
Abstract
Background
The objective of this study was to analyze factors influencing the risk and timing of Alzheimer's disease (AD) in central Norway. The APOE ε4 allele is the only consistently identified risk factor for late onset Alzheimer's disease (LOAD). We have described the allele frequencies of the apolipoprotein E gene (APOE) in a large population of patients with AD compared to the frequencies in a cognitively-normal control group, and estimated the effect of the APOE ε4 allele on the risk and the age at onset of AD in this population.
Methods
376 patients diagnosed with AD and 561 cognitively-normal control individuals with no known first degree relatives with dementia were genotyped for the APOE alleles. Allele frequencies and genotypes in patients and control individuals were compared. Odds Ratio for developing AD in different genotypes was calculated.
Results
Odds Ratio (OR) for developing AD was significantly increased in carriers of the APOE ε4 allele compared to individuals with the APOE ε3/ε3 genotype. Individuals carrying APOE ε4/ε4 had OR of 12.9 for developing AD, while carriers of APOE ε2/ε4 and APOE ε3/ε4 had OR of 3.2 and 4.2 respectively. The effect of the APOE ε4 allele was weaker with increasing age. Carrying the APOE ε2 allele showed no significant protective effect against AD and did not influence age at onset of the disease. Onset in LOAD patients was significantly reduced in a dose dependent manner from 78.4 years in patients without the APOE ε4 allele, to 75.3 in carriers of one APOE ε4 allele and 72.9 in carriers of two APOE ε4 alleles. Age at onset in early onset AD (EOAD) was not influenced by APOE ε4 alleles.
Conclusion
APOE ε4 is a very strong risk factor for AD in the population of central Norway, and lowers age at onset of LOAD significantly.
Similar content being viewed by others
Background
Apolipoprotein E (APOE) is a multifunctional circulating lipoprotein consisting of 299 amino acids, synthesized in various organs, including liver, spleen, kidney and brain [1]. Three common isoforms of the protein are identified as ApoE 2, ApoE 3 and ApoE 4. These isoforms are coded for by three different alleles (denoted by a specific combination of two coding SNPs), located within the APOE gene on chromosome 19q13.2. APOE ε2, APOE ε3 and APOE ε4 account for more than 99% of the variation of APOE [2]. The frequencies of these three APOE alleles are highly variable in different populations [3–5]. Globally, APOE ε3 is the most frequently occurring allele, constituting 60–90% of the allelic variation. APOE ε2 constitutes 0–20% and APOE ε4 10–20% of allelic variation [5, 6]. APOE ε3 and APOE ε4 are also found to be inversely correlated in European populations with the APOE ε4 allele found more frequently in populations in northern Europe than in southern Europe [4].
Carrying the APOE ε4 allele is a risk factor for early onset Alzheimer's disease (AD), and is the only consistently identified risk factor for late-onset Alzheimer's disease (LOAD) [7–11]. APOE ε4 acts in a dose-dependant manner: carriers of two APOE ε4 alleles have a higher risk and earlier onset of AD than heterozygous subjects [12–14].
Only a few studies have examined the APOE allele frequencies in AD patients in Scandinavia [15–19]. In the present study we describe APOE genotypes and allele frequencies in AD patients and a cohort of controls from central Norway where the population is comparatively genetically homogeneous.
Methods
The clinical material consisted of 376 patients diagnosed with probable or possible AD and 561 cognitively-normal controls, all ethnic Norwegians. The patients were neurological and geriatric patients recruited from the University Hospital of Trondheim, the district hospital in Namsos and patients from nursing homes in central Norway. The inclusion period lasted from May 2003 until September 2006. Patients or suitable proxies were asked about case history, including a family history of dementia. Neurological examination was completed by one neurologist (SBS) in all patients and included the Mini-Mental State Examination (MMSE) [20] and Clock Drawing Test [21]. Blood screening was performed, and secondary causes of dementia were excluded. Additional tests including EEG, lumbar puncture or an olfactory test were performed in subgroups [22]. Blood samples for DNA extraction and genetic testing were obtained from all patients and control subjects.
