have potential therapeutic implications.
Conclusion
In summary, the presence of mtDNA mutations in cancer cells is consistent with the intrinsic susceptibility of mtDNA to damage and constitutive oxidative stress. Alterations in respiratory activity and mtDNA abnormalities appear to be a general feature of malignant cells. Mutations and deletions in mtDNA and abnormal expression of mtDNA-encoded proteins have been observed in various solid tumors and hematological malignancies. However, studies to date have focused on identification and characterization of the altered mtDNA, with only limited insight into functional consequences and clinical relevance. Future research to address the functional aspects of mtDNA mutations in cancer development and therapeutic response is likely to be fruitful and have significant clinical implications in the prevention and treatment of cancer. Important questions to be addressed include (1) What is the exact role of mtDNA in cancer initiation and progression? (2) How do heteroplasmic mtDNA mutations arise and evolve to a homoplasmic state in cancer cells? (3) Is there a mechanistic link between mtDNA mutations, changes in respiration and ROS generation, and alterations in apoptosis? (4) What are the most important mtDNA mutations that confer drug resistance and growth/metastasis advantage in clinically relevant cancer cells? (5) How can we effectively kill those cancer cells that carry mtDNA mutations and become resistant to conventional anticancer agents? The answers to these questions would undoubtedly advance our understanding of mitochondrial biology in cancer, as well as provide a basis for designing new strategies for effective cancer prevention, diagnosis, and treatment.
References
Warburg O: The metabolism of tumors. London Constable Co. Ltd. 1930
Warburg O: On the origin of cancer cells. Science. 1956, 123: 309-314.
Hockenbery DM: A mitochondrial achilles heel in cancer?. Cancer Cell. 2002, 2: 1-2. 10.1016/S1535-6108(02)00087-9
Schwartz RM, Dayhoff MO: Origins of prokaryotes, eukaryotes, mitochondria, and chloroplasts. Science. 1978, 199: 395-403.
Whatley JM, John P, Whatley ER: From extracellular to intracellular: the establishment of mitochondria and chloroplasts. Proc R Soc Lond B Biol Sci. 1979, 204: 165-87.
Gray MW, Doolittle WF: Has the endosymbiont hypothesis been proven?. Microbiol Rev. 1982, 46: 1-42.
Saybasili H, Yuksel M, Haklar G, Yalcin AS: Effect of mitochondrial electron transport inhibitors on superoxide radical generation in rat hippocampal and striatal slices. Antioxid Redox Signal. 2001, 3: 1099-1104. 10.1089/152308601317203602
Staniek K, Gille L, Kozlov AV, Nohl H: Mitochondrial superoxide radical formation is controlled by electron bifurcation to the high and low potential pathways. Free Radic Res. 2002, 36: 381-387. 10.1080/10715760290021225
Hileman EO, Achanta G, Huang P: Superoxide dismutase: an emerging target for cancer therapeutics. Expert Opin Ther Targets. 2001, 5: 697-710.
Krebs HA: The history of the tricarboxylic acid cycle. Perspect Biol Med. 1970, 14: 154-170.
Kornburg HL: Tricarboxylic acid cycles. Bioessays. 1987, 7: 236-238.
Watford M: The urea cycle: a two-compartment system. Essays Biochem. 1991, 26: 49-58.
King MP, Attardi G: Human cells lacking mtDNA: repopulation with exogenous mitochondria by complementation. Science. 1989, 246: 500-503.
Wang X: The expanding role of mitochondria in apoptosis. Genes Dev. 2001, 15: 2922-2933.
Liu X, Kim CN, Yang J, Jemmerson R, Wang X: Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell. 1996, 86: 147-157.
