
Abstract
Background
In Drosophila, male flies require the expression of the male-specific Fruitless protein (FRUM) within the developing pupal and adult nervous system in order to produce male courtship and copulation behaviors. Recent evidence has shown that specific subsets of FRUM neurons are necessary for particular steps of courtship and copulation. In these neurons, FRUM function has been shown to be important for determining sex-specific neuronal characteristics, such as neurotransmitter profile and morphology.
Results
We identified a small cohort of FRUM interneurons in the brain and ventral nerve cord by their co-expression with the transcription factor Engrailed (En). We used an En-GAL4 driver to express a fruM RNAi construct in order to selectively deplete FRUM in these En/FRUM co-expressing neurons. In courtship and copulation tests, these males performed male courtship at wild-type levels but were frequently sterile. Sterility was a behavioral phenotype as these En-fruMRNAi males were less able to convert a copulation attempt into a stable copulation, or did not maintain copulation for long enough to transfer sperm and/or seminal fluid.
Conclusions
We have identified a population of interneurons necessary for successful copulation in Drosophila. These data confirm a model in which subsets of FRUM neurons participate in independent neuronal circuits necessary for individual steps of male behavior. In addition, we have determined that these neurons in wild-type males have homologues in females and fru mutants, with similar placement, projection patterns, and neurochemical profiles.
Similar content being viewed by others


Background
The genes that govern behavior and how these genes function to create specific neural circuits that underlie behavior can be addressed in the model organism, Drosophila melanogaster, which has both well-documented stereotyped behaviors and a wealth of genetic information available. In Drosophila, male reproductive behaviors are dependent primarily on the activity of the fruitless (fru) and doublesex (dsx) genes, outputs directly regulated by the sex determination hierarchy [1–7]. The male-specific functions of fru derive from transcripts generated from the distal-most fru promoter (P1) [8–11]. In males, but not in females, P1 fru transcripts are translated into male-specific FRU proteins (FRUM) [8–11]. FRUM proteins are members of the BTB/ZnF (Broad complex-Tramtrack-Bric-a-brac/Zinc Finger) family, likely function as transcription factors, and are expressed in 2000–4000 neurons in the central nervous system (CNS) and a subset of peripheral sensory neurons [10, 12–17]. FRUM neurons are distributed throughout the brain, ventral nerve cord, and peripheral nervous system, in regions previously implicated in male courtship behavior [18–23].
Based on the expression pattern of FRUM and the fact that individual steps of courtship and copulation behavior are differentially affected in specific fru mutant genotypes, FRUM function fits both necessary and sufficiency criteria as a regulator of the development and function of neurons that participate exclusively in neuronal circuits used during male courtship and copulation behavior [1, 3, 6]. For example, males lacking FRUM exhibit abnormalities specifically in sexually dimorphic behaviors (for example, [8, 9, 11, 13, 24–29]). When paired with females, mutant males with a complete loss of FRUM function do not produce male courtship behaviors, such as courtship song and attempted copulation, however these males do show some male-male orientation and following behaviors, termed chaining [8, 11, 30]. Males bearing weaker fru mutations display courtship, but have reduced fertility including copulation and sperm transfer defects [26, 30, 31]. Thus, it has been inferred that subsets of FRUM neurons are organized in circuits for the execution of behavioral subroutines, whereas other FRUM neurons act as command neurons exerting more global control over the timing or sequence of male sexual behaviors. Recent studies using enhancer trap GAL4 insertion lines to deplete the expression of FRUM in subsets of neurons support this model of nervous system organization [32–34]. Although neurons with roles in some aspects of courtship behaviors have been identified, the role of most FRUM-expressing neurons in male-specific behaviors, including later behaviors like copulation, has not been established.
To determine the role of other FRUM neurons in male reproductive behavior, we identified and characterized a small subset of neurons by their co-expression with Engrailed (En), a homeodomain transcription factor [35, 36]. En has well-known functions in patterning the posterior domains of segments and compartments of imaginal discs (reviewed in [35, 37]). In addition, En contributes to the identity of eight neuroblasts and their progeny in each gnathal, thoracic and abdominal hemisegment and in the brain (reviewed in [38–42]) En neurons are found within regions of the CNS known to be important for courtship behavior [18, 21]. We identified a small cohort of En and FRUM co-expressing neurons distributed in a segmentally restricted pattern in the brain and ventral nerve cord. Depletion of FRUM in these neurons by En-GAL4-driven expression of an inhibitory-RNA transgene directed against male-specific fru transcripts resulted in males that courted females vigorously but were frequently unable to successfully copulate or maintain copulation long enough to transfer sperm and accessory materials. Thus, these En/FRUM neurons form part of a specific neuronal network involved in copulation behaviors, supporting the model in which behavioral subroutines are directed by particular neuronal circuits.
Results
A subset of FRUMneurons is defined by co-expression with En
FRUM neurons contribute to a variety of male-specific reproductive behaviors by their involvement in neuronal circuits mediating courtship and copulation actions. Small groups of FRUM neurons are distributed throughout the brain and ventral nerve cord (VNC; Figure 1A; [8, 11, 12, 15, 17, 28]). En-positive neurons in the brain are confined to: three compact groups located dorso- medio-, and ventro-laterally within the anterior supraesophageal region, four to five individual neurons near the optic lobes, and four to five clusters of neurons in the subesophageal ganglion (SOG). In the ventral nerve cord, En-positive neurons are found coalesced along the ventral midline in the pro-, meta-, and mesothoracic ganglia (T1, T2 and T3, respectively) and in a large ventral group in the abdominal ganglion (AbG, Figure 1B).

FRUMand En are co-expressed in a subset of neurons throughout development. The expression patterns of FRUM and En were analyzed by immunohistochemistry using anti-FRUM (green) and anti-En (magenta) in early pupae through adults and the number of co-expressing neurons counted (Table 1). (A-B) Doubly-labeled 50 hr APF pupal CNS, anti-FRUM (A) and anti-En (B) are expressed and co-expressed (C) throughout the brain and ventral nerve cord. (D) Schematic of En/FRUM co-expression. Pink circles represent En-positive and black-rimmed white circles represent En/FRUM neurons in the brain (box i), T1 midline (arrow, iii), T1-T2-T3 medial cells (arrowheads, ii, iv, v, respectively), and abdominal ganglion (vi). (E-G) In optical image overlays of regions i, iii, and vi (noted in panel D), En/FRUM co-expression appears white inside cells of the brain (E), T1 ganglion (F), and abdominal ganglion (G). (H-J) In the anterior mediolateral brain (Box i) from a 27 hr APF pupa (H), a 50 hr APF pupa (I) and an adult (J), En/FRUM are co-expressed (arrowheads) at all stages. (K-M) In the VNC (arrowheads, ii and iii) from a 27 hr APF pupa (K), a 50 hr APF pupa (L) and an adult (M), FRUM/En are co-expressed (arrowheads) in midline neurons of T1 and medial neurons in T1-T3 at all stages. Images are confocal z-stacks through the entire CNS (A, B, C) or stacks of a subset of z-slices (E-M). Size bar = 100 μm (A) for panels A and B, 20 μm (E, H, K) for panels E-M.
By co-expression of En, we have defined a subset of 53 ± 0.9 FRUM neurons in the brain and VNC, termed En/FRUM neurons (Figure 1C, Table 1; cf. [12, 15]). In the brain, En/FRUM co-expressing neurons were found in nearly half of the approximately 50 En-positive neurons in the medial brain En groups (E/F-brain, Figure 1C, D box i, Table 1), thus, 18–22 En/FRUM neurons in total are detected in the brain. In the VNC, about 33 En/FRUM neurons are detected. The greatest number of En/FRUM-expressing neurons is found as a compact group of about 14 neurons at the midline of the first thoracic ganglion (E/F-VNCmid, Figure 1C, D arrow iii, E, J-L; Table 1). Distinct from the midline group is a set of 3–4 large medial co-labeled neurons in each of the three thoracic ganglia (E/F-VNCmed, Figure 1C, D arrowheads ii, iv, v). In the abdominal ganglion, there are a small number of co-labeled neurons, usually flanking the midline (E/F-AbG, Figure 1C D box vi, G). Thus, we have identified roughly 53 En/FRUM co-expressing neurons distributed among brain and VNC populations of En neurons.
The expression of En developmentally precedes that of FRUM in En/FRUM neurons whether visualized by fru-GAL4 expression pattern or anti-FRUM antibody label. By 8 hours after puparium formation (APF), anti-FRUM antibody labels E/F-brain and E/F-VNCmid, and by 12 hours APF the E/F-VNCmed and E/F-AbG (data not shown). The number of anti-FRUM neurons labeled from early pupal stages into adulthood is consistent, suggesting that a population of neurons has persistent FRUM expression at levels high enough to be detected by the anti-FRUM antibody (Figure 1H-M). Using a GAL4 driver that recapitulates endogenous FRUM expression, fruP1-GAL4 driving membrane-bound GFP (UASmcd8::GFP, FBtp0002652) or a nuclear GFP reporter (UASGFPnls, FBtp0001204) with anti-En staining, we confirm the 53 En/FRUM co-expressing neurons. This pattern is also detected with fruP1.LexA[32] driving expression of GFP (data not shown).