The guidelines given in the International Classification of Diseases tenth revision (ICD-10) were applied for diagnosing dementia. Patients diagnosed with Alzheimer's disease met the Criteria for probable or possible AD from the National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer's disease and Related Disorders Association (NINCDS-ADRDA) [23].
Healthy spouses and members of societies for retired people in central Norway were recruited for the cognitively-normal control group. Only ethnic Norwegians without known dementia in first degree relatives were included. Both patients and controls were asked for place of birth, place of residence during childhood, number of years of education, and former occupation. Informed consent was obtained from patients or suitable proxies and from all control subjects. The study was approved by the Regional Committee for Medical Research Ethics in central Norway.
Genotyping
Overall APOE genotype was determined by the combinations of genotypes at two single nucleotide polymorphisms (SNPs), rs7412 and rs429358. Genotyping was performed using predesigned Taqman Assays on the Applied Biosystems 7900 HT Fast Real Time PCR system, and genotype calls were made using the SDS v2.2 software (Applied Biosystems). Any sample in which the Taqman assays gave inconclusive allele calls was re-genotyped using a restriction fragment length polymorphism (RFLP) method [24]. Deviation from Hardy Weinberg Equilibrium (HWE) was calculated for the APOE locus, and both cases (p = 0.86) and controls (p = 0.36) were found to be in HWE.
Statistical analyses
Data analysis was chiefly performed with the SPSS, version 13. Categorical variables were compared using Pearson's chi-square test. Age at onset in different groups was compared by Wilcoxon-Mann-Whitney's test. Odds ratios (OR) were calculated for each genotype by binary logistic regression, using the ε3/ε3 genotype as reference value. Logistic regression was performed with exact conditional maximum likelihood and median unbiased estimation in LogXact. Two-sided p-value < 0.05 was considered significant. Standard deviation (SD) is given after mean value as ± SD.
Results
264 patients were diagnosed with probable AD (70.2%), and 112 with possible AD (29.8%) according to the NINCDS-ADRDA criteria. 263 were women (69.9%) and 113 were men (30.1%). 213 of the patients had first degree relatives with dementia (56.6%), and 149 (39.6%) had no known dementia in parents, siblings or children. Family history was missing in 14 patients. Mean age at inclusion for patients was 79.5 ± 8.2 years. The median MMSE score was 17 (interquartile range 11–22). MMSE scores were missing in four patients.
The control group consisted of 561 subjects; 338 women (60.2%) and 223 men (39.8%) (Table 1). Age at inclusion for control individuals was 75.1 ± 7.3 years.
Allele frequencies and genotypes were calculated for patients and controls, and for gender in each group (Table 1). The allele frequencies for men and women did not differ significantly either in controls or patients. The differences in allele frequencies in patients and controls were significant. Allele frequencies were also calculated for patients and control individuals according to age (Table 2). The frequency of the APOE ε4 allele in patients was highest in those with age at onset 60–69 years (51.4%). The oldest patients with onset ≥ 80 years had the lowest proportion of the APOE ε4 allele (24.8%). While 64.1% (n = 241) of the AD patients had one or two APOE ε4 alleles, only 26.4% (n = 148) of the control individuals carried an APOE ε4 allele (p < 0.001). In patients with dementia in first degree relatives 70.9% (n = 151) had one or two APOE ε4 alleles, compared to 56.4% (n = 84) in patients with no known history of dementia in first degree relatives (p = 0.004).
The odds ratios (OR) for developing AD were calculated for each APOE genotype, using ε3/ε3 as the reference value (Table 3). These analyses were also carried out separately for patients with and without dementia in first degree relatives. Significantly increased ORs were found in all genotypes containing the APOE ε4 allele, both for the group with dementia in first degree relatives and for those with a negative family history. Odds Ratios for genotypes containing an APOE ε4 allele were increased in the group with a positive family history compared to those without dementia in first degree relatives. Logistic regression with the number of APOE ε2 and APOE ε4 alleles as covariates showed no significant protective effect of either one APOE ε2 allele (OR = 0.82; p = 0.32) or two APOE ε2 alleles (OR = 0.73; p = 0.78). Logistic regression analysis with APOE ε4 and age as covariates, showed a non-significant (p = 0.14) interaction, with a reduced effect of APOE ε4 in the older patients.