Susin SA, Lorenzo HK, Zamzami N, Marzo I, Snow BE, Brothers GM, Mangion J, Jacotot E, Costantini P, Loeffler M, Larochette N, Goodlett DR, Aebersold R, Siderovski DP, Penninger JM, Kroemer G: Molecular characterization of mitochondrial apoptosis-inducing factor. Nature. 1999, 397: 441-446. 10.1038/17135
Singh KK, Russell J, Sigala B, Zhang T, Williams J, Keshav KF: Mitochondrial DNA determines the cellular response to cancer therapeutic agents. Oncogene. 1999, 18: 6641-6646. 10.1038/sj.onc.1203056
Cai J, Wallace DC, Zhivotovsky B, Jones DP: Separation of cytochrome c-dependent caspase activation from thiol-disulfide redox change in cells lacking mitochondrial DNA. Free Radic Biol Med. 2000, 29: 334-42. 10.1016/S0891-5849(00)00312-9
Joshi B, Li L, Taffe BG, Zhu Z, Wahl S, Tian H, Ben-Josef E, Taylor JD, Porter AT, Tang DG: Apoptosis induction by a novel anti-prostate cancer compound, BMD188 (a fatty acid-containing hydroxamic acid), requires the mitochondrial respiratory chain. Cancer Res. 1999, 59: 4343-4355.
Hayashi JI, Takemitsu M, Nonaka M: Recovery of the missing tumorigenicity in mitochondrial DNA-less HeLa cells by introduction of mitochondrial DNA from normal human cells. Somatic Cell Mol Genet. 1992, 18: 123-129.
Morais R, Zinkewich P, Parent M, Wang H, Babai F, Zollinger M: Tumor-forming ability in athymic nude mice of human cell lines devoid of mitochondrial DNA. Cancer Res. 1994, 54: 3889-3996.
Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, Young IG: Sequence and organization of the human mitochondrial genome. Nature. 1981, 290: 457-465.
Grivell LA: Mitochondrial DNA. Sci Am. 1983, 248: 78-89.
Blanchard JL, Schmidt GW: Mitochondrial DNA migration events in yeast and humans: integration by a common end-joining mechanism and alternative perspectives on nucleotide substitution patterns. Mol Biol Evol. 1996, 13: 537-548.
Zischler H: Nuclear integrations of mitochondrial DNA in primates: inference of associated mutational events. Electrophoresis. 2000, 21: 531-536. 10.1002/(SICI)1522-2683(20000201)21:3<531::AID-ELPS531>3.3.CO;2-G
Wallace DC: Diseases of the mitochondrial DNA. Annu Rev Biochem. 1992, 61: 1175-1212. 10.1146/annurev.bi.61.070192.005523
Wallace DC: Mitochondrial diseases in man and mouse. Science. 1999, 283: 1482-1488. 10.1126/science.283.5407.1482
Singh KK, : Mitochondrial DNA Mutations in Aging, Disease, and Cancer. Springer New York NY. 1998
Zastawny TH, Dabrowska M, Jaskolski T, Klimarczyk M, Kulinski L, Koszela A, Szczesniewicz M, Sliwinska M, Witkowski P, Olinski R: Comparison of oxidative base damage in mitochondrial and nuclear DNA. Free Rad Biol Med. 1998, 24: 722-725. 10.1016/S0891-5849(97)00331-6
Belliard G, Vadel F, Pelletier G: Mitochondrial recombination in cytoplasmic hybrids of Nicotina tabacum by protoplast fusion. Nature. 1979, 281: 401-403.
Dujon B, Slonimski PP, Weill L: Mitochondrial genetics IX: a model for recombination and segregation of mitochondrial genomes in Saccharomyces cerevisiae. Genetics. 1974, 78: 415-437.
Ono T, Isobe K, Nakada K, Hayashi JI: Human cells are protected from mitochondrial dysfunction by complementation of DNA products in fused mitochondria. Nat Genet. 2001, 28: 272-275. 10.1038/90116
Tan DJ, Bai RK, Wong LJ: Comprehensive scanning of somatic mitochondrial DNA mutations in breast cancer. Cancer Res. 2002, 62: 972-976.
Parrella P, Xiao Y, Fliss M, Sanchez-Cespedes M, Mazzarelli P, Rinaldi M, Nicol T, Gabrielson E, Cuomo C, Cohen D, Pandit S, Spencer M, C Rabitti, Fazio VM, Sidransky D: Detection of mitochondrial DNA mutations in primary breast cancer and fine-needle aspirates. Cancer Res. 2001, 61: 7623-7626.
Bianchi MS, Bianchi NO, Bailliet G: Mitochondrial DNA mutations in normal and tumor tissues from breast cancer patients. Cytogenet Cell Genet. 1995, 71: 99-103.
