
Abstract
Background
Staphylococcus aureus is unrestrictedly found in humans and in animal species that maintain thermal homeostasis. Inadequate cleaning of processing equipment or inappropriate handling can contaminate processed food and cause severe food poisoning. Staphylococcal enterotoxin B (SEB), a potent superantigenic exotoxin, is produced by 50% of clinical isolates of S. aureus and is associated with massive food poisoning and with the induction of toxic shock syndrome.
Results
A gene sequence encoding a recombinant SEB (rSEB), devoid of superantigenic activity, was successfully cloned and expressed in a cytoplasmic or a secreted form in the food-grade lactic acid bacterium Lactococcus lactis. The recombinant protein detected in the cytoplasm or in the culture medium exhibited the expected molecular mass and was recognized by a SEB-polyclonal antibody. Oral immunization with the recombinant L. lactis strains induced a protective immune response in a murine model of S. aureus infection. Immunized mice survived intraperitoneal challenge with an S. aureus SEB-producer strain. Counts of S. aureus in the spleen of rSEB-immunized mice were significantly reduced. The rSEB-immunized mice showed significant titers of anti-SEB IgA and IgG in stools and serum, respectively. Both recombinant L. lactis strains were able to elicit cellular or systemic immune responses in mice, with no significant difference if rSEB was produced in its cytoplasmic or secreted form. However, recombinant L. lactis expressing the cytoplasmic rSEB increased the survival rate of the challenged mice by 43%.
Conclusions
These findings show the vaccine efficacy of L. lactis carrying an attenuated SEB, in a murine model, following lethal S. aureus challenge.
Similar content being viewed by others


Background
Staphylococcus aureus is a Gram-positive opportunistic pathogen of humans and warm-blooded animals, and is part of the commensal microbiota of the skin and nares in a significant proportion of the human population. It is a leading cause of bloodstream, lower respiratory tract, and skin and soft-tissue infections. Moreover, S. aureus has a wide range of virulence factors, including superantigens such as staphylococcal enterotoxins (SEs). These heat-stable toxins cause a self-limiting gastrointestinal intoxication, but parenteral exposures can cause a potentially fatal toxic shock syndrome [1–3]. Staphylococcal enterotoxin type B (SEB) is a single polypeptide of approximately 27 kDa, highly resistant to proteases [4]. As a superantigen, SEB is capable of massive activation of CD4+ lymphocytes, with subsequent secretion of cytokines and systemic inflammation [5]. Because of its remarkable toxicity and stability, SEB is considered a prime threat as a biological weapon of mass destruction [6–8]. Bacterial superantigens can be inactivated by rational site-directed mutagenesis, and these genetically altered constructs can be used for vaccine purposes [9, 10]. Additionally, various vaccination regimens of an attenuated SEB mutant protein containing L45R, Y89A and Y94A were effective in a primate model against aerosolized wild-type SEB, with a correlation between survival of rhesus monkeys, antibody titers, and neutralizing antibody [11].
Lactic acid bacteria (LAB) have been considered good candidates for controlled and targeted administration of heterologous proteins to the mucosal immune system [12, 13]. Lactococcus lactis is widely used as a starter in the dairy industry and is considered a model organism for LAB. Several genetic tools for the model LAB, Lactococcus lactis, were developed: transformation protocols, cloning- or screening-vectors and mutagenesis [14] (for a review) and the complete genome sequence is now available for several strains [15]. L. lactis has been extensively used for antigen delivery [12, 16–18] thanks to genetic tools allowing antigen production in different cellular compartments (intracellular, secreted, or anchored to the cell wall) [8, 19]. L. lactis has also been used to efficiently produce, secrete, and deliver therapeutic proteins to the mucosal tissues, specifically through the intranasal, oral, or genital mucosal surfaces [12, 17]. Sufficient data are now available to support the use of recombinant LAB, in particular L. lactis, to deliver therapeutic proteins to the mucosal tissues [8, 20]. Mucosal vaccination is able to stimulate an immune response at the site of invasion, providing a first line of defense against pathogens [13, 21]. In addition, oral immunization frequently evokes both local and systemic immune responses, resulting in the effective elimination of foreign invaders [13].
The objective of this study was to explore the efficacy of an oral vaccine in C57Bl/6 mice, using L. lactis strains to deliver a recombinant SEB protein lacking superantigenic activity [10]. This rSEB variant was obtained after mutations in a hydrophobic binding loop, polar binding pocket, and disulfide loop (L45R, Y89A, and Y94A, respectively) without affecting the antigenic characteristics of SEB [10, 11]. The humoral immune response against rSEB in mice was characterized, and its protective effect was evaluated through a challenge infection using a live SEB-producer strain of S. aureus.
