
Abstract
A new locality Kimiltei, Irkutsk oblast, where unmineralized remains of arthropods were buried en masse, is described. The U–Pb zircon dating yielded a Cambrian age with the youngest values of 492 ± 4 Ma. Kimiltei arthropods are represented by Chasmataspidida, Synziphosurina, and Euthycarcinoidea. The last two groups have not been known in the Siberian Platform, and the first one has been described in this area only from Devonian rocks. The Irkutsk findings are the most ancient representatives of their groups, and their distribution areas turn out to be wider than previously known. Their co-burial is noteworthy, because the representatives of these groups have never been found together before. Taking into account the fact that the descendants of all three groups gave rise to the terrestrial lines of chelicerates and insects, the Kimiltei assemblage is the entire fauna of the land arthropod ancestors. Their cohabitation is indicative of the fact that they started their expansion on land from some close ecological niches in the intertidal zone.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
INTRODUCTION
The middle–late Cambrian arthropods (arthropods excluding trilobites are meant) were extremely diverse and widespread throughout the world. However, only few findings are known from deposits of this age in the Siberian Platform. It is hard to imagine that, in contrast to the middle–late Cambrian flourishing of arthropods throughout the world, their diversity and abundance were so negligible in the large paleoarea of the Siberian Platform. It is more logical to assume that they simply have not been studied and described properly. The latter is supported, in particular, by the information on fossil arthropods from the Cambrian of Irkutsk oblast in the southern part of Eastern Siberia. Arthropod findings with an unclear taxonomic status were noted in this area, in five middle–late Cambrian localities, in the course of geological research in the middle of the twentieth century [1, 2]. These data are not taken into account in any way in the world compendiums on the Cambrian fauna. In other words, as many as five localities, covering the impressive geographic region with the corresponding fauna, are completely left out of the discussion on the diversity and evolutionary history of this group. Meanwhile, the middle and upper Cambrian is the period of arthropod formation and radiation, and any information on their diversity is therefore very relevant.
It should be noted that the arthropod findings in Irkutsk oblast are not taken into account by the researchers, primarily because of the lack of any meaningful description of them and clear images [1, 2]. Unfortunately, according to our investgations, the collections from the localities indicated in the publications noted above and assigned to various scientific museums are currently not available or have been lost. Therefore, it is impossible to re-study them. In addition, due to the reorganization of scientific geological institutes, the location of the field logs compiled in the course of the expeditions (listed in [2]) is unknown. These logs recorded the findings of unusual middle–late Cambrian arthropods. We made an attempt to find one of these localities according to the instructions in the publication of V.S. Voropinov [1]. However, all landmarks noted in the publication no longer exist, and they are also absent on the old topographic maps. Hence, the probability of getting to the right points of the old expeditions seems negligible currently. In this regard, new Siberian localities with fossil arthropods of this age are of great interest.
In this paper, we describe arthropods from a new locality situated near the village of Kimiltei in Irkutsk oblast. This locality was discovered in 2006 by a local amateur paleontologist Aleksandr Klepikov. In 2019, he visited this site together with scientists from the Institute of the Earth’s Crust, Siberian Branch, Russian Academy of Sciences, and Borissiak Paleontological Institute, Russian Academy of Sciences, and took active part in the collection of samples.
In this paper, we give descriptions of the findings, which make it possible to assign them to specific taxa (classes and orders). Our identifications turned out to demonstrate very unexpectedly the exceptional importance of arthropod from this area for understanding the entire evolution of non-trilibite arthropods. Due to the fact that the known species of the identified groups—Chasmataspidida, Synziphosurina (early relatives of Chelicerae), and Euthycarcinoidea (insect ancestors)—are common in the Ordovician–Carboniferous, but not in the Cambrian, we considered it necessary to confirm their stratigraphic age using U‒Pb isotope dating of detrital zircon grains. The estimates obtained made it possible to date the sediments as the late Cambrian in the range from 485 to 497 Ma. Hence, already in the Late Cambrian, a whole assemblage of the most ancient ancestors of the future colonizers of the land co-existed in the Kimiltei paleoaquatic area. In contrast to the available data, it turned out that they lived together and in close habitats of shallow lagoons or intertidal pools, so that their descendants began to expand in the coastal zone in parallel with the same initial geographical and ecological position.
SECTION DESCRIPTION AND FACIES FEATURES OF THE LAYER WITH FOSSILS
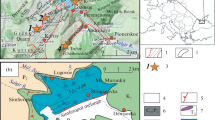
The Kimiltei section is located one and a half kilometers east of Kimiltei (54°9′49.99″ N, 102°1′22.44″ E) in Irkutsk oblast in the southern part of Eastern Siberia (Fig. 1). In the local coastal cliff, Verkholensk rocks are exposed. The Verkholensk Group is widespread within the Irkutsk Amphitheatre of the Siberian Platform [3]. Historically, the Verkholensk Group was subdivided into six successive formations (from bottom to top): Osinsk, Balagansk, Ust’-Talkin, Mikhailov, Ryutin, and Kardin [3, 4]. According to the current concepts, in the Angara–Lena region, the last four are combined into the Baitog Formation [2].