Sharp MG, Adams SM, Walker RA, Brammar WJ, Varley JM: Differential expression of the mitochondrial gene cytochrome oxidase II in benign and malignant breast tissue. J Pathol. 1992, 168: 163-168.
Liu VW, Shi HH, Cheung AN, Chiu PM, Leung TW, Nagley P, Wong LC, Ngan HY: High incidence of somatic mitochondrial DNA mutations in human ovarian carcinomas. Cancer Res. 2001, 61: 5998-6001.
Polyak K, Li Y, Zhu H, Lengauer C, Willson JK, Markowitz SD, Trush MA, Kinzler KW, Vogelstein B: Somatic mutations of the mitochondrial genome in human colorectal tumours. Nat Genet. 1998, 20: 291-293. 10.1038/3108
Alonso A, Martin P, Albarran C, Aquilera B, Garcia O, Guzman A, Oliva H, Sancho M: Detection of somatic mutations in the mitochondrial DNA control region of colorectal and gastric tumors by heteroduplex and single-strand conformation analysis. Electrophoresis. 1997, 18: 682-685.
Chester KA, Robson L, Begent RH, Pringle H, Primrose L, Talbot IC, Macpherson AJ, Owen SL, Boxer G, Malcolm AD: In situ and slot hybridization analysis of RNA in colorectal tumours and normal colon shows distinct distributions of mitochondrial sequences. J Pathol. 1990, 162: 309-315.
Yamamoto A, Horai S, Yuasa Y: Increased level of mitochondrial gene expression in polyps of familial polyposis coli patients. Biochem Biophys Res Commun. 1989, 159: 1100-1106.
Lu X, Walker T, MacManus JP, Seligy VL: Differentiation of HT-29 human colonic adenocarcinoma cells correlates with increased expression of mitochondrial RNA: effects of trehalose on cell growth and maturation. Cancer Res. 1992, 52: 3718-3725.
Savre-Train I, Piatyszek MA, Shay JW: Transcription of deleted mitochondrial DNA in human colon adenocarcinoma cells. Hum Mol Genet. 1992, 1: 203-204.
Habano W, Nakamura S, Sugai T: Microsatellite instability in the mitochondrial DNA of colorectal carcinomas: evidence for mismatch repair systems in mitochondrial genome. Oncogene. 1998, 17: 1931-1937. 10.1038/sj.onc.1202112
Habano W, Sugai T, Yoshida T, Nakamura S: Mitochondrial gene mutation, but not large-scale deletion, is a feature of colorectal carcinomas with mitochondrial microsatellite instability. Int J Cancer. 1999, 83: 625-629. 10.1002/(SICI)1097-0215(19991126)83:5<625::AID-IJC10>3.3.CO;2-E
Maximo V, Soares P, Seruca R, Rocha AS, Castro P, Sobrinho-Simoes M: Microsatellite instability, mitochondrial DNA large deletions, and mitochondrial DNA mutations in gastric carcinoma. Genes Chromosomes Cancer. 2001, 32: 136-143. 10.1002/gcc.1175
Tamura G, Nishizuka S, Maesawa C, Suzuki Y, Iwaya T, Sakata K, Endoh Y, Motoyama T: Mutations in mitochondrial control region DNA in gastric tumours of Japanese patients. Eur J Cancer. 1999, 35: 316-319. 10.1016/S0959-8049(98)00360-8
Burgart LJ, Zheng J, Shu Q, Strickler JG, Shibata D: Somatic mitochondrial mutation in gastric cancer. Am J Pathol. 1995, 147: 1105-1111.
Nomoto S, Yamashita K, Koshikawa K, Nakao A, Sidransky D: Mitochondrial D-loop mutations as clonal markers in multicentric hepatocellular carcinoma and plasma. Clin Cancer Res. 2002, 8: 481-487.
Nishikawa M, Nishiguchi S, Shiomi S, Tamori A, Koh N, Takeda T, Kubo S, Hirohashi K, Kinoshita H, Sato E, Inoue M: Somatic mutation of mitochondrial DNA in cancerous and noncancerous liver tissue in individuals with hepatocellular carcinoma. Cancer Res. 2001, 61: 1843-1845.