Results
Engineering of recombinant L. lactis strains producing cytoplasmic and secreted forms of rSEB
Since the immune response to an antigen depends on its presentation [12] we engineered L. lactis for the intracellular delivery of rSEB or secretion of rSEB by L. lactis to the intestinal mucosa. Two expression vectors were initially constructed, pCYT:rSEB and pSEC:rSEB for the cytoplasmic expression or secretion of rSEB, respectively (see Methods for details). The pCYT:rSEB vector harbors a transcriptional fusion between the ribosome-binding site (RBSusp45) of the usp45 gene [22] and the DNA sequence encoding the mature moiety of rSEB, and the pSEC:rSEB harbors a transcriptional fusion between RBSusp45 and the DNA sequence encoding the signal peptide (SPusp45) of Usp45 plus rSEB. In both cases, rSEB expression was under the control of the xylose-inducible promoter, PxylT[14]. The ability of L. lactis to secrete rSEB or to accumulate rSEB intracellularly was examined using L. lactis (pSEC:rSEB) and L. lactis (pCYT:rSEB), respectively, after induction with 1% xylose. Cell pellet and supernatant protein samples from L. lactis carrying pCYT:rSEB or pSEC:rSEB, respectively, were prepared from late exponential-phase cultures (cells harvested at OD600 1.5), resolved by SDS-PAGE and transferred onto a PVDF membrane. Production of rSEB, in the culture medium or in the whole-cell extracts, was analyzed by Western blotting, using a commercial polyclonal anti-staphylococcal enterotoxin B antibody (anti-SEB) as a probe. A single band with the expected size for rSEB (27 kDa) – available in the Protein families database Pfam [http://www.sanger.ac.uk/Software/Pfam/search.shtml] - was identified in the cell pellet (Figure 1, lane 2) and in culture supernatants (Figure 1, lane 4) of induced L. lactis (pCYT:rSEB) and L. lactis (pSEC:rSEB), respectively. The 27 kDa polypeptide band was not detected in the samples prepared from non-induced recombinant L. lactis (Figure 1, lanes 3 and 5). Protein extracts of S. aureus ATCC 14458 (an SEB producer) and of a non-SEB producer S. aureus (strain from our laboratory collection) were used as positive (Figure 1, lanes 6, 7 and 8) and negative (Figure 1, lane 9) controls, respectively. Taken together, the results showed that L. lactis carrying (pCYT:rSEB) or (pSEC:rSEB) was able to produce rSEB intracellularly and also to secrete rSEB, both of which were properly recognized by the anti-SEB antibody.

Expression of rSEB in Lactococcus lactis pSEC:rSEB (secreted) and pCYT:rSEB (cytoplasm) after induction by 1% xylose, assayed in supernatants or cell pellets, respectively. rSEB expression was analyzed by Western blot using a commercial polyclonal anti-staphylococcal enterotoxin B antibody (anti-SEB), as a probe. rSEB (27 kDa) is indicated by an arrow. Lane 1 – prestained protein molecular weight markers (Fermentas); lanes 2 and 4, -induced (+ xyl); lanes 3 and 5, non-induced (+ xyl) cultures; lanes 6, 7 and 8, cell pellets from S. aureus cultures + SEB (ATCC 14458 strain/SEB producer strain); lane 9, – SEB (29 strain/SEB non-producer strain). The arrow indicates the SEB of an apparent molecular mass of 27 kDa.
Protective effect of oral immunization with rSEB
To determine the efficacy of the recombinant L. lactis strains as a live vaccine against S. aureus, mice were orally immunized with 109 CFU of each strain expressing rSEB, either in the cytoplasm or secreted in the intestinal mucosa. The rSEB expression was induced in each strain prior to oral administration, as described in the Methods section. Control groups were immunized with wild-type L. lactis NCDO2118 or with PBS. Mice received a booster immunization for three consecutive days. Fourteen days after the last immunization, the mice were challenged with a lethal dose of S. aureus ATCC 14458, a SEB producer strain (7 × 108 CFU per mouse), by intraperitoneal (i.p.) administration (Figure 2). Fourteen days after the S. aureus challenge, 100% of the mice vaccinated with L. lactis pCYT:rSEB and 70% of those vaccinated with L. lactis pSEC:rSEB survived. In contrast, only 10% of the mice injected with PBS or L. lactis NCDO2118 (wt strain) survived the lethal challenge (Figure 3). Three days after inoculation, the number of bacterial cells in the spleens was determined. The rSEB-immunized mice presented significantly fewer S. aureus cells in the spleens compared to the control mice (Figure 4). Bacterial counts were not significantly different between the animals that were immunized with the cytoplasmic or the secreted form of rSEB (p < 0.01). There was no significant difference in the counts of S. aureus in the spleen of control mice that received PBS or L. lactis NCDO 2118. These results indicate that each animal immunized with L. lactis pCYT:rSEB and L. lactis pSEC:rSEB developed protection against systemic S. aureus ATCC 14458 infection.