The Kimiltei locality and its structure. (a) A simplified geological map showing the distribution of uneven-aged rocks in the south of the Siberian Platform. Deposits: (1) Jurassic, (2) Silurian, (3) Ordovician, (4) upper Cambrian (Verkholensk Group), (5) early–middle Cambrian, (6) Vendian, (7) Archean–Proterozoic. (b) Geological structure of the Kimiltei section with marks of sampling sites for analysis of detrital zircons (1957, K-21). Deposits: (8) silty sandstone, (9) aleuropelite (siltstone and mudstone).
The Verkholensk Group is predominantly composed of red–brown and variegated rocks characteristic in appearance and composition. Its deposits are composed of sulfate–carbonate terrigenous rocks and carbonates formed in the shallow seashore conditions. This Group covers a long-time interval from the beginning of the Mayan stage of the middle Cambrian to the end of the upper Cambrian [3–5].
The rocks of the Baitog Formation exposed in the Kimiltei section occur as polymictic medium- and fine-grained silty sandstones and sandstones with carbonate cement. Thick sandstone members are rhythmically interbedded with chocolate-brown thin-platy siltstone and claystone interlayers (Figs. 1b, 2a). There are traces of ripples and mud cracks at different levels of the section.

Location and facies features of a lens with fossils. (a) General view of the section indicating the location of a lens with fossils (red color) and samples on panels (b) and (c), (b) mud cracks in silty sandstone, (c) ripple marks in silty sandstone.
The horizon with fossils is located in the upper part of the section (Fig. 2a). It is a thin layer of fine-platy chocolate-brown and sometimes turquoise siltstones and mudstones. It extends along the strike for 30‒40 m and then wedges out. A silty sandstone layer overlies mudstones. A surface of brown claystone and silty sandstones is marked by distinct filled mud cracks (Fig. 2b). At the same stratigraphic level as a lens with fossils, traces of ripples can also be found in some parts of the section (Fig. 2c).
Based on the mineralogy of lens rocks studied by combined calorimetric and thermogravimetric analysis, the silty sandstone layer is mainly composed of quartz (45 wt %) and calcite (30 wt %) with an impurity of kaolinite (10 wt %) and illite (5 wt %). The finely dispersed silty layer consists mainly of kaolinite (35–40 wt %), quartz (17–20 wt %), and chlorite, chlorite–montmorillonite (25–27 wt %). The latter is characteristic of a terrigenous drift. The chocolate shade of the clays is due to goethite and hematite (5 wt %). All layers are depleted in siderite and pyrite. Many arthropod casts have a thin coating of red iron oxides.
Lithofacies features of the layer with fossils are indicative of the fact that it was a small shallow pool most likely cut off from the sea or a large lagoon by a coastal bar. Animals could have been thrown behind the bank during a storm or a high tide, then the water reservoir dried up and exposed the bottom silt. The animals died in one way or another and dried up together with the clay crust: this suggestion is confirmed by an animal “skin” torn by a mud crack into two parts that fit each other (Fig. 4a). After drying, their body remains were soon buried under a fine- and medium-grained silty sand layer which pressed a chitinous skin into a finely dispersed clay matrix. As a result, detailed casts of the adjacent surface of the animals were formed on the clay (mainly the ventral side). The overlying upper layer of fine-grained sand did not interfere with the oxygenation of sediments with the remains of buried animals. As a result, their soft tissues were decomposed and disappeared relatively quickly and iron oxidized in the clay layer formed brown oxides. Decaying organic materials effectively bound Fe-containing products; as a result, a thin reddish coating of oxidized iron was formed on the surface of the casts. Meanwhile, some thin three-dimensional microdetails of the structure, in particular, of chelicerae and spines of pedipalps on one of the studied specimens, could hardly be due to pressing into the clay matrix. Hence, the slower replacement of hard chitinous parts by the surrounding rocks is also relatively possible. When the bottom of the compact silty sandstone layer was exposed in the section, a loosely consolidated mudstone layer crumbled down from its surface, exposing the underside of three-dimensional casts.

Histograms and probability density curves for the age of detrital zircon grains from samples of the Kimiltei section: (a) no. 1957, (b) no. K21, and the Nora section: (c) no. 1974.

Two of nine slabs with numerous arthropod casts. (a) PIN 58915891; specimens 41 and 43 are probably a whole body cut in two by a desiccation crack. (b) PIN 5891: a desiccation crack limits the top (here) edge of the slab.