The anti-En antibody recognizes both En and the closely related Invected (Inv) proteins [43–45]. In the embryo, some neurons express only one of these proteins. We assessed whether En/FRUM neurons in the adult expressed both of these proteins by labeling male CNSs from enXho2-lacZ animals and invXba2-lacZ animals with anti-βgal and anti-FRUM. We found complete congruence of FRUM expression in En and Inv neurons suggesting that En/FRUM neurons produce both En and Inv proteins (data not shown).
We further analyzed the En expression pattern in the CNS and other tissues to define the full pattern of En and FRUM co-expression. The neurons expressing En-GAL4 completely overlapped with the anti-En antibody showing that the driver line recapitulates the normal En pattern in the CNS (data not shown), consistent with other results using the same En-GAL4 driver and anti-En antibody [46].
Outside the CNS, En-GAL4 expresses in the En pattern in the posterior epidermis of each body segment, two direct flight muscles, the posterior compartment epithelia of imaginal discs and their adult derivatives, including the genitalia, and sensory neurons derived from the epithelia of the antenna, legs, and genitalia (cf. data not shown). Peripheral sensory neurons in a variety of imaginal derivatives express FRUM proteins at least transiently [15–17, 27, 28, 47]. To determine whether we could find peripheral neurons that co-expressed FRUM and En, we examined external tissues, including genitalia, legs, proboscis, antenna, abdominal body wall and thoracices, from pupal and adult En-GAL4-membrane GFP animals that were labeled with both anti-En and FRUM antibodies. We did not find co-expressing sensory neurons at the stages we examined (data not shown). Recent studies have shown that En is expressed in the anterior lobe of the male genital disc that contributes to the development of the internal genitalia and is not expressed in the parts of the disc that contribute to the male external genitalia. Instead, cubitus interruptus is expressed in the region from which the genital arch, lateral plate, clasper and hypandrium are derived and from which the FRUM-positive sensory neurons are produced [48]. Thus no co-expression of FRUM and En is detected outside of the CNS.
Expression of fruM-RNAi selectively depletes FRUM within En/FRUMneurons
Expression of two copies of a fruM-inhibitory RNA (fruM-RNAi) transgene construct (UAS-fruMIR[29]) driven by En-GAL4 was sufficient to reduce FRUM to very low levels in En/FRUM neurons in males raised at 29°C (En-fruM-RNAi males, Figure 2). The ratio of anti-FRUM pixel intensity in depleted En/FRUM neurons to that of adjacent control FRUM-only reference neurons was used to normalize data for neurons sampled in each group (see Methods). In En/FRUM neurons from wild-type males, the anti-FRUM signal was lower than the reference neuron for all neuronal groups. In En-fruM-RNAi males, FRUM expression was significantly reduced in all En/FRUM neurons compared to wild-type levels with decreases of 86% in E/F-brain, 81% in E/F-VNCmid, 77% in both E/F-VNCmed and E/F-AbG (Figure 2). This reduction was significant for neurons in all regions (p < 0.001 for E/F-brain and all E/F-VNC neurons, p < 0.005 for E/F-AbG neurons analyzed by paired t-tests after arcsine transformation of the ratios). En expression begins prior to FRUM expression in all En/FRUM groups. We measured a significant reduction in FRUM levels in 2–3 day old En-fruM-RNAi males suggesting that in depleted adults, the manipulated En/FRUM neurons developed and functioned with only about 14-20% of normal FRUM levels.

FRUMis significantly reduced in En-fruMRNAi males. CNSs of wild-type (A, C, C’ E-G) and En-fruMRNAi (B, D, D’, H-J) males labeled for anti-FRUM (green) and anti-En (magenta) expression. (A) FRUM expression in E/F-brain neurons. (B) FRUM is extremely reduced in the E/F-brain neurons of En-fruMRNAi males. In the brain, FRUM is reduced by 86% (n = 3, compare A to B, see results for methods). (C) FRUM expression in E/F-VNCmid T1 (arrow) and E/F-VNCmed T1 and T2 (arrowheads) neurons. (D) FRUM is reduced by 81% in midline (n = 3) and by 77% in medial (n = 3, arrowheads) neurons in En-fruMRNAi males. (E-G) FRUM expression in ventral (E), ventromedial (F) and medial (G) AbG neurons (arrows). (H-I) FRUM is reduced by 77% in AbG neurons (n = 3) of En-fruMRNAi males. To clearly show that FRUM has been reduced in all AbG neurons, panels E, F, and G are at three different z-axis positions of the AbG in a WT male and panels H, I, and J are at similar z-axis positions in an En-fruMRNAi male. Arrowheads in panels H-J denote neurons that appear to co-express En/FRUM (but do not) because of overlap of two separate neurons in adjacent z-sections. (C’-D’) Examples of FRUM-only reference neurons (arrowheads) and En/FRUM neurons (arrows) are shown in panels C’ (WT male) and D’ (En-fruMRNAi male). Images are either confocal z-stacks (A-D, E-J) or overlays of single z-sections (C’, D’). Size bars = 20 μm (A) for panels A-D, and (E) for panels E-J.
En/FRUMneurons function in male fertility
Fewer En-fruM-RNAi males were fertile when housed with several virgin females for one week at 29°C compared to control males (61%, Table 2). In vials with fertile En-fruM-RNAi males, we observed that many progeny were produced suggesting that at least some individual males were able to mate and were apparently as fecund as control males. As a second approach to deplete FRUM levels, we used males expressing UAS-GAL4 in addition to drive the UAS-fruM-construct, and found that fewer En-fruM-RNAi/UAS-GAL4 males were fertile compared with control males. Males of two of the three En-fruM-RNAi/UAS-GAL4 lines were additionally less fertile than En-fruM-RNAi males (Table 2). To assess the relationship between mating frequencies in single pair tests to fertility results from week-long tests, twenty-three individual En-fruM-RNAi males were transferred directly from the 10-minute courtship assay to food vials with 2–3 virgin females for a one-week general fertility test. Only fourteen of these males (61%) were fertile. The percentages of fertile En-fruM-RNAi and control males from this experiment were the same as that found in the original one-week fertility tests. Failures in courtship and copulation account for most of the sterile phenotypes of fru mutant males, however, fertile matings do occur in certain hypomorphic genotypes associated with copulation abnormalities [6, 30, 31, 49].
Males with low FRUM levels in En/FRUMneurons have normal courtship but fail to copulate
To test the hypothesis that En-fruM-RNAi males produced abnormal courtship, we paired individual males with single females in small courtship chambers (see Methods). These males exhibited the full range of courtship behaviors, including orientation to and following the female, wing extension and vibration, licking, and tapping. By two important criteria, the courtship of En-fruM-RNAi males was robust: 1) These mutant males initiated courtship with the same latency as WT and control males (Figure 3A); 2) The courtship index (CI) of En-fruM-RNAi males, a measure of the amount of time spent courting, was not significantly different from WT and control males (Additional file 1: Table S1). However, fewer En-fruM-RNAi males mated (57%, Table 2). Likewise, fewer En-fruM-RNAi/UAS-GAL4 males mated (19%, 35%, 55% En-fruM-RNAi/UAS-GAL4-I, En-fruM-RNAi/UAS-GAL4-II, En-fruM-RNAi/UAS-GAL4-III, respectively; Table 2). These En-fruM-RNAi males did attempt copulation with an average of 12 unsuccessful copulation attempts with 21% of these males making over 20 attempts. By comparison, WT and control males average 1.5 attempts and 40% of WT and control males were successful on their first attempt (13/35 WT; 17/40 En-GAL4/+; 13/38 UAS-fruMRNAi /+; Figure 3B). These data confirm that En-fruM-RNAi males produce very active courtship and suggest that male sterility is due at least in part to the failure to make the transition from male–female genital contact to a stable copulation stance. Further, these findings demonstrate that the copulation defects in En-fruM-RNAi males are due specifically to depletion of FRUM in En neurons and not to genetic background effects. Moreover, these data suggest that further reduction of FRUM leads to more severe, but not additional, copulation phenotypes.