Immunization, serum and stool collection, and challenge schedule. To evaluate the rSEB-associated immune response in a murine model, four groups of 10 C57Bl/6 mice were tested, designated PBS, Lactococcus lactis, pCYT:rSEB, and pSEC:rSEB administered as shown in the schedule. Each group was immunized orally on days 0/1/2, and boosted on days 14/15/16 and on days 28/29/30. During the experiment, mice were bled three times, on days 14, 28 and 42. Stools were collected on days 15, 24 and 38. Animals were challenged by 7 × 108 CFU Staphylococcus aureus ATCC 14458. The animals were killed at the end of the experiment on day 48, before splenectomy.

Survival rates of immunized mice. 10 animals in each group previously immunized with Lactococcus lactis pCYT:rSEB, L. lactis pSEC:rSEB, L. lactis NCDO2118, or PBS were challenged with 7 × 108 CFU of Staphylococcus aureus ATCC 14458 (SEB producer) 14 days after the last boost.

Enumeration of Staphylococcus aureus in the spleen of immunized mice. 10 animals in each group were immunized with Lactococcus lactis pCYT:rSEB, L. lactis pSEC:rSEB, L. lactis NCDO2118, or PBS and then challenged with 7 × 108 CFU of S. aureus ATCC 14458 (SEB producer) on the 14th day after the last boost, as described in the Methods section. Significant results are marked with asterisks: *** p < 0.05.
In sum, oral immunization with rSEB delivered by L. lactis appeared to provide efficient protection and promoted survival against infection by a lethal S. aureus SEB producer.
Production of anti-SEB antibody in orally immunized mice
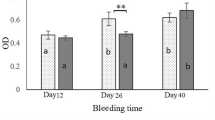
The production of SEB-specific IgG in serum and IgA antibody in feces was determined by specific ELISA during the time course of the immunization (Figure 5). Each animal responded with a strong, stable antibody response to SEB after the initial immunization (data not shown). The IgG antibody titer showed a significant increase 42 days after the first immunization (Figure 5A). Interestingly, immunization with L. lactis pSEC:rSEB induced an anti-SEB antibody production, which was stronger than that induced by L. lactis pCYT:rSEB, at days 14 and 28. However, specific anti-SEB antibody titers reached the same levels at day 42, in both immunizations. Fecal SEB-IgA antibodies increased significantly after immunization with rSEB-producing L. lactis strains with either the cytoplasmic or secreted form (Figure 5B). It appears that the intracellular presentation of rSEB by L. lactis or the presentation in a secreted form to the mucosal site results in a similar stimulation of local IgA antibodies.

Specific SEB antibodies. Sera (A) and stools (B) from the different groups of mice were collected and analyzed by ELISA after the first oral administration. Immune response was assayed in mice orally immunized with recombinant Lactococcus lactis pSEC:rSEB or pCYT:rSEB. Significant results are marked with asterisks: *, p < 0.05; ** p < 0.01. Values were recorded as standard deviation of samples from 10 mice per group.
The data presented here demonstrated that the oral administration of recombinant L. lactis strains carrying pSEC:rSEB or pCYT:rSEB stimulated both the serum-SEB IgG and mucosal IgA specific antibodies in mice.