It should be noted that the late Cambrian remains of shallow-water arthropods are characterized by a similar fossilization type: three-dimensional casts on a clay matrix with fine structural details preserved [6]. However, the data are still insufficient to understand the nature of this similarity: it can be indicative of specific features of the burial time, environmental and taphonomic conditions, or peculiar features of arthropods in this period.
AGE OF FOSSILS IN THE KIMILTEI AREA
As mentioned above, the Kimiltei section is part of the Baitog Formation of the Verkholensk Group commonly considered to be middle–upper Cambrian in age. The Baitog Formation is overlain by the Early–Middle Ordovician Ust’-Kut and Ilga formations. The dates of 512 ± 10 and 502 ± 2 Ma were previously obtained for the Verkholensk Group (review in [5]).
In 2020, sample no. 1957 was taken from the silty sandstone layer with casts to clarify the age of this locality (Fig. 1b). Eighty-nine grains of detrital zircon were analyzed in the sample studied. All dates with a discordance of |D| > 10% were excluded from consideration. The remaining sixty-eight dates were used to construct a histogram and the probability density curve (PDC) of U–Pb isotopic dates from this sample (Fig. 3a). The PDC has a pronounced peak (22 grains) corresponding to the age of 511 ± 6 Ma [5]. The youngest zircon in this sample was 491 ± 6 Ma. In the Early Ordovician, the mountains were actively formed within the Irkutsk Amphitheatre. As a result, more detrital material was accumulated in the sediments, including that from the destroyed Early Ordovician volcanic rocks. Therefore, a peak of zircons corresponding to the Early Ordovician age appears in the Ordovician sedimentary deposits in this region. Comparison of the obtained age sets of detrital zircon grains from sample no. 1957 with similar spectra for rocks of both lower and higher stratigraphic levels in the southern part of the Siberian Platform makes it possible to demonstrate a similarity with those characteristic of the middle–upper Cambrian deposits, rather than the Early Ordovician [5].
Due to the fact that the found arthropods had an Ordovician–Devonian appearance, we considered it necessary to confirm the previously obtained Cambrian dates with additional samples taken from different stratigraphic levels of the Kimiltei section and combined them into one sample no. K-21 (Fig. 1b). The combination made it possible, by expanding the range of drift sources, to increase the probability of getting younger, in this case, Ordovician, detrital zircon grains into the sample. In addition, a sample from the Nory section located at a distance of about 20 km from Kimiltei [2] was analyzed. This close section also belongs to the Verkholensk Group (Fig. 1a).
Absolute Dating of Zircons from the Kimiltei Section, Sample K-21
The U‒Pb isotope analysis of zircons manually selected from crushed silty sandstone samples was carried out by the laser ablation with an Element XR mass spectrometer (Thermo Fisher Scientific) at the Geospectrum Center of the Geological Institute SB RAS (Ulan-Ude). The measurement parameters were standard; the whole procedure, the equipment used, and the statistical methods were described in detail earlier [7].
Each zircon grain corresponded to one point on the graph; its value was calculated from two measurements controlled by accepted procedural standards (91 500, Plešovice, and GJ-1). The difference between the two measurements obtained should not have exceeded 10%, and each peak on the graph was determined by at least three points with similar age values. The arrays of values were processed by means of the GLITTER, ISOPLOT, and AgePick and K-S Test software.
Analysis of the Kimiltei Age Ranges
Ninety-one grains were analyzed, out of which thirty were discordant. No Ordovician grains were found, and the youngest zircon from sample K-21 was dated at 492 ± 4 Ma. The maximum peak (ten grains) corresponded to 517 Ma, typical for the Cambrian rocks of the Verkholensk Group in the southern part of the Siberian Platform.
Samples 1957 and K-21 demonstrate a high similarity of age ranges (Figs. 3a, 3b). It can be related to the fact that the entire Kimiltei sequence was accumulated relatively quickly, so that the provenance sources did not noticeably change during this time.
The zircon drift topography began to change in the Early Ordovician. It can be seen from the analysis of a sample from the Nory section. The maximum peak in the Nory sample is of Early Ordovician age (485 Ma) (Fig. 3c). The later local deposits formed from the Middle Ordovician to the Carboniferous demonstrate a different PDC. They are characterized by peaks at 450 Ma and 1830 Ma being equal due to the activation of magmatism and erosion of the earlier buried Proterozoic rocks [5]. The Kimiltei samples are distinguished neither by the Early Ordovician peak identified in the Nory section, nor the above-mentioned pronounced bimodal form of the PDC, characteristic of the Ordovician–Carboniferous deposits in the southern part of the Siberian Platform.