Courtship and copulation phenotypes of males expressing a FRUMinhibitory RNA transgene compared to control males. Measurements from 10-minute videotaped courtship tests (see Methods) include A) Latencies to courtship initiation (black), first attempted copulation (white) and copulation (stippled), B) Number of copulation attempts, and C) Copulation duration. The number of animals tested (n) is shown above each bar on the graph (A) and genotype labels are indicated beneath graph (C). For genotypes missing stippled bars in (A) or black bars in (C), none of those males achieved copulation. En-fruMRNAi males’ values for Latency to courtship initiation (A-black) are not statistically different from controls (p = 0.265). The average durations for En-fruMRNAi males measured as Latency to the first attempted copulation (A-white), Latency to copulation (A-stippled) are significantly different from durations measured for En-Gal4/+ and fruMRNAi/+ controls (p < 0.001)*. En-fruMRNAi males have more copulation attempts (B) and shorter copulation durations (C), which are significantly different from values for all other genotypes (p < 0.001)**. All data are shown as mean ± SEM.
Expression of FRUMin En neurons is necessary for maintenance of successful copulations
In the course of these courtship/copulation experiments, we discovered that En-fruM-RNAi males often had abnormal courtship durations compared to control males (Figure 3C). The average copulation duration for control males was around 16 minutes, consistent with previously published results [30], while En-fruM-RNAi males mated with a wide range of copulation durations. The mean duration was11 minutes, which is significantly different from that of controls (Figure 3C).
To understand better the copulation phenotypes of En-fruM-RNAi males, copulation durations were measured in a separate set of experiments (Figure 4A-C). Control and En-fruM-RNAi males were paired individually with mature virgin females, allowed 30 minutes maximum to initiate copulation, and duration determined. Nearly 50% of the En-fruM-RNAi males in these experiments failed to mate within the 30 minute period (see Figure 4 legend). For the En-fruM-RNAi males that did mate, there was a wide range of copulation durations, from five minutes to 27 minutes, whereas control males had more uniform copulation durations (13–22 minutes, Figure 4A) similar to the copulation durations observed after 10-minute courtship and copulation tests (see Figure 3C).

En-fruMRNAi males have abnormal copulation durations. En-fruMRNAi males’ copulation durations vary and often failed to transfer sperm and mating plug material. (A) Scatter plot of individual copulation durations. Mean copulation durations with wild-type females ± SEM are WT, 16.3 ± 0.6 min; En-GAL4/+, 15.7 ± 0.4 min; fruMRNAi/+, 18.3 ± 0.7 min; En-fruMRNAi, 15.2 ± 2.2 min. (B) Individual durations on two consecutive days, each with a fresh wild-type female. For controls, mean ± SEM duration on day 1 and day 2 was not statistically different between genotypes or days (p = 0.622); WT = 16.2 ± 0.8 min, 15.3 ± 0.8 min; En-GAL4/+ = 17.2 ± 0.7 min, 17.0 ± 0.5 min; fruMRNAi /+ = 16.0 ± 0.6 min, 16.4 ± 0.7 min. Durations for En-fruMRNAi males were different between day 1 and day 2, 11.0 ± 2.5 min, 4.2 ± 1.6 min; four En-fruMRNAi males did not mate on either day. Regression analysis for Day 1 versus Day 2 durations show that Day 1 duration does not predict Day 2 duration: control genotypes grouped, R2 = 0.006, p = 0.744; En-fruMRNAi males R2 = 0.137, p = 0.193. (C) Sperm and mating plug transfer was assessed in relation to copulation duration. WT (n = 32) and En-fruMRNAi (n = 38) males were used; all males that copulated are shown. Here, 84% of WT males mated with a mean duration of 16.5 ± 0.5 minutes, compared to only 55% of En-fruMRNAi males mated, with a mean duration of 7.5 ± 0.9 minutes. Males that transferred sperm and a mating plug are filled symbols (both genotypes). In the case of En-fruMRNAi males, those that transferred a mating plug, but no sperm are open symbols and those that did not transfer sperm or a mating plug are gray symbols.
Because of the variability in the copulation durations measured for En-fruM-RNAi males, we determined whether individual males consistently had short duration copulations or repeatedly failed to copulate by measuring the durations of serial copulations for a given male (Figure 4B). Of the fourteen males tested, only three En-fruM-RNAi males mated on consecutive days, seven males mated on only one of the two days and four males did not mate either day. By contrast, all control males mated on both days. The mean copulation durations for En-fruM-RNAi males were 11.0 ± 2.5 minutes on day 1 and 4.2 ± 1.6 minutes on day 2, significantly shorter than control values on both days, around 16 minutes (Figure 4B). Thus, the copulation phenotype of En-fruM-RNAi males shows both variable expressivity and penetrance (Figure 4).
Our data raised the possibility that some En-fruM-RNAi males might be sterile due to the failure to copulate long enough to transfer sperm, seminal fluids, or both components to females [50]. To more thoroughly assess the relationship between copulation duration and fertility, females from matings with known copulation durations (Figure 4B) were transferred singly to food vials. Females mated with control males (n = 20) for durations ranging from 12 to 21 minutes produced offspring. Only two matings by control males resulted in females that did not produce offspring, and both of these males had fertile matings on the second day. Of the females that mated with En-fruM-RNAi males (Day 1, n = 8; Day 2, n = 4), only two females produced offspring and these mated for 16 and 20 minutes (Figure 4B). When the reproductive tracts of females that mated but did not produce offspring (n = 10) were dissected, no sperm was present (data not shown). Seven of these sterile copulations were shorter than the 7–10 minute period when sperm is normally transferred [50] however sperm was not transferred during some copulations that lasted for 11, 15 or 17 minutes, well within the range of successful copulation durations of control males.
To determine the timing for transfer of components during copulation, reproductive tracts were removed within 60 minutes after copulation and viewed under fluorescent optics to visualize mating plug material and by differential interference contrast (DIC) optics to visualize sperm (Figure 4C). Control males had copulation durations of 12 to 25 minutes and always transferred both the sperm plug and sperm to the female. En-fruM-RNAi males that mated with copulations of three minutes or less did not transfer either sperm or sperm plug material, males that mated with copulation durations between 4–11 minutes only transferred sperm plug material and males that mated with copulations lasting at least 13 minutes transferred both sperm and sperm plug material. These times are in general agreement with the timeline of when components are transferred during wild-type copulations [50].
Examining the data regarding both the fertility and the transfer of sperm and seminal fluids, 30% of En-fruM-RNAi males that mated with wild-type durations (≥12 minutes, n = 10) did not transfer sperm, even though these En-fruM-RNAi males manufactured apparently wild-type levels of sperm plug material and motile sperm (Additional file 2: Figure S1). These findings suggest that some copulating En-fruM-RNAi males fail or have delays transferring sperm and mating plug material compared with controls, but generally, if copulation lasted long enough, both sperm and sperm plug material could be transferred by these males. We find that although En-fruM-RNAi males make normal levels of sperm and accessory fluids, they have several defects associated with achieving a successful/stable copulatory position as well as differences in the timing or failure to transfer sperm and accessory material transfer, resulting in a higher frequency of sterile matings. These data suggest that only a small fraction (23%) of copulations by En-fruM-RNAi males would be expected to be fertile. When coupled with the high rate of failed copulation attempts, typically 50-57%, it is surprising that males reach an overall fertility rate of 60 percent (Table 2). We suggest that over the one-week period for the fertility test, males must have copulated at least twice to reach frequency of 61% fertility.
Although there was no overlap of FRUM and En protein expression in the peripheral nervous system, we carried out an additional control to address the possibility that the copulation phenotypes could be due to disruption of peripheral nervous system function. En-fruM-RNAi/Cha-GAL80 males were generated, with the Cha-GAL80 transgene acting to repress GAL4 activity in cholinergic neurons, including all primary sensory neurons [51]. Thus in En-fruM-RNAi /Cha-GAL80 males, the FRUM-RNAi construct is not transcribed in the peripheral sensory neurons, leading to wild-type FRUM levels in peripheral neurons but depleted FRUM in central En/FRUM neurons En-fruM-RNAi/,Cha-GAL80 males were paired individually with wild-type virgin females and examined for courtship and copulation (see Methods). These males did not have courtship defects but did have abnormalities in copulation and fertility, similar to the phenotypes of En-fruM-RNAi males (Figure 3, Table 2). This finding indicates that the copulation phenotypes of En-fruM-RNAi males are not due to FRUM depletion in the peripheral nervous system but to a defect originating within the CNS neurons described here.
The copulation defects and sterility in En-fruM-RNAi males are not due to abnormal locomotor activity levels or abnormal reproductive structures
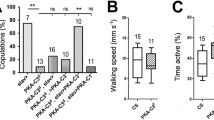
Post-mating sterility of certain fruitless mutant males has been linked to defects in a group of serotonergic neurons in the dorsal posterior abdominal ganglion, which are either missing or fail to express serotonin in females and fru mutants [26, 49]. In a wild-type male, the axons of these serotonergic neurons project down the main abdominal nerve to form terminal arborizations on the accessory glands, testicular ducts, seminal vesicles, vas deferens, and anterior ejaculatory duct [26]. To determine whether the sterility of En-fruM-RNAi males might be due to loss of this serotonergic innervation, reproductive tracts were labeled with anti-serotonin antibody and serotonergic nerve terminals were present and appeared to be at wild-type levels on the same set of reproductive structures (Additional file 3: Figure S2). Therefore, the defects in copulation in En-fruM-RNAi males do not appear to be due to defects in the serotonergic innervation on the male internal reproductive organs.