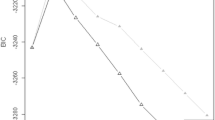
Age at onset in patients with LOAD was significantly reduced by the APOE ε4 allele in a dose dependent manner, while it had no lowering effect in patients with onset before 65 years (early onset AD, EOAD) (Table 4). In LOAD patients without the APOE ε4 allele, mean age at onset was 78.4 years, whereas those carrying one APOE ε4 allele had onset at 75.3 years (p = 0.005). For patients with two APOE ε4 alleles, age at onset was further reduced to 72.9 years. The difference in onset between carriers of one and two APOE ε4 alleles was also significant (p = 0.002). Figure 1 shows onset in carriers of zero, one and two APOE ε4 alleles. The presence of an APOE ε2 allele had no significant effect on age at onset.

Cumulative proportion of diseased patients.
Discussion
In this study we have described the APOE allele frequencies in 376 AD patients and 561 healthy controls from central Norway. Individuals carrying the APOE ε4 allele had an increased OR for developing AD, both in homozygous (12.9) and heterozygous individuals (4.2 for ε3/ε4, 3.2 for ε2/ε4). Age at onset in LOAD patients was significantly lowered by about three years for every APOE ε4 allele the patient carried. The APOE ε2 allele did not affect age at onset of AD and had no significant protective effect on the risk of AD.
While smaller studies in this field have been published from Norway, to our knowledge this is the first report of APOE allele frequencies and APOE genotypes in a large Norwegian AD population and the largest study on APOE allele frequencies in AD patients from Scandinavia to date. The study was performed in central Norway where the migration of the population for several centuries has been very modest. The population is therefore quite genetically homogeneous. The frequency of APOE ε4 in patients in the present study was similar to the result obtained in a large meta-analysis of 5107 Caucasian AD patients, where the APOE ε4 frequency was found to be 36.7% [25]. Age dependent variation in frequency of the APOE ε4 allele in AD patients has been reported earlier [26] and was also present in our study. In North America, the estimated APOE ε4 allele frequency is 60% in the AD population (15% ε4/4 and 40% with ε3/4 and < 5% with ε2/4) [27]. In our study, we found that 64.1% of the patients carried one or more APOE ε4 alleles (15.2% had ε4/4, 42.3% had ε3/ε4 and 6.6% had ε2/4). Our findings are thus in keeping with previous estimates in Caucasians.
The frequency of APOE ε4 in control individuals in the present study was also similar to the frequency in a meta-analysis of 6262 Caucasian control subjects from 1997 (13.7%) [25], though less than previously reported in a Norwegian study (19.8%) [28]. However, the latter study was conducted in 798 healthy blood donors aged < 40 years. A decrease in the frequency of the APOE ε4 allele with increasing age has been reported in healthy individuals [29]. As our control population was significantly younger than the AD patients, the APOE ε4 frequency might actually be slightly increased compared to controls of the same age as the patients. However, only control individuals without known dementia in first degree relatives were included in the present study, to increase the probability of detecting genetic differences between AD patients and control individuals. This selection may have caused a lower proportion of the APOE ε4 allele than in an unselected control population, as earlier findings suggest that offspring of AD patients have a frequency of the APOE ε4 allele that exceeds population estimates [30]. The frequency of patients reporting dementia in one or more first degree relatives was high in this population (56.6%). In most cases, both patients and next of kin were asked about relatives with cognitive impairment, and this may have increased the trend to report family members with dementia. Other studies have also reported a high occurrence of AD patients with dementia in first degree relatives [19, 31].