The maximum peaks of samples 1957 and K-21 from the Kimiltei section are 511 and 517 Ma, respectively. The minimum age of zircons from the samples was 491 ± 6 and 492 ± 4 Ma. Hence, the re-dating of the total sample K-21 from the Kimiltei section confirmed the Late Cambrian age of the deposits. The Ordovician age of fossils in this section is excluded with a high probability. Based on these data, we make the assumption that the Kimiltei rocks can be dated at 488‒494 Ma. It should be emphasized that a reliable age determination is very important for this study, because the fossils found are, apparently, the most ancient ancestors of land arthropods represented, in fact, by a whole fauna assemblage.
BRIEF DESCRIPTION OF ARTHROPODS FROM THE KIMILTEI SECTION
Material
The fossil arthropods are located on nine massive slabs of compact gray silty sandstone. These slabs were broken out where possible, from the section wall, from one horizontal layer. The fossils occurred in the lower (reverse) side of the layer. Among the scree lying at the section foot, there were also fragments with fossils with a similar appearance.
Fossils are three-dimensional casts on a thin mudstone layer underlying a massive silty sandstone layer (Figs. 4a, 4b). All casts have a gray tint of silty sandstone, but some specimens are covered by a very thin reddish coating of rust. No bodily remains are observed inside the consolidated silty sandstone, on fracture planes and chips. In addition, no limb casts are seen anywhere, although one specimen retained casts of two front pairs of limbs (this specimen is not discussed in this paper).
In total, forty-three casts of various preservation were found on the slabs. Out of them, thirty-six seem to belong to the same species (this dominant species will be described in detail in our next paper). Two casts were found together with them. They were drastically different in morphology from the dominant species and from each other. Five other specimens were poorly preserved and unidentifiable. Most casts showed the animals’ ventral side. Hence, our collection has a dominant species and one representative each of the other two taxa.
The collection is kept under no. PIN 5891 at the Borissiak Paleontological Institute, Russian Academy of Sciences.
Morphology and Identifications
Representatives of the Kimiltei fauna assemblage are shown in Figs. 5 and 6. According to our determinations, they belong to Synziphosurina (Fig. 5a), Euthycarcinoidea (Figs. 5b, 5c), and Chasmataspidida (Fig. 6). Only one specimen was found for each of the first two groups, while chazmataspidids were buried en masse in Kimiltei. Therefore, based on the available data on two single specimens, only their high taxonomic category can be understood. The latter, chasmataspidids, apparently should be described by a new genus and species, and we will devote a separate publication to a detailed description. In this paper, we provide brief morphological characteristics with emphasis on those features by which the found forms can be attributed to the corresponding higher taxa.

Kimiltei fossils: (a) Sinziphosurina, specimen PIN 5891 no. 8, (b) Euthycarcinoidea, specimen PIN 5891 no. 48, (c) enlarged fragment of the head with an antenna. (a', b', and c') Corresponding drawings arranged from photo images with different lighting angles: (act) Anterior head tergite; (appII–VI) grooves corresponding to limbs of segments III–VI; (ant) antenna; (bck) dorsal shield; its segmentation is shown on the right, (chi?) probable chilaria, (dst) paired sternites; (epst) epistome; (gapp) sex processes; (go) sex cover; (gut) gut; (lb) lip (labrum), (mo) mouth, (mnd) mandibles, (pro) prosoma, (psg) posterior (without longitudinal grooves) segments, (spp/e) spherical process (eyes), (st) sternites, (T) telson, caudal spine, (7-16?) opisthosoma segments.

Chasmataspidida. (a) Specimen PIN 5891 no. 19; ventral plates are present. (b) Chasmataspidida, specimen PIN 5891 no. 2; ventral plates are not preserved. (a', b') corresponding drawings: (appII–VI, appVI) Grooves corresponding to limbs of segments II–VI; (bck) dorsal shield; its segmentation is seen in some places; (brapp) gill limbs, (chel) chelicerae, (epst) epistome, (go) sex cover (?), (mo) mouth; (pro) prosoma; (preT) pretelson, (st) sternites; (T) telson; (vp9-11) ventral plates of segments 9–11; (vr) ventral keels; (7–19) opisthosoma segments.
