
Abstract
Background
Recent reports suggest that vitamin B1 (thiamine) participates in the processes underlying plant adaptations to certain types of abiotic and biotic stress, mainly oxidative stress. Most of the genes coding for enzymes involved in thiamine biosynthesis in Arabidopsis thaliana have been identified. In our present study, we examined the expression of thiamine biosynthetic genes, of genes encoding thiamine diphosphate-dependent enzymes and the levels of thiamine compounds during the early (sensing) and late (adaptation) responses of Arabidopsis seedlings to oxidative, salinity and osmotic stress. The possible roles of plant hormones in the regulation of the thiamine contribution to stress responses were also explored.
Results
The expression of Arabidopsis genes involved in the thiamine diphosphate biosynthesis pathway, including that of THI1, THIC, TH1 and TPK, was analyzed for 48 h in seedlings subjected to NaCl or sorbitol treatment. These genes were found to be predominantly up-regulated in the early phase (2-6 h) of the stress response. The changes in these gene transcript levels were further found to correlate with increases in thiamine and its diphosphate ester content in seedlings, as well as with the enhancement of gene expression for enzymes which require thiamine diphosphate as a cofactor, mainly α-ketoglutarate dehydrogenase, pyruvate dehydrogenase and transketolase. In the case of the phytohormones including the salicylic, jasmonic and abscisic acids which are known to be involved in plant stress responses, only abscisic acid was found to significantly influence the expression of thiamine biosynthetic genes, the thiamine diphosphate levels, as well as the expression of genes coding for main thiamine diphosphate-dependent enzymes. Using Arabidopsis mutant plants defective in abscisic acid production, we demonstrate that this phytohormone is important in the regulation of THI1 and THIC gene expression during salt stress but that the regulatory mechanisms underlying the osmotic stress response are more complex.
Conclusions
On the basis of the obtained results and earlier reported data, a general model is proposed for the involvement of the biosynthesis of thiamine compounds and thiamine diphosphate-dependent enzymes in abiotic stress sensing and adaptation processes in plants. A possible regulatory role of abscisic acid in the stress sensing phase is also suggested by these data.
Similar content being viewed by others

Background
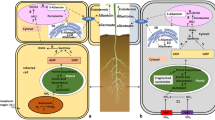
Vitamin B1 (thiamine) is essential for the proper functioning of all living organisms as it plays important roles in carbohydrate catabolism, NADPH and ATP synthesis and in the formation of nucleic acids. The biologically active form of thiamine is its diphosphate analog (TDP) which functions as a cofactor for a number of critical enzymes (Figure 1A) including pyruvate decarboxylase (PDC), pyruvate dehydrogenase (PDH), α-ketoglutarate dehydrogenase (KGDH), branched-chain α-ketoacid dehydrogenase (BKDH), transketolase (TK), acetolactate synthase (AHAS) and 1-deoxy-D-xylulose-5-phosphate synthase (DXPS) [1]. Plants can synthesize TDP de novo from simple precursors via biosynthetic pathways that bear the hallmarks of the systems utilized by both bacteria and yeast [2–5]. In this regard, the early stages of TDP biosynthesis include two parallel pathways. One is similar to the mechanism found in bacteria and leads from 5-aminoimidazole ribonucleotide (AIR) to the pyrimidine moiety of thiamine (4-amino-2-methyl-5-hydroxymethylpyrimidine monophosphate, HMP-P) and engages only a single enzyme, a product of the THIC gene that has been characterized in Arabidopsis thaliana [6]. In the second of the parallel pathways, the thiazole moiety of thiamine (4-methyl-5-(2-hydroxyethyl)-thiazole phosphate, HET-P) is synthesized from glycine, NAD+ and a sulfur donor protein, in a similar manner to the system utilized in yeast [7]. The major enzyme involved in the HET-P formation is encoded by the THI1 gene, which has now been characterized in both Arabidopsis and Zea mays [8, 9].

Schematic representation of thiamine biosynthesis in plants (A) and subcellular localization of the major thiamine diphosphate-incorporating enzymes (B). The Arabidopsis genes encoding the enzymes which catalyze the biosynthetic reactions shown are specified. AIR, 5-aminoimidazole ribonucleotide; HET-P, 4-methyl-5-(2-hydroxyethyl)thiazole phosphate; HMP-P/HMP-PP, 4-amino-2-methyl-5-hydroxymethylpyrimidine monophosphate/diphosphate; TA, thiamine; TMP, thiamine monophosphate; TDP, thiamine diphosphate; PDC, pyruvate decarboxylase; PDH, pyruvate dehydrogenase; KGDH, α-ketoglutarate dehydrogenase; BKDH, branched-chain α-ketoacid dehydrogenase; TK, transketolase; DXPS, 1-deoxy-D-xylulose-5-phosphate synthase.
Two further thiamine biosynthetic steps include an additional phosphorylation of HMP-P to 4-amino-2-methyl-5-hydroxymethylpyrimidine diphosphate (HMP-PP) and the condensation of HMP-PP with HET-P to form thiamine monophosphate (TMP). These two reactions are catalyzed by a single protein, the product of the TH1 gene in Arabidopsis [10] or THI3 gene in Z. mays [11], which is thus a bifunctional enzyme with both HMP-P kinase and TMP synthase activities. TMP does not possess any recognized physiological function to date but constitutes a thiamine resource or transitional stage for further TDP synthesis. In the latter case, an unphosphorylated form of thiamine is required. As has been shown previously in Z. mays [12], TMP is dephosphorylated by a relatively unspecific phosphatase which can also use TDP as a substrate. All of the steps described above proceed in chloroplasts [13], but the process of thiamine conversion to TDP occurs in the cytosol and is catalyzed by thiamine pyrophosphokinase, encoded by the TPK genes [14]. To fulfill its biological function, TDP must be transported to chloroplasts or mitochondria but the required transporters have not yet been identified. The thiamine biosynthesis pathway in plants is precisely regulated through TDP-dependent riboswitch mechanisms on the THIC gene and in ancient plant taxa also the THI1 gene levels [15].