Discussion
Production of several staphylococcal proteins in L. lactis has been reported. However, these studies were dedicated to the development of expression-secretion systems, e.g., staphylococcal nuclease used as a reporter protein [23]; to the characterization of staphylococcal virulence factors (e.g., ClfA and FnbA [24], ClfB [25], IsdA [26]); or to increase adhesion properties of recombinant L. lactis strains [27]. In addition, several staphylococcal antigen candidates have been tested alone or in combination for immunization in animal models [28, 29]. Proteins such as ClfA [30], recombinant forms of SEB [2] or SEC [31], as well as non-protein molecules such as capsular polysaccharides [29, 32, 33] are the most widely investigated S. aureus antigens. To our knowledge, this study reports, for the first time, the production of a staphylococcal antigen in a recombinant LAB strain to be used for oral vaccination. The two recombinant L. lactis strains constructed here allow for the production of a non-superantigenic rSEB, either intracellularly or secreted in the intestinal mucosa of mice, which stimulates the production of specific anti-SEB IgG antibodies in the serum and specific IgA in feces [12, 34]. There is an apparent discrepancy between survival and anti-SEB IgG production when compared the CYT:rSEB and SEC:rSEB groups. It should be considered that the antigen presentation pathway (secreted versus intracellular) may also impact the quality of neutralizing antibodies, the reason that these alternative forms were tested. The immune response can reflect the production of different classes of IgG or the production of IgGs against rSEB epitopes with distinct neutralizing abilities Oral immunization with the recombinant strains induced a protective immune response against a lethal challenge with S. aureus ATCC 14458, an SEB producer strain, in a murine model. Interestingly, analysis of the S. aureus ATCC 14458 exoproteome revealed that this strain produces and secretes, in addition to SEB, other enterotoxin-like proteins such as SElK and SElQ, together with other toxins such as alpha-hemolysin and gamma-hemolysin [35]. Thus, it seems that oral immunization with a recombinant L. lactis producing rSEB confers a protective immunity against a strain of Staphylococcus that produces a variety of enterotoxins. Alternatively, one might consider that SEB, as a potent superantigen, is preponderant in the pathogenesis of SEB-producing strains. TSST1 and SEB production was indeed shown to repress exotoxin synthesis in S. aureus[36]. Similarly, encouraging results were obtained using a recombinant SEC mutant vaccine against mastitis. Intramuscular immunization resulted in significant protection against a challenge with live SEC-producer S. aureus strains in dairy cows [31]. In addition to these SE-oriented vaccines, other immunization strategies against S. aureus were developed, based on other staphylococcal antigens that are widely distributed among S. aureus strains, such as adhesins [32], IsdB [28], ClfA or exopolysaccharides [30].
Previous studies reported a protective immunization against lethal challenge with SEB after oral or nasal immunization using SEBv, a non-superantigenic variant of SEB. Significant protection was observed only when cholera toxin was used as an adjuvant [26].
The effect of the vaccination route on immunogenicity has to be considered. In a previous study, Stiles et al.[2] showed that immunization via the nasal route yielded higher titers of circulating IgG when compared to the oral route. An intranasal immunization route yielded a better immune response, compared to the oral route, in a live-vaccine strategy using L. lactis producing the HPV16 E7 antigen [12]. Whether nasal immunization with our recombinant L. lactis strains would confer an even better protection remains to be tested.
Conclusions
Lactococcus lactis, a food-grade organism that carries and expresses an attenuated form of enterotoxin B (rSEB) appears to be an efficient delivery vehicle for immunization in mice against the toxic shock provoked by Staphylococcus aureus. The live vaccine can be administered orally and can be considered a promising tool to develop strategies for prevention of life-threatening toxic-shock syndrome.
Methods
Animals
Six- to eight-week-old C57Bl/6 mice were purchased from Cecal, Fiocruz, Rio de Janeiro, Brazil. Mice were housed in plastic cages under specific pathogen-free conditions at the Department of Immunology, UFRJ. The daily cycle consisted of 12 h light and 12 h darkness, and food and water were available at all times. The experiments were approved by the Institutional Animal Welfare Committee (approval ID: CEUA/CCS/UFRJ/IMPPG 011).
Bacterial strains, growth conditions and plasmids
Escherichia coli TG1 was aerobically grown at 37°C in Luria-Bertani medium. L. lactis NCDO2118 was grown anaerobically at 30°C in M17 medium supplemented with 0.5% glucose. When required, antibiotics were added as follows: ampicillin (100 μg/mL) for E. coli and chloramphenicol (10 μg/mL) for L. lactis. rSEB was cloned into lactococcal plasmids pCYT:Nuc and pSEC:Nuc under the regulation of a xylose-inducible promoter, PxylT[17]. rSEB was produced either in the cytoplasm (CYT) or into the extracellular medium (SEC). For the sake of simplicity, these constructions are hereafter referred to as pCYT:rSEB, and pSEC:rSEB, referring to the location of the recombinant protein.
DNA manipulations
General DNA manipulation techniques were carried out according to standard procedures [37]. DNA restriction and modification enzymes were used as recommended by the suppliers. DNA fragments were isolated from agarose gels with the Concert™ Rapid Gel Extraction System (Gibco BRL). PCR amplifications were performed using Taq DNA polymerase (Invitrogen) in a DNA thermocycler (MJ Research, Inc.). DNA plasmids from E. coli and L. lactis were isolated as previously described [37, 38].