Yamamoto H, Tanaka M, Katayama M, Obayashi T, Nimura Y, Ozawa T: Significant existence of deleted mitochondrial DNA in cirrhotic liver surrounding hepatic tumor. Biochem Biophys Res Commun. 1992, 182: 913-920.
Miyazono F, Schneider PM, Metzger R, Warnecke-Eberz U, Baldus SE, Dienes HP, Aikou T, Hoelscher AH: Mutations in the mitochondrial DNA D-Loop region occur frequently in adenocarcinoma in Barrett's esophagus. Oncogene. 2002, 21: 3780-3783. 10.1038/sj.onc.1205532
Hibi K, Nakayama H, Yamazaki T, Takase T, Taguchi M, Kasai Y, Ito K, Akiyama S, Nakao A: Mitochondrial DNA alteration in esophageal cancer. Int J Cancer. 2001, 92: 319-321. 10.1002/ijc.1204
Jones JB, Song JJ, Hempen PM, Parmigiani G, Hruban RH, Kern SE: Detection of mitochondrial DNA mutations in pancreatic cancer offers a "mass"-ive advantage over detection of nuclear DNA mutations. Cancer Res. 2001, 61: 1299-1304.
Horton TM, Petros JA, Heddi A, Shoffner J, Kaufman AE, Graham SD, Gramlich T, Wallace DC: Novel mitochondrial DNA deletion found in a renal cell carcinoma. Genes Chromosomes Cancer. 1996, 15: 95-101. 10.1002/(SICI)1098-2264(199602)15:2<95::AID-GCC3>3.0.CO;2-Z
Selvanayagam P, Rajaraman S: Detection of mitochondrial genome depletion by a novel cDNA in renal cell carcinoma. Lab Invest. 1996, 74: 592-599.
Faure VH, Heddi A, Giraud S, Chautard D, Stepien G: Expression of oxidative phosphorylation genes in renal tumors and tumoral cell lines. Mol Carcinog. 1996, 16: 165-172. 10.1002/(SICI)1098-2744(199607)16:3<165::AID-MC7>3.0.CO;2-G
Simonnet H, Alazard N, Pfeiffer K, Gallou C, Beroud C, Demont J, Bouvier R, Schagger H, Godinot C: Low mitochondrial respiratory chain content correlates with tumor aggressiveness in renal cell carcinoma. Carcinogenesis. 2002, 23: 759-768. 10.1093/carcin/23.5.759
Jessie BC, Sun CQ, Irons HR, Marshall FF, Wallace DC, Petros JA: Accumulation of mitochondrial DNA deletions in the malignant prostate of patients of different ages. Exp Gerontol. 2001, 37: 169-174. 10.1016/S0531-5565(01)00153-X
Jeronimo C, Nomoto S, Caballero OL, Usadel H, Henrique R, Varzim G, Oliveira J, Lopes C, Fliss MS, Sidransky D: Mitochondrial mutations in early stage prostate cancer and bodily fluids. Oncogene. 2001, 20: 5195-5198. 10.1038/sj.onc.1204646
Liang BC, Hays L: Mitochondrial DNA copy number changes in human gliomas. Cancer Lett. 1996, 105: 167-173. 10.1016/0304-3835(96)04276-0
Liang BC: Evidence for association of mitochondrial DNA sequence amplification and nuclear localization in human low-grade gliomas. Mutat Res. 1996, 354: 27-33. 10.1016/0027-5107(96)00004-8
Reid RA: Can migratory DNA activate oncogenes. Trends Biochem Sci. 1983, 8: 190-191. 10.1016/0968-0004(83)90205-0. 10.1016/0968-0004(83)90205-0
Kirches E, Michael M, Woy C, Schneider T, Warich M-Kirches, Schneider-Stock R, Winkler K, Wittig H, Dietzmann K: Loss of heteroplasmy in the displacement loop of brain mitochondrial DNA in astrocytic tumors. Genes Chromosomes Cancer. 1999, 26: 80-83. 10.1002/(SICI)1098-2264(199909)26:1<80::AID-GCC11>3.0.CO;2-4
Stefaneanu L, Tasca C: An electron-microscopic study of human thyroid cancer. Endocrinologie. 1979, 17: 233-239.