It has been well documented that thiamine biosynthesis is activated during plant adaptation responses to persistent abiotic stress conditions such as salting and flooding [16], cold, heat and drought [17, 18], and oxidative stress [19, 20]. However, it has not yet been determined how thiamine biosynthesis and activation responds to the above stimuli at the early stages of the corresponding stress responses (sensing). In our current study, we further characterize the role of thiamine biosynthesis upregulation in the early response to oxidative, osmotic and salt stress in plants. In the experiments, we analyze changes in the expression levels of all thiamine-biosynthetic genes and of genes encoding the major TDP-dependent enzymes such as TK, KGDH, PDH, PDC and DXPS in Arabidopsis seedlings. We also show for the first time that abscisic acid (ABA), but not salicylic acid (SA), is involved in the upregulation of thiamine biosynthesis in the plant response to abiotic stress.
Results
Increased expression of thiamine biosynthesis genes during early response of Arabidopsis seedlings to oxidative, salt and osmotic stress
The expression of the four main genes involved in thiamine biosynthesis in A. thaliana was analyzed during different phases of the responses (early sensing and later adaptation) to oxidative (0.25 μM paraquat; PQ), salt (200 mM NaCl) and osmotic (200 mM sorbitol; SOR) stress conditions. These tests assessed the genes responsible for the biosynthesis of the pyrimidine and thiazole moieties of thiamine (THIC and THI1, respectively), the coupling of HET-P and HMP-PP to form TMP (TH1), and the activation of thiamine to TDP (TPK). A fast sensing response (within 2 h) by Arabidopsis seedlings to environmental changes (Figure 2) was observed through increases in the expression of two genes involved in early steps of thiamine biosynthesis, THI1 and THIC (2.9- and 2.2-fold increases, respectively) in the presence of high concentrations of salt or SOR, but not PQ. In the case of a longer stress duration of up to 48 h, the higher expression of THIC in the presence of SOR was similar (2-fold higher expression as compared with unstressed plants) but was found to be dramatically decreased under persistent high salt conditions. The stress-induced changes in the expression of THI1 showed a similar time-dependence, with the notable exception of osmotic stress for 48 h during which time the expression of this gene decreased in contrast to the response of the THIC gene. The expression of TH1 was also altered by these stress conditions with a time profile similar to that of THIC but with increases of no more than 1.5-fold. The highest differential expression response (1.5- to 3.0-fold) of the TPK gene was observed after 2-4 h from the application of the stress stimulus (SOR or NaCl) to the growth medium.

Time-dependent response of Arabidopsis seedlings to oxidative, salt and osmotic stress revealed through thiamine biosynthesis gene expression changes. Nine day old seedlings of A. thaliana ecotype Columbia were transferred onto MS medium supplemented with 0.2 μM paraquat (PQ), 200 mM NaCl or 200 mM sorbitol (SOR) for the indicated times. Transcripts of the THI1 (coding for HET-P synthase), THIC (coding for HMP-P synthase), TH1 (encoding thiamine monophosphate synthase/HMP-P kinase bifunctional enzyme) and TPK (coding for thiamine pyrophosphokinase) genes were quantified in seedling leaves by RT-PCR and expressed relative to the corresponding gene expression levels in the control seedlings grown under stress-free conditions. The asterisk indicates a statistically significant difference when compared with the control samples (t-test, P < 0.05).
Supplemental experiments were performed that investigated changes in the overall thiamine contents in the leaves of Arabidopsis seedlings grown under stress conditions for 48 h (Figure 3). The total thiamine levels, i.e. the sum of the thiamine, TMP and TDP contents, were found to increase in the seedlings under stress conditions by 160-170%. Whilst TDP predominated over the other thiamine compounds under all of the tested conditions, its level increased by only 120-130% during the plant adaptation stress response. Hence, the observed increase in the total thiamine level is principally due to the large increases in TMP and free thiamine (by 200-380%, as compared with unstressed plants).

Changes in the levels of thiamine and its phosphate esters in Arabidopsis seedlings in response to abiotic stress conditions. Nine day old seedlings were maintained for 48 h on growth medium supplemented with 0.2 μM PQ, 200 mM NaCl or 200 mM SOR. Thiamine compounds were assayed in seedling leaves by RP-HPLC via post-column oxidation to thiochrome derivatives and fluorometric detection. The asterisk indicates a statistically significant difference when compared with the control samples (t-test, P < 0.05).
Early response of TDP-dependent enzymes to oxidative, salt and osmotic stress
In an attempt to correlate the stress-induced increases in the expression of plant genes involved in thiamine biosynthesis with the actual requirement of the plant for TDP under different stress conditions, we analyzed the expression level of several genes encoding TDP-dependent enzymes that play critical roles in cellular metabolism (KGDH, PDH, TK and DXPS). In these experiments, KGDH appeared to be most sensitive to environmental changes. In the case of this enzyme, a 2 to 3-fold increase in expression was observed by 2 h (salt) or 6 h (SOR) after exposure to stress conditions (Figure 4). Over a longer time scale (24 h), this expression returned rapidly to a basal level. The upregulation of TK genes (about 2-fold) was also observed with a similar time-dependence under salt stress, but in the presence of SOR this expression increase was prolonged for 24 h. PDH genes were also found to be involved in salt and SOR stress sensing. However, the expression increases in these genes (about 2-fold) were highest after 6 h under both stress conditions. In contrast, only a marginal (1.2-fold) increase in DXPS gene expression was observed after 6 h of salt or SOR treatment, and these levels then remained constant for 24 h.

Time-dependent changes in the expression of TDP-dependent enzyme genes: E1 components of α-ketoglutarate dehydrogenase (KGDH) and pyruvate dehydrogenase (PDH) complexes, transketolase (TK) and 1-deoxy-D-xylulose-5-phosphate synthase (DXPS). Gene expression was determined by RT-PCR analysis of the leaves of nine day-old seedlings maintained for the specified time on medium supplemented with PQ, NaCl or SOR or without any stressor (C). The asterisk indicates a statistically significant difference when compared with the control (C) samples (t-test, P < 0.05).