To rule out the possibility that En-fruM-RNAi males had some defect in general activity or a physical defect in sex-specific and/or reproductive structures that might be responsible for the copulation phenotypes, the locomotor activity levels and anatomy of En-fruM-RNAi were compared with controls. There were no differences in overall activity of En-fruM-RNAi compared to control males measured over a twelve day period (p = 0.214, One-way ANOVA); En-fruM-RNAi males (n = 8) made 18.7 ± 1.9 line crossings per half hour, compared to 15.2 ± 1.6 for En-GAL4/+ (n = 8) and 15.2 ± 1.1 for fruMRNAi /+ males (n = 8). In addition, since En-fruM-RNAi males perform courtship as robustly as control males as measured by CI (Additional file 1: Table S1), it is unlikely that reduced locomotion accounts for the lower copulation success of En-fruM-RNAi males.
Finally, the external and internal genital cuticular structures and abdominal-genital musculature of control and En-fruM-RNAi males were examined and no differences were found in the morphology of these structures or in the Muscle of Lawrence (MOL), a male-specific dorsal abdominal muscle that is missing or defective in fru mutant males (data not shown).
Females and fru mutant males have homologues to WT male En/FRUMneurons
Having identified a set of En/FRUM neurons in males, we considered the possibility that these neurons might be present exclusively in males as found for some anterior brain neurons [33] or whether females and fru mutants might also have these neurons. We used two approaches to assess the presence of these neurons: 1) we counted the number of En neurons at different developmental stages in males, females and fru mutants (Table 3), and 2) we labeled CNSs from En-GAL4 expressing membrane-bound GFP with anti-FRUM (Figure 5).

En/FRUMneurons have the same initial projections in males, females, and fru -mutant males. CNSs of wild-type males (B, F), wild-type females (C, G), and fru-mutant males (D, H) were labeled for anti-FRUM (green) and En-GAL4 expressing membrane-bound GFP (magenta). (A, E) Engrailed neuronal projections in the brain and VNC of wild-type males. Bracketed regions indicate zoom areas for panels B-D and F-H. (B) E/F-brain neurons appear to be in two distinct groups, which project anteriorly, then bifurcate. The same neurons are seen in females (C) and fru-mutant males (D) which show the same projection pattern but do not express FRUM. (F) E/F-VNC neurons in T1 include the midline group in T1 and the medial groups in T1 and T2. Although the projection patterns are difficult to discern, similar neurons are clearly present in females (G) and fru-mutant males (H). Size bars = 20 μm (D) for panels B-D, and (F) for panels F-H.
We counted the numbers of En-positive neurons labeled by anti-Engrailed antibody in the medial brain and first thoracic segment (T1) of the VNC at several pupal stages and in adults. In both sexes and fru mutants, the number of En neurons increases during larval and early pupal stages of development (Table 3). There were no consistent statistical differences in the numbers of En neurons between males and females or between WT and fru mutant animals at any developmental stages (Table 3), indicating that FRUM does not affect the number of En-positive neurons. This is consistent with the finding that FRUM expression begins at 8 hr APF in neurons that already express En.
Examination of CNSs from En-membrane GFP males labeled with anti-FRUM antibody revealed overlap of Engrailed and FRUM neurons in similar locations and numbers in females and fru males as found in wild-type males (Figure 5). We additionally examined the neurochemical profile of the En/FRUM neurons, to look for any difference between sexes or in fru mutants. We labeled adult CNSs with antibodies to gamma-Aminobutyric acid (GABA) or Glutamic acid Decarboxylase (GAD), an enzyme used in GABA synthesis. Most Engrailed neurons are GABAergic, including those that co-express FRUM (Additional file 4: Figure S3), even in females and fru mutant males. Because some En neurons may also be serotonergic, we also labeled CNSs with serotonin (5HT) or used a GAL4 driver that expresses in Dopa Decarboxylase (Ddc)-positive neurons [51, 52]. We found that the E/F-VNCmed neurons are labeled with Ddc-GAL4-driven expression but not 5HT in wild-type males. In females and fru mutant males, the Ddc-GAL4 expression was much weaker and less consistent, noted by the fact that the staining was lighter in these animals and depending on the CNS, not all four of these cells could be observed routinely in each thoracic segment in females and fru males (Additional file 4: Figure S3).
In sum, these data confirm that females and fru mutant males have the same set of En neurons, which are equivalent to the En/FRUM neurons in males. Thus, the presence of the same cohort of En neurons in females means that the male-specific functions of En/FRUM neurons must be due to their sexually dimorphic differentiation or function as opposed to the absence of these neurons in females.
Discussion
En-fruM-RNAi males have mutant phenotypes similar to phenotypes of known frumutants
We have shown that FRUM expression in En/FRUM neurons is necessary in order for males to reliably initiate and maintain copulation leading to frequent male sterility. These copulatory phenotypes have also been described for certain hypomorphic fru mutants, even though the phenotypes are often more severe than those of En-fruM-RNAi males. For example fru1, fru3, and fru4 homozygous mutant males never attempt copulation and also fail to produce, or have abnormalities in, courtship song [53, 54]. Some fru males, for example fru1 homozygotes, perform male-male courtship, a phenotype that is not present in En-fruM-RNAi males. Other fru mutant combinations, however, generate males with behavioral phenotypes that are similar to En-fruM-RNAi males. Hypomorphic fru mutant males that are able to copulate, for example fru1/fru3, have extremely long copulation durations, and often fail to transfer either or both sperm and seminal fluids [26, 31]. Interestingly, fru1/fru3 males lack expression of FRUM in about 50% of AbG neurons [49]. fru mutant males, including fru1/fru3 males, have defects in terminal serotonergic neurons that project out of the central nervous system [49]. Since these serotonergic neurons in the terminal ganglion do not express En, we did not anticipate inactivation of FRUM in these neurons. As expected En-fruM-RNAi males had serotonergic nerve terminals on their reproductive tracts indicating that these neurons were producing their expected neurotransmitter, thus behavioral effects are due to upstream neurons. Although many fewer neurons have lost FRUM expression in En-fruM-RNAi males than in, for example, fru1/fru3 males, the similarity in copulation defects suggests that the same neuronal circuits are affected in these genotypes. Thus using a targeted reduction of FRUM in a defined set of neurons, we have been able to identify a small population of En neurons in the CNS that function to provide robustness in the neuronal circuit that mediates a specific subroutine of male copulation. In normal males this is contingent on the production of early courtship behaviors and was inaccessible in fru mutants that show no courtship.
Phenotypes of En-fruM-RNAi males are similar to other copulation mutants
Other genes besides fru have been implicated in the regulation of male copulation. Mutants that have phenotypes similar to En-fruM-RNAi males, in that they alter the duration of copulation, include stuck (sk, [55]), coitus interruptus (coi, [55]), okina and fickle (fic, reviewed in [56]), lingerer (lig; [57]). Recently, males with mutations in the period (per) and timeless (tim) genes were also found to have longer than average copulation durations [58]. Male fic mutants have phenotypes most similar to En-fruM-RNAi males in that they initiate, but do not maintain, copulation [59]. These fic males have difficulty in sustained aedeagus extension via the protractor muscles, although these males also have internal structural problems with the apodeme [59]. We detected no anatomical cuticular or muscular defects in En-fruM-RNAi males; however, lack of neuronal integration to sustain protractor muscle contraction is a possibility. In a few cases, En-fruM-RNAi males were observed to have difficulty disengaging from the female prior to the end of copulation (KLL unpublished observations), a phenotype associated with sk and lig mutants. Males expressing the effects of hypomorphic lig mutations have difficulty terminating copulation, and interestingly, lig null males make many attempts but never achieve copulation [57]. lig encodes a set of cytoplasmic proteins expressed in cells of the CNS, and so may be expressed in En/FRUM neurons[60]. Thus, lig and fru may work in at least partially overlapping sets of neurons to regulate copulation.
En/FRUMneurons functions as interneurons within a copulatory neuronal circuit
En/FRUM neurons are likely to be interneurons, since their processes do not leave the CNS, and based on their position, size, and similarity to En neurons in other insects. These En interneurons could belong to one or more classes of interneurons including 1) sensory interneurons processing incoming primary sensory information, 2) pre-motor interneurons that contribute to the activity of motorneurons affecting copulation behavior, 3) neurosecretory cells that modulate the activity of other neurons via release of neurochemicals, or 4) neurons forming part of a descending control pathway in which higher centers in the CNS influence the function of neurons in the thoracic and abdominal ganglia [52, 61–63].
