
Abstract
Background
Highland areas with naturally less intense malaria transmission may provide models of how lowland areas might become if transmission was permanently reduced by sustained vector control. It has been argued that vector control should not be attempted in areas of intense transmission.
Methods
Mosquitoes were sampled with light traps, pyrethrum spray and window exit traps. They were tested by ELISA for sporozoites. Incidence of malaria infection was measured by clearing existing infections from children with chlorproguanil-dapsone and then taking weekly blood samples. Prevalence of malaria infection and fever, anaemia and splenomegaly were measured in children of different age groups. All these measurements were made in highland and lowland areas of Tanzania before and after provision of bednets treated with alphacypermethrin.
Results
Entomological inoculation rates (EIR) were about 17 times greater in a lowland than a highland area, but incidence of infection only differed by about 2.5 times. Malaria morbidity was significantly less prevalent in the highlands than the lowlands. Treated nets in the highlands and lowlands led to 69–75% reduction in EIR. Malaria morbidity showed significant decline in younger children at both altitudes after introduction of treated nets. In children aged 6–12 the decline was only significant in the highlands
Conclusions
There was no evidence that the health benefits to young children due to the nets in the lowlands were "paid for" by poorer health later in life. Our data support the idea of universal provision of treated nets, not a focus on areas of natural hypo-endemicity.
Similar content being viewed by others

Background
The policy of the Roll Back Malaria programme of WHO has put strong emphasis on the introduction of insecticide treated nets throughout Africa [1]. However, this is contrary to the view common until quite recently, and is still held by some [2, 3], that any attempts at vector control should be focused on hypo-endemic areas e.g. in highlands. It is suggested that, in areas of stable malaria with intense transmission in lowland Africa, attempts to control malaria by intervention against vectors would be futile, or even counter-productive. It is considered that, in such areas, reducing transmission would interfere with normal build-up of immunity so that early reduction in morbidity would be "paid for" by postponement of morbidity until a later age, so that the lifetime burden of malaria would not decline and might actually increase. Furthermore, reduction in transmission might convert stable malaria to epidemic malaria, which might exact a higher death toll.
The existing data on insecticide-treated nets are mostly based on short-term trials [4], but Maxwell et al [5] have reported encouraging results in eight Tanzanian villages which have had community-wide use of nets (re-treated annually) for up to 4 years. Another approach to settling the issue of the effects of long-term exposure to either intense, or more moderate, transmission is to study malaria morbidity in different age groups in nearby areas which differ naturally and semi-permanently in vector density, due to a difference in altitude. Ellman et al [6] compared malaria morbidity in the East Usambara mountains of Tanzania with that in culturally similar people in the nearby lowlands, which have much greater densities of vectors. They found markedly less morbidity in the highlands and a clear decline of prevalence of morbidity with age at both altitudes. However, children were only studied up to 5 years of age which, it could be argued, was not enough to completely exclude the delay of morbidity into early adolescence, reported by Trape and Rogier [2].
This paper reports a study, in the same highland and lowland areas where Ellman et al [6] worked, which compared the areas by entomological measures of transmission, incidence of Plasmodium falciparum infection after clearing existing infections and malaria morbidity up to the age of 12 years. It also reports on the impact of introducing insecticide-treated nets in both areas to determine whether better results would be obtained by focusing anti-vector measures on areas of lower initial rates of malaria transmission.
Clearance for the studies was granted by the ethical committees of the Tanzanian National Institute for Medical Research and the London School of Hygiene & Tropical Medicine.