The vectors to produce cytoplasmic or secreted rSEB, pCYT:rSEB, and pSEC:rSEB, respectively, were obtained as follows: A 721-bp DNA fragment encoding rSEB was PCR-amplified from a plasmid previously described [10] and subcloned into pGEM-T (Promega) cloning vector, resulting in pGEM:rSEB. A primer pair was then designed based on rSEB sequence: 5'GGCTGCAG AGAGTCAACCAGATCCT-3' for the coding strand containing a Pst I restriction site (underlined) and 5'GGGAATTC TCACTTTTTCTTTGTCGT-3' for the complementary strand, containing an EcoR I restriction site (underlined). The rSEB fragment resulting from Pst I and Eco RI digestion of pGEM:rSEB was cloned into a Nsi I-Eco RI-cut and the purified backbone of the pCYT:Nuc expression vector, replacing the DNA sequence encoding NucB. The pSEC:rSEB vector was constructed to target the rSEB protein to the extracellular medium (i.e., secreted) of L. lactis, using the following procedures: the rSEB gene was PCR-amplified from pGEM:rSEB using 5'GGCTGCAG AGAGTCAACCAGATCCT-3' (including a Pst I site; underlined) as forward primer and 5'GGGAATTC TCACTTTTTCTTTGTCGT-3' (including an EcoR I site; underlined) as reverse primer. The PCR product was then Pst I- and Eco RI-digested and cloned, in frame with the signal peptide coding sequence, into the purified backbone isolated from the Nsi I-EcoR I-cut pSEC:Nuc expression vector, again replacing the DNA sequence encoding for NucB. In both cases, pCYT:rSEB and pSEC:rSEB were first obtained in E. coli TG1 and then transferred into L. lactis NCDO2118. All constructions were confirmed by DNA sequencing.
Conditions of xylose induction
L. lactis strains harboring pCYT:rSEB or pSEC:rSEB were grown at 30°C overnight in 10 mL M17, containing 0.5% glucose. For xylose induction, 1 μL of the culture was transferred into 10 mL M17 medium supplemented with 1% xylose in the presence of chloramphenicol (10 ng/mL) [14].
Protein sample preparation and Western blotting analysis
Protein samples from L. lactis cultures were prepared as previously described [39], except for the use of protease inhibitors and mild protein precipitation procedures. Briefly, protein extracts were prepared from 2 mL of cultures centrifuged at 17,500 × g at 4°C for 10 min. The cell pellet and supernatant were treated separately. To inhibit proteolysis in supernatant samples, 1 mM phenylmethylsulfonyl fluoride (PMSF) and 10 mM dithiothreitol (DTT) were added. Proteins were then precipitated by addition of 100 μL of 100% trichloroacetic acid. The supernatant proteins were incubated for 1 h on ice, and collected by centrifugation (17,500 × g at 4°C for 20 min). Supernatant proteins were resuspended in 50 mM NaOH, 1 mM PMSF, and 1.2 μL DTT-LB (4 mL charge buffer solution, consisting of 0.1 Tris–HCl pH 6.8, 4% SDS, 10 ml glycerol, 0.2% bromophenol blue, 50 mL distilled water q.s.p., and 1 mL of 1 M DTT). The cell pellets were resuspended in 120 μL of TES-Lys buffer (25% sucrose, 1 mM EDTA, 50 mM Tris–HCl pH 8.0 and 1 mg/mL of lysozyme), 1.2 μL DTT and 1 mM PMSF. The suspension was incubated for 30 min at 37°C, and 20% SDS, 1.2 μL DTT-LB and 1 mM PMSF were added. Supernatant or pellet proteins were separated in a 12% PAGE-SDS [40]. The gel-resolved proteins were transferred onto a PVDF membrane and probed with a commercial polyclonal anti-staphylococcal enterotoxin B antibody (anti-SEB, Sigma-Aldrich). Immunodetection was carried out using peroxidase conjugates (Sigma-Aldrich).
Immunization and antibody detection
The animal study was conducted according to current Good Scientific Practice-principles (2000) and approved by the Ethical Committee of the UFRJ. Groups of C57Bl/6 mice (8–10 mice per group) were orally immunized with 109 CFU of each recombinant strain suspended in 100 μL of sterile PBS for 3 consecutive days. The immunization protocol was repeated twice at 2-week intervals. Control groups received sterile PBS or a plasmidless L. lactis NCDO2118, in the same conditions. Blood samples were drawn on days 0, 14, 28 and 42 after immunization. Serum samples were examined by ELISA for IgG titers with specific antigen-binding activity after coating 1 μg/mL SEB in carbonate buffer (S4881 - Sigma-Aldrich) onto microplates. Anti-Mouse IgG Peroxidase antibody (Sigma-Aldrich) diluted 1:1000 was used as the secondary antibody. Stool samples were obtained 0, 15, 24, and 38 days after immunization and were examined by ELISA for IgA titers with specific antigen-binding activity after coating the microplate in the same way as for IgG ELISA. The secondary antibody, Anti-Mouse IgA Peroxidase antibody (A4789, Sigma-Aldrich) was diluted 1:1000.