The examination of En neurons throughout development and the timing of FRUM expression suggests that most of the FRUM/En neurons are born post-embryonically, as opposed to embryonic neurons that persist into adulthood and are remodeled for adult-specific functions. Based on the small numbers of embryonic En neurons in the brain, the increase in neurons added during larval and pupal development, and their small size, the E/F-brain and E/F-VNCmid are very likely post-embryonic. With these properties, we speculate that these neurons function as local circuit interneurons within a single ganglion. E/F-brain neurons likely belong to the MC2 lineage described by Kumar et al. [64] based on their location and projection pattern. E/F-VNCmid are almost surely progeny of the median neuroblast, lineage 0 [65], Drosophila homologues of neurons identified in the grasshopper as neuronal progeny of the median neuroblast. These neurons express En, use the neurotransmitter gamma-aminobutyric acid (GABA), and are spiking local-circuit interneurons [52, 61–63] . At the stages for which we could detect FRUM expression, these midline clusters are part of much larger En groups, approximately 120 neurons in T1, representing multiple en-positive lineages [61].
We expect that the local circuit interneurons in the prothoracic ganglion coordinate the movement of the legs and/or wings and that the local circuit interneurons in the abdominal ganglion mediate bending of the abdomen and/or activity of the reproductive organs. A previous study of FruM-teashirt co-expressing local circuit interneurons suggested that these neurons were the primary source of FRUM function in directing courtship song phenotypes [66]. Because the latency to the first copulation attempt were not different between the En-fruM-RNAi and control males, it does not appear that the thoracic En/FRUM neurons are necessary components of the FRUM-teashirt courtship song neuronal circuit. The En/FRUM neurons in the brain may not be directly involved in copulatory behavior since they are found in regions deemed necessary for male courtship but not copulation behavior [15, 16], however we cannot rule out an indirect role in copulation behavior for E/F-brain neurons.
Based on the larger size of E/F-VNCmed neurons and the presence of few post-embryonic neuroblasts, these and E/F-AbG may be embryonic. These larger, E/F-VNCmed neurons may be part of interganglionic interneuronal circuits [63]. As interganglionic neurons, we speculate that these neurons may have a role in coordination of local neuronal circuits, such as facilitating the movement of legs and abdomen for successful copulation [52, 63, 64]. Given these two different types of interneurons (local circuit and interganglionic), FRUM may have different roles in the sex-specific differentiation in these two classes of neurons. From our studies, we speculate that the loss of FRUM function in these larger interganglionic neurons maybe the reason that En-fruM-RNAi males are less successful at copulation and have reduced fertility.
Conclusions
We have identified a small subset of FRUM neurons distributed in the brain and ventral cord of males by their co-expression with in En neurons. FRUM expression begins during early to mid-pupal period in neurons already expressing En. Most En/FRUM neurons have a distinctive segmental pattern and contribute to only a part of an En neuronal lineage in the prothoracic midline, anterior medial brain or abdominal ganglia. Only in the four large medial neurons is FRUM expressed in the T1, T2, and T3 segmental homologs. Our data further suggests that the En/FRUM neurons are not unique to males but present in females and fru mutant males. The En/FRUM neurons in males are GABAergic or show Ddc-GAL4 expression indicating that for these characteristics they do not differ from other En neurons in males or females. Based on these observations, we suggest that FRUM likely functions in some aspect of sex-specific differentiation of these neurons, perhaps in their physiology or distal projections, that makes them different than other neurons in the lineage in males and the homologs in females.
Our data strongly support a model in which different groups of FRUM neurons regulate different aspects of courtship and copulation behavior [1, 3, 4]. In such a model, the nervous system is modular, with designated clusters of interconnected neurons responsible for particular behavioral outputs. It is interesting to speculate what behavioral functions might be served by the En/FRUM neurons in females or the consequence of mis-expression of FRUM in these neurons. Female flies in which FRUM is expressed in all of the fru-positive neurons have been shown to produce male courtship behavior but do not have male-like attempted copulation or copulation, perhaps a function of the different size and shape of the abdomen [13, 15]. However, it is possible that these neurons are involved in circuits responsible for female reproductive functions since females also coordinate walking movements with mating and egg-deposition.
Our findings show that the loss of FRUM expression in the small cohort of En/FRUM neurons results in a high frequency of male sterility. Because some En-fruM-RNAi males are fertile, have apparently normal levels of sperm and capable of transferring sperm and sperm plug material to females, male sterility is not likely due to insufficient sperm. Instead, En-fruM-RNAi male sterility is accounted for by a reduced frequency of copulation and/or the failure to adequately transfer sperm and seminal fluids during copulation. Our data from the dual mating experiments also shows that individual males may have a successful copulation and unsuccessful copulation attempts. The lower frequency with which En-fruM-RNAi males were able to successfully mate and be fertile has two potential explanations. One, FRUM activity in these En/FRUM neurons is necessary for fine-tuning the neuronal circuit responsible for copulation and, in its absence, the neuronal circuit has a much lower success rate. Two, residual expression of FRUM in these En/FRUM neurons permits the neuronal circuit to work well enough for some successful copulations to occur. We cannot rule out the second possibility. It is possible that additional experiments expressing UAS-dicer in conjunction with RNAi might lead to a complete loss of FRUM function in these En/FRUM neurons leading to complete male sterility. Our data suggest that the role of FRUM in these neurons is to shift their differentiation to a male-specific fate. Additional studies, at the individual-cell level, will be important to elucidate how the ability to perform courtship and copulation is built into the nervous system during development and how such circuits are maintained and function in the adult fly.
Methods
Fly stocks and crosses
Fly stocks were reared in a 12h: 12h light: dark (12h L/D) cycle at 25°C on standard dextrose medium supplemented with 0.1% Nipagen (p-hydroxybenzoic acid methyl ester; Sigma, St. Louis MO) to inhibit mold. The Canton-S strain, CS-A (from Jeffrey Hall, Brandeis University, Waltham, MA), was the source of wild-type (WT) males and females. To create different fru mutants, we used Df(3R)fru4-40(fru4-40), from which full length non-sexspecific FRU transcripts, encoded by the P3 and P4 promoters, but no sex-specific transcripts are made [30]; Df(3R)frusat15(frusat15), from which fru protein coding sequences are deleted so no fru transcripts are made [11]; and fruP1-GAL4, in which the GAL4 protein coding region is inserted directly downstream of the P1 translational start site, thus blocking the production of FRUM proteins [15]. For FRUM depletion experiments, we used a strain carrying two RNA mediated interference transgenes, UAS-fruMIR/CyO; UAS-fruMIR, which targets the 5′ coding sequences of male-specific fru transcripts [29]. Additional strains for these experiments included three independent recombinant UAS-GAL4, UAS-fruMIR/CyO; UAS-fruMIR, lines I, II, and III (denoted as UAS-GAL4 I, II, III), and a UAS-fruMIR, Cha-GAL80 line (from Devanand Manoli (Stanford University, Palo Alto, CA). The Engrailed and Invected expression pattern, respectively, was determined using the enXho25 and invXba21 lines ([44]; from Chihiro Hama, RIKEN Center for Developmental Biology, Kobe, Japan). We used an engrailed-GAL4 line, en-GAL4e16E (en-GAL4; from Andrea Brand, University of Cambridge, Cambridge, United Kingdom) to drive expression of UAS-GFP-lacZnls and UAS-mCD8GFP reporters (Bloomington Stock Center). For neuronal labeling experiments, we used a Ddc-GAL4 line (from Jay Hirsh, University of Virginia).
Immunohistochemistry
Central nervous systems (CNSs) from sexed larvae, pupae and adults were processed for immunohistochemistry according to standard techniques [42]. For staged pupae, white pre-pupae (0 hr after puparium formation [APF]) were collected and aged at 25°C. CNSs were dissected in Phosphate Buffered Saline (PBS), fixed in 4% paraformaldehyde, washed in PBS + 0.1% Triton-X (PBS-Tx), blocked in PBS-Tx + 10% normal goat serum (NGS), incubated in primary antibody overnight at 4°C and then in secondary antibody for 2 to 4 hrs at room temperature before mounting. To eliminate cross reactivity, CNSs were processed for antigens detected by anti-mouse antibodies, blocked with anti-mouse FAB fragments (Sigma) for one hour then processed for antigens detected by anti-rat antibodies. We used the following primary antibodies: rat anti-FRUM (1:400, [11, 14, 31], rabbit anti-β-galactosidase (βgal) (1:10,000, Cappel, Durham, NC); rabbit anti-serotonin (5HT) (1:500, Sigma); mouse anti-aquorea fluorescent protein (AFP) (1:200, Q-Biogene, Inc., Carlsbad, CA); mouse anti-Engrailed/ Invected (mAb 4D9, 1:5 or 1:10, Development Studies Hybridoma Bank, Iowa City, IA; [42]). Secondary antibodies were conjugated to Alexa-488, -555, -594, or −647 fluorophores (Molecular Probes, Eugene OR), or to horseradish peroxidase (HRP, Jackson ImmunoResearch Laboratories, West Grove, PA) for visualization of the color reaction with diaminobenzidine (DAB, Sigma, St Louis, MO). Fluorescently labeled CNSs were mounted in Prolong (Molecular Probes, Eugene OR) and DAB-labeled preparations were dehydrated in alcohol and mounted in Permount (Sigma, St Louis, MO).
Imaging and image analysis
Confocal images were obtained on a Zeiss 510-Meta confocal scanning microscope while DIC images were captured from an Olympus Vanox-TX microscope with a Sony DKC-5000 digital camera. Images were subsequently processed for contrast using PhotoShop 5.0.2 (Adobe Systems Inc., San Jose, CA).
Neuronal analysis
The number of En neurons was counted in anti-En labeled CNSs visualized by DAB and analyzed by Kruskal-Wallis One-Way Analysis of Variance (ANOVA) followed by Multiple Comparison Procedures, Dunn’s Method (SigmaStat, version 2.03, SPSS Inc., Chicago, IL). Fluorescently labeled neurons were counted from stacks of confocal images.
We assessed the anti-FRUM signal in En/FRUM neurons to gauge the level of FRUM depletion in En-fruM-RNAi adult male CNSs. In single confocal sections, we assigned a pixel intensity value (ImagePro, Media Cybernetics) to the FRUM signal in En/FRUM neurons and a neighboring, distinct FRUM-only neuron, present in the same section, and recognizable in all preparations; the ratio of these values gives a normalized pixel intensity measurement for each En/FRUM neuron. The average FRUM signal was determined for four different neuronal En/FRUM groups in 2 or 3 day old WT (n = 3) and En-fruM-RNAi (n = 3) adult male CNSs and analyzed using paired t-tests after arcsine transformation of the ratios (SigmaStat, Version 2.03, SPSS Inc., Chicago, IL).
Fertility assays
Single virgin males aged for 3–7 days or males used in courtship tests were placed with 2–4 virgin CSA females. Fertility was scored by the presence of progeny after seven days.
Courtship assays and data analysis
Males for the behavioral tests were reared at 29°C on a 12h L/D cycle, collected within 24 hours of eclosion and aged singly at 29°C for 4–6 days to promote the most effective expression of the RNAi transgene [29]. The En-fruM-RNAi males were generated from UAS-fruMIR/CyO; UAS-fruMIR females crossed to en-GAL4/CyO males. For control males, wild-type females were crossed to en-GAL4 males, to UAS-fruMIR/CyO; UAS-fruMIR males or to wild-type males. Virgin CSA female flies were reared at 25°C on a 12h L/D cycle, collected under light CO2 anesthesia within 12 hours of eclosion and aged for 3–5 days en masse.
Courtship assays were performed between 6 to 10 hours after lights on. A female and then a male fly were aspirated into a courtship chamber (1.0 cm diameter × 0.5 cm high) the pair was video-recorded until copulation occurred or for 10 minutes. We measured the latency to courtship initiation (lat Courtin) as the interval after adding the male to his first wing extension. After courtship initiation, the percent of time the male courts is defined as the courtship index (CI; cf[67] ). For males who initiated courtship, the CI was measured for the entire interval between courtship initiation and copulation if the period was less than three minutes, for a three-minute interval prior to copulation or for the last three minutes of the recording period, if males did not copulate. The latency to first attempted copulation (lat 1st Copatt) is the interval between courtship initiation and the first instance of genital-genital contact. The number of attempted copulations (# Copatt) was counted from the onset of courtship through the entire recording period, or until copulation occurred. The latency to copulation (lat Cop) was measured from courtship initiation until the flies achieved copulation. Before statistical analysis, the raw data for lat Courtin and lat 1st Copatt were transformed to the square root of the data and CI and # Copatt were transformed to rank order since these were not normally distributed. The differences between genotypes were analyzed using a Kruskal-Wallis One-Way ANOVA with the source of significant difference determined with Tukey Test for multiple comparisons (Tukey Test; SigmaStat, version 2.03, SPSS Inc.).
Copulation assays
For some experiments, copulation durations were determined for pairs video recorded for 30 minutes. Statistical comparisons of copulation duration intervals were carried out with a Kruskal-Wallis One-Way ANOVA on Ranks, followed by All Pairwise Multiple Comparison Procedures (Dunn’s Method). After copulation, female reproductive tracts were dissected within 60 minutes of copulation and examined for the presence of sperm and mating plug [31, 50].
Activity assays
General locomotion was quantified in a TriKinetic DAMSystem Drosophila Activity Monitor (TriKinetics, Inc., Waltham, MA). Individual males were loaded into single capped tubes and the number of midline crossings was recorded in 30-minute intervals for 12 days at 29°C.
Abbreviations
- 5HT:
-
Serotonin
- AbG:
-
Abdominal ganglion
- APF:
-
After puparium formation
- CI:
-
Courtship index
- CNS:
-
Central nervous system
- Cop:
-
Copulation
- CSA:
-
Canton-S strain A
- Ddc:
-
Dopa Decarboxylase
- En:
-
Engrailed
- en-GAL4:
-
Engrailed-GAL4 e16E insert
- E/F-brain:
-
En/FRUM co-expression neurons in the medial brain Engrailed groups
- E/F-VNCmid:
-
En/FRUM co-expression neurons in the midline first thoracic ganglion Engrailed group
- E/F-VNCmed:
-
En/FRUM co-expression neurons in the medial ventral nerve cord Engrailed groups
- E/F-AbG:
-
En/FRUM co-expression neurons in the abdominal ganglion Engrailed groups
- FRUM:
-
Male-specific products of the fruitless gene
- En/FRUM:
-
Neurons expressing both En and FRUM proteins
- GABA:
-
Gamma-Aminobutyric acid
- GAD:
-
Glutamic acid Decarboxylase
- Lat:
-
Latency
- T1:
-
First thoracic ganglion
- T2:
-
Second thoracic ganglion
- T3:
-
Third thoracic ganglion
- SOG:
-
Subesophageal ganglion
- UAS:
-
fruMIR parent
- VNC:
-
Ventral nerve cord
- WT:
-
Wild-type.
References
Baker BS, Taylor BJ, Hall JC: Are complex behaviors specified by dedicated regulatory genes? Reasoning from Drosophila. Cell. 2001, 105 (1): 13-24. 10.1016/S0092-8674(01)00293-8.
Dauwalder B: The roles of fruitless and doublesex in the control of male courtship. Int Rev Neurobiol. 2011, 99: 87-105.
Dickson BJ: Wired for sex: the neurobiology of Drosophila mating decisions. Science. 2008, 322 (5903): 904-909. 10.1126/science.1159276.
Manoli DS, Meissner GW, Baker BS: Blueprints for behavior: genetic specification of neural circuitry for innate behaviors. Trends Neurosci. 2006, 29 (8): 444-451. 10.1016/j.tins.2006.06.006.
Siwicki KK, Kravitz EA: Fruitless, doublesex and the genetics of social behavior in Drosophila melanogaster. Curr Opin Neurobiol. 2009, 19 (2): 200-206. 10.1016/j.conb.2009.04.001.
Villella A, Hall JC: Neurogenetics of courtship and mating in Drosophila. Adv Genet. 2008, 62: 67-184.
Yamamoto D: Brain sex differences and function of the fruitless gene in Drosophila. J Neurogenet. 2008, 22 (3): 309-332. 10.1080/01677060802298491.
Ryner LC, Goodwin SF, Castrillon DH, Anand A, Villella A, Baker BS, Hall JC, Taylor BJ, Wasserman SA: Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell. 1996, 87 (6): 1079-1089. 10.1016/S0092-8674(00)81802-4.
Usui-Aoki K, Ito H, Ui-Tei K, Takahashi K, Lukacsovich T, Awano W, Nakata H, Piao ZF, Nilsson EE, Tomida J: Formation of the male-specific muscle in female Drosophila by ectopic fruitless expression. Nat Cell Biol. 2000, 2 (8): 500-506. 10.1038/35019537.
Goodwin SF, Taylor BJ, Villella A, Foss M, Ryner LC, Baker BS, Hall JC: Aberrant splicing and altered spatial expression patterns in fruitless mutants of Drosophila melanogaster. Genetics. 2000, 154 (2): 725-745.
Ito H, Fujitani K, Usui K, Shimizu-Nishikawa K, Tanaka S, Yamamoto D: Sexual orientation in Drosophila is altered by the satori mutation in the sex-determination gene fruitless that encodes a zinc finger protein with a BTB domain. Proc Natl Acad Sci USA. 1996, 93 (18): 9687-9692. 10.1073/pnas.93.18.9687.