Methods
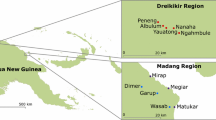
Twenty one hamlets (vitongoji in Ki-Swahili, i.e. sub-communities of politically defined villages (vijiji)) were included in the study which fell into distinct categories of 11 highland hamlets (altitudes 784 to 1148 metres) and 10 lowland hamlets (199 to 300 metres). In the first (baseline) year, the vector population, incidence of infections and measures of morbidity were studied in the two areas without any vector control. In the second year, bednets treated with 20 mg alphacypermethrin/m2 of netting were provided free of charge to protect all beds and sleeping mats in five of the highland and five of the lowland hamlets. In some hamlets the aqueous mix of insecticide was provided by appropriate dilution of Fendona SC and, in others, of granular Fastac. Bioassays were carried out on treated nets by wrapping them round wire frames and inserting wild collected Anopheles funestus inside and observing either (a) % mortality 24 hrs after a 3 minute exposure, or (b) the median time for knock-down with continuous exposure [7]. The nets were not re-treated until 15 months after the first treatment, i.e. after the end of the observations on the vector populations reported in this paper.
Mosquito populations were monitored in each hamlet by CDC light traps set beside occupied, untreated bednets [8] monthly in each of eight "sentinel" bedrooms per hamlet. The anophelines caught by these indoor traps were found by morphological examination to consist almost entirely of Anopheles gambiae s.l., An. funestus and Anopheles marshallii s.l., all of which have been incriminated as vectors in this area [9]. The heads and thoraces of these species were tested by ELISA for P. falciparum circumsporozoite protein (CSP) [10]. In the second year of the study, resting mosquitoes were collected by pyrethrum spray catch monthly in five bedrooms per lowland hamlet and in window exit-traps on these bedrooms. In the netted hamlets these collections were made in bedrooms with treated bednets. Anophelines were classified as bloodfed (or semi-gravid) or unfed and the total numbers and the numbers of bloodfed individuals were compared in the netted and untreated villages. After adjusting for the differences in the hamlet populations of anophelines found by the light traps, the ratio of the numbers of fed mosquitoes was used as a measure of the protection from biting conferred to the individual net users by the treated nets.
In the wet season (April-June) of the baseline year, incidence of infection was tested in four lowland and four highland hamlets after clearing existing infections with chlorproguanil-dapsone (lap-dap), as in several previous studies [11–13]. Thick bloodfilms were taken from 60 children per hamlet each week for eight weeks after lap-dap treatment. The results were expressed as numbers converting from parasite negativity to positivity per child week at risk.
Morbidity was monitored monthly by requesting the attendance of 50 children from each hamlet at monitoring sessions. The children's names for each session were picked at random from our census lists, after excluding those who had attended in the previous month. The following parameters were measured on each child attending: (a) malaria parasite presence and density per 200 w.b.c. in thick blood films, (b) core body temperature as measured with a Thermoscan thermometer placed first in one ear and then the other, (c) haemoglobin density measured with a Hemocue machine, (d) weight and (e) presence and grade of splenomegaly on the Hackett scale, as assessed by palpation by an experienced medical aide. The age of each child was determined, where available, from the hospital birth registration or MCH card or, if it was not available, by questioning the parent or guardian. They were also questioned about occurrence of fever episodes in the child in the previous two days.
Children found positive for infection were given anti-malarial treatment according to Tanzanian guidelines.
Results and Discussion
Entomological data
The granular Fastac formulation of alpahcypermethrin was found preferable to the concentrated Fendona SC liquid for use in the field because, in the event of spillage on to the skin, the granules could be easily brushed off but the concentrated liquid needs to be carefully washed off. With both formulations, bioassays involving 3 minutes exposure and 24 hrs holding yielded mortalities very close to 100% even after 15 months domestic use and about 3 washes (Table 1). The mean median times for knockdown 15 months after net treatment suggested that the nets treated with Fendona had higher insecticidal activity than those treated once with Fastac and that re-treatment boosted insecticidal activity of the latter. However, the 95% confidence limits of the means overlapped, so no definite conclusion can be drawn about apparent differences in insecticidal activity.