Inoculum for lethal challenge
S. aureus ATCC 14458, an SEB-producer strain, was first grown on BHI agar (Difco) for 18 h. Freshly grown colonies were suspended in BHI broth (Difco) and incubated overnight at 37°C. Bacteria were harvested by centrifugation at 4000 × g for 10 min at 4°C, and cell density was adjusted to 7 × 108 CFU/mL. The inoculum size was confirmed by serial dilutions and quantitative subcultures on Baird Parker agar, and the infecting dose was established based on constructed lethality curves. The challenge dosage was established by a lethality assay, where the dosage ranged from 109 CFU to 5 × 108 CFU of S. aureus ATCC14458 administered to 6 animals with similar body mass. The concentration of 7×108 CFU was able to kill approximately 70% of animals.
Challenge tests
10 mice in each group (PBS, L. lactis, pSECrSEB, and pCYTrSEB) were challenged 14 days after the last booster with a lethal dose of S. aureus ATCC 14458 by intraperitoneal (i.p.) injection. Untreated mice (control groups) were infected in the same way. Mice were killed on the third day after challenge (day 3) and the spleen was excised, weighed, and homogenized in 1 mL sterile PBS. Homogenates were analyzed by plating 10-fold serial dilutions, in duplicate, on Baird Parker agar supplemented with egg yolk tellurite emulsion. Colonies were counted after incubation at 37°C for 18 h. Results were expressed as log of CFU/g of spleen. Mouse deaths were recorded for 15 days.
Statistical analysis
Data were analyzed by ANOVA, followed by Bonferroni’s test to determine the significance of differences in antibody titers between the control and experimental groups. A plot of the Kaplan-Meier method was used to estimate the survival fractions, and significance was determined by a log rank test. Differences in bacterial counts in the spleen were evaluated by Student’s t test and Tukey’s multiple comparison test.
Authors’ information
Yves Le Loir share credit for senior co-authorship of this work.
References
Casey AL, Lambert PA, Elliott TS: Staphylococci. Int J Antimicrob Agents. 2007, 29: 23-32.
Stiles BG, Garza AR, Ulrich RG, Boles JW: Mucosal vaccination with recombinantly attenuated staphylococcal enterotoxin B and protection in a murine model. Infect Immun. 2001, 69: 2031-2036. 10.1128/IAI.69.4.2031-2036.2001.
Trilla A, Miro JM: Identifying high risk patients for Staphylococcus aureus infections: skin and soft tissue infections. J Chemother. 1995, 7: 37-43.
Swaminathan S, Furey W, Pletcher J, Sax M: Crystal structure of staphylococcal enterotoxin B, a superantigen. Nature. 1992, 359: 801-806. 10.1038/359801a0.
Madsen JM: Toxins as weapons of mass destruction: a comparison and contrast with biological-warfare and chemical-warfare agents. Clin Lab Med. 2001, 21: 593-605.
Ahanotu E, Alvelo-Ceron D, Ravita T, Gaunt E: Staphylococcal enterotoxin b as a biological weapon: recognition, management, and surveillance of staphylococcal enterotoxin. Appl Biosafety. 2006, 11: 120-126.
Franz DR, Jahrling PB, Friedlander AM, McClain DJ, Hoover DL, Bryne WR, Pavlin JA, Christopher GW, Eitzen EM: Clinical recognition and management of patients exposed to biological warfare agents. JAMA. 1997, 278: 399-411. 10.1001/jama.1997.03550050061035.
Leggiadro RJ: The threat of biological terrorism: a public health and infection control reality. Infect Control Hosp Epidemiol. 2000, 21: 53-56. 10.1086/501700.
Bavari S, Dyas B, Ulrich RG: Superantigen vaccines: a comparative study of genetically attenuated receptor-binding mutants of staphylococcal enterotoxin. Am J Infect Dis. 1996, 174: 338-345. 10.1093/infdis/174.2.338.
Ulrich RG, Olson MA, Bavari S: Development of engineered vaccines effective against structurally related bacterial superantigens. Vaccine. 1998, 16: 1857-1864. 10.1016/S0264-410X(98)00176-5.
Boles JW, Pitt ML, LeClaire RD, Gibbs PH, Torres E, Dyas B, Ulrich RG, Bavari S: Generation of protective immunity by inactivated recombinant staphylococcal enterotoxin B vaccine in nonhuman primates and identification of correlates of immunity. Clin Immunol. 2003, 108: 51-59. 10.1016/S1521-6616(03)00066-4.
Bermúdez-Humarán LG, Kharrat P, Chatel J-M, Langella P: Lactococci and lactobacilli as mucosal delivery vectors for therapeutic proteins and DNAvaccines. Microbial Cell Factories. 2011, 10: 1-10. 10.1186/1475-2859-10-1.