Involvement of plant hormones in the up-regulation of thiamine biosynthesis during early response to stress
The results of control analyses of thiamine pools in Arabidopsis seedlings grown for 48 h on standard medium supplemented with plant growth regulators, including ABA, jasmonic acid (JA) and SA, are presented in Figure 5A. The total thiamine content was markedly increased in the presence of ABA (360% of the control levels), but was much less enhanced (140%) by JA and did not exceed control levels in the presence of SA. TMP, produced de novo under the influence of ABA, was found to accumulate mostly in a free thiamine pool (8.8-fold increase over the control), probably owing to the activation of unspecific phosphatases. However, the TDP level was essentially maintained as only a 1.5-fold increase was observed (Figure 5B). These results were further confirmed by determinations of the thiamine biosynthetic gene transcript levels after 6 h following ABA supplementation (Figure 6). Remarkably, the expression of all analyzed genes increased in seedlings grown in ABA-containing medium: 2-to 2.2-fold increases were noted for the THI1, THIC and TPK genes and a slightly lower (1.5-fold) upregulation was observed for the TH1 gene. In contrast, over the same time scale, the presence of SA in the medium did not affect the thiamine biosynthesis processes and only a slightly increased expression (1.2- to 1.5-fold) of THI1, THIC and TH1, but not TPK, was observed after supplementation of the medium with JA.

Changes in the levels of total thiamine (A) and of individual thiamine compounds (B) in the leaves of Arabidopsis seedlings treated for 48 h with various plant hormones. Seedlings were treated with 100 μM abscisic acid (ABA), 100 μM jasmonic acid (JA) or 200 μM salicylic acid (SA) and assayed for thiamine compounds as described in Figure 3. Seedlings maintained on growth medium without any added hormone served as the control (C). The asterisk indicates a statistically significant difference when compared with the control samples (t-test, P < 0.05).

Effects of plant hormones on the expression of thiamine biosynthesis genes in Arabidopsis seedlings. Nine day old seedlings were kept for 6 h on growth medium supplemented with 100 μM ABA, 100 μM JA or 200 μM SA. Gene expression in seedling leaves was analyzed by RT-PCR. The asterisk indicates a statistically significant difference when compared with the control samples (t-test, P < 0.05).
An analysis of the effects of medium supplementation with ABA on the transcription of TDP-dependent enzymes (Figure 7) revealed KGDH and TK (2.6- and 2.0-fold expression increases, respectively) as the enzymes likely to be regulated by ABA. However, we did not observe any significant influence of ABA on PDH expression in our present experiments.

Effects of ABA on the expression of genes coding for TDP-dependent enzymes in Arabidopsis seedlings. Nine day old seedlings were maintained for the specified times on medium supplemented with 100 μM ABA. Gene expression in seedling leaves was analyzed by RT-PCR. The asterisk indicates a statistically significant difference when compared with the control samples (t-test, P < 0.05).
To further confirm the involvement of ABA in the upregulation of thiamine biosynthesis during water stress sensing, a mutant defective in ABA biosynthesis (aba1) was analyzed. The expression of thiamine biosynthesis genes in aba1 mutant plants treated with SOR showed a similar pattern to that in the wild type plants (Figure 8), with the only noted difference being that the response was slower in the mutant plants (6 h versus 2 h in the wild plants). In contrast, under salt treatment, the THI1 and THIC genes were not significantly affected in this mutant plant and TH1 and TPK expression showed a wild-type pattern.

Changes in the expression of thiamine biosynthesis genes in the seedlings of an ABA-defective Arabidopsis mutant subjected to salt or osmotic stress or treated with exogenous ABA. Nine day old seedlings of the aba1 mutant of A. thaliana (ecotype Landsberg erecta) or of the corresponding wild-type plant (C) were maintained for the indicated time on medium supplemented with NaCl, SOR or ABA. Gene expression in the seedling leaves was analyzed by RT-PCR. The asterisk indicates a statistically significant difference when compared with the control samples (t-test, P < 0.05).
Discussion
Because their growth and development are affected by environmental fluctuations, plants require fast responses to these changes for the maintenance of internal homeostasis. Hence, a number of sensitive mechanisms have evolved in plants to allow rapid sensing of environmental cues and facilitate growth modifications for adaptation and survival [21, 22]. As stress has an impact on the carbon fixation and overall energy status of plants, most previous genetic, proteomic and metabolic studies of this area have dealt with changes in glycolysis, the tricarboxylic acid cycle, the pentose phosphate pathway and photosynthesis [23–25]. However, these earlier reports have presented data that was obtained under very different growth or stress conditions, and also over quite different time scales and using different measurement parameter. This makes it difficult to compare the results from different laboratories in a meaningful way.
In our current study, which for the first time evaluates the early response stage ("stress sensing") rather than the long-term adaptation to stress in plants, we compared (i) the expression levels of the genes involved in thiamine/TDP biosynthesis pathways; (ii) the expression levels of genes coding for the most important TDP-requiring enzymes; and (iii) the thiamine and its phosphate ester contents, in Arabidopsis seedlings treated with abiotic stressors including PQ, NaCl and SOR. The application of NaCl or SOR into the plant growth media during the first few hours of treatment is sensed by the plants as general water stress rather than as salt- or SOR-specific effects [26]. It is generally accepted, that stress-generated damage involves the most important pathways and hence an obvious need to upregulate the main biosynthetic processes, including the biosynthesis of TDP. Indeed, we observed in our present experiments that the expression of all TDP-biosynthesis genes had already been increased at 2 h after the application of two water-stress agents (Figure 2). Interestingly, these genes showed decreased expression under salt stress over a longer time scale of several days, suggesting an extensive ion penetration into plant tissues [26] that could possibly generate new stimuli and adaptation processes. However, in our SOR stress experiments, the most of the early observed changes, particularly those involving the THIC gene, were maintained over a long time scale. This may indicate that a hormonal signaling mechanism rather than water availability controls plant growth under these conditions [27]. Surprisingly, we did not observe any marked influence of PQ on the expression of TDP biosynthesis genes over a time scale of a few hours, which is in contrast to the findings of an earlier study of long-lasting PQ-generated oxidative stress [20].
Our observed increases in gene expression during initial stages of the plant stress responses may indicate that (i) the translated proteins are susceptible to damage under unfavorable conditions and an increase of transcription is therefore required to maintain the proper thiamine cellular levels; (ii) the TDP-dependent enzymes are destroyed and, parallel to their re-synthesis, a new requirement for their cofactors emerges; and/or (iii) thiamine production is necessary to overcome the consequences of stress or to develop new adaptation strategies. Our analysis of thiamine and its phosphate analogs in stressed plants (Figure 3) showed that the observed changes in the expression of thiamine biosynthetic genes correlated with the activity of the encoded enzymes. The increase in the thiamine and TDP contents during the early Arabidopsis response (sensing) to water stress is consistent with similar changes recently reported in Z. mays seedlings subjected to long-lasting osmotic, oxidative and salinity stress [19]. This suggests that during stress sensing and adaptation in plants, the biosynthesis of thiamine is tightly regulated. Moreover, this regulation does not seem to be specific to the type of stress and could benefit from two different mechanisms previously identified in plants. The first type of possible regulation occurs at the genetic level and can apply to the THIC gene promoter as it contains a TDP-dependent riboswitch [15]. The other regulation process can operate at the protein level, as was demonstrated in a study of Z. mays, where TMP synthase activity was strongly inhibited by an increased supply of HMP-PP, produced through the activity of the kinase domain of the same protein, THI3 [11].
To test the hypothesis that the effects of stress conditions on thiamine compound pools in plant tissues arise from the increased requirement for TDP from the TDP-dependent main catabolic pathways, we analyzed the stress-dependent changes in the expression of several genes encoding TDP-dependent enzymes (Figure 4). The mitochondrial production of acetyl CoA and the tricarboxylic acid cycle are assumed to be the first plant sensors of oxidative stress [23, 28, 29]. Our current analyses identified KGDH and PDH as the main source of stress perturbation. However, the upregulation of the expression of these two mitochondrial TDP-dependent enzymes can result not only from the increased requirement for mitochondrial potential restoration, respiration control, nitrogen metabolism or glutamate signaling [30] but also reflect the activation of a stress signaling response to a metabolic imbalance as a result of a KGDH side reaction resulting in reactive oxygen species production [29].
The TK enzyme, which operates in the chloroplastic Calvin-Benson cycle (CBC), is involved in carbon fixation [31] and its activity is a limiting factor for sucrose production and photosynthesis [32]. As it functions also in the pentose phosphate pathway, TK is additionally important for the generation of NADPH, cooperating with a variety of oxidant-scavenging systems [23, 33–35]. The early reported increase in TK transcripts in Arabidopsis agrees with the changes in TK activity observed in Z. mays under salt and oxidative stress [19] and supports the hypothesis that this enzyme is involved in plant stress protection. DXPS is another TDP-dependent enzyme which is involved in the production of powerful antioxidants, the carotenoids [36, 37]. The activation of carotenoid synthetic pathways has been observed during osmotic, salt and drought stress [38–40]. In our present study, however, no significant changes in the expression of DXPS-encoding gene were observed over 24 h of stress treatment, possibly because its regulation occurs at the posttranscriptional level [41].
The well characterized and essential roles of the phytohormones in a wide range of stress adaptive processes in plants [42, 43] prompted us to search for candidate regulators of the activation of thiamine biosynthesis and TDP-dependent metabolic processes during salt and osmotic stress sensing and in the later adaptation phase. Our focus on SA was based on previous reports of an activation of a cabbage THIJ-like gene product involved in HMP phosphorylation via the SA-dependent signaling pathway [44] and an involvement of SA in thiamine-induced plant resistance to pathogen attacks [45]. However, our treatments of Arabidopsis plants with exogenous SA did not cause any observable changes in expression of genes involved in thiamine biosynthesis (Figures 5 and 6).
JA is associated with the defense against necrotrophic pathogens and herbivorous insects [46] but its involvement in salt stress-dependent gene regulation has also been suggested [47]. In our current study, supplementation of growth medium with JA caused a slightly increased thiamine content in Arabidopsis seedlings, resulting from the activation of thiamine biosynthetic genes, mostly TH1 (Figures 5 and 6). Although the mechanism underlying thiamine biosynthesis regulation by JA requires further analysis, it is noteworthy that a cis-acting element T/GBOXATPIN2 (AACGTG) was identified in the TH1 promoter region [48] which is responsible for activating this gene in response to JA upon wounding [49].
ABA regulates nearly 10% of the protein-coding genes in plants [50] and commonly participates in the transcriptional regulation processes that operate under salt and osmotic stress conditions [51–54]. The upregulation of the THI1, THIC, TH1 and TPK genes and the 3.5-fold increase in the total thiamine levels in Arabidopsis seedlings under the influence of an external ABA supply that we observed in this study provide the first evidence for the involvement of ABA in thiamine biosynthesis under stress conditions (Figures 5, 6). These findings are also interpretable in the light of previous analyses of THI1and THIC promoters that showed the presence of an ABA-responsive element (ABRE) in the THI1 promoter [16] and putative stress-related elements in the THIC promoter, including ABRE and drought-response element [20]. In silico analysis of the TPK promoter has identified a dehydration and ABA-inducible expression element MYBATRD22 (CTAACCA) and of the TH1 promoter an ABRE - ABRELATERD1 (ACGTG) element.
Our current analysis of the expression of TDP-dependent enzymes in seedlings grown on ABA-containing medium showed a transcript accumulation for KGDH- and TK-encoding genes but no significant effects on PDH gene transcripts. These data are consistent with our previous observations of the increased ABA content and TK activity in Z. mays seedlings treated with NaCl and polyethylene glycol [19]. Taken together, these data are consistent with the well established hypothesis regarding the linkages between ABA and plant mitochondrial functionality [55–57].
Although the application of exogenous ABA has often been used to mimic water stress responses in plants [58], it is generally accepted that both ABA-dependent and ABA-independent processes are involved in these types of responses [59]. In an attempt to further validate the hypothesis that ABA is responsible for thiamine biosynthesis in the early response (sensing) of plants to water stress generated by salt or SOR, a mutant Arabidopsis plant (aba1) was employed in our present experiments. This mutant is defective in the production of zeaxanthin epoxidase, an enzyme that catalyzes two early steps in the generation of the epoxycarotenoid precursor as part of the ABA biosynthetic pathway [60]. In the analysis, aba1 showed different behaviors in response to salt and osmotic stress. Under salt stress, a decrease in the THI1 and THIC expression levels in the aba1 mutant plants was observed, confirming the regulation of these thiamine biosynthetic steps by ABA. The expression of TH1 and TPK was found to be maintained as in the wild type plant. In contrast, in the presence of SOR the activation of thiamine biosynthetic genes was found not to be markedly different from wild type plants, suggesting that some additional, e.g. post-transcriptional, regulatory processes for thiamine biosynthesis operate under these conditions, although the details remain to be clarified.
Our present findings suggest a contribution of ABA to the transcriptional regulation of the two first genes that function in thiamine biosynthesis (THI1 and THIC) in Arabidopsis seedlings subjected to salt and osmotic stress, particularly at the early stages of the stress response (i.e. stress sensing). However, other as yet unidentified endogenous mediators must also be involved and the relative importance of ABA becomes less significant over a longer time scale (i.e. during the plant adaptation to stress). Moreover, the TH1 and TPK genes appear to be regulated by endogenous factors other than ABA, but also by ABA when it is present in the growth medium.
On the basis of the data obtained in this study as well as the results of previous reports, a role for thiamine biosynthesis and TDP-dependent metabolic pathways in abiotic stress sensing and adaptation can be hypothesized (Figure 9). Briefly, abiotic stressors which appear in the plant environment begin to inactivate the major metabolic pathways, the crucial enzymes of which can act as stress sensing centers (i.e. KGDH, TK). The urgent requirement for the rapid reactivation of these factors generates the secondary processes or stimuli that initiate a further adaptation phase. The activation of TDP biosynthesis processes leads to thiamine/TDP overproduction, which supports defense responses such as NADPH production or carotenoid biosynthesis. TDP production and the subsequent activities of TDP-dependent enzymes come under ABA control but is not the sole regulator of these processes. However, thiamine compounds themselves can also act as a significant source of antioxidants [61, 62] or play a stress alarmone function as has been suggested in bacteria under carbon starvation conditions [63].

A proposed role of TDP biosynthesis and TDP-dependent enzymes in stress sensing and adaptation in plants. Environmental stress damages the main TDP-dependent enzymes in plants (TK, PDH, KGDH, DXPS) causing them to function as sensors of these stimuli. The further propagation of stress sensing processes activates the expression of thiamine/TDP biosynthetic enzymes (THI1, THIC, TH1 and TPK) that are necessary for the regeneration of the TDP-dependent pathways such as the Calvin-Benson cycle (CBC), the pentose phosphate pathway (PPP), the tricarboxylic acid cycle (TCAC), and the isoprenoid phosphate biosynthesis pathway (IPP). This in turn facilitates the activation of certain defense mechanisms and the production of stress protecting molecules including thiamine which can function as an antioxidant or as a stress alarmone.
Conclusions
We here demonstrate for the first time that TDP biosynthesis processes are quickly activated during the early phase (sensing) of the Arabidopsis response to salt and osmotic stress. The produced TDP is incorporated into TDP-dependent enzymes that are up-regulated at the same time such as KGDH, PDH, TK, all of which are involved in the main metabolic pathways that respond to stress conditions in plants. ABA seems to be involved in the activation of genes encoding enzymes that function in thiamine and TDP biosynthesis as well as the TDP-dependent KGDH and TK genes. We propose a working model for the contribution of thiamine to the stress sensing and adaptation processes in plants.
Materials and methods
Plant materials, growth conditions and stress treatments
The seeds of Arabidopsis thaliana ecotype Columbia (Lehle, Round Rock, TX, USA) were used for all gene expression analysis performed for the detection of thiamine biosynthesis and activation processes. The seeds of the ABA-deficient mutant aba1 (CS21) were obtained from the Arabidopsis Biological Resource Center (ABRC, Columbus, OH, USA). For comparative analyses using this mutant, wild-type Arabidopsis plants of the background ecotype Landsberg erecta were used.
Arabidopsis seeds (100 mg) were surface sterilized by treatment for 5 min with 0.5% Tween-20, for 5 min with 70% ethanol and for 7 min with a 15% bleach solution. Water-rinsed seeds were stratified for three days at 4°C and then grown on Murashige and Skoog (MS) agar plates supplemented with 1% sucrose but depleted of thiamine, under a 12 h light/12 h dark cycle at 22°C, with a constant light intensity of 100 μmol m-2 s-1. Nine-day old seedlings were then transferred and grown for 2-48 h on MS media containing 0.2 μM PQ, 200 mM NaCl, 200 mM SOR, 100 μM ABA, 100 μM JA or 200 μM SA. All experiments were performed with 3-5 replicates of 20 seedlings per treatment.
RNA isolation and quantitative real-time PCR
Total RNAs were isolated from liquid nitrogen-disrupted seedling leaves using Trizol reagent (Invitrogen, Carlsbad, CA, USA), and by treatment with RNase-free DNase I (Promega, Madison, WI, USA). Further purification was performed using a PureLink RNA Mini Kit (Invitrogen). The RNA concentrations were determined using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc., Wilmington, DE, USA) and the integrity of the RNA was assessed on a 1% (w/v) agarose gel. cDNAs were generated from 1 μg of total RNA using Invitrogen SuperScript III Reverse Transcriptase, as recommended by the manufacturer. Quantitative RT-PCR was carried out using an Rotor-Gene 6,000 system (Corbett Life Science, USA) and SYBR Green JumpStart Taq Ready Mix, in accordance with the manufacturer's instructions (Sigma, St. Louis, MO, USA). The gene-specific primer pairs, listed in Table 1 were designed by using a Primer Express 3.0 software (Applied Biosystems, Bedford, MA, USA)). The specificity of the reactions was verified by melting curve analysis. The relative mRNA level for each gene was calculated using ΔΔCT values in comparison with unstressed seedlings and the EF-1 gene was used as an internal control for normalization. Quantitative RT-PCR analysis was performed using at least three replicates of each cDNA sample.
HPLC analysis of thiamine and its phosphate analogs
Assays of thiamine and its phosphate analogs in Arabidopsis seedlings were carried out using a previously described method [19]. Briefly, plant extracts were prepared by 12% TCA treatment of frozen, ground Arabidopsis seedlings for 5 min at 4°C. After removal of the protein pellet and TCA disposal by ethyl ether treatment, the supernatant was analyzed by reverse-phase high performance liquid chromatography (RP-HPLC) with post-column oxidation using 90 μM sodium hexacyanoferrate (III) in 0.56 M NaOH and fluorescence detection of the formed thiochrome and thiochrome phosphates [64]. For RP-HPLC separation, gradient elution (0-90%B, 16 min) was used, where solvent A contained 15 mM ammonium citrate (pH 4.2) and solvent B consisted of 0.1 M formic acid and 55 mM diethylamine. The applied HPLC equipment contained a Shimadzu (Kyoto, Japan) model LC-9A HPLC pump with Shimadzu FCV-AL proportioning valve, a Knauer (Bad Homburg, Germany) model A0263 manual injector, a Merck cartridge LiChrosphere 100RP-18 column (250 × 3.4 mm), a Shimadzu model RF-535 fluorescence monitor (excitation at 365 nm, emission at 430 nm) and Shimadzu Class-VP software (version 4) for pump control, data acquisition and analysis.
References
Frank RA, Leeper FJ, Luisi BF: Structure, mechanism and catalytic duality of thiamine-dependent enzymes. Cell Mol Life Sci. 2007, 64: 892-905. 10.1007/s00018-007-6423-5.
Jurgenson CT, Begley TP, Ealick SE: The structural and biochemical foundations of thiamin biosynthesis. Ann Rev Biochem. 2009, 78: 569-603. 10.1146/annurev.biochem.78.072407.102340.
Kowalska E, Kozik A: The genes and enzymes involved in the biosynthesis of thiamin and thiamin diphosphate in yeasts. Cell Mol Biol Lett. 2008, 13: 271-282. 10.2478/s11658-007-0055-5.
Goyer A: Thiamine in plants: aspects of its metabolism and functions. Phytochemistry. 2010, 71: 1615-1624. 10.1016/j.phytochem.2010.06.022.
Rapala-Kozik M: Vitamine B1 (thiamine): a cofactor for enzymes involved in the main metabolic pathways and an environmental stress protectant. Adv Bot Res. 2011, 58: 37-90.
Raschke M, Burkle L, Muller N, Nunes-Nesi A, Fernie AR, Arigoni D, Amrhein N, Fitzpatrick TB: Vitamin B1 biosynthesis in plants requires the essential iron sulfur cluster protein, THIC. Proc Natl Acad Sci USA. 2007, 104: 19637-19642. 10.1073/pnas.0709597104.
Chatterjee A, Schroeder FC, Jurgenson CT, Ealick SE, Begley TP: Biosynthesis of the thiamin-thiazole in eukaryotes: identification of a thiazole tautomer intermediate. J Am Chel Soc. 2008, 130: 1394-11398.
Machado CR, de Oliveira RL, Boiteux S, Praekelt UM, Meacock PA, Menck CF: Thi1, a thiamine biosynthetic gene in Arabidopsis thaliana, complements bacterial defects in DNA repair. Plant Mol Biol. 1996, 31: 585-593. 10.1007/BF00042231.
Belanger F, Leustek T, Chu B, Kirz A: Evidence for the thiamine biosynthetic pathway in higher plant plastids and its developmental regulation. Plant Mol Biol. 1995, 29: 809-821. 10.1007/BF00041170.
Ajjawi I, Tsegaye Y, Shintani D: Determination of the genetic, molecular, and biochemical basis of the Arabidopsis thaliana thiamin auxotroph th1. Arch Biochem Biophys. 2007, 459: 107-114. 10.1016/j.abb.2006.11.011.
Rapala-Kozik M, Olczak M, Ostrowska K, Starosta A, Kozik A: Molecular characterization of the thi3 gene involved in thiamine biosynthesis in Zea mays: cDNA sequence and enzymatic and structural properties of the recombinant bifunctional protein with 4-amino-5-hydroxymethyl-2-methylpyrimidine (phosphate) kinase and thiamine monophosphate synthase activities. Biochem J. 2007, 408: 149-159. 10.1042/BJ20070677.
Rapala-Kozik M, Golda A, Kujda M: Enzymes that control the thiamine diphosphate pool in plant tissues. Properties of thiamine pyrophosphokinase and thiamine-(di)phosphate phosphatase purified from Zea mays seedlings. Plant Physiol Biochem. 2009, 47: 237-242. 10.1016/j.plaphy.2008.12.015.
Julliard JH, Douce R: Biosynthesis of the thiazole moiety of thiamin (vitamin B1) in higher plant chloroplasts. P Natl Acad Sci USA. 1991, 88: 2042-2045. 10.1073/pnas.88.6.2042.
Ajjawi I, Rodriguez Milla MA, Cushman J, Shintani DK: Thiamin pyrophosphokinase is required for thiamin cofactor activation in Arabidopsis. Plant Mol Biol. 2007, 65: 151-162. 10.1007/s11103-007-9205-4.
Bocobza S, Aharoni A: Switching the light on plant riboswitches. Trends Plant Sci. 2008, 13: 526-533. 10.1016/j.tplants.2008.07.004.
Ribeiro DT, Farias LP, de Almeida JD, Kashiwabara PM, Ribeiro AF, Silva-Filho MC, Menck CF, Van Sluys MA: Functional characterization of the thi1 promoter region from Arabidopsis thaliana. J Exp Bot. 2005, 56: 1797-1804. 10.1093/jxb/eri168.
Ferreira S, Hjerno K, Larsen M, Wingsle G, Larsen P, Fey S, Roepstorff P, Salome Pais M: Proteome profiling of Populus euphratica Oliv. upon heat stress. Ann Bot. 2006, 98: 361-377. 10.1093/aob/mcl106.
Wong CE, Li Y, Labbe A, Guevara D, Nuin P, Whitty B, Diaz C, Golding GB, Gray GR, Weretilnyk EA, Griffith M, Moffatt BA: Transcriptional profiling implicates novel interactions between abiotic stress and hormonal responses in Thellungiella, a close relative of Arabidopsis. Plant Physiol. 2006, 140: 1437-1450. 10.1104/pp.105.070508.
Rapala-Kozik M, Kowalska E, Ostrowska K: Modulation of thiamine metabolism in Zea mays seedlings under conditions of abiotic stress. J Exp Bot. 2008, 59: 4133-4143. 10.1093/jxb/ern253.
Tunc-Ozdemir M, Miller G, Song L, Kim J, Sodek A, Koussevitzky S, Misra AN, Mittler R, Shintani D: Thiamin confers enhanced tolerance to oxidativestress in Arabidopsis. Plant Physiol. 2009, 151: 421-432. 10.1104/pp.109.140046.
Taylor NL, Tan Y-F, Jacoby RP, Millar AH: Abiotic environmental stress induced changes in the Arabidopsis thaliana chloroplast, mitochondria and peroxisome proteomes. J Proteomics. 2009, 72: 367-378. 10.1016/j.jprot.2008.11.006.
Baena-Gonzalez E: Energy signaling in the regulation of gene expression during stress. Mol Plant. 2010, 3: 300-313. 10.1093/mp/ssp113.
Baxter CJ, Redestig H, Schauer N, Repsilber D, Patil KR, Nielsen J, Selbig J, Liu J, Fernie AR, Sweetlove LJ: The metabolic response of heterotrophic Arabidopsis cells to oxidative stress. Plant Physiol. 2007, 143: 312-325.
Williams TC, Poolman MG, Howden AJ, Schwarzlander M, Fell DA, Ratcliffe RG, Sweetlove LJ: A genome-scale metabolic model accurately predicts fluxes in central carbon metabolism under stress conditions. Plant Physiol. 2010, 154: 311-323. 10.1104/pp.110.158535.
Obata T, Matthes A, Koszior S, Lehmann M, Araújo WL, Bock R, Sweetlove LJ, Fernie AR: Alteration of mitochondrial protein complexes in relation to metabolic regulation under short-term oxidative stress in Arabidopsis seedlings. Phytochemistry. 2011, 72: 1081-1091. 10.1016/j.phytochem.2010.11.003.
Munns R: Comparative physiology of salt and water stress. Plant Cell Einviron. 2002, 25: 239-250. 10.1046/j.0016-8025.2001.00808.x.
Munns R, Passioura JB, Guo J, Chazen O, Cramer GR: Water relations and leaf expansion: importance of time scale. J Exp Bot. 2000, 51: 1495-1504. 10.1093/jexbot/51.350.1495.
Sweetlove LJ, Heazlewood JL, Herald V, Holtzapffel R, Day DA, Leaver CJ, Millar AH: The impact of oxidative stress on Arabidopsis mitochondria. Plant J. 2002, 32: 891-904. 10.1046/j.1365-313X.2002.01474.x.
Taylor NL, Day DA, Millar AH: Targets of stress-induced oxidative damage in plant mitochondria and their impact on cell carbon/nitrogen metabolism. J Exp Bot. 2004, 55: 1-10.
Bunik VI, Fernie AR: Metabolic control exerted by the 2-oxoglutarate dehydrogenase reaction: a cross-kingdom comparison of the crossroad between energy production and nitrogen assimilation. Biochem J. 2009, 422: 405-421. 10.1042/BJ20090722.
Raines CA: The Calvin cycle revisited. Photosynth Res. 2003, 75: 1-10. 10.1023/A:1022421515027.
Henkes S, Sonnewald U, Badur R, Flachmann R, Stitt M: A small decrease of plastid transketolase activity in antisense tobacco transformants has dramatic effects on photosynthesis and phenylpropanoid metabolism. Plant Cell. 2001, 13: 535-551.
Arora A, Sairam RK, Srivastava GC: Oxidative stress and antioxidative system in plants. Curr Sci. 2002, 82: 1227-1238.
Couée I, Sulmon C, Gouesbet G, El Amrani A: Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J Exp Bot. 2006, 57: 449-459. 10.1093/jxb/erj027.
Valderrama R, Corpas FJ, Carreras A, Gómez-Rodríguez MV, Chaki M, Pedrajas JR, Fernández-Ocaña A, Del Río LA, Barroso JB: A dehydrogenase-mediated recycling of NADPH is a key antioxidant system against salt-induced oxidative stress in olive plants. Plant Cell Environ. 2006, 29: 1449-1459. 10.1111/j.1365-3040.2006.01530.x.
Estevez JM, Cantero A, Reindl A, Reichler S, Leon P: 1-Deoxy-D-xylulose-5-phosphate synthase, a limiting enzyme for plastidic isoprenoid biosynthesis in plants. J Biol Chem. 2001, 276: 22901-22909. 10.1074/jbc.M100854200.
Lois LM, Rodriguez-Concepcion M, Gallego F, Campos N, Boronat A: Carotenoid biosynthesis during tomato fruit development: regulatory role of 1-deoxy-D-xylulose5-phosphate synthase. Plant J. 2000, 22: 503-513. 10.1046/j.1365-313x.2000.00764.x.
Paterami I, Kanellis AK: Stress and developmental responses of terpenoid osynthetic genes in Cistus creticus subsp. creticus. Plant Cell Rep. 2010, 29: 629-641. 10.1007/s00299-010-0849-1.
Meier S, Tzfadia O, Vallabhaneni R, Gehring C, Wurtzel ET: A transcriptional analysis of carotenoid, chlorophyll and plastidial isoprenoid biosynthesis genes during development and osmotic stress responses in Arabidopsis thaliana. BMC Syst Biol. 2011, 19: 77.
Zhu JK: Salt and drought stress signal transduction in plants. Ann Rev Plant Biol. 2002, 53: 247-273. 10.1146/annurev.arplant.53.091401.143329.