Lee G, Foss M, Goodwin SF, Carlo T, Taylor BJ, Hall JC: Spatial, temporal, and sexually dimorphic expression patterns of the fruitless gene in the Drosophila central nervous system. J Neurobiol. 2000, 43 (4): 404-426. 10.1002/1097-4695(20000615)43:4<404::AID-NEU8>3.0.CO;2-D.
Demir E, Dickson BJ: fruitless splicing specifies male courtship behavior in Drosophila. Cell. 2005, 121 (5): 785-794. 10.1016/j.cell.2005.04.027.
Billeter JC, Rideout EJ, Dornan AJ, Goodwin SF: Control of male sexual behavior in Drosophila by the sex determination pathway. Curr Biol. 2006, 16 (17): R766-776. 10.1016/j.cub.2006.08.025.
Manoli DS, Foss M, Villella A, Taylor BJ, Hall JC, Baker BS: Male-specific fruitless specifies the neural substrates of Drosophila courtship behaviour. Nature. 2005, 436 (7049): 395-400.
Dornan AJ, Gailey DA, Goodwin SF: GAL4 enhancer trap targeting of the Drosophila sex determination gene fruitless. Genesis. 2005, 42 (4): 236-246. 10.1002/gene.20143.
Cachero S, Ostrovsky AD, Yu JY, Dickson BJ, Jefferis GS: Sexual dimorphism in the fly brain. Curr Biol. 2010, 20 (18): 1589-1601. 10.1016/j.cub.2010.07.045.
Hall JC: The mating of a fly. Science. 1994, 264 (5166): 1702-1714. 10.1126/science.8209251.
Hall JC: Portions of the central nervous system controlling reproductive behavior in Drosophila melanogaster. Behav Genet. 1977, 7 (4): 291-312. 10.1007/BF01066800.
Hall JC: Control of male reproductive behavior by the central nervous system of Drosophila: dissection of a courtship pathway by genetic mosaics. Genetics. 1979, 92 (2): 437-457.
Ferveur JF, Greenspan RJ: Courtship behavior of brain mosaics in Drosophila. J Neurogenet. 1998, 12 (4): 205-226. 10.3109/01677069809108559.
Ferveur JF, Stortkuhl KF, Stocker RF, Greenspan RJ: Genetic feminization of brain structures and changed sexual orientation in male Drosophila. Science. 1995, 267 (5199): 902-905. 10.1126/science.7846534.
O’Dell KM, Armstrong JD, Yang MY, Kaiser K: Functional dissection of the Drosophila mushroom bodies by selective feminization of genetically defined subcompartments. Neuron. 1995, 15 (1): 55-61. 10.1016/0896-6273(95)90064-0.
Meissner GW, Manoli DS, Chavez JF, Knapp JM, Lin TL, Stevens RJ, Mellert DJ, Tran DH, Baker BS: Functional dissection of the neural substrates for sexual behaviors in Drosophila melanogaster. Genetics. 2011, 189 (1): 195-211. 10.1534/genetics.111.129940.
Lee G, Hall JC: A newly uncovered phenotype associated with the fruitless gene of Drosophila melanogaster: aggression-like head interactions between mutant males. Behav Genet. 2000, 30 (4): 263-275. 10.1023/A:1026541215546.
Lee G, Villella A, Taylor BJ, Hall JC: New reproductive anomalies in fruitless-mutant Drosophila males: extreme lengthening of mating durations and infertility correlated with defective serotonergic innervation of reproductive organs. J Neurobiol. 2001, 47 (2): 121-149. 10.1002/neu.1021.
Stockinger P, Kvitsiani D, Rotkopf S, Tirian L, Dickson BJ: Neural circuitry that governs Drosophila male courtship behavior. Cell. 2005, 121 (5): 795-807. 10.1016/j.cell.2005.04.026.
Yu JY, Kanai MI, Demir E, Jefferis GS, Dickson BJ: Cellular organization of the neural circuit that drives Drosophila courtship behavior. Curr Biol. 2010, 20 (18): 1602-1614. 10.1016/j.cub.2010.08.025.
Manoli DS, Baker BS: Median bundle neurons coordinate behaviours during Drosophila male courtship. Nature. 2004, 430 (6999): 564-569. 10.1038/nature02713.
Anand A, Villella A, Ryner LC, Carlo T, Goodwin SF, Song HJ, Gailey DA, Morales A, Hall JC, Baker BS: Molecular genetic dissection of the sex-specific and vital functions of the Drosophila melanogaster sex determination gene fruitless. Genetics. 2001, 158 (4): 1569-1595.
Villella A, Peyre JB, Aigaki T, Hall JC: Defective transfer of seminal-fluid materials during matings of semi-fertile fruitless mutants in Drosophila. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2006, 192 (12): 1253-1269. 10.1007/s00359-006-0154-1.
Mellert DJ, Knapp JM, Manoli DS, Meissner GW, Baker BS: Midline crossing by gustatory receptor neuron axons is regulated by fruitless, doublesex and the Roundabout receptors. Development. 2010, 137 (2): 323-332. 10.1242/dev.045047.
Kimura K, Ote M, Tazawa T, Yamamoto D: Fruitless specifies sexually dimorphic neural circuitry in the Drosophila brain. Nature. 2005, 438 (7065): 229-233. 10.1038/nature04229.
Villella A, Ferri SL, Krystal JD, Hall JC: Functional analysis of fruitless gene expression by transgenic manipulations of Drosophila courtship. Proc Natl Acad Sci USA. 2005, 102 (46): 16550-16557. 10.1073/pnas.0507056102.
Kornberg T: Engrailed: a gene controlling compartment and segment formation in Drosophila. Proc Natl Acad Sci USA. 1981, 78 (2): 1095-1099. 10.1073/pnas.78.2.1095.
Poole SJ, Kauvar LM, Drees B, Kornberg T: The engrailed locus of Drosophila: structural analysis of an embryonic transcript. Cell. 1985, 40 (1): 37-43. 10.1016/0092-8674(85)90306-X.
Hidalgo A: The roles of engrailed. Trends Genet. 1996, 12 (1): 1-4. 10.1016/0168-9525(96)81373-4.
Bhat KM: Segment polarity genes in neuroblast formation and identity specification during Drosophila neurogenesis. BioEssays. 1999, 21 (6): 472-485. 10.1002/(SICI)1521-1878(199906)21:6<472::AID-BIES4>3.0.CO;2-W.
Brower DL: Posterior-to-anterior transformation in engrailed wing imaginal disks of Drosophila. Nature. 1984, 310 (5977): 496-497. 10.1038/310496a0.
Cui X, Doe CQ: ming is expressed in neuroblast sublineages and regulates gene expression in the Drosophila central nervous system. Development. 1992, 116 (4): 943-952.
Bhat KM, Schedl P: Requirement for engrailed and invected genes reveals novel regulatory interactions between engrailed/invected, patched, gooseberry and wingless during Drosophila neurogenesis. Development. 1997, 124 (9): 1675-1688.
Patel NH, Schafer B, Goodman CS, Holmgren R: The role of segment polarity genes during Drosophila neurogenesis. Genes Dev. 1989, 3 (6): 890-904. 10.1101/gad.3.6.890.
Coleman KG, Poole SJ, Weir MP, Soeller WC, Kornberg T: The invected gene of Drosophila: sequence analysis and expression studies reveal a close kinship to the engrailed gene. Genes Dev. 1987, 1 (1): 19-28. 10.1101/gad.1.1.19.
Hama C, Ali Z, Kornberg TB: Region-specific recombination and expression are directed by portions of the Drosophila engrailed promoter. Genes Dev. 1990, 4 (7): 1079-1093. 10.1101/gad.4.7.1079.
Gustavson E, Goldsborough AS, Ali Z, Kornberg TB: The Drosophila engrailed and invected genes: partners in regulation, expression and function. Genetics. 1996, 142 (3): 893-906.
Blagburn JM: Engrailed expression in subsets of adult Drosophila sensory neurons: an enhancer-trap study. IN. 2008, 8 (3): 133-146.
Billeter JC, Goodwin SF: Characterization of Drosophila fruitless-gal4 transgenes reveals expression in male-specific fruitless neurons and innervation of male reproductive structures. J Comp Neurol. 2004, 475 (2): 270-287. 10.1002/cne.20177.
Chen EH, Baker BS: Compartmental organization of the Drosophila genital imaginal discs. Development. 1997, 124 (1): 205-218.
Lee G, Hall JC: Abnormalities of male-specific FRU protein and serotonin expression in the CNS of fruitless mutants in Drosophila. J Neurosci. 2001, 21 (2): 513-526.
Lung O, Wolfner MF: Identification and characterization of the major Drosophila melanogaster mating plug protein. Insect Biochem Mol Biol. 2001, 31 (6–7): 543-551.
Lundell MJ, Hirsh J: Temporal and spatial development of serotonin and dopamine neurons in the Drosophila CNS. Dev Biol. 1994, 165 (2): 385-396. 10.1006/dbio.1994.1261.
Siegler MV, Pankhaniya RR, Jia XX: Pattern of expression of engrailed in relation to gamma-aminobutyric acid immunoreactivity in the central nervous system of the adult grasshopper. J Comp Neurol. 2001, 440 (1): 85-96. 10.1002/cne.1371.
Villella A, Gailey DA, Berwald B, Ohshima S, Barnes PT, Hall JC: Extended reproductive roles of the fruitless gene in Drosophila melanogaster revealed by behavioral analysis of new fru mutants. Genetics. 1997, 147 (3): 1107-1130.
Wheeler DA, Kulkarni SJ, Gailey DA, Hall JC: Spectral analysis of courtship songs in behavioral mutants of Drosophila melanogaster. Behav Genet. 1989, 19 (4): 503-528. 10.1007/BF01066251.
Hall JC, Tompkins L, Kyriacou CP, Siegel RW, von Schilcher F, Greenspan RJ: Higher behavior in Drosophila analyzed with mutations that disrupt the structure and function of the nervous system. Basic Life Sci. 1980, 16: 425-455.
Yamamoto D, Jallon JM, Komatsu A: Genetic dissection of sexual behavior in Drosophila melanogaster. Annu Rev Entomol. 1997, 42: 551-585. 10.1146/annurev.ento.42.1.551.
Kuniyoshi H, Baba K, Ueda R, Kondo S, Awano W, Juni N, Yamamoto D: lingerer, a Drosophila gene involved in initiation and termination of copulation, encodes a set of novel cytoplasmic proteins. Genetics. 2002, 162 (4): 1775-1789.
Beaver LM, Giebultowicz JM: Regulation of copulation duration by period and timeless in Drosophila melanogaster. Curr Biol. 2004, 14 (16): 1492-1497. 10.1016/j.cub.2004.08.022.
Baba K, Takeshita A, Majima K, Ueda R, Kondo S, Juni N, Yamamoto D: The Drosophila Bruton’s tyrosine kinase (Btk) homolog is required for adult survival and male genital formation. Mol Cell Biol. 1999, 19 (6): 4405-4413.
Kuniyoshi H, Usui-Aoki K, Juni N, Yamamoto D: Expression analysis of the lingerer gene in the larval central nervous system of Drosophila melanogaster. J Neurogenet. 2003, 17 (2–3): 117-137.
Kumar A, Fung S, Lichtneckert R, Reichert H, Hartenstein V: Arborization pattern of engrailed-positive neural lineages reveal neuromere boundaries in the Drosophila brain neuropil. J Comp Neurol. 2009, 517 (1): 87-104. 10.1002/cne.22112.
Truman JW, Schuppe H, Shepherd D, Williams DW: Developmental architecture of adult-specific lineages in the ventral CNS of Drosophila. Development. 2004, 131 (20): 5167-5184. 10.1242/dev.01371.
Thompson KJ, Siegler MV: Anatomy and physiology of spiking local and intersegmental interneurons in the median neuroblast lineage of the grasshopper. J Comp Neurol. 1991, 305 (4): 659-675. 10.1002/cne.903050409.
Siegler MV, Pankhaniya RR: Engrailed protein is expressed in interneurons but not motor neurons of the dorsal unpaired median group in the adult grasshopper. J Comp Neurol. 1997, 388 (4): 658-668. 10.1002/(SICI)1096-9861(19971201)388:4<658::AID-CNE11>3.0.CO;2-H.
Jia XX, Siegler MV: Midline lineages in grasshopper produce neuronal siblings with asymmetric expression of Engrailed. Development. 2002, 129 (22): 5181-5193.
Rubinstein CD, Rivlin PK, Hoy RR: Genetic feminization of the thoracic nervous system disrupts courtship song in male Drosophila melanogaster. J Neurogenet. 2010, 24 (4): 234-245. 10.3109/01677063.2010.519805.
Villella A, Hall JC: Courtship anomalies caused by doublesex mutations in Drosophila melanogaster. Genetics. 1996, 143 (1): 331-344.
Acknowledgements
We would like to thank B. Baker, K. Cromack, Jr., M. Foss, J. Fowler, J. Giebultowicz, J. Hall, J. Hatzidakis, Y. Liu, D. Manoli, G. Meisner, A. Villella, and V. Weis, and for readings and comments. B. Baker, A. Brand, T. Carlo, J. Hall, C. Hama, J. Hirsh, M. Landgraf, D. Manoli, N. Patel and A. Villella graciously provided reagents. K. Cromack, Jr., D. Lutterschmidt, and H. Waye gave insight on statistical analyses. C. Beach assisted with activity assays. The Engrailed antibody developed by N. Patel was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biology, Iowa City, IA 52242. FlyBase provided information on stocks, supported by a grant from the National Human Genome Research Institute at the U.S. National Institutes of Health P41-HG000739. This publication was made possible in part by grant number 1S10RR107903-01 from the National Institutes of Health. The authors wish to acknowledge the Confocal Microscopy Facility of the Center for Genome Research and Biocomputing and the Environmental and Health Sciences Center at Oregon State University. This work was funded by NIH-R01 NS33352 to B. Baker, J. Hall, and B. Taylor and NIH RO1-GM085818 to B. Taylor.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interest
We have no conflicts of interest in presenting this manuscript.
Authors’ contributions
KLL designed the study, carried out the fly breeding, immunohistochemistry, microscopy and image analysis, behavioral assays, data analysis, and drafted the manuscript. YSL carried out additional immunohistochemistry and behavioral assays. BJT participated in study design and coordination, and helped to draft the manuscript. All authors read and approved the final manuscript.
Electronic supplementary material
12868_2012_2896_MOESM1_ESM.docx
Additional file 1: Table S1: En-fruMRNAi males have normal courtship index (CI) values. Measurements from 10-minute videotaped courtship tests (see Methods) include courtship index (a measure of time spent performing wing courtship song). All genotypes were not statistically different for courtship index (One-Way ANOVA, p = 0.019). (DOCX 77 KB)
12868_2012_2896_MOESM2_ESM.docx
Additional file 2: Figure S1: En-fruMRNAi males make and store sperm and mating plug material. In dissected reproductive tracts from wild-type (A, B) and En-fruMRNAi (C, D) males, sperm was viewed by differential interference contrast microscopy and mating plug material was visible under ultraviolet light. Sperm and mating plug material levels appeared to be normal, and sperm were motile. (DOCX 75 KB)
12868_2012_2896_MOESM3_ESM.docx
Additional file 3: Figure S2: En-fruMRNAi males have normal serotonergic innervation. Serotonergic nerve terminals innervating the internal reproductive organs were examined in wild-type and en-GAL4/UAS-fruMIR males by immunohistochemistry with anti-serotonin (5HT). A) In a wild-type male, serotonergic nerve terminals are present on the seminal vesicles (sv), accessory glands (ag) and ejaculatory duct (ed). B) In an En-fruMRNAi male, serotonergic terminals are present on the same organs, similar to wild-type males. Images are confocal z-stacks through the male internal reproductive tract. Size bar = 200 um. (DOCX 179 KB)
12868_2012_2896_MOESM4_ESM.docx
Additional file 4: Figure S3: Wild-type males, females, and fru mutant males have similar neurotransmitter profiles for brain and VNC-T1midline neurons, but vary for VNC-medial neurons in T1, T2, and T3. CNSs of wild-type males (A, C, F, G, H), wild-type females (B, D, I), and fru-mutant males (E, J) were labeled for anti-FRUM (green) and anti-Engrailed (magenta), and a neurochemical marker (blue). (A, B) E/F-VNCmid neurons in males, and the equivalent in females, express gamma-Aminobutyric acid (GABA) neurotransmitter as labeled by anti-GABA antibody. (C, D, E) In males, females, and fru mutant males E/F-VNCmid neurons are also labeled by anti-GAD antibody (GAD = Glutamic Acid Decarboxylase, an enzyme for GABA synthesis, localized exclusively to GABAergic neurons). (F, G) E/F-brain (F) and E/F-AbG (G) neurons are also labeled by GAD in wild-type males. (H, I, J) Males expressed a Ddc-GAL4; UAS-mcd8::GFP in E/F-VNCmed neurons of T1, T2 (H) and T3 (not shown). Females (I) and fru-mutant males (J) express this driver much more faintly, in with variable penetrance in the equivalent neurons. (Ddc = dopa Decarboxylase, an enzyme for serotonin/5HTsynthesis). (K, L, M) Schematic indicating neurotransmitter profile of En/FRUM neurons in the brain (K), T1/T2 segments of the VNC (L), and T3 and AbG (M). (DOCX 16 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Latham, K.L., Liu, YS. & Taylor, B.J. A small cohort of FRUM and Engrailed-expressing neurons mediate successful copulation in Drosophila melanogaster. BMC Neurosci 14, 57 (2013). https://doi.org/10.1186/1471-2202-14-57
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2202-14-57