The present study confirmed that individuals carrying the APOE ε4 allele are at increased risk for developing AD. The ORs found in our study was close to those found previously in a large meta-analysis [25]. As the ORs calculated in the present study may slightly overestimate the effect of the APOE ε4 allele because of selection of the control group, we performed analyses of the population of patients with and without first degree relatives separately, using the same 561 selected controls for both analyses. This calculation showed increased ORs in the group consisting of patients with dementia in first degree relatives compared to the group without a known family history of dementia. The true estimate of OR is in between these estimates.
Age at onset in LOAD was strongly influenced by the number of APOE ε4 alleles in this study, and decreased by about three years for every APOE ε4 allele the patient carried. The decrease in age at onset of AD in carriers of the APOE ε4 allele is well known [12, 13], though the magnitude of the decrease in onset varies. A Finnish study found that age at onset decreased from 76 to 69 years in LOAD as the number of APOE ε4 alleles increased from 0 to 2 [32], whereas a twin study from Norway demonstrated no effect on age at onset of the APOE ε4 allele, probably due to the small number of patients included [17].
Occurrence of the APOE ε4 allele did not influence age at onset in patients with EOAD in the present study. Similar results are reported from another study with a larger sample size [29]. The occurrence of the APOE ε2 allele in control individuals in the present study was 11.3%, while the frequency in AD patients was 7.3%. Both of these APOE ε2 frequencies are increased compared to frequencies described in a meta-analysis [25], where the frequency for control individuals was 8.4% and for patients 3.9%. The increased APOE ε2 frequency may be a characteristic unique to this Norwegian population, as previous populations may have been too small to observe this increase [15, 17].
How the APOE ε4 allele exerts its influence is not fully understood, and the magnitude of the influence is also disputed. In the Framingham study [33] an increased risk for AD was found both in homozygous and heterozygous carriers of the APOE ε4 allele. However, most APOE ε4-carriers in the Framingham study did not develop AD. The authors emphasized that around half of all AD cases is not caused by APOE ε4. Others consider the APOE ε4 allele to be responsible for as much as 95% of the AD cases in North America [34]. However, not all patients with AD carry an APOE ε4 allele, and not all carriers of the APOE ε4 allele develop AD [35]. In the present study 35.9% of AD patients had no APOE ε4 allele, and 26.4% of the control population carried one or two APOE ε4 alleles. This confirms that the APOE ε4 allele is neither necessary nor sufficient for developing AD.
The incidence of AD increases by increasing age [36]. In our study, analysis of the interaction of APOE ε4 by age indicated that the effect of the APOE ε4 allele was weaker with increasing age. The frequency of the APOE ε4 allele decreased after 80 years in AD patients while the frequency of the APOE ε2 allele increased. This suggests that the reported increase in AD in individuals aged ≥ 80 years is likely due to genetic or environmental factors other than the APOE ε4 [36, 37]. Consequently, factors influencing the risk of AD in this age group may be interesting for further studies.
The APOE ε4 allele is the only known genetic risk factor for LOAD. The present study demonstrates that also in this Norwegian population the APOE ε4 allele is a strong risk factor for dementia, similar to what is seen in other Caucasian populations. The effect of two APOE ε4 alleles is stronger than of one, regarding both risk and age at onset. Because of the similarities seen with respect to genetic risk of APOE ε4 in this Norwegian and other Caucasian populations and with the likely increased genetic homogeneity due to population demographics, this Norwegian population may serve as an ideal population to search for additional genetic risk factors contributing to risk of developing of AD.
References
Mahley RW: Apolipoprotein E: cholesterol transport protein with expanding role in cell biology. Science. 1988, 240 (4852): 622-630. 10.1126/science.3283935.
Zannis VI, Breslow JL: Human very low density lipoprotein apolipoprotein E isoprotein polymorphism is explained by genetic variation and posttranslational modification. Biochemistry. 1981, 20 (4): 1033-1041. 10.1021/bi00507a059.
Gerdes LU, Klausen IC, Sihm I, Faergeman O: Apolipoprotein E polymorphism in a Danish population compared to findings in 45 other study populations around the world. Genet Epidemiol. 1992, 9 (3): 155-167. 10.1002/gepi.1370090302.