Pouwels PH, Leer RJ, Shaw M, Heijne den Bak-Glashouwer MJ, Tielen FD, Smit E, Martinez B, Jore J, Conway PL: Lactic acid bacteria as antigen delivery vehicles for oral immunization purposes. Int J Food Microbiol. 1998, 41: 155-167. 10.1016/S0168-1605(98)00048-8.
Le Loir Y, Azevedo V, Oliveira SC, Freitas DA, Miyoshi A, Bermúdez-Humarán LG, Nouaille S, Ribeiro LA, Leclercq S, Gabriel JE, Guimaraes VD, Oliveira MN, Charlier C, Michel Gautier M, Philippe Langella P: Protein secretion in Lactococcus lactis: an efficient way to increase the overall heterologous protein production. Microbial Cell Factories. 2005, 4: 2- 10.1186/1475-2859-4-2.
Bolotin A, Wincker P, Mauger S, Jaillon O, Malarne K, Weissenbach J, Dusko Erlich S, Sorokin A: The Complete Genome Sequence of the Lactic Acid Bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11: 731-753. 10.1101/gr.GR-1697R.
Frossard CP, Steidler L, Eigenmann PA: Oral administration of an IL-10-secreting Lactococcus lactis strain prevents food-induced IgE sensitization. J Allergy Clin Immunol. 2007, 119: 952-959. 10.1016/j.jaci.2006.12.615.
Medina M, Villena J, Salva S, Vintiñi E, Langella P: Nasal administration of Lactococcus lactis improves local and systemic immune responses against Streptococcus pneumoniae. Microbiol Immunol. 2008, 52: 399-409. 10.1111/j.1348-0421.2008.00050.x.
Wells JM, Mercenier A: Mucosal delivery of therapeutic and prophylactic molecules using lactic acid bacteria. Nat Rev Microbiol. 2008, 6: 349-362. 10.1038/nrmicro1840.
Le Loir Y, Azevedo V, Oliveira SC, Freitas DA, Miyoshi A, Bermúdez-Humarán LG, Nouaille S, Ribeiro LA, Leclercq S, Gabriel JE, Guimaraes VD, Oliveira MN, Charlier C, Gautier M, Langella P: Protein secretion in Lactococcus lactis: an efficient way to increase the overall heterologous protein production. Microb Cell Fact. 2005, 4: 2- 10.1186/1475-2859-4-2.
Bermúdez-Humarán LG: Lactococcus lactis as a live vector for mucosal delivery of therapeutic proteins. Hum Vacc. 2009, 5: 264-267. 10.4161/hv.5.4.7553.
Streatfield S: Mucosal immunization using plant-based oral vaccines. Methods. 2006, 38: 150-157. 10.1016/j.ymeth.2005.09.013.
van Asseldonk M, Rutten G, Oteman M, Siezen RJ, de Vos WM, Simons G: Cloning of usp45, a gene encoding a secreted protein from Lactococcus lactis subsp. lactis MG1363. Gene. 1990, 95: 155-160. 10.1016/0378-1119(90)90428-T.
Le Loir Y, Gruss A, Ehrlich SD, Langella P: Direct screening of recombinants in Gram-positive bacteria using the secreted staphylococcal nuclease as a reporter. J Bacteriol. 1994, 176: 5135-5139.
Que YA, François P, Haefliger JA, Entenza JM, Vaudaux P, Moreillon P: Reassessing the role of Staphylococcus aureus clumping factor and fibronectin-binding protein by expression in Lactococcus lactis. Infect Immun. 2001, 69: 6296-6302. 10.1128/IAI.69.10.6296-6302.2001.
Clarke SR, Walsh EJ, Dufrêne YF, Foster TJ, Foster SJ: Iron-regulated surface determinant protein A mediates adhesion of Staphylococcus aureus to human corneocyte envelope proteins. Infect Immun. 2009, 77: 2408-2416. 10.1128/IAI.01304-08.
Innocentin S, Guimarães V, Miyoshi A, Azevedo V, Langella P, Chatel JM, Lefèvre F: Lactococcus lactis expressing either Staphylococcus aureus fibronectin-binding protein A or Listeria monocytogenes internalin A can efficiently internalize and deliver DNA in human epithelial cells. Appl Environ Microbiol. 2009, 75: 4870-4878. 10.1128/AEM.00825-09.
Harro CD, Bett RF, Hartzel JS, Onorato MT, Lipka J, Smugar SS, Kartsonis NA: The immunogenicity and safety of different formulations of a novel Staphylococcus aureus vaccine (V710): Results of two Phase I studies. Vaccine. 2012, 30: 1729-1736. 10.1016/j.vaccine.2011.12.045.