Table 2 shows the data from CDC light trap catches in rooms with untreated bednets in the four quarters of each year of the study. Data are also presented on the proportion of mosquitoes found CSP positive; there were few mosquitoes for testing in the dry seasons and no obvious seasonal trend in CSP rates, so the CSP data are presented for each whole year. The hamlets are grouped into those assigned in the second year in the highlands and in the lowlands to receive treated bednets and those assigned not to receive them. The following points are evident:-
-
Anopheline densities were more than 12 times greater in the lowlands than the highlands.
-
Anopheline densities peaked in April-June, the main rainy season.
-
An. marshallii s.l. formed a considerable proportion of the total indoor anopheline populations in the highlands, especially between July and December; in the same months An. funestus was the majority species at both altitudes; An. gambiae s.l. dominated the anopheline populations in the main rainy season, when anopheline populations were largest, especially in the lowlands. The sibling species of the An. gambiae s.l. were not identifed during this study as they are generally found to be all An. gambiae s.s.[14] in this area except for some An. arabiensis which were found after very heavy El Nino associated rainfall [9].
-
The CSP (sporozoite) rate appeared to be slightly (but not significantly) greater in the lowlands than the highlands.
-
In the first year the means of the population densities and CSP rates, and hence the EIR values, for each of the groups of hamlets in the highlands and the lowlands showed that these groups were comparable.
-
In the second year, anopheline population densities were less at each altitude even where no nets were introduced, presumably for climatic reasons; the CSP rates did not change in these groups of villages without nets in the second year.
-
Where treated nets were introduced, the anopheline densities showed significant reductions compared with the untreated contemporary controls in 6 out of 8 of the comparisons at the same season and altitude. In the other 2 cases the differences were in the expected direction but were not significant.
-
The apparent impact of the SC formulation of alphacypermethrin was greater than that of the Fastac formulation in 5 out of 8 of the comparisons at the same season and altitude. In the other 3 cases the difference was in favour of Fastac and thus no convincing difference between the impact of the two formulations was demonstrated.
-
The CSP rates appeared to be reduced due to the introduction of nets, but the effect did not reach statistical significance with the data pooled over all seasons as shown in Table 2. However, if account was taken of possible heterogeneity in CSP rate by altitude and season and a Mantel Haenszel χ2 was applied to the CSP rates stratified by altitude and season, the CSP rates were found significantly less in hamlets with nets than in those without them [16].
-
Converting the trap catches to estimates of bites per person [8] and multiplying by the CSP rates gave estimates of the entomological inoculation rate per year; these differed about 17 fold between lowland and highland and showed 69–75% reduction as a result of the "mass" (or community) effect on the vector populations due to introducing treated nets at each altitude.
Table 3 shows the much reduced numbers of blood-fed anophelines in, and exiting from, lowland bedrooms with treated nets compared with corresponding catches in lowland villages with no nets. We must draw attention to two sources of error in these data. One is that the exit trap did not cover all possible mosquito escape routes and, if we missed many of the exiting mosquitoes, that would have exaggerated the difference in feeding rates, with or without treated nets since, as shown in Table 3, there was a larger difference in the feeding rate in those resting indoors than in those exiting. On the other hand, some of the fed mosquitoes may have entered to rest after feeding outside the house and this would have led to underestimation of the protection provided by the nets. The latter source of error could be avoided by molecular matching [17] of all mosquito blood meals with the blood of the sleepers under the nets in the same houses. We have not had the resources to undertake this yet. Meanwhile, we note that these two types of error would have tended to cancel out and we consider that further analysis of these data is worthwhile. The number of blood-fed mosquitoes was about 89% less (0.455 versus 4.15) in the treated rooms compared with the untreated rooms in untreated hamlets. Part of this difference is undoubtedly because of the differences in mosquito population densities of the treated and untreated hamlets due to the mass or community effect. This effect was corrected for by dividing by the contemporary light trap catches (Table 2) which, in all hamlets, were in rooms with untreated nets. The results lead to an estimate that the personal protection effect of the treated nets, on its own, gives a 73.8% reduction in biting. The scale of this effect was very similar to the 69–75% reduction in EIR due to the mass effect as demonstrated in Table 2. We conclude from this that applying treated nets in such a way as to exclude a mass effect, would reduce the total benefit to the individual net user by about half, compared to the full potential of treated nets when there is comprehensive coverage, as in the present trial. The relative importance of the mass, compared with the personal protection, effect was less than in two of our previous studies (data of both tabulated in [5]). However, all three studies demonstrated the need for a distribution system which ensures a high percentage coverage with effectively treated nets so as to maximise mosquito killing in the community.