Synziphosurina (Fig. 5a). This specimen, open from the ventral side, is characterized by the following morphological features. The body is subdivided into a head section and a trunk which ends with a long tail spine. The head section, a prosoma, is wide at the base, narrowing to a triangle towards the anterior margin. It is impossible to determine whether the prosoma has a marginal limbus or a doublure in this specimen. At its anterior end, a triangular protrusion, an epistome, is distinguished by grooves, at the end of which there is an oval labrum. Under it there is a hole; it appears to be a mouth. A longitudinal medial groove extends down from the labrum. To the right and left of it, there are curved V-shaped grooves or relief structures; there are five of them, and we interpret them as remains of coxal prosomal limbs II–VI. At the end of this groove, an interesting element is clearly visible in the form of two close (or connected) small rounded parts. Behind them there is a short horizontal strip. Further, a large eighth segment consists of two symmetrical parts. Their inner edges diverge semicircularly to the side corners. The medial structure is placed in the resulting triangle. Its surface is uneven, with embossed horizontal bulges. It can be assumed that these are articulation remains of this structure. It is relatively long, extending to the posterior margin of the next, ninth, segment. The tenth segment is also divided along the center of symmetry by a medial groove; two small forked elements are distinguishable on the inner edge of the halves.
Asymmetric extensions on the sides of the tenth and eleventh segments look like lateral spikes, but they are, in fact, an unfortunate cleavage of clayey rock which is very fragile in this area. Therefore, it is not possible to describe the shape of the posterior margin of the animal’s shield.
Five or six segments following one after another decrease in width, forming a triangular posterior end of the body. The first three of these posterior segments are divided into symmetrical halves by a medial groove, while the last two or three segments are solid. A part of the segment in front of the long tail spine is destroyed in this specimen, and it is impossible to judge whether it had another narrow terminal sixth segment or whether the body ended in a relatively wide fifth segment.
Summarizing the above-mentioned morphological description, it is important to pay attention to tagmas of this animal. The head section included at least five pairs of distinct grooves which, in their shape, can be interpreted as limb casts. It should be noted that the first pair of limbs, cheliceras, are extremely rarely preserved in fossil chelicerae, so most likely they were not preserved in this case either. Presumably, this animal could have six pairs of head limbs. The anterior part also includes two segments, the seventh and eighth ones. The seventh segment is observed as a short transverse strip without a medial groove; it can be correlated with the seventh microsegment of the cephalothorax characteristic of some basal chelicerates. If so, then the paired symmetrical element on this microsegment can be correlated with the chilaria of Xiphosura. It is logical, therefore, to assume that the next eighth segment bears large symmetrical genital oprecula. The opisthosoma of this animal consists of seven or eight segments, out of which the last two or three segments are fused together and with the tail spine.
This tagmatization corresponds to Xiphosura and Synziphosurina. The cephalothorax of horseshoe crabs consists of eight segments, and the next part, a toracetron, combines five gill segments; two fused posterior segments end in a tail spine. When comparing the Xiphosura tagmata with the Kimiltei specimen, we still should pay attention to the medial, likely articulated structure between the branchial opercula. Neither present-day nor fossil Xiphosura has such structure. It is also uncommon that this taxonomic version is devoid of any traces of a dublura.
In Synziphosurina, the head tagma includes six segments and the seventh segment can bear a pair of walking legs or chilaria (the latter are known from Dibasterium). A doublure is also not necessary for representatives of this group [8]. The middle (mesosomal) part of the synziphosurin’s body combines six segments with gill limbs, and the next metasomal part includes four segments without limbs. The telson can be expressed by a tail spine. The “Synziphosurina” hypothesis requires an explanation for the smaller (three rather than four) number of metasomal segments. However, the boundaries between the segments were likely poorly preserved or not expressed in this specimen. In early chelicerates and their ancestors, the calculating of metasomal segments is generally difficult due to their indistinct of fused boundaries. This problem was discussed in the redescription of Houia, a possible relative of chasmataspidids and eurypterids.
Eurypterids are also similar to xiphosurans and synziphozurins. They, however, have five metasomal (postabdominal) segments; i.e., their number is even greater than that of the Kimiltei specimen. Therefore, despite a medial structure between the halves of the eighth segment—and this, as mentioned above, is an analogue of genital appendages—it is hardly necessary to associate the Kimiltei specimen with eurypterids.
Based on the combination of features (characteristic tagmata, absence of a doublura and limbus), the Kimiltei form is the most similar to synziphozurins. It is interesting to note probable chilaria and genital appendages of the eighth segment. Thus far no forms are known in which chilaria and genital appendages would be present simultaneously. However, in order to discuss this phenomenon seriously, additional specimens of this species are needed to confirm it. In addition, due to this reason, we will be careful to speculate on the phylogenetic relationships of the Kimiltei form with other basal species of Eucherli-cerata.
Euthycarcinoidea (Figs. 5b, 5c). The second form also represented by a single specimen is attributed to euthycarcinoids. It should be emphasized that this specimen (its ventral side) is very poorly preserved, so that only a limited set of morphological features can be reliably distinguished on it. Nevertheless, they make it possible to maintain the “Euthycarcinoidea” taxonomic version.