Schaffer AC, Lee JC: Staphylococcal vaccines and immunotherapies. Infect Dis Clin N Am. 2009, 23: 153-171. 10.1016/j.idc.2008.10.005.
Schlievert PM: Use of intravenous immunoglobulin in the treatment of staphylococcal and streptococcal toxic shock syndromes and related illnesses. J Allergy Clin Immunol. 2001, 108: 107-110. 10.1067/mai.2001.117820.
Nanra JS, Timofeyeva Y, Buitrago SM, Sellman BR, Dilts DA, Fink P, Nunez L, Hagen M, Matsuka YV, Mininni T, Zhu D, Pavliak V, Green BA, Jansen KU, Anderson AS: Heterogeneous in vivo expression of clumping factor A and capsular polysaccharide by Staphylococcus aureus: implications for vaccine design. Vaccine. 2009, 27: 3276-3280. 10.1016/j.vaccine.2009.01.062.
Chang BS, Moon JS, Kang HM, Kim Y-I, Lee H-K, Kim J-D, Lee B-S, Koo HC, Park YH: Protective effects of recombinant staphylococcal enterotoxin type C mutant vaccine against experimental bovine infection by a strain of Staphylococcus aureus isolated from subclinical mastitis in dairy cattle. Vaccine. 2008, 26: 2081-2091. 10.1016/j.vaccine.2008.02.043.
Castagliuolo I, Piccinini R, Beggiao E, Palù G, Mengoli C, Ditadi F, Vicenzoni G, Zecconi A: Mucosal genetic immunization against four adhesins protects against Staphylococcus aureus-induced mastitis in mice. Vaccine. 2006, 24: 4393-4402. 10.1016/j.vaccine.2006.02.055.
Tuchscherr LP, Buzzola FR, Alvarez LP, Lee JC, Sordelli DO: Antibodies to capsular polysaccharide and clumping factor A prevent mastitis and the emergence of unencapsulated and small-colony variants of Staphylococcus aureus in mice. Infect Immun. 2008, 76: 5738-5744. 10.1128/IAI.00874-08.
Reveneau N, Geoffroy MC, Locht C, Chagnaud P, Mercenier A: Comparison of the immune responses induced by local immunizations with recombinant Lactobacillus plantarum producing tetanus toxin fragment C in different cellular locations. Vaccine. 2002, 20: 1769-1777. 10.1016/S0264-410X(02)00027-0.
Pocsfalvi G, Cacace G, Cuccurullo M, Serluca G, Sorrentino A, Schlosser G, Blaiotta G, Malorni A: Proteomic analysis of exoproteins expressed by enterotoxigenic Staphylococcus aureus strains. Proteomics. 2008, 8: 2462-2476. 10.1002/pmic.200700965.
Vojtov N, Ross HF, Novick RP: Global repression of exotoxin synthesis by staphylococcal superantigens. Proc Natl Acad Sci U S A. 2002, 99: 10102-10107. 10.1073/pnas.152152499.
Sambrook J, Fritsch EF, Maniatis T: Molecular cloning: a laboratory manual. 1989, New York: Cold Spring Harbor
Langella P, Le Loir Y, Ehrlich SD, Gruss A: Efficient plasmid mobilization by pIP501 in Lactococcus lactis subsp. lactis. J Bacteriol. 1993, 175: 5806-5813.
Le Loir Y, Gruss A, Ehrlich SD, Langella P: A nine-residue synthetic propeptide enhances secretion efficiency of heterologous proteins in Lactococcus lactis. J Bacteriol. 1998, 180: 1895-1903.
Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970, 227: 680-685. 10.1038/227680a0.
Acknowledgments
This work was supported by FAPERJ, CNPq, and CAPES - COFECUB project 359–09. GFA received a CNPq scholarship, and YLL is the recipient of CNPq grant PVE Nivel1.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
GFA performed all the experimental procedures and was the primary author of the manuscript; NFFS carried out the western-blotting assays; NFFFD, DDF, and MTB participated in immunization studies and challenge tests; KM, VACA, AM, and RU participated in molecular biology studies; JTS participated in the study design and data analysis; YLL participated in the coordination of the study and drafted the manuscript; VMFP conceived the study, participated in its design and coordination, and drafted the manuscript. RU critically revised the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Asensi, G.F., de Sales, N.F.F., Dutra, F.F. et al. Oral immunization with Lactococcus lactis secreting attenuated recombinant staphylococcal enterotoxin B induces a protective immune response in a murine model. Microb Cell Fact 12, 32 (2013). https://doi.org/10.1186/1475-2859-12-32
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2859-12-32