Corbo RM, Scacchi R: Apolipoprotein E (APOE) allele distribution in the world. Is APOE*4 a 'thrifty' allele?. Ann Hum Genet. 1999, 63 (Pt 4): 301-310. 10.1046/j.1469-1809.1999.6340301.x.
Singh PP, Singh M, Mastana SS: APOE distribution in world populations with new data from India and the UK. Ann Hum Biol. 2006, 33 (3): 279-308. 10.1080/03014460600594513.
Gerdes LU, Gerdes C, Hansen PS, Klausen IC, Faergeman O, Dyerberg J: The apolipoprotein E polymorphism in Greenland Inuit in its global perspective. Hum Genet. 1996, 98 (5): 546-550. 10.1007/s004390050257.
Strittmatter WJ, Saunders AM, Schmechel D, Pericak-Vance M, Enghild J, Salvesen GS, Roses AD: Apolipoprotein E: high-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer disease. Proc Natl Acad Sci U S A. 1993, 90 (5): 1977-1981. 10.1073/pnas.90.5.1977.
Saunders AM, Strittmatter WJ, Schmechel D, George-Hyslop PH, Pericak-Vance MA, Joo SH, Rosi BL, Gusella JF, Crapper-MacLachlan DR, Alberts MJ: Association of apolipoprotein E allele epsilon 4 with late-onset familial and sporadic Alzheimer's disease. Neurology. 1993, 43 (8): 1467-1472.
Okuizumi K, Onodera O, Tanaka H, Kobayashi H, Tsuji S, Takahashi H, Oyanagi K, Seki K, Tanaka M, Naruse S: ApoE-epsilon 4 and early-onset Alzheimer's. Nat Genet. 1994, 7 (1): 10-11. 10.1038/ng0594-10b.
van Duijn CM, de Knijff P, Cruts M, Wehnert A, Havekes LM, Hofman A, Van Broeckhoven C: Apolipoprotein E4 allele in a population-based study of early-onset Alzheimer's disease. Nat Genet. 1994, 7 (1): 74-78. 10.1038/ng0594-74.
Pearson JV, Huentelman MJ, Halperin RF, Tembe WD, Melquist S, Homer N, Brun M, Szelinger S, Coon KD, Zismann VL, Webster JA, Beach T, Sando SB, Aasly JO, Heun R, Jessen F, Kolsch H, Tsolaki M, Daniilidou M, Reiman EM, Papassotiropoulos A, Hutton ML, Stephan DA, Craig DW: Identification of the genetic basis for complex disorders by use of pooling-based genomewide single-nucleotide-polymorphism association studies. Am J Hum Genet. 2007, 80 (1): 126-139. 10.1086/510686.
Corder EH, Saunders AM, Strittmatter WJ, Schmechel DE, Gaskell PC, Small GW, Roses AD, Haines JL, Pericak-Vance MA: Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer's disease in late onset families. Science. 1993, 261 (5123): 921-923. 10.1126/science.8346443.
Locke PA, Conneally PM, Tanzi RE, Gusella JF, Haines JL: Apolipoprotein E4 allele and Alzheimer disease: examination of allelic association and effect on age at onset in both early- and late-onset cases. Genet Epidemiol. 1995, 12 (1): 83-92. 10.1002/gepi.1370120108.
Poirier J, Davignon J, Bouthillier D, Kogan S, Bertrand P, Gauthier S: Apolipoprotein E polymorphism and Alzheimer's disease. Lancet. 1993, 342 (8873): 697-699. 10.1016/0140-6736(93)91705-Q.
Benjamin R, Leake A, McArthur FK, Candy JM, Ince PG, Edwardson JA, Torvik A, Morris CM, Bjertness E: Apolipoprotein E genotype and Alzheimer's disease in an elderly Norwegian cohort. Neurodegeneration. 1996, 5 (1): 43-47. 10.1006/neur.1996.0006.
Johannsen P, Jakobsen JK, Jørgensen AL: [Apolipoprotein E genotypes in patients investigated for dementia]. Ugeskr Laeger. 1997, 159 (38): 5658-5661.