This arthropod had a small head (Fig. 5c) and a long, segmented body. The remains of a narrow curved strip of small segments are traced to the right of the head; its morphology and location correspond to the antenna. The posterior end of the body is destroyed, so it is impossible to judge the presence of a telson. In the anterior part of the head, there are remains of the protruding tergite; it apparently covered the anterior end of the head from the dorsal side. Two symmetrical club-shaped processes set on sides of the anterior tergite protruding proximally. They are similar in their form and position to the so-called spherical processes of euthycarcinoids. These processes are interpreted as eyes. Behind them there is a clear two-lobed symmetrical structure like butterfly wings. They are separated by a medial groove. Anterior parts have internal semicircular pits indicating the complex structure of this element. It is similar to the mandibles of known euthycarcinoids, for example, Apankura [9], Heterocrania [10], and Euthycarcinus [11, 12]. A small depression is seen in the medial groove between these elements; it is probably a mouth. The posterior lobes of this structure are also symmetrical and separated by a groove. The nature of these element is not clear. In some species, it is interpreted as a buccal pharyngeal complex involved in food processing or as a pair of cephalon limbs following the mandibles [11].
Behind the two-lobed structure, there is a triangular element directed at an obtuse angle to the medial groove. In this specimen, it is impossible to determine whether this structure is a trunk segment or a part of the head tagma, because the head is not clearly separated from the body on the ventral side. It should be noted for comparison that S. Shlyutka [12, Fig. 6] showed a similar element in Euthycarcinus ibbenburensis, considering it as an analogue of the labrum and including it in the head tagma. Meanwhile, in other euthycarcinoids, for example, Sottyxerxes multiplex and Apankura machu, two ventral plates are located behind the mandibles, but there is no labrum. Their head section is thus composed of only two segments [9, 13].
The trunk of euthycarcinoids is subdivided into segments. It is impossible to determine the number of segments in this specimen, because the posterior end of the body is smoothed and the boundaries of the segments are not visible. In the anterior part, the segments are narrower (sagittally) than in the posterior one. A closer examination of the segmentation makes it possible to reveal that the segments are relatively wide in the anterior part of the body: the boundaries of the wide segments are clear on the sides, but each segment is divided in half by a horizontal groove in its central part. The gut running along the body also has boundaries of wide segments in the anterior part, such as those on the sides of the body.
In this specimen, six anterior body segments are divided in half horizontally. This segmentation pattern can be interpreted as casts of large tergites protruding laterally and superimposed on narrow (in a saggittal and horizontal planes) sternites; two sternites correspond to each tergite. This is a segmentation of euthycarcinoids.
Hence, this animal has identifiable segmented antenna, large eyes on processes, mandibles on a relatively small head covered by at least one large tergite, and the segmented body with large dorsal tergites and ventral sternites narrower anteriorly. According to these features, this animal can be attributed, relatively confidently, to euthycarcinoids.
Chasmataspidida (Figs. 6a, 6b). In the specimens studied, the body is subdivided into an anterior part, closed by a shield, and a posterior, clearly segmented part. The shield covers the prosoma and preabdomen; they are separated on the dorsal side by a narrow microtergite; on the ventral side, the microtergite is not distinct. Prosoma has a triangular epistome extending from the apical margin [14, 15] with a small oval labrum at its end. The prosoma bears short chelicerae bent downwards and small pedipalps with a spine on the terminal segment; both the chelicerae and pedipalps are directed toward the mouth. The four following prosomal limbs, judging from the relief grooves on the prosoma, are relatively large; the sixth pair is the largest. The coxae of these four limbs form a medial groove along the centre line.
The preabdomen is covered by a shield which, on the ventral side, is turned down from both sides in wide folds forming cavities for the preabdominal limbs. It, as far as can be judged from the ventral casts, consists of five segments, the first of which corresponds to the narrow seventh segment and the microtergite on the dorsal side. The eighth segment is large with a cover divided into symmetrical halves. Long genital appendages are located along the inner edge of these halves. The ninth–eleventh segments are covered by plates not divided along the symmetry axis. They are most likely merged, because no movable joints are noted anywhere between them. Movable articulations of the segments are present only starting from the boundary between the eleventh and twelfth segments. In the specimen shown here, the plates are absent on the ninth–eleventh preabdominal segments, but instead fossilized limbs of the preabdomen are seen clearly. They are wrinkled, with folds, i.e., not rigid, so it is logical to assume that these are soft branchial limbs. It is possible that the limbs of the eighth segment were also completed with gills.
There are eight postabdominal segments (12‒19). All of them have movable joints. A symmetrical ventral keels run along sternites 14‒19; these segments have one longitudinal keel on the dorsal side. The short pretelson bears the anus, and the telson itself, apparently, is short, lanceolate. It was not preserved in full in the specimens we have.