Yeh JJ, Lunetta KL, van Orsouw NJ, Moore FD, Mutter GL, Vijg J, Dahia PL, Eng C: Somatic mitochondrial DNA (mtDNA) mutations in papillary thyroid carcinomas and differential mtDNA sequence variants in cases with thyroid tumours. Oncogene. 2000, 19: 2060-2066. 10.1038/sj.onc.1203537
Maximo V, Soares P, Lima J, Cameselle-Teijeiro J, Sobrinho-Simoes M: Mitochondrial DNA somatic mutations (point mutations and large deletions) and mitochondrial DNA variants in human thyroid pathology: a study with emphasis on Hurthle cell tumors. Am J Pathol. 2002, 160: 1857-1865.
Fliss MS, Usadel H, Caballero OL, Wu L, Buta MR, Eleff SM, Jen J, Sidransky D: Facile detection of mitochondrial DNA mutations in tumors and bodily fluids. Science. 2000, 287: 2017-2019. 10.1126/science.287.5460.2017
Sanchez-Cespedes M, Parrella P, Nomoto S, Cohen D, Xiao Y, Esteller M, C Jeronimo, Jordan RC, Nicol T, Koch WM, Schoenberg M, Mazzarelli P, Fazio VM, Sidransky D: Identification of a mononucleotide repeat as a major target for mitochondrial DNA alterations in human tumors. Cancer Res. 2001, 61: 7015-7019.
Clayton DA, Vinograd J: Circular dimer and catenate forms of mitochondrial DNA in human leukaemic leucocytes. Nature. 1967, 216: 652-657.
Clayton DA, Vinograd J: Complex mitochondrial DNA in leukemic and normal human myeloid cells. Proc Natl Acad Sci U S A. 1969, 62: 1077-1084.
Robberson DL, Gay ML, Wilkins CE: Genetically altered human mitochondrial DNA and a cytoplasmic view of malignant transformation. in Genes, chromosomes, and neoplasia. Edited by: Arrighi FE, Rao PN, Stubblefield E. 1981, 125-156.
Ivanova R, Lepage V, Loste MN, Schachter F, Wijnen E, Busson M, Cayuela JM, Sigaux F, Charron D: Mitochondrial DNA sequence variation in human leukemic cells. Int J Cancer. 1998, 76: 495-498. 10.1002/(SICI)1097-0215(19980518)76:4<495::AID-IJC9>3.3.CO;2-3
Gattermann N: From sideroblastic anemia to the role of mitochondrial DNA mutations in myelodysplastic syndromes. Leuk Res. 2000, 24: 141-151. 10.1016/S0145-2126(99)00160-5
Gattermann N, Retzlaff S, Wang YL, Hofhaus G, Heinisch J, Aul C, Schneider W: Heteroplasmic point mutations of mitochondrial DNA affecting subunit I of cytochrome c oxidase in two patients with acquired idiopathic sideroblastic anemia. Blood. 1997, 90: 4961-4972.
Reddy PL, Shetty VT, Dutt D, York A, Dar S, Mundle SD, Allampallam K, Alvi S, Galili N, Saberwal GS, Anthwal S, Shaikh M, Suleman S, Kamal SY, Raza A: Increased incidence of mitochondrial cytochrome c-oxidase gene mutations in patients with myelodysplastic syndromes. Br J Haematol. 2002, 116: 564-575. 10.1046/j.0007-1048.2001.03323.x
Jia L, Liu KZ, Newland AC, Mantsch HH, Kelsey SM: Pgp-positive leukaemic cells have increased mtDNA but no increased rate of proliferation. Br J Haematol. 1999, 107: 861-869. 10.1046/j.1365-2141.1999.01771.x
Grandjean F, Bremaud L, Robert J, Ratinaud MH: Alterations in the expression of cytochrome c oxidase subunits in doxorubicin-resistant leukemia K562 cells. Biochem Pharmacol. 2002, 63: 823-831. 10.1016/S0006-2952(01)00865-6
Boultwood J, Fidler C, Mills KI, Frodsham PM, Kusec R, Gaiger A, Gale RE, Linch DC, Littlewood TJ, Moss PA, Wainscoat JS: Amplification of mitochondrial DNA in acute myeloid leukaemia. Br J Haematol. 1996, 95: 426-431. 10.1046/j.1365-2141.1996.d01-1922.x
LaBiche RA, Yoshida M, Gallick GE, Irimura T, Robberson DL, Klostergaard J, Nicolson GL: Gene expression and tumor cell escape from host effector mechanisms in murine large cell lymphoma. J Cell Biochem. 1988, 36: 393-403.
