
Abstract
The carbonaceous macrofossils which belong to multicellular algae and which were previously known from the younger (>570 Ma) Miaohe biota are found for the first time in the Lower Vendian (Middle Ediacaran, ~580 Ma) rocks of the Ura Formation of the Patom Basin in Siberia. The Ura macrofossils include phytoleims of unbranched ribbon-like thalli preserved on the surfaces of silty mudstones. Among the fossils, the species Liulingjitaenia alloplecta and Jiuqunaoella simplicis are identified. The features of their structure are considered. The in vivo forms of the organisms and their colonies are interpreted. It is shown that these organic macrofossils were a noticeable detritus-forming component of the Ura Basin. The Ura macrophytes coexisted with the Early Ediacaran microbiota and composed a common ecosystem of the paleobasin, which formed shortly before the Shuram event and was significantly transformed after it.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The largest negative Shuram δ13C anomaly (EN3) in the Earth’s history, which is confined approximately to the middle of the Ediacaran, is known from sedimentary successions of many paleocontinents of that period (Grotzinger et al., 2011). In spite of contradictory estimates of its precise age and duration (Gong et al., 2017; Witkosky and Wernicke, 2018; Xiao and Narbonne, 2020), it is most likely that this event occurred in the interval of 571–562 Ma and lasted 8–9 m.y. (Canfield et al., 2020; Gong et al., 2017; Minguez et al., 2015). In the Ediacaran paleontological record, the Shuram anomaly (namely, its base) is a main paleoecological boundary that divides the end of the predominance of the Early Ediacaran unicellular eukaryotes and the first occurrence of the first Late Ediacaran multicellular animals. It is suggested that the observed evolutionary innovations and ecosystem turnover at this boundary were related to the oxygenation of the ocean of that period (Canfield et al., 2007; Fan et al., 2020; Narbonne, 2010; Sahoo et al., 2016; Zhang et al., 2019).
It is evident that the Shuram event was also a crucial moment in evolution of other groups of organisms, the remains of which are typically preserved on rock surfaces as macroscopic carbonaceous compressions (phytoleims; Burgess Shale-type preservation) and impressions, the majority of which is associated with macroalgae. The remains of these organisms are known as a Miaohe type of fossil macrobiota, which is most completely preserved in the post-Shuram Ediacaran rocks of South China (Xiao et al., 2002; Ye et al., 2017). The taxonomically poor assemblages of this macrobiota are also known beyond China: in Western Mongolia (Dornbos et al., 2016), the United States (Rowland and Rodriguez, 2014), and Australia (Xiao et al., 2020), as well as Siberia (Grazhdankin et al., 2008) and the Urals (Grazhdankin et al., 2007; Marusin et al., 2011). As with the Ediacaran “soft-bodied organisms,” the typical representatives of the Miaohe biota are confined to the post-Shuram rocks that lack assemblages of the Early Ediacaran acanthomorphs. In this paper, we report on a new locality of some taxa of the Miaohe biota in the pre-Shuram strata of the Ura Formation in Siberia, where these fossils are closely associated with widely known Ura acanthomorphic biota of the Doushantuo–Pertatataka type (Moczydłowska and Nagovitsin, 2012; Sergeev et al., 2011; Vorob’eva and Sergeev, 2018).
PATOM BASIN AND THE AGE OF ITS ROCKS
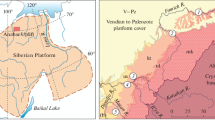
The Patom Basin, which encircles the southern margin of the Siberian Craton (Fig. 1), formed after the breakup of Rodinia Supercontinent in the end of the Neoproterozoic (Metelkin et al., 2012; Powerman et al., 2015; Sovetov, 2002). The lower part of terrigenous-carbonate succession of this basin has been distinguished as a thick (>6 km) Patom Supergroup, which includes (bottom to top) the Ballaganakh, Dal’nyaya Taiga, and Zhuya groups (Chumakov et al., 2007). The Bolshoi Patom glacial horizon, which is a thick (~1 km) succession of diamictites traditionally compared with Marinoan glaciation (635 Ma), occurs at the base of the Dal’nyaya Taiga Group without features of a long stratigraphic hiatus (Chumakov, 2015; Chumakov et al., 2013). The age of the Bolshoi Patom diamictites (as well as the entire Dal’nyaya Taiga Group), however, remains a matter of debate (Rud’ko et al., 2020). The data of basin analysis (Petrov, 2018a, 2018b) along with paleontological data (Leonov and Rud’ko, 2012; Vorob’eva and Petrov, 2020) and, partly, with δ13C data (Petrov and Pokrovsky, 2020; Rud’ko et al., 2017) suggest a younger (<580 Ma) age of the Dal’nyaya Taiga rocks, as well as the Vendian (Ediacaran) age of the entire Patom Supergroup. Recent Pb–Pb isochron ages of high-Sr limestones of the Dal’nyaya Taiga Group of 581 ± 16 and 575 ± 20 Ma (Barakun and Kalancha formations, respectively) are consistent with this suggestion (Rud’ko et al., 2021). In this interpretation, the thick sequences of the Ballaganakh and Dal’nyaya Taiga groups compose a single terrigenous–carbonate transgressive-regressive cycle, the sequence of which was interrupted during the most intense basin subsidence by a eustatic fall of sea level related to the Bolshoi Patom glacial episode. During the glacial maximum, only proximal areas of the Patom Basin were eroded in contrast to continuous sedimentation in the internal deep-sea area.

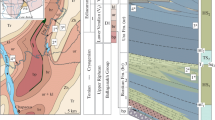
Region of study (a) and location of analyzed fossil organisms in Vendian (Ediacaran) rocks of the Ura Uplift (b). The isotopic-geochemical data are after (Pelechaty, 1998; Petrov and Pokrovsky, 2020; Pokrovsky and Buyakaite, 2015; Rud’ko et al., 2015), stratigraphy is after (Chumankov et al., 2013; Petrov, 2018a, 2018b), and paleontological data are after (Khomentovsky et al., 2004; Kochnev and Karlova, 2010; Leonov and Rud’ko, 2012; Moczydłowska and Nagovitsin, 2012; Sergeev et al., 2011; Vorob’eva and Petrov, 2020). Groups: Bl, Ballaganakh; Dt, Dal’nyaya Taiga; Zu, Zhuya; Tr, Trekhversta; formations: bp, Bol’shoi Patom; br, Barakun; ur, Ura; kl, Kalancha; vl, Valyukhta; nk, Nikolskaya; cn, Chencha; zr, Zherba; tn, Tinnaya; nh, Nokhtui; Є, Cambrian; О, Ordovician. EN3, Shuram δ13C negative anomaly. Anabarites trisulcatus and Nochoroicyathus sunnaginicus (Aldanocyathus sunnaginicus): zones of the Nemakit–Daldynian Stage of the Vendian and Lower Cambrian (after Russian and International Stratigraphic Charts, respectively).
Postglacial deposits of the Dal’nyaya Taiga Group on the Ura Uplift include a succession of the Barakun, Ura, and Kalancha formations (Chumakov et al., 2013). This succession >2 km thick contains two terrigenous-carbonate cycles of the second order: Lower Barakun and Upper Ura–Kalancha (Petrov, 2018b). Toward the center of the basin, the latter is replaced by relatively homogeneous and more clayey deposits of the Valyukhta Formation (Fig. 1b). The sandstones of the lower part of the Barakun Formation contain impressions of the Vendian fossils Beltanelloides sorichevae Sokolov (Leonov and Rud’ko, 2012), which are ascribed to Beltanelliformis brunsae Menner in the current classification (Ivantsov et al., 2014). The acanthomorphic biota of the Ura and Barakun formations contains taxa typical of the upper two assemblage zones of Australia and China (Vorob’eva and Petrov, 2020).
The Dal’nyaya Taiga Group is unconformably overlain by a siliciclastic-carbonate succession of the Zhuya Group with a negative δ13C excursion significant in amplitude (down to –11.9‰) and thickness (>800 m) (EN3, Fig. 1b), which corresponds to the Shuram global anomaly (Melezhik et al., 2005, 2009; Pokrovsky and Buyakaite, 2015; Pokrovsky et al., 2006a, 2006b) that, according to the latest data, began just after 571 Ma (Canfield et al., 2020) or 574 ± 4.7 Ma (Rooney et al., 2020). This anomaly is also traced in the internal regions of the Siberian Platform in the upper part of the Nepa Horizon (Kochnev et al., 2018). The unconformity at the base of the Zhuya Group is a main sequence boundary of the Patom Basin (Petrov, 2018b) and is an erosion surface without features of angular unconformity, however, with striking erosion incision with amplitude of up to 100 m. The erosional topography in the early Zhuya time was smoothed by sandy sediments, which now include a local basal member of the Nikolskaya Formation (Chumakov et al., 2013). This member 80 m thick was previously assigned to the Kullekin Formation (Bobrov, 1964, 1979; Kolosov, 1975). It is evident that erosion and further sedimentation was a result of a short-term fall and rise of the sea level significant in amplitude. These fluctuations could have been initiated by tectonic reconstruction of the basin (third-order cyclicity 0.5–3.0 m.y. in length; Cloetingh et al., 1985) or were of glacioeustatic nature similar to the last glacial event of the Late Pleistocene in amplitude and duration (Lambeck et al., 2014). At the top, the Zhuya Group is truncated by a similar erosion surface and is overlapped by terrigenous-carbonate deposits of the Trekhversta Group including the Zherba and Tinnaya formations (Fig. 1b) (Chumakov et al., 2013). The clayey-carbonate rocks of the upper part of the Tinnaya Formation contain small-shell fauna of the Anabarites trisulcatus Zone of the Upper Vendian Nemakit–Daldynian Stage (Khomentovsky et al., 2004), and the overlying Nokhtui Formation hosts numerous fauna of the Nochoroicyathus sunnaginicus Zone of the Lower Cambrian Tommotian Stage (Khomentovsky et al., 2004; Kochnev and Karlova, 2010). Thus, no stratigraphically significant hiatuses are observed in the sedimentary succession of the entire Patom Supergroup, and the age of the Dal’nyaya Taiga Group is constrained to a period of 580–570 Ma.
MATERIALS AND METHODS
The main paleontological material was collected from the upper part of the Ura Formation that is exposed at the right bank of the Ura River, 5.5 km upstream from the mouth (GPS: 60.26753° N, 117.10835° E) (Fig. 1b). A few samples were taken from the middle part of the section of the formation, which is located in the upper stream of the Ura River, upstream from the mouth of the Ulakhan-Iligir River (GPS: 60.40280° N, 117.35112° E). The Ura silty mudstones are characterized by ubiquitous concentric and disordered linear cleavage oriented at various angles to sedimentary surfaces, which created serious difficulties in preparation of macrofossils. For example, macrofossils are lost in many cases as a result of self-induced (without mechanical impact) decomposition of samples during its repeated wetting. To avoid the decomposition of samples (especially, during preparