
Abstract
Background
Ears of Brn3c null mutants develop immature hair cells, identifiable only by certain molecular markers, and undergo apoptosis in neonates. This partial development of hair cells could lead to enough neurotrophin expression to sustain sensory neurons through embryonic development. We have therefore investigated in these mutants the patterns of innervation and of expression of known neurotrophins.
Results
At birth there is a limited expression of BDNF and NT-3 in the mutant sensory epithelia and DiI tracing shows no specific reduction of afferents or efferents that resembles neurotrophin null mutations. At postnatal day 7/8 (P7/8), innervation is severely reduced both qualitatively and quantitatively. 1% of myosin VIIa-positive immature hair cells are present in the mutant cochlea, concentrated in the base. Around 20% of immature hair cells exist in the mutant vestibular sensory epithelia. Despite more severe loss of hair cells (1% compared to 20%), the cochlea retains many more sensory neurons (46% compared to 15%) than vestibular epithelia. Even 6 months old mutant mice have some fibers to all vestibular sensory epithelia and many more to the cochlear apex which lacks MyoVIIa positive hair cells. Topologically organized central cochlea projections exist at least until P8, suggesting that functional hair cells are not required to establish such projections.
Conclusion
The limited expression of neurotrophins in the cochlea of Brn3c null mice suffices to support many sensory neurons, particularly in the cochlea, until birth. The molecular nature of the long term survival of apical spiral neurons remains unclear.
Similar content being viewed by others

Background
Brn3c is a POU domain factor that is crucial for inner ear hair cell development. Targeted null Brn3c mutants have no mature hair cells [1, 2]. Close examination has revealed that some 'immature' hair cells form in Brn3c null mutants and express cellular markers such as Myosin VI and VIIa, calretinin and parvalbumin [3]. Furthermore, these immature hair cells of Brn3c null mutants undergo apoptosis in neonates [3]. Consistent with an apparent absence of mature hair cells, initial work suggested that all vestibular and most spiral ganglion cells are lost by postnatal day 14 (P14; [1]). However, more detailed quantification by others [3] reported that at P4 about 77% of vestibular neurons and only 29% of spiral neurons are lost. It was suggested that there is possibly a complete loss in adults [3]. Other than these preliminary statements, no data exists concerning the detailed pattern of loss of innervation in Brn3c null mutants.
The initial development and partial differentiation of hair cells in Brn3c mutants could possibly lead to some neurotrophin expression in these cells to sustain sensory neurons through embryonic development and beyond. Data on various single and compound neurotrophin null mutants have shown that the loss of a specific neurotrophin leads to topologically restricted loss of sensory neurons in the embryonic ear [4, 5]. Such selective loss in Brn3c null mutants would therefore indicate reduction of a specific neurotrophin in immature hair cells. Moreover, recent work shows that in embryos NT-3 is primarily expressed in supporting cells, moving only around birth into hair cells [6, 7]. In fact, the selective loss of vestibular as compared to cochlear sensory neurons (77% versus 29%; [3]) suggests that NT-3 expression may be less downregulated in Brn3c null mutants than BDNF [6, 8, 9], provided that at least some differentiation of supporting cells takes place. In the ear [4] as well as elsewhere [10] neurotrophins are progressively downregulated in postnatal mammals and possibly replaced by other factors [11].
We have investigated in detail the pattern of innervation in the Brn3c mutants, as well as the expression of NT-3 and BDNF. We want to evaluate a possible correlation between the topology of sensory neuron loss and absence of a specific neurotrophin or topological loss of hair cells at birth and in older animals. This information could be important for an in-depth evaluation of the human deafness related to the Pou4f3 gene, DFNA15 [12].
We report here long term retention of cochlear sensory neurons for at least 6 months, in particular in the cochlear apex, in Brn3c null mutant mice. This retention of afferents and efferents is unrelated to hair cell differentiation as not even immature hair cells can be detected at early postnatal stages with MyoVII immunocytochemistry in this part of the cochlea. This retention of apical spiral neurons is also largely unrelated to neurotrophins which are known to be reduced in their expression in neonatal rodents [4].
Results
To appreciate the effects of the Brn3c null mutation on the pattern of the inner ear innervation, we first want to present the effects of BDNF and NT-3 null mutations at birth [6, 13, 14]. Null mutants of BDNF or its receptor trkB lose all innervation to the semicircular canals and have a reduced innervation to the utricle, saccule and apical turn of the cochlea. In contrast, null mutations of either NT-3 or its receptor trkC result in loss of spiral neurons in the basal turn with formation of an inner spiral bundle of afferents extending to the basal tip. Our null hypothesis for this study would be that Brn3c null mice show severe compromised production of these neurotrophins and should therefore show a comparable pattern of nerve fiber loss.
Brn3c null mutants at birth (P0)
Vestibular ganglia are smaller in Brn3c null mutants (Fig. 1b) than in control littermates (Fig. 1a), but larger than in BDNF or trkB null mutants of the same age [13, 15]. The reduction in apparent size of the vestibular ganglia is in agreement with quantitative data published previously [3, 13]. These data suggest a loss of 80–85% of vestibular sensory neurons in BDNF and trkB null mutants [13] and of 77% of vestibular sensory neurons in P4 Brn3c null mutants [3]. Thus, the size reduction in the vestibular sensory ganglion could be compatible with a loss of BDNF production in the immature hair cells.

Size variations of vestibular ganglia in control and mutant littermates labeled with DiI. In newborn animals, the vestibular ganglion shows a dramatic reduction in Brn3c null mutants (b) compared to control littermates (a). Abbreviations for this and other figures: AC, anterior crista; ggl, ganglion; c, spiral capillary; Genic. ggl., geniculate ganglion; GER, greater epithelial ridge; HC, horizontal crista; HaC, vestibular hair cell; IHC, inner hair cell; iHC, immature hair cell; IGSB, intraganglionic spiral bundle; OHC, outer hair cell; PC, posterior crista; S, saccule; SG, spiral ganglion; TM, tectorial membrane; U, utricle; VCN, ventral cochlear nucleus; VG, vestibular ganglion. Bar indicates 1000 μm.
However, the Brn3c null mutants show only a reduced density of afferent and efferent fibers to all vestibular sensory epithelia. There is no specific loss of all afferent and efferent innervation to the canal crista (Fig. 2a), a hallmark of both BDNF and trkB null mutations [13, 15]. In fact, the reduction of fibers seems to be rather uniform throughout a given sensory epithelium with the crista innervation being qualitatively no more reduced than the innervation of the utricle and saccule. No loss in specific areas, comparable to that in BDNF null mutant mice, is apparent in the saccule or utricle of Brn3c null mice. Similar patterns of innervation are obtained using acteylated tubulin immunocytochemistry (Fig. 2b).

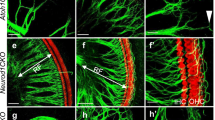
Innervation of Brn3c null and control ears are shown for newborn mice. There is no specific loss of fibers to any vestibular endorgan, as visualized by DiI labeling (a) or acetylated tubulin immunoreactivity (b). No major differences in pattern of projection through radial fibers are found in the cochlea of Brn3c null mutants (d,f) as compared to control littermates (c). Note, however, the lack of orderly fiber outgrowth to the outer hair cells (c, d). Efferent fibers to the ear show a well developed intraganglionic spiral bundle (IGSB) with no detectable differences compared to controls (e). Bar indicates 100 μm.
Consistent with the finding of Xiang et al. [3] of only a 29% loss of spiral sensory neurons at P4, our data show little difference in the pattern of innervation of the cochlea in P0 Brn3c null mutants (Fig. 2c,2d). No selective loss of spiral neurons is observed in Brn3c null mutants in the basal turn, a feature of either NT-3 or trkC loss [6, 9, 14]. Likewise, the innervation of the apex (Fig. 2f) shows no detectable abnormality in overall pattern of innervation compared to control animals (data not shown), an indication that BDNF could be expressed in the apex [6]. In addition, the pattern of efferent innervation shows no deviation from normal either (Fig. 2e), whereas they show the same pattern of loss as afferent fibers in neurotrophin null mutants [16]. These data suggest that the spiral sensory neurons develop qualitatively normal at least until P0 and therefore allow normal pathfinding of efferents. Most interestingly, there is no increase in radial fiber spacing in the apex, a specific problem of BDNF null mutants [6, 13].
However, there is one qualitative difference not recognized in any single neurotrophin null mutant. Afferents reach all three rows of outer hair cells in the basal turn of control wildtype littermates (Fig. 2c), but both afferent and efferent outgrowth is disorganized to outer hair cells in Brn3c null mutants (Fig. 2d) and does not show any clear organization into three distinct longitudinal fiber bundles paralleling the three rows in the outer hair cell region. These data suggest that fiber organization in the outer hair cell region is partly disrupted in Brn3c null mutant.
Brn3c null mutants at P7/8
Reduction of vestibular innervation is more pronounced than at P0. Loss of afferents and, even more clearly of efferents, is most pronounced in the utricle (Fig. 3a,3b) and saccule (data not shown). In contrast, the innervation to the canal cristae, while reduced, has changed little from P0. In particular the horizontal canal receives a rather dense afferent and efferent innervation in Brn3c null mutants.

Reduction of innervation of the ear is shown in 8-day old mice. While all vestibular sensory epithelia retain some afferent (a) and efferent (b) innervation, there is a more pronounced reduction, in particular of efferent fibers, to the utricle (a,b) as compared to the canals. In the cochlea there is an increased spacing between radial fiber bundles in the basal turn (c,e). No afferents can be traced to the three outer spiral bundles along outer hair cells of mutant littermates (c, e). The apex shows little change, except for the absence of fibers to the area of outer hair cells and the formation of inner spiral bundles passing along the few immature hair cells (d,f,g). The pattern obtained with DiI tracing is identical to that obtained by acetylated tubulin immunoreactivity (f,g). Bar indicates 1000 μm in a, 100 μm in b-g.
The cochlea at P8 shows more qualitative deviations from the normal pattern of innervation. For example, all three rows of outer hair cells now receive both afferent and efferent innervation in the base of control littermates (Fig. 3c). However, no fibers extend to the outer hair cell region in Brn3c null mutants (Figs. 3d,3e,3f). In control animals there is dense innervation of inner hair cells, whereas Brn3c null mutants show a curious aggregation of fibers near focal spots around the habenula perforata. The apex shows less pronounced deviations from normal (Fig. 3f,3g). However, as in the base, afferents and efferents extend sparingly to the outer hair cell region. There is formation of an inner spiral bundle communicating both afferents and efferents between the focal points where fibers appear to pass through the habenula perforata (Fig. 3f,3g). As in newborn animals, efferent fibers closely follow the pattern of innervation displayed by afferents (data not shown).
The limited qualitative effect of Brn3c null mutations on fiber loss is particularly obvious in the P8 Brn3c null mutant apical turn (Fig. 3g). Immunocytochemical and histochemcial data show the presence of only a few immature hair cells in the cochlea (Fig. 4), which amounted to about 1% (Table 1). Not a single immature hair cell forms in the apex of the cochlea. In contrast, more immature hair cells are found in the appropriate position of hair cells in the vestibular endorgans (Table 1). In fact, the vestibular end organs show no overt morphological defect other than the reduced presence of these immature hair cells. Nerve fibers can be traced into the sensory epithelium of P0 and P8 Brn3c null mutants past the habenula perforata and there is a normal location of spiral sensory neurons in Rosenthal's canal in the apex of P8 Brn3c null mutants (Fig. 3d,3g). The cochlea shows other defects as well. For example, the spiral capillary is not found underneath the organ of Corti but in the spiral limbus nearby (Fig. 4). The lack of immature hair cells seems to affect also the formation of pillar cells (undifferentiated almost throughout the cochlea; Fig. 4) and Deiters' cells (not recognizable based on their morphology), a suggestion consistent with recent molecular data on the interaction of hair cells and supporting cells [17, 18].

The distribution of hair cells as identified with MyoVII immunocytochemistry is shown for 7-day old whole mounted cochlea (a-d) and the appearance of cytological differentiation is shown as revealed with epoxy resin sections in 7-day old (e-g) and 6-month old (h) control (a,c,e) and Brn3c mutant cochleae (b,d,f-h). Note the uniform staining with this antibody of all three rows of outer hair cells (OHC) and the single row of inner hair cells (IHC) in the wildtype (a,c). In contrast, only few immature hair cells (iHC) are found in the basal turn of the mutant littermate (b) and only an occasional cell is MyoVII immunopositive in the middle turn (d). No MyoVII positive immature hair cells exist in the apex. Sections show the degree of differentiation of hair cells and their innervating fibers stained with tubulin immunocytochemistry (arrow; e-h). Note the presence of a spiral capillary (C) under the tunnel of Corti separating the inner and outer hair cells in the P7 wildtype (e). In contrast, no such capillary is present underneath the organ of Corti in mutants (f-h) and no tunnel of Corti is found either. A tectorial membrane forms (TM) but otherwise there is little cyotological differentiation. Note that tubulin-positive fibers can be traced to the immature cells until 6 months (h). Bar in c indicates 100 μm.
Comparison of numbers of immature hair cells in the vestibular and cochlear areas (Fig. 4) with the numbers of surviving vestibular and cochlear sensory neurons does not show any correlation (Table 1; Fig. 5). Only a few undifferentiated hair cells were detected by MyoVIIa immunostaining and only in the basal region of the cochlea. And yet only about 50% of cochlear sensory neurons are lost overall with a less severe loss in the apex in which we could not detect any immature hair cells with MyoVIIa immunocytochemistry. This does not exclude the possibility that even less differentiated hair cells may form in the apex.

This graph shows the reduction in sensory neurons (top) and in hair cells (bottom) between 7-day old wildtype and Brn3c null mutant littermates. Note the sharp reduction to 1% in the cochlear immature hair cells (bottom) whereas spiral sensory neurons (SG) are only reduced by 50% (top). In contrast, 20% immature hair cells could be identified in vestibular sensory epithelia (saccule has been selected; bottom) whereas only around 15% of the vestibular sensory neurons survive to this age (VG; top). For details on the data see Table 1.
The central projection of DiI and DiA traces fibers to the cochlear nuclei (Fig. 6) where they show a clear segregation of apical turn afferents (Fig. 6, green) and basal turn afferents (Fig. 6, red) in both the entering cochlear nerve (Fig. 6a) and the ventral cochlear nucleus (VCN) (Fig. 6b). Qualitative comparable central projections are found in wildtype control littermates (data not shown) and suggest that the central projection to cochlear nuclei is at least until P8 in mice not critically dependent on auditory information or other hair cell-mediated influences, in agreement with recent suggestions [5, 19].

These confocal images show the projection pattern of a P8 Brn3c null mutant mouse. DiI (red) was inserted into the basal turn, DiA (green) was inserted into the apical turn. Note that the fibers run in discrete bundles inside the cochlear nerve (a), spiral around each other near the root and project to distinct medial and lateral divisions of the ventral cochlear nucleus (b). These data suggest that a crude cochleotopic projection can form independently of any signaling from the immature hair cells of the cochlea. Arrow indicates the efferent bundle as it leaves the afferent fiber tract. Bar indicates 100 μm.
6 months Brn3c null mutants
Very few fibers (both afferent and efferent) to the vestibular epithelia remain (Fig. 7a), thus displaying a further reduction of the pattern of innervation found in P8 Brn3c null mutants. Notably, there are still a number of fibers extending from the very few vestibular sensory neurons to the canal epithelia, in particular the horizontal canal (Fig. 7a,7b). Only very few fibers extend to the utricle and saccule with the reduction of efferents to these epithelia being almost complete (Fig. 7a and data not shown). Comparable to control mice, there is some transcellular labeling with DiI to cells apparently contacted by the afferent fibers in the Brn3c null mutant utricle (Fig. 7b). However, these cells could not be clearly identified as immature hair cells based on their morphology.

The innervation of sensory epithelia is shown in 6-month old Brn3c null mutant mice. Compared to P8 animals there is a further reduction of fiber density to vestibular sensory epithelia, but fibers can be traced to all sensory epithelia (a,b). Transcellular labeling with DiI (b) suggest contacts between these afferents and immature hair cells. The cochlea shows a much reduced density of radial bundles, even in the apex (c,d). However, spiral sensory neurons are present in Rosenthal's canal (c,d) and fibers extend to the organ of Corti where they form longitudinal tracts in the immature epithelium. Bar indicates 100 μm.
The cochlea, most prominently the apex, receives many afferent fibers, which form an interrupted inner spiral bundle (Fig. 7c). Some of the radial fiber bundles are myelinated (Fig. 7d). Only an occasional fiber extends beyond the inner spiral bundle into the area of the outer hair cells (Fig. 7d). Even at this late stage in maturation there are still numerous spiral neurons present in the apex (Fig. 7c,7d).
In summary, these tracing data suggest that mature hair cells are not necessary to direct fiber outgrowth and to maintain some fibers at least up to 6 months in the apex of the cochlea. Importantly, the spatio-temporal loss of innervation does not follow the pattern of loss known from neurotrophin and neurotrophin receptor null mutations as it starts in Brn3c mutant mice after birth.
In situhybridization confirms neonatal neurotrophin expression
We processed four ears of P0 and P8 Brn3c null and control littermates for expression of BDNF and NT-3 using the riboprobes previously described [7]. We did not analyze neurotrophin expression in 6-month old animals as neurotrophins are downregulated in neonates [20] and no expression can be found even in normal animals [4]. In the canal cristae we expected, and found, only a BDNF signal. This signal was very strong over hair cells of the P0 control animals (Fig. 8b). In contrast, only a faint signal was detected in the Brn3c null mutant (Fig. 8a) and the few silver grains were located predominantly over the area of the immature hair cells. The BDNF signal in the utricle was clearly centered on the hair cells in the control animal but some aggregation of silver grains appeared also in the Brn3c null mutants (data not shown).

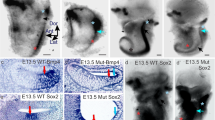
The expression of BDNF and NT-3 mRNA is shown in the P0 vestibular sensory epithelia. Silver grains are false colored as red to enhance visibility against the grey Nissl-stained background. Canal cristae contain only BDNF (a,b), but gravistatic endorgans, such as saccule and utricle, contain both BDNF and NT-3 (c-f). Note the prominent BDNF expression in the hair cell region (HaC) of control canal (b) and saccule (d) and low levels of expression in the Brn3c null mice (a,c) over immature hair cells (iHC). The NT-3 expression is much more diffuse in both control and Brn3c mutant littermates. Bar indicates 100 μm.
In the saccule, we directly compared the BDNF (Fig. 8c,8d) and NT-3 signal (Fig. 8e,8f) of the Brn3c null mutant (Fig. 8c,8e) and control littermates (Fig. 8d,8f). A strong BDNF signal was found over the hair cells of control animals and an occasional immature hair cell also showed a BDNF signal in the Brn3c null mutants. The NT-3 signal was more diffuse in both the control and the Brn3c null littermates but showed some slightly above background signal in the area of the sensory epithelium even in the Brn3c null mutant.
In the cochlea, the BDNF signal was very weak in the base in both the control (Fig. 9b) and the Brn3c null littermates (Fig. 9a). However, there was a surprisingly strong NT-3 signal over the organ of Corti in the apex of Brn3c null littermates (Fig. 9c) that closely matched the signal found in the control littermates (Fig. 9d). The NT-3 signal in the area of the cochlea that corresponds to the IHC's persisted at least until P8 in the apex of Brn3c null mice (Fig. 9e,9f).

BDNF and NT-3 mRNA expression is shown in the cochlea of P0 (a-d) and P8 (e,f) Brn3c null and control littermates. Note the limited expression of BDNF in the basal turn of both wildtype and mutant littermates (a,b) and the much stronger expression of NT-3 in the apex over the region of the inner hair cells in both wildtype and control littermates (c,d). NT-3 expression persists at least until P8 in the apex of Brn3c null mice in an area that topologically compares to the inner hair cells of control animals (e,f, arrows). Bar indicates 100 μm.
Discussion
We will explore five points in this discussion: How the limited expression of neurotrophins relates to the apparent survival of primary neurons until P8; how the known absence of apical hair cells and of classical neurotrophins can be related to the presence of large numbers of apical turn spiral neurons; how absence of differentiated hair cells affects afferent and efferent targeting; and how these data possibly relate to other mutant animals and to children born with profound hearing loss.
Immature hair cells and limited expression of neurotrophins rescue afferent projection until P8
The neurotrophins have long been implicated as the mediators for target mediated cell death. In essence, this theory of regulation of neuronal connections via neurotrophic support implies that only limited quantities of neurotrophins are released by the target tissue to support only properly connected sensory neurons by providing only these with a critical amount of neurotrophins [21]. Past experiments, which eliminated one or more neurotrophins entirely, only partially tested the basic assumption of this theory in vivo. For example, in BDNF heterozygotic animals there is a small decrease in the number of sensory neurons in the ear [13] but no change in the overall pattern of innervation. Moreover, all of the neurotrophin/neurotrophin receptor null mutants studied to date eliminate expression in both the peripheral target and the brain simultaneously and achieve their effects in late embryos. In contrast, Brn3c is barely expressed during embryogenesis in the CNS [22] thus arguing that the survival of ear primary neurons depends crucially on the periphery.
We suggest that limited expression of neurotrophins in the immature target leads to retention of primary neurons and their afferent fibers in newborn and early postnatal Brn3c null mice. In agreement with this suggestion, our in situ data show a severe reduction in expression of BDNF (Figs. 8, 9). Despite this reduction, some, apparently biologically significant low levels of neurotrophins are apparently expressed in all sensory epithelia in embryonic Brn3c null mutants. It appears that even these low levels of neurotrophins in the vestibular sensory epithelia can sustain normal fiber outgrowth and a limited maintenance until P0. The comparatively high level of NT-3 expression in Brn3c null mutant cochlea, which has been shown to be the neurotrophin most prominently supporting spiral sensory neurons [6, 9, 14], is in agreement with the normal development of cochlear innervation in newborn Brn3c null mutants.
Since BDNF appears to be exclusively produced by hair cells [6, 7, 20], the lack of terminal differentiation of hair cells appears to be accompanied by only limited expression of BDNF, as is the case in the canal cristae (Fig. 8). Past research has shown that in BDNF and trkB null mutants this innervation to canal cristae is lost before birth [6, 13]. That even these low levels of expression are significant is clear from the fact that all canal sensory epithelia receive an innervation by both afferents and efferents at birth and later (Figs. 2, 3). The situation in the utricle, saccule and cochlea is less clear as both BDNF and NT-3 are expressed [6, 7]. In fact, it appears that the levels of NT-3 expression hardly differ from those in control littermates (Figs. 8, 9). The latter is particularly obvious in the apex of the cochlea. In fact, in the cochlea of embryos and neonates as well as in the saccule and utricle of embryos, NT-3 is predominantly expressed in supporting cells [6]. This expression of NT-3 in supporting cells in embryos and neonates may even preserve in Brn3c null mutants the numerous spiral sensory neurons in the apex of 8-day old animals (Figs. 3, 4).
Long term survival of apical sensory neurons is unrelated to differentiated hair cells and likely is independent of neurotrophins
Past work has shown that BDNF and NT-3 appear to shift in their expression into inner hair cells around birth or are lost in the ear and other parts of the nervous system [6, 10, 20, 23]. It has been suggested that other neurotrophins may come into play in the ear [11]. However, the apex of Brn3c null mutants, which retains most of the spiral neuron afferents at 6-month, shows no formation of mature hair cells at any time surveyed here (P0, P8, P10). Thus the factor(s) cannot be released by differentiated hair cells but there is an unexplored possibility of generalized expression of neurotrophins at low levels in the undifferentiated epithelia. However, known neurotrophins are largely absent in the adult cochleae [4] and significant amounts of neurotrophin expression appear to develop only in postnatal animals in the CNS [24]. Such expression in the CNS could possibly offset to some extent the peripheral loss of neutrophins in targeted null mutants. This would work only in cases in which the loss of primary neurons would not already be completed long before birth as is the case in the ear [6, 13].
It therefore remains unclear what supports the many afferents in the apex of the Brn3c null mutants until 6 months of age or longer, unknown peripheral neurotrophic substances or neurotrophic support provided by the CNS. Comparable long term retention of apical spiral sensory neurons was described for the deaf white cat [25] and may also be the case in humans with congenital deafness.
Growth of fibers to the cochlea does not require mature hair cells
Formation of radial fibers that bring peripheral processes of spiral neurons to the organ of Corti seems to be rather normal, even in the apex of the cochlea which does not even develop immature hair cells recognizable by Myo VIIa. This does not preclude that even less differentiated hair cell precursors may form in the apex or that those precursors have died before birth. Interestingly, the growth to outer hair cells is most affected in Brn3c null mutant cochleae. Instead of extending radially through the tunnel of Corti to outer hair cells, afferents appear to stall and extend in longitudinal directions as inner spiral bundles (Figs. 3, 7). It has been shown that the differentiation of pillar cells depends on activation of Fgfr3 [26] by FGF's, probably Fgf8, a factor produced by developing and mature inner hair cells [18, 27]. The formation of supporting cells also appears to depend on the proper expression of various bHLH factors such as Hes 1 and 5 which appear to be regulated by the Notch signaling pathway [17, 28, 29]. It is possible that the apparent inability of fibers to extend along outer hair cells is related to the lack of differentiation of supporting cells in the absence of mature hair cells. Probing for the proper expression of supporting cell specific markers in Brn3c null mutants is necessary to further evaluate these suggestions.
Previous work has suggested that efferent fibers, derived from facial branchial motoneurons, follow afferents and for that reason reflect in detail all connectional deviations of afferents [16, 30–32]. Our data agree with this scenario but extend it. Specifically, efferent fibers may suffer the same loss of their molecularly unknown neurotrophic support as do afferents and therefore show the same spatio-temporal profile of loss (Fig. 3). Alternatively, efferent fibers can sustain established connections only as long as afferent fibers are present. More information on the actual molecular support of efferent fibers in the ear is needed before those two possibilities can be distinguished.
The onset of cochleotopic projections has been investigated in detail in only very few mammals [19]. These data suggest that a cochleotopic projection develops before onset of hearing. Our data show that at least a cochleotopic projection in the cochlear nerve and the cochlear nuclei develops even in Brn 3c null mutants (Fig. 6). Those data are also consistent with the formation of a crude cochleotopic projection in NT-3 null mutants [6, 14]. Those mutants have a much more severely reduced density of innervation in the developing cochlea [6, 14] than the reduction we found in Brn3c null mutants at P0. In this context, it would be interesting to see whether the recently generated transgenic mouse in which BDNF is expressed under NT-3 promoter control [33] could actually perform an even more pronounced rescue in a Brn3c null mutant background.
Relevance of our findings to other mutations with hair cell loss
Vahava et al. [12] and Frydman et al. [34] have demonstrated that mutation in a single human allele of Pou4f3 (Brn3c) results in a late onset of high frequency hearing loss. Thus far no data are available that correlate the late onset of hearing loss with morphological defects in the cochlea. In fact, an investigation of Brn3c heterozygotic mice showed no additional effect on hearing loss due to haploinsufficiency in these mice [35]. Our data on Brn3c null mouse mutants nevertheless suggest that the high to low frequency progressive hearing loss in humans may correlate with the unknown factor(s) that mediate apical spiral neuron survival into adulthood. Quantitative data on human Pou4f3 cochleae is needed to verify that spiral neuron loss is more pronounced in the basal, high frequency area of these hearing-deficient patients. Similar data are also needed for other congenitally deaf children [36] and in the deaf white cat [25]. Further work on the recently available mutant mice with specific absence of all hair cells [37], outer hair cells in the base [38] or apex [39] could further help to clarify the role of hair cells in forming and maintaining afferent and efferent innervation in these model mice.
Conclusion
The progressive loss of afferent and efferent innervation in Brn3c null mutants shows neither in spatial nor in temporal pattern a resemblance to losses reported in simple BDNF or NT-3 null mutations [6]. Null mutants of ear specific neurotrophins have completed the loss of sensory neurons before birth. In contrast, our results suggest a slow loss of afferent and efferent innervation between P0 and at least 6 months. This late sensory neuron loss is likely not related to known neurotrophin signaling which becomes reduced in neonatal wildtype animals. Other factors, such as GDNF [11, 40] need to be investigated in their expression in these mutants and their functional role needs to be assessed, in particular in the cochlear apex.
The long term retention of afferents in the cochlear apex in the absence of any apparent differentiation of hair cells raises hopes for cochlear implants in deaf-born children. Similar long term retention of apical afferents exists in the deaf white cat [25] and should be explored in other model systems with embryonic and neonatal hair cell loss [37, 38].
Methods
Breeding and genotyping
Brn3c mice were bred as previously described [22]. Mice were genotyped and Brn3c null mutants were raised to a specific age (P0, N = 8; P7, N = 6; P8, N = 6; 6 months old, N = 2). The animals were deeply anesthetized with pentobarbital and transcardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Ears were postfixed in the same fixative for at least one day prior to dissection.
Tracing of nerve fibers
DiI tracing from the brainstem was performed as previously described [41]. Small filter stripes soaked with saturated DiI were implanted into the efferent fiber bundle near the floor plate or into the ascending or descending afferents in the alar plate [42]. After appropriate diffusion time of the dye, the ears were dissected and mounted flat for visualization in an epifluorescent microscope. Images were taken on black and white film or acquired with a cooled CCD camera and processed using ImagePro software (Media Cybernetics).
Ears were subsequently reacted for acetylated tubulin to reveal the pattern of innervation in addition to the DiI tracing with a different technique as previously described [14]. Briefly, dissected ears were incubated with 1:500 anti-acetylated tubulin antibodies (Sigma, St. Louis) followed by secondary antibodies conjugated to HRP. Dissected inner ears were reacted with DAB and H2O2 for HRP distribution and subsequently viewed as whole mounts.
After photographing as whole mounts, the ears were embedded in epoxy resin and sectioned to reveal the distribution of fibers inside the sensory epithelia in more detail.
Cochleotopic projection was evaluated by inserting DiI soaked filter strips into the base and DiA soaked filter strips into the apical turn of 2 P0 and 4 P8 Brn3c null mutants and a similar number of wildtype littermates. After appropriate diffusion time [43], the brains with the attached cochlear nerve were embedded in gelatin, hardened in 4% PFA over night, sectioned coronally on a vibratome (100 μm thickness) and viewed with a Biorad Radiance 2000 confocal system attached to a Nikon Eclipse 800 microscope. Image stacks were collapsed to view the entire projection in one section in one focal plane.
In situhybridization
Four ears each of Brn3c null mutants (P0 and P8) and control littermates were embedded in paraffin, sectioned and probed for the presence of BDNF and NT-3 mRNA using the in situ technique previously described [7]. Sections were lightly counterstained and viewed with bright and dark field microscopy. Images were acquired with a CCD camera and displayed as false color images as previously described [18].
Immunocytochemistry of hair cells and quantification of hair cells and neurons
Immunostaining of P7 cochlear whole mounts using the hair cell specific-myosin VII (MyoVIIa) antibody was performed as previously described [2]. Images of labeled organs of Corti were then acquired using a SPOT digital camera (Diagnostic Instruments Inc.) and the number of hair cells was scored from acquired images. To determine the number of hair cells or neurons in vestibular endorgans or sensory ganglia, serial sections were stained with Cresyl Violet and images were similarly acquired and scored. Every other section was scored for each sensory epithelium and every forth section was scored for each ganglion. Only neurons or cells with a clear nucleus and nucleoli were counted and 4–6 samples were counted for each epithelium and ganglion. All data were tested for significance using two-sample Student's t-test with unequal variances.
References
Erkman L, McEvilly RJ, Luo L, Ryan AK, Hooshmand F, O'Connell SM, Keithley EM, Rapaport DH, Ryan AF, Rosenfeld MG: Role of transcription factors Brn-3.1 and Brn-3.2 in auditory and visual system development. Nature. 1996, 381: 603-606. 10.1038/381603a0.
Xiang M, Gan L, Li D, Chen ZY, Zhou L, O'Malley BW, Klein W, Nathans J: Essential role of POU-domain factor Brn-3c in auditory and vestibular hair cell development. Proc Natl Acad Sci U S A. 1997, 94: 9445-9450. 10.1073/pnas.94.17.9445.
Xiang M, Gao WQ, Hasson T, Shin JJ: Requirement for Brn-3c in maturation and survival, but not in fate determination of inner ear hair cells. Development. 1998, 125: 3935-3946.
Fritzsch B, Pirvola U, Ylikoski J: Making and breaking the innervation of the ear: neurotrophic support during ear development and its clinical implications. Cell Tissue Res. 1999, 295: 369-382. 10.1007/s004410051244.
Rubel EW, Fritzsch B: AUDITORY SYSTEM DEVELOPMENT: Primary Auditory Neurons and Their Targets. Annu Rev Neurosci. 2002, 25: 51-101. 10.1146/annurev.neuro.25.112701.142849.
Farinas I, Jones KR, Tessarollo L, Vigers AJ, Huang E, Kirstein M, de Caprona DC, Coppola V, Backus C, Reichardt LF, Fritzsch B: Spatial shaping of cochlear innervation by temporally regulated neurotrophin expression. J Neurosci. 2001, 21: 6170-6180.
Pirvola U, Ylikoski J, Palgi J, Lehtonen E, Arumae U, Saarma M: Brain-derived neurotrophic factor and neurotrophin 3 mRNAs in the peripheral target fields of developing inner ear ganglia. Proc Natl Acad Sci U S A. 1992, 89: 9915-9919.
Ernfors P, Van De Water T, Loring J, Jaenisch R: Complementary roles of BDNF and NT-3 in vestibular and auditory development. Neuron. 1995, 14: 1153-1164.
Farinas I, Jones KR, Backus C, Wang XY, Reichardt LF: Severe sensory and sympathetic deficits in mice lacking neurotrophin-3. Nature. 1994, 369: 658-661. 10.1038/369658a0.
Cosgaya JM, Chan JR, Shooter EM: The neurotrophin receptor p75NTR as a positive modulator of myelination. Science. 2002, 298: 1245-1248. 10.1126/science.1076595.
Hashino E, Dolnick RY, Cohan CS: Developing vestibular ganglion neurons switch trophic sensitivity from BDNF to GDNF after target innervation. J Neurobiol. 1999, 38: 414-427. 10.1002/(SICI)1097-4695(19990215)38:3<414::AID-NEU9>3.0.CO;2-Y.
Vahava O, Morell R, Lynch ED, Weiss S, Kagan ME, Ahituv N, Morrow JE, Lee MK, Skvorak AB, Morton CC, Blumenfeld A, Frydman M, Friedman TB, King MC, Avraham KB: Mutation in transcription factor POU4F3 associated with inherited progressive hearing loss in humans. Science. 1998, 279: 1950-1954. 10.1126/science.279.5358.1950.
Bianchi LM, Conover JC, Fritzsch B, DeChiara T, Lindsay RM, Yancopoulos GD: Degeneration of vestibular neurons in late embryogenesis of both heterozygous and homozygous BDNF null mutant mice. Development. 1996, 122: 1965-1973.
Fritzsch B, Farinas I, Reichardt LF: Lack of neurotrophin 3 causes losses of both classes of spiral ganglion neurons in the cochlea in a region-specific fashion. J Neurosci. 1997, 17: 6213-6225.
Fritzsch B, Silos-Santiago I, Bianchi LM, Farinas I: The role of neurotrophic factors in regulating the development of inner ear innervation. Trends Neurosci. 1997, 20: 159-164. 10.1016/S0166-2236(96)01007-7.
Fritzsch B, Barbacid M, Silos-Santiago I: The combined effects of trkB and trkC mutations on the innervation of the inner ear. Int J Dev Neurosci. 1998, 16: 493-505. 10.1016/S0736-5748(98)00043-4.
Zine A, Aubert A, Qiu J, Therianos S, Guillemot F, Kageyama R, de Ribaupierre F: Hes1 and Hes5 activities are required for the normal development of the hair cells in the mammalian inner ear. J Neurosci. 2001, 21: 4712-4720.
Pirvola U, Ylikoski J, Trokovic R, Hebert J, McConnell S, Partanen J: FGFR1 Is Required for the Development of the Auditory Sensory Epithelium. Neuron. 2002, 35: 671.
Leake PA, Snyder RL, Hradek GT: Postnatal refinement of auditory nerve projections to the cochlear nucleus in cats. J Comp Neurol. 2002, 448: 6-27. 10.1002/cne.10176.
Wheeler EF, Bothwell M, Schecterson LC, von Bartheld CS: Expression of BDNF and NT-3 mRNA in hair cells of the organ of Corti: quantitative analysis in developing rats. Hear Res. 1994, 73: 46-56. 10.1016/0378-5955(94)90281-X.
Levi-Montalcini R: The nerve growth factor 35 years later. Science. 1987, 237: 1154-1162.
Xiang M, Gan L, Li D, Zhou L, Chen ZY, Wagner D, O'Malley BW, Klein W, Nathans J: Role of the Brn-3 family of POU-domain genes in the development of the auditory/vestibular, somatosensory, and visual systems. Cold Spring Harb Symp Quant Biol. 1997, 62: 325-336.
Ylikoski J, Pirvola U, Moshnyakov M, Palgi J, Arumae U, Saarma M: Expression patterns of neurotrophin and their receptor mRNAs in the rat inner ear. Hear Res. 1993, 65: 69-78. 10.1016/0378-5955(93)90202-C.
Tierney TS, T PD, Xia G, Moore DR: Development of brain-derived neurotrophic factor and neurotrophin-3 immunoreactivity in the lower auditory brainstem of the postnatal gerbil. Eur J Neurosci. 2001, 14: 785-793. 10.1046/j.0953-816x.2001.01690.x.
Heid S, Hartmann R, Klinke R: A model for prelingual deafness, the congenitally deaf white cat – population statistics and degenerative changes. Hear Res. 1998, 115: 101-112. 10.1016/S0378-5955(97)00182-2.
Colvin JS, Bohne BA, Harding GW, McEwen DG, Ornitz DM: Skeletal overgrowth and deafness in mice lacking fibroblast growth factor receptor 3. Nat Genet. 1996, 12: 390-397.
Pirvola U, Spencer-Dene B, Xing-Qun L, Kettunen P, Thesleff I, Fritzsch B, Dickson C, Ylikoski J: FGF/FGFR-2(IIIb) signaling is essential for inner ear morphogenesis. J Neurosci. 2000, 20: 6125-6134.
Eddison M, Le Roux I, Lewis J: Notch signaling in the development of the inner ear: lessons from Drosophila. Proc Natl Acad Sci U S A. 2000, 97: 11692-11699. 10.1073/pnas.97.22.11692.
Fritzsch B, Beisel KW, Bermingham NA: Developmental evolutionary biology of the vertebrate ear: conserving mechanoelectric transduction and developmental pathways in diverging morphologies. Neuroreport. 2000, 11: R35-44.
Kim WY, Fritzsch B, Serls A, Bakel LA, Huang EJ, Reichardt LF, Barth DS, Lee JE: NeuroD-null mice are deaf due to a severe loss of the inner ear sensory neurons during development. Development. 2001, 128: 417-426.
Karis A, Pata I, van Doorninck JH, Grosveld F, de Zeeuw CI, de Caprona D, Fritzsch B: Transcription factor GATA-3 alters pathway selection of olivocochlear neurons and affects morphogenesis of the ear. J Comp Neurol. 2001, 429: 615-630. 10.1002/1096-9861(20010122)429:4<615::AID-CNE8>3.0.CO;2-F.
Ma Q, Anderson DJ, Fritzsch B: Neurogenin 1 null mutant ears develop fewer, morphologically normal hair cells in smaller sensory epithelia devoid of innervation. J Assoc Res Otolaryngol. 2000, 1: 129-143.
Coppola V, Kucera J, Palko ME, Martinez-De Velasco J, Lyons WE, Fritzsch B, Tessarollo L: Dissection of NT3 functions in vivo by gene replacement strategy. Development. 2001, 128: 4315-4327.
Frydman M, Vreugde S, Nageris BI, Weiss S, Vahava O, Avraham KB: Clinical characterization of genetic hearing loss caused by a mutation in the POU4F3 transcription factor. Arch Otolaryngol Head Neck Surg. 2000, 126: 633-637.
Keithley EM, Erkman L, Bennett T, Lou L, Ryan AF: Effects of a hair cell transcription factor, Brn-3.1, gene deletion on homozygous and heterozygous mouse cochleas in adulthood and aging. Hear Res. 1999, 134: 71-76. 10.1016/S0378-5955(99)00070-2.
Yagi M, Kanzaki S, Kawamoto K, Shin B, Shah PP, Magal E, Sheng J, Raphael Y: Spiral ganglion neurons are protected from degeneration by GDNF gene therapy. J Assoc Res Otolaryngol. 2000, 1: 315-325.
Bermingham NA, Hassan BA, Price SD, Vollrath MA, Ben-Arie N, Eatock RA, Bellen HJ, Lysakowski A, Zoghbi HY: Math1: an essential gene for the generation of inner ear hair cells. Science. 1999, 284: 1837-1841. 10.1126/science.284.5421.1837.
Liberman MC, Gao J, He DZ, Wu X, Jia S, Zuo J: Prestin is required for electromotility of the outer hair cell and for the cochlear amplifier. Nature. 2002, 419: 300-304. 10.1038/nature01059.
Li S, Price SM, Cahill H, Ryugo DK, Shen MM, Xiang M: Hearing loss caused by progressive degeneration of cochlear hair cells in mice deficient for the Barhl1 homeobox gene. Development. 2002, 129: 3523-3532.
Qun LX, Pirvola U, Saarma M, Ylikoski J: Neurotrophic factors in the auditory periphery. Ann N Y Acad Sci. 1999, 884: 292-304.
Fritzsch B, Nichols DH: DiI reveals a prenatal arrival of efferents at the differentiating otocyst of mice. Hear Res. 1993, 65: 51-60. 10.1016/0378-5955(93)90200-K.
Fritzsch B, Christensen MA, Nichols DH: Fiber pathways and positional changes in efferent perikarya of 2.5- to 7-day chick embryos as revealed with DiI and dextran amines. J Neurobiol. 1993, 24: 1481-1499.
Maklad A, Fritzsch B: The developmental segregation of posterior crista and saccular vestibular fibers in mice: A carbocyanine tracer study using confocal microscopy. Develop Brain Research. 2002, 135: 1-17. 10.1016/S0165-3806(01)00327-3.
Acknowledgements
Supported by the National Eye Institute (EY12020, MX), the March of Dimes Birth Defects Foundation (MX), the Egyptian Government (AM), the Juselius Foundation (UP), the NIDCD (2 P01 DC00215, BF; DC04594, M.X.), the Taub foundation (BF) and NASA (01-OBPR-06; BF).
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
MX generated the null mutation, bred all the mice used for this study and generated the statistical data and the immunocytochemical data with MyoVII, AM conducted the tract tracing studies of cochlear afferents and helped with the histological analysis and assembly of image plates, UP carried out the in situ hybridization for neurotrophins and assembled the plates for these data, BF wrote the manuscript, carried out most of the neuronal tracing and immunocytochemistry for acetylated tubulin and data presentation. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Xiang, M., Maklad, A., Pirvola, U. et al. Brn3c null mutant mice show long-term, incomplete retention of some afferent inner ear innervation. BMC Neurosci 4, 2 (2003). https://doi.org/10.1186/1471-2202-4-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2202-4-2