Cordoba E, Porta H, Arroyo A, San Román C, Medina L, Rodríguez-Concepción M, León P: Functional characterization of the three genes encoding 1-deoxy-D-xylulose 5-phosphate synthase in maize. J Exp Bot. 2011, 62: 2023-2038. 10.1093/jxb/erq393.
Santner A, Estelle M: Recent advances and emerging trends in plant hormone signalling. Nature. 2009, 459: 1071-1078. 10.1038/nature08122.
Peleg Z, Blumwald E: Hormone balance and abiotic stress tolerance in crop plants. Curr Opin Plant Biol. 2011, 14: 290-295. 10.1016/j.pbi.2011.02.001.
Oh KJ, Park YS, Lee KA, Chung YJ, Cho TJ: Molecular characterization of a thiJ-like gene in Chinese cabbage. J Biochem Mol Biol. 2004, 37: 343-350. 10.5483/BMBRep.2004.37.3.343.
Ahn IP, Kim S, Lee YH: Vitamin B1 functions as an activator of plant disease resistance. Plant Physiol. 2005, 138: 1505-1515. 10.1104/pp.104.058693.
Bari R, Jones JD: Role of plant hormones in plant defense responses. Plant Mol Biol. 2009, 69: 473-488. 10.1007/s11103-008-9435-0.
Walia H, Wilson C, Wahid A, Condamine P, Cui X, Close TJ: Expression analysis of barley (Hordeum vulgare L.) during salinity stress. Funct Integr Genomics. 2006, 6: 143-156. 10.1007/s10142-005-0013-0.
Higo K, Ugawa Y, Iwamoto M, Korenaga T: Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27: 297-300. 10.1093/nar/27.1.297.
Vandepoele K, Quimbaya M, Casneuf T, De Veylder L, Van de Peer Y: Unraveling transcriptional control in Arabidopsis using cis-regulatory elements and coexpression networks. Plant Physiol. 2009, 50: 535-546.
Nemhauser JL, Hong F, Chory J: Different plant hormones regulate similar processes through largely nonoverlapping transcriptional responses. Cell. 2006, 126: 467-475. 10.1016/j.cell.2006.05.050.
Arbona V, Argamasilla R, Gómez-Cadenas A: Common and divergent physiological, hormonal and metabolic responses of Arabidopsis thaliana and Thellungiella halophila to water and salt stress. J Plant Physiol. 2010, 167: 342-1350.
Divi UK, Rahman T, Krishna P: Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biol. 2010, 10: 151-10.1186/1471-2229-10-151.
Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR: Abscisic acid: emergence of a core signaling network. Annu Rev Plant Biol. 2010, 61: 651-79. 10.1146/annurev-arplant-042809-112122.
Fujita Y, Fujita M, Shinozaki K, Yamaguchi-Shinozaki K: ABA-mediated transcriptional regulation in response to osmotic stress in plants. J Plant Res. 2011, 124: 509-525. 10.1007/s10265-011-0412-3.
Prasad TK, Anderson MD, Stewart CR: Acclimation, Hydrogen Peroxide, and Abscisic Acid Protect Mitochondria against Irreversible Chilling Injury in Maize Seedlings. Plant Physiol. 1994, 105: 619-627.
Millar AH, Heazlewood JL: Genomic and proteomic analysis of mitochondrial carrier proteins in Arabidopsis. Plant Physiol. 2003, 131: 443-453. 10.1104/pp.009985.
Kharenko OA, Boyd J, Nelson KM, Abrams SR, Loewen MC: Identification and characterization of interactions between abscisic acid and mitochondrial adenine nucleotide translocators. Biochem J. 2011, 437: 117-123. 10.1042/BJ20101898.
Bartels D, Sour E: Molecular responses of higher plants to dehydration. Plant responses to abiotic stress. Edited by: Heribert H, Shinozaki K. Springer, Berlin;2004:13-37.
Shinozaki K, Yamaguchi-Shinozaki K, Seki M: Regulatory network of gene expression in the drought and cold stress responses. Curr Opin Plant Biol. 2003, 6: 410-417. 10.1016/S1369-5266(03)00092-X.
Barrero JM, Piqueras P, González-Guzmán M, Serrano R, Rodríguez PL, Ponce MR, Micol JL: A mutational analysis of the ABA1 gene of Arabidopsis thaliana highlights the involvement of ABA in vegetative development. J Exp Bot. 2005, 56: 2071-2083. 10.1093/jxb/eri206.
Hu ML, Chen YK, Lin YF: The antioxidant and prooxidant activity of some B vitamins and vitamin-like compounds. Chem Bio Interact. 1995, 97: 63-73. 10.1016/0009-2797(95)03608-8.
Lukienko PI, Mel'nichenko NG, Zverinskii IV, Zabrodskaya SV: Antioxidant properties of thiamine. Bull Exp Biol Med. 2000, 130: 874-876.
Bettendorff L, Wins P: Thiamin diphosphate in biological chemistry: new aspects of thiamin metabolism, especially triphosphate derivatives acting other than as cofactors. FEBS J. 2009, 276: 2917-2925. 10.1111/j.1742-4658.2009.07019.x.
Lee BL, Ong HY, Ong CN: Determination of thiamine and its phosphate esters by gradient-elution high-performance liquid chromatography. J Chrom. 1991, 567: 71-80. 10.1016/0378-4347(91)80311-Y.
Acknowledgements
The authors thank Professor Andrzej Kozik for helpful discussions and critical reading of the manuscript. This work was supported in part by the Polish Ministry of Science and Higher Education (the grant No. NN303 320937 to M. R-K) and by Jagiellonian University (statutory funds No. DS15/WBBiB).
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
MRK designed and coordinated the study, participated in the HPLC and molecular biology experiments and prepared the manuscript; NW carried out the molecular biology experiments; MK participated in thiamine compound HPLC analysis; AKB participated in molecular biology studies and plant propagation. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Rapala-Kozik, M., Wolak, N., Kujda, M. et al. The upregulation of thiamine (vitamin B1) biosynthesis in Arabidopsis thaliana seedlings under salt and osmotic stress conditions is mediated by abscisic acid at the early stages of this stress response. BMC Plant Biol 12, 2 (2012). https://doi.org/10.1186/1471-2229-12-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2229-12-2