LaBiche RA, Demars M, Nicolson GL: Transcripts of the mitochondrial gene ND5 are overexpressed in highly metastatic murine large cell lymphoma cells. In Vivo. 1992, 6: 317-324.
DiMauro S, Schon EA: Mitochondrial DNA mutations in human disease. Am J Med Genet. 2001, 106: 18-26. 10.1002/ajmg.1392
Penta JS, Johnson FM, Wachsman JT, Copeland WC: Mitochondrial DNA in human malignancy. Mutat Res. 2001, 488: 119-133. 10.1016/S1383-5742(01)00053-9
Copeland WC, Wachsman JT, Johnson FM, Penta JS: Mitochondrial DNA alterations in cancer. Cancer Invest. 2002, 20: 557-569. 10.1081/CNV-120002155
Modica-Napolitano JS, Singh KK: Mitochondria as targets for detection and treatment of cancer. Exp Rev Mol Med. 2002, 11 April
Coller HA, Khrapko K, Bodyak ND, Nekhaeva E, Herrero-Jimenez P, Thilly WG: High frequency of homoplasmic mitochondrial DNA mutations in human tumors can be explained without selection. Nat Genet. 2001, 28: 147-150. 10.1038/88859
Lee HC, Yin PH, Chi CW, Wei YH: Increase in mitochondrial mass in human fibroblasts under oxidative stress and during replicative cell senescence. J Biomed Sci. 2002, 9: 517-526. 10.1159/000064724
Liu SL, Lin X, Shi DY, Cheng J, Wu CQ, Zhang YD: Reactive oxygen species stimulated human hepatoma cell proliferation via cross-talk between PI3-K/PKB and JNK signaling pathways. Arch Biochem Biophys. 2002, 406: 173-182. 10.1016/S0003-9861(02)00430-7
Zhou Y, Hileman EO, Keating MJ, Huang P: Correlation between superoxide generation and cellular response to the anticancer agent 2-methoxyestradiol in chronic lymphocytic leukemia cells. Proc Am Ass Cancer Res. 2002, 43: 798-
Talarico T, Cullinane CM, Gray PJ, Webster LK, Deacon GB, Phillips DR: Nuclear and mitochondrial distribution of organoamidoplatinum(II) lesions in cisplatin-sensitive and -resistant adenocarcinoma cells. Anticancer Drug Des. 2001, 16: 135-141.
Olivero OA, Chang PK, Lopez DM-Larraza, Semino MC-Mora, MC Poirier: Preferential formation and decreased removal of cisplatin-DNA adducts in Chinese hamster ovary cell mitochondrial DNA as compared to nuclear DNA. Mutat Res. 1997, 391: 79-86. 10.1016/S0165-1218(97)00037-2
Huang P, Feng L, Oldham EA, Keating MJ, Plunkett W: Superoxide dismutase as a target for the selective killing of cancer cells. Nature. 2000, 407: 390-395. 10.1038/35030140
Acknowledgments
This work was supported in part by grants CA77339 and CA85563 from the National Cancer Institute, National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
JSC conducted literature review and wrote this article. PH provided suggestions and finalized the manuscript. All authors read and approved the final manuscript.
Electronic supplementary material
12943_2002_9_MOESM1_ESM.xls
Additional File 1: Carew & Huang Table1.xls, Microsoft Excel table providing a summary of mutations found in the non-coding region and coding regions of the mtDNA in various types of human cancer cells (XLS 20 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Carew, J.S., Huang, P. Mitochondrial defects in cancer. Mol Cancer 1, 9 (2002). https://doi.org/10.1186/1476-4598-1-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-4598-1-9