Incidence of infection
Table 4 shows data on recurrence of infection of children in four lowland and four highland hamlets after clearing their existing infections with lap-dap. The observed mean rate of recurrence of infection per child week at risk was more than twice as great in the lowlands as the highlands. The hamlet with the highest recurrence rate in the highlands (M11) had a lower rate (0.327) than the hamlet (B8) with the lowest rate (0.385) in the lowlands. A significance test on the rates for the four highland versus the four lowland hamlets gave t (6 d.f.) = 4.53, P < 0.01.
Although the significance and direction of difference between the two altitudes was in the direction expected from the entomological data (Table 2), the scale of the difference in rate of recurrence of infection appeared smaller than would have been expected. However, a proper comparison requires that account be taken of the fact that there is occasional recrudescence of infection after lap-dap treatment. The best estimate for the rate of recrudescence, based on treated lowland children taken to stay at 1700 m where there was no transmission, was 0.013 per child week at risk [13]. This estimate did not differ significantly from one made by the more commonly used, but possibly less reliable, method of genotyping infections before and after drug treatment [18].
For infection not to recur requires that there is neither incidence of new infection from infective mosquito bites, nor recrudescence. The observed mean rate of recurrence of infection in the highlands of 0.220 implies a probability that recurrence will not occur of 0.780. On the reasonable assumption of independence of the processes of recrudescence and new infection:
(1-rate of new infection per child week) = 0.780/(1-0.013),
thus the rate of new infection can be calculated to have been 0.210 in the highlands. By the same method it was calculated to have been 0.415 in the lowlands.
These rates are based on weekly monitoring for infection and, in some cases, there could have been two or more separate new infection events in the same week; these would have been indistinguishable by our slide reading without molecular testing. Assuming independent occurrence of infection events there would be a Poisson distribution of weeks with 0,1,2,3... infections. Thus, if the probability of incidence of an infection is p, the proportion of child weeks with no infection would be 1/ep.
Therefore the above estimates, after correction for recrudescence, can be further corrected for multiple infection in the same week by calculating for the highlands that the probability of incidence of a new infection per week is:
ln (1/(1-0.210)) = 0.236; similarly the incidence in the lowlands = 0.536,
Note that without either of these corrections applied the estimates for the highlands and lowlands were 0.220 and 0.423, i.e. the corrections are of more than academic interest in the intense transmission conditions in the lowlands.
Table 2 shows that in the months of April to June 1999 the mean light trap catch in the highlands was 3.80 per night, i.e. 26.6 per week, corresponding to a rate of biting per person per week of 39.9, assuming the same conversion factor from light trap catches to human biting as was found in the lowlands [8]. With the observed CSP rate for the highlands of 3.62%, this corresponds to an entomological inoculation rate of 1.45 CSP positive bites per person per week. Thus the estimated probability that a CSP positive bite in the highlands would lead to a patent infection was 0.236/1.45 = 0.163. Similarly, with the data from Table 2 for the same time period in the lowlands, there were 605 bites per person per week and 26.7 CSP positive bites per person per week. Thus, in the lowlands, the probability that a CSP positive bite would lead to a patent infection was only 0.536/26.7 = 0.020.
This is a further example of a phenomenon reported, for example, in [12, 13] and [19] that, as the number of infective bites per unit time decreased (for natural reasons or due to vector control measures), so the probability that each bite would cause a patent infection tended to increase. This would seem to be a response of the immune system to less frequent inoculations of sporozoites [20].