Bergem AL, Lannfelt L: Apolipoprotein E type epsilon4 allele, heritability and age at onset in twins with Alzheimer disease and vascular dementia. Clin Genet. 1997, 52 (5): 408-413.
Qiu C, Kivipelto M, Agüero-Torres H, Winblad B, Fratiglioni L: Risk and protective effects of the APOE gene towards Alzheimer's disease in the Kungsholmen project: variation by age and sex. J Neurol Neurosurg Psychiatry. 2004, 75 (6): 828-833. 10.1136/jnnp.2003.021493.
Huang W, Qiu C, von Strauss E, Winblad B, Fratiglioni L: APOE genotype, family history of dementia, and Alzheimer disease risk: a 6-year follow-up study. Arch Neurol. 2004, 61 (12): 1930-1934. 10.1001/archneur.61.12.1930.
Folstein MF, Folstein SE, McHugh PR: "Mini-Mental State": A practical method for grading the cognitive state of patients for clinician. J Psychol Res. 1975, 12 (3): 189-198. 10.1016/0022-3956(75)90026-6.
Brodaty H, Moore CM: The Clock Drawing Test for dementia of the Alzheimer's type: A comparison of three scoring methods in a memory disorders clinic. Int J Geriatr Psychiat. 1997, 12 (6): 619-627. 10.1002/(SICI)1099-1166(199706)12:6<619::AID-GPS554>3.0.CO;2-H.
Kjelvik G, Sando SB, Aasly J, Engedal KA, White LR: Use of the Brief Smell Identification Test for olfactory deficit in a Norwegian population with Alzheimer's disease. Int J Geriatr Psychiat. 2007, 22 (10): 1020-1024. 10.1002/gps.1783.
McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan EM: Clinical diagnosis of Alzheimer's disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer's Disease. Neurology. 1984, 34 (7): 939-944.
Henderson JN, Crook R, Crook J, Hardy J, Onstead L, Carson-Henderson L, Mayer P, Parker B, Petersen R, Williams B: Apolipoprotein E4 and tau allele frequencies among Choctaw Indians. Neurosci Lett. 2002, 324 (1): 77-79. 10.1016/S0304-3940(02)00150-7.
Farrer LA, Cupples LA, Haines JL, Hyman B, Kukull WA, Mayeux R, Myers RH, Pericak-Vance MA, Risch N, van Duijn CM: Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA. 1997, 278 (16): 1349-1356. 10.1001/jama.278.16.1349.
Bickeböller H, Campion D, Brice A, Amouyel P, Hannequin D, Didierjean O, Penet C, Martin C, Pérez-Tur J, Michon A, Dubois B, Ledoze F, Thomas-Anterion C, Pasquier F, Puel M, Demonet JF, Moreaud O, Babron MC, Meulien D, Guez D, Chartier-Harlin MC, Frebourg T, Agid Y, Martinez M, Clerget-Darpoux F: Apolipoprotein E and Alzheimer disease: genotype-specific risks by age and sex. Am J Hum Genet. 1997, 60 (2): 439-446.
Ashford JW, Mortimer JA: Non-familial Alzheimer's disease is mainly due to genetic factors. J Alzheimers Dis. 2002, 4 (3): 169-177.
Kumar T, Liestøl K, Maehlen J, Hiorth A, Jettestuen E, Lind H, Brorson SH: Allele frequencies of apolipoprotein E gene polymorphisms in the protein coding region and promoter region (-491A/T) in a healthy Norwegian population. Hum Biol. 2002, 74 (1): 137-142. 10.1353/hub.2002.0006.
Davidson Y, Gibbons L, Pritchard A, Hardicre J, Wren J, Stopford C, Julien C, Thompson J, Payton A, Pickering-Brown SM, Pendleton N, Horan MA, Burns A, Purandare N, Lendon CL, Neary D, Snowden JS, Mann DM: Apolipoprotein E epsilon4 allele frequency and age at onset of Alzheimer's disease. Dement Geriatr Cogn Disord. 2007, 23 (1): 60-66. 10.1159/000097038.