A characteristic feature of the order Chasmataspidida is the division of a body into a prosoma of six segments and an opisthosoma of thirteen segments, the latter consisting of a four-segmented preabdomen and a nine-segmented postabdomen. However, a form described from the late Cambrian of Texas is also referred to as Chasmataspididae (the species has not yet been described formally). It appears to have had seven or eight postabdominal segments and five or six pre-abdominal segments [16]. The Kimiltei species has the same tagmata. In the Texas form, the dorsal shield covered segments of the preabdomen in the same manner as in the Kimiltei species. In addition, a distinctive feature of chasmataspidids is a merged ventral plate on segments 8‒10 of the body; it is present in many specimens of the Irkutsk species, but reaches the middle of the eleventh segment. On the basis of this feature and tagmatization of the chasmataspidids, we classify the Kimiltei form as a chazmataspidid.
DISCUSSION
The most ancient representative of euthycarcinoids, Apankura machu, was found in the Garganta del Diablo locality. The age of the member with this find in the Santa Rosita Formation, according to trilobites Neoparabolina frequens argentina and Plicatolina scalpta found in this area, is considered to be the uppermost of the late Cambrian; this is the South American Neoparabolina frequens argentina zone [13]. Facies features of the layer with the fossils correspond to open-marine, likely subtidal, deposits. Another representative of this group, Mosineia macnaughtoni, was found in Wisconsin, United States, in the Elk Mound Group deposits, where the age range of rocks with fossils is wide: from the Terranuvian to the lower half of the Furongian. Euthycarcinoids Mictomerus melochevillensi are known from Canada, the Potsdam Group deposits; their age is also within the range from the middle Cambrian to the lower half of the Furongian [6]. The age of the Kimiltei findings is comparable with the upper age limits of all these fossils: i.e., the Furongian, its lower half. In addition, according to the U‒Pb isotope dating of detrital zircon grains (485‒495 Ma), the Kimiltei form belongs to the Furongian division. It should be noted that the fossils from Argentina can be both contemporaries of the North American and Irkutsk euthycarcinoids, and somewhat younger than them. Hence, euthycarcinoids appeared to be present in the middle of the late Cambrian in at least three areas: tropical Laurentia in the southern hemisphere, equatorial waters of the Siberian Platform, and temperate or subpolar Gondwana in the southern hemisphere. If this group is monophyletic (and we still do not have any facts for a different opinion), then it should have formed and passed through the initial stages of their evolution before the beginning of the late Cambrian. However, focusing on the wide geography of the late Cambrian findings, it is hard to guess where their ancient homeland was located.
Kimiltei synziphosurins are the most ancient of all known representatives of this group. Synziphosurins are currently considered as a combined group: one part of them is similar to the Xiphosura, and the other is attributed to the basal Euchelicerata. The earliest representative of the “horseshoe crab” line is Lunataspis from the Late Ordovician of Canada; and among the basal Eucherlicerata with synziphosurine features, a form from the Early Ordovician of Morocco (Fezoata biota) is known [17, 18]. The Kimiltei form, whatever line it belongs to, is dated at no younger than the late Cambrian, so it is older than both the Canadian and Moroccan species.
The most ancient chasmataspidids [16] are described based on casts from Texas, United States, from Dresbachian deposits globally comparable with the lower half of the Furongian. The Kimiltei species apparently co-existed with the Texas one. However, it is still hard to compare their morphology due to the very different preservation. Therefore, it is impossible to conclude how close these forms were. Anyhow in the late Cambrian, chasmataspidids, like euthycarcinoids, had already passed the initial stage of dispersal and radiation. Alongside with that, some of the most ancient representatives of all three groups such as chasmataspidids, euthycarcinoids, and synziphosurins co-habited in the Kimiltei paleoaquatic area.
In Kimiltei, the Texas Hickory Sandstone locality, and the Wisconsin Blackberry Hill, the fossil facies represented shallow dry zones. Therefore, all animals from the discussed groups lived, most likely, in the littoral zone. Euticarcinoids began to inhabit land already in the late Cambrian. This assumption is based on the fact that on the bedding plane with fossils at Blackberry Hill there are numerous trackways left, as it is assumed, by euthycarcinoids [19]. Subsequently, the evolution of this group led to the formation of terrestrial insects. Synziphosurina, as noted above, does not form a single clade, it is a combined group at the base of the Euchelicerata tree, and its part is related to Xiphosura. The present-day Xiphosura, horseshoe crabs, live in coastal sea waters, but come to land for breeding. The present-day Euchelicerata (arachnids) are mostly terrestrial, and fossil eurypterids, also classified as Euchelicerata, could come ashore above the tidal zone. In other words, the Synziphosurina descendants are tended to terrestrial life. The Kimiltei shallow-water synziphosurines appeared to have had genital appendages, which are thought to be pre-adaptation for land mating. This information is consistent with the further evolution of their descendants who began to inhabit the land. During their evolution in the Ordovician–Devonian, chazmataspidids inhabited brackish and fresh waters [20]; this fact is hard to imagine without the ability to safely survive draining the drying.
Hence, the middle–late Cambrian Kimiltei fauna includes representatives of those groups which, in the course of their evolution, began to inhabit the coastal zone and land. Apparently, in the late Cambrian, they were already preadapted to this ecological task. They were solving it in parallel, each using its own morphological peculiarities. However, they put off on this long evolutionary journey at the same time and from the same ecological start.
Change history
21 April 2024
An Erratum to this paper has been published: https://doi.org/10.1134/S1028334X23070322
REFERENCES
V. S. Voropinov, Dokl. Akad. Nauk SSSR 114 (6), 1291–1293 (1957).
S. Yu. Garina and L. V. Ogienko, Stratigraphy and Trilobites of the Cambrian of the Siberian Platform (Nauchn. mir, Moscow, 2001) [in Russian].
Resolutions of the 4th Stratigraphic Conference on the Refinement of Stratigraphic Schemes for the Vendian and Cambrian of the Interior of the Siberian Craton (Siberian Res. Inst. Geol., Geophys., Mineral Raw Materials, Novosibirsk, 1989) [in Russian].
N. V. Mel’nikov, Yu. Ya. Shabanov, and O. S. Shabanova, Russ. Geol. Geophys. 51 (6), 672–683 (2010).
D. P. Gladkochub, Z. L. Motova, T. V. Donskaya, V. B. Khubanov, and A. V. Sizov, J. Asian Earth Sci. X 8, 100107 (2022). https://doi.org/10.1016/j.jaesx.2022.100107
J. H. Collette and J. W. Hagadorn, J. Paleont. 84 (4), 646–667 (2010).
V. B. Khubanov, M. D. Buyantuev, and A. A. Tsygankov, Russ. Geol. Geophys. 57 (1), 190–205 (2016).
J. C. Lamsdell, Zool. J. Linn. Soc. 167 (1), 1–27 (2013).
N. Vaccari, G. Edgecombe, and C. Escudero, Nature 430, 554–557 (2004). https://doi.org/10.1111/j.1096-3642.2012.00874.x
L. I. Anderson and N. H. Trewin, Palaeontology 46, 467–509 (2003).
J.-C. Gall and L. Grauvogel, Ann. Paleontol. Invert. 50, 1–18 (1964).
S. Schultka, Palaontol. Z. 65 (3-4), 319–332 (1991). https://doi.org/10.1007/bf02989847
P. R. Racheboeuf, J. Vannier, F. R. Schram, D. Chabard, and D. Sotty, Earth Environ. Sci. Trans. R. Soc. Edinburgh 99 (1), 11–25 (2008).
J. C. Lamsdell, The Eurypterid Stoermeropterus conicus from the Lower Silurian of the Pentland Hills, Scotland (The Palaeontogr. Soc., London, 2011).
J. C. Lamsdell, G. O. Gunderson, and R. C. Meyer, BMC Evol. Biol. 19 (8), 1–24 (2019).
J. A. Dunlop, L. I. Anderson, and S. J. Braddy, Trans. R. Soc. Edinburgh Earth Sci. 94, 207–225 (2004).
P. Van Roy, D. E. Briggs, and R. R. Gaines, J. Geol. Soc. 172 (5), 541–549 (2015).
L. Lustri, P. Gueriau, P. Van Roy, and A. C. Daley, in Proc. 18th Swiss Geoschi. Meet. (Zurich, 2020).
S. J. Braddy, K. C. Gass, and T. C. Gass, Geology Today 38, 25–31 (2022).
J. C. Lamsdell and D. G. Briggs, Geol. Mag. 154 (1), 175–180 (2017).
ACKNOWLEDGMENTS
We thank N.M. Boeva, Institute of Geology of Ore Deposits, Petrography, Mineralogy, and Geochemistry, Russian Academy of Sciences, who carried out a range of mineralogical analyses for this work with a high professional responsibility.
Funding
This work was carried out under a State Assignment of the Borissiak Paleontological Institute, Russian Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by E. Maslennikova
Publisher’s Note.
Pleiades Publishing remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: Due to a retrospective Open Access order.
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Naimark, E.B., Sizov, A.V. & Khubanov, V.B. Kimiltei Is a New Late Cambrian Lagerstätte with the Faunistic Complex of Arthropods (Euthycarcinoidae, Synziphosurina, and Chasmataspidida) in the Irkutsk Region. Dokl. Earth Sc. 512, 859–870 (2023). https://doi.org/10.1134/S1028334X2360127X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1028334X2360127X