Prevalence of malaria infection and mild malaria morbidity
Table 5 shows data by age group from monthly surveys in the pre-intervention year which agree with Ellman et al [6] in showing highly significantly greater prevalence of malaria infection and mild malaria morbidity in the lowlands than the highlands, up to age 5 years. This was also true for three of the parameters in the 6–12 age group which was not studied by Ellman et al [6]. For malaria fever, there was no significant difference between highland and lowland in the oldest age group, but there was no evidence that the benefit to the young children of the reduced transmission in the highlands was "paid for" by more malaria fever later in childhood. In fact there was far less malaria fever and anaemia in the older children than the younger ones.
Table 6 shows the impact of introducing treated nets in hamlets at both altitudes. For the youngest children this gave highly significant benefits in all respects measured, and the 95% confidence limits of the odds ratios in the highlands and lowlands overlapped. Thus there was no evidence that the benefit of the nets was any less in the initially intensely malarious lowlands than in the more moderate conditions in the highlands. In the 6–12 age group, significant benefits were seen in the highlands for all parameters measured. In this age group in the lowlands the benefits were not significant. However, the 95% confidence limits of the odds ratios for the two altitudes overlapped and, in no case, approached evidence for a disadvantage to older children of the intervention which reduced the intense transmission in the lowlands. The concentration of the benefit of treated nets among the younger children was also found 3–4 years after introduction of treated nets into nearby villages [5].
We are still working on data on acute malaria episodes from villages with and without nets as recorded at the District Hospital and via resident village reporters.
Conclusions
Our data on mild malaria morbidity is re-assuring in indicating that treated nets are about equally beneficial to young children, and not disadvantageous to older children, whether the initial transmission is intense or more moderate. Thus we consider that the present WHO policy of encouraging high coverage with effectively treated nets all over Africa is the correct one and there is no justification for focusing only on hypo-endemic highland areas.
References
World Health Organization/UNICEF: The Africa Malaria Report 2003. WHO/CDS/MAL/203.1093.
Trape JE, Rogier C: Combating malaria morbidity and mortality by reducing transmission. Parasit Today. 1996, 12: 236-240. 10.1016/0169-4758(96)10015-6.
Touré YT, Coluzzi M: The challenges of doing more against malaria, particularly in Africa. Bull World Health Organ. 2000, 78: 1376-
Lengeler C: Insecticide-treated bednets and curtains forpreventing malaria. Cochrane Library, issue 1. Oxford University update software. 2001
Maxwell CA, Msuya E, Sudi M, Njunwa KJ, Carneiro IA, Curtis CF: Effect on malaria morbidity of 3–4 years community-wide use of insecticide treated nets in Tanzanian villages. Trop Med Int Hlth. 2002, 7: 1003-1008. 10.1046/j.1365-3156.2002.00966.x.
Ellman R, Maxwell C, Finch R, Shayo D: Malaria and anaemia at different altitudes in the Muheza district of Tanzania: childhood morbidity in relation to level of exposure to infection. Ann Trop Med Parasit. 1998, 92: 741-753. 10.1080/00034989858989.
World Health Organization Pesticide Evaluation Scheme: Report of the W.H.O. Informal Consultation on the evaluation and testing of insecticides. CTD/WHOPES/IC/96.1. 1996
Lines JD, Curtis CF, Wilkes TJ, Njunwa KJ: Monitoring human biting mosquitoes in Tanzania with light-traps hung beside mosquito nets. Bull Ent Res. 1991, 81: 77-84.
Curtis CF, Pates HV, Takken W, Maxwell CA, Myamba J, Priestman A, Akinpelu O, Yayo AM, Hu JT: Biological problems with the replacement of a vector population by Plasmodium-refractory mosquitoes. Parassitologia. 1999, 41: 479-481.
Burkot TR, Williams JL, Schneider I: Identification of Plasmodium falciparum infected mosquitoes by double antibody enzyme linked immunosorbent assay. Am J Trop Med Hyg. 1984, 33: 783-788.