Sager MA, Hermann B, La Rue A: Middle-aged children of persons with Alzheimer's disease: APOE genotypes and cognitive function in the Wisconsin Registry for Alzheimer's Prevention. J Geriatr Psychiatry Neurol. 2005, 18 (4): 245-249. 10.1177/0891988705281882.
Olarte L, Schupf N, Lee JH, Tang MX, Santana V, Williamson J, Maramreddy P, Tycko B, Mayeux R: Apolipoprotein E epsilon4 and age at onset of sporadic and familial Alzheimer disease in Caribbean Hispanics. Arch Neurol. 2006, 63 (11): 1586-1590. 10.1001/archneur.63.11.1586.
Lehtovirta M, Helisalmi S, Mannermaa A, Soininen H, Koivisto K, Ryynänen M, Riekkinen P: Apolipoprotein E polymorphism and Alzheimer's disease in eastern Finland. Neurosci Lett. 1995, 185 (1): 13-15. 10.1016/0304-3940(94)11213-3.
Myers RH, Schaefer EJ, Wilson PW, D'Agostino R, Ordovas JM, Espino A, Au R, White RF, Knoefel JE, Cobb JL, McNulty KA, Beiser A, Wolf PA: Apolipoprotein E epsilon4 association with dementia in a population-based study: The Framingham study. Neurology. 1996, 46 (3): 673-677.
Raber J, Huang Y, Ashford JW: ApoE genotype accounts for the vast majority of AD risk and AD pathology. Neurobiol Aging. 2004, 25 (5): 641-650. 10.1016/j.neurobiolaging.2003.12.023.
Hyman BT, Hedley-Whyte ET, Rebeck GW, Vonsattel JP, West HL, Growdon JH: Apolipoprotein E epsilon4/4 in a neuropathologically normal very elderly individual. Arch Neurol. 1996, 53 (3): 215-
Launer LJ, Andersen K, Dewey ME, Letenneur L, Ott A, Amaducci LA, Brayne C, Copeland JR, Dartigues JF, Kragh-Sorensen P, Lobo A, Martinez-Lage JM, Stijnen T, Hofman A: Rates and risk factors for dementia and Alzheimer's disease: results from EURODEM pooled analyses. EURODEM Incidence Research Group and Work Groups. European Studies of Dementia. Neurology. 1999, 52 (1): 78-84.
Ott A, Breteler MM, van Harskamp F, Claus JJ, van der Cammen TJ, Grobbee DE, Hofman A: Prevalence of Alzheimer's disease and vascular dementia: association with education. The Rotterdam study. Bmj. 1995, 310 (6985): 970-973.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2377/8/9/prepub
Acknowledgements
The authors are grateful to patients and their relatives for participation in this study. We are grateful to Dr. Wenche Frogn Sellæg for kindly help with recruitment of patients from Namsos hospital. The study was supported by the Research Council of Norway (153487/V50; SBS held a three year PhD scholarship) and the National Institutes of Health, NSRA fellowship #AG 24030 to SM and National Institute of Aging grant PO1 AG17216 and the Mayo Foundation to MH. The funding bodies did not influence the content of the study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author(s) declare that they have no competing interests.
Authors' contributions
SBS has done the clinical and neurological evaluation of all included patients, recruited the control individuals, done some of the statistics and drafted the manuscript. AC has carried out genotyping of the samples. SM has been involved in conception and design of the study, carried out genotyping of the samples, and has revised the manuscript. IS and OS have made substantial contribution to conception and design and have recruited patients for the study. SL has made substantial work with analysing and interpretation of the data, and has been involved in revising the manuscript. JAa and LW have made important contribution to conception and design of the study and to drafting and revising the manuscript. MH has made substantial contribution to conception and design of the study. All authors have read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Sando, S.B., Melquist, S., Cannon, A. et al. APOE ε4 lowers age at onset and is a high risk factor for Alzheimer's disease; A case control study from central Norway. BMC Neurol 8, 9 (2008). https://doi.org/10.1186/1471-2377-8-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2377-8-9