Trigg J, Mbwana H, Chambo O, Hills E, Watkins W, Curtis CF: Resistance to pyrimethamine-sulfadoxine in Plasmodium falciparum in 12 villages in north-east Tanzania and a test of chlorproguanil-dapsone. Acta Trop. 1997, 63: 185-189. 10.1016/S0001-706X(96)00617-1.
Curtis CF, Maxwell CA, Finch RJ, Njunwa KJ: A comparison of use of a pyrethroid either for house spraying or for bednet treatment against malaria vectors. Trop Med lnt Hlth. 1998, 3: 619-631.
Maxwell CA, Myamba J, Njunwa KJ, Greenwood BM, Curtis CF: Comparison of bednets impregnated with different pyrethroids for their impact on mosquitoes and on re-infection with malaria after clearance of pre-existing infections with chlorproguanil-dapsone. Trans R Soc Trop Med Hyg. 1999, 93: 4-11.
Mnzava AEP, Kilama W: Observations on the distribution of the Anopheles gambiae complex in Tanzania. Acta Tropica. 1986, 43: 277-282.
Malima R: Sporozoite rates and species identity of mosquitoes collected from highland and lowland Tanzania. M.Sc. thesis,. 1999, University of London.
Tsang L: Tests of Mosquitoes collected in Tanzania for CSP by ELISA. M.Sc. thesis,. 2000, University of London.
Gokool S, Curtis CF, Smith DF: Analysis of mosquito bloodmeals by DNA profiling. Med Vet Ent. 1993, 7: 208-215.
Curtis J, Maxwell CA, Msuya FHM, Mkongewa S, Alloueche A, Warhurst D: Mutations at dhfr in Plasmodium falciparum infections selected by chlorproguanil-dapsone treatment. J Inf Dis. 2002, 186: 1861-1864. 10.1086/345765.
Charlwood JD, Smith T, Lyimo E, Kitua AY, Masanja H, Booth M, Alonso PL, Tanner M: Incidence of Plasmodium falciparum infection in infants in relation to exposure to sporozoite-infected anophelines. Amer J Trop Med Hyg. 1998, 59: 243-251.
Metzger WG, Maxwell CA, Curtis CF: Anti-sporozoite immunity and impregnated bednets in Tanzanian villages. Ann Trop Med Parasit. 1998, 92: 727-729. 10.1080/00034989859195.
Stata Corporation: Stata Statistical Software. Stata Corp, College Station, Texas.
Acknowledgements
We thank the people of the 21 hamlets for their excellent co-operation throughout this project. We thank Rukia Ahmed, Oluyomi Akinpelu, Mwantum Kabanda, Steven Mkongewa, Fikirini Msuya, Donald Mwanjaluka, Mzee Mziray, Constantine Njunwa, Ahmed Nuru, Deogratias Shayo, Musa Sudi and Martin Zuwakuu for their skilful and conscientious work.
Financial support from the British Medical Research Council is gratefully acknowledged. The alphacypermethrin was provided by Cyanamid/BASF.
Chlorproguanil-dapsone was provided by W Watkins.
CAM and CFC have clearance from the Tanzanian Commission of Science and Technology for their work in Tanzania. This paper is published by permission of the Director General of the Tanzanian National Institute for Medical Research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
Caroline Maxwell: organised the data collection in the field.
William Chambo, Mathew Mwalimu and Frank Magogo: led the field teams.
Ilona Carneiro: statistical analysis of malaria morbidity data.
Christopher Curtis: strategic planning of the trial and analysis of entomological and malaria incidence data.
Competing interests
None declared.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Maxwell, C.A., Chambo, W., Mwaimu, M. et al. Variation of malaria transmission and morbidity with altitude in Tanzania and with introduction of alphacypermethrin treated nets. Malar J 2, 28 (2003). https://doi.org/10.1186/1475-2875-2-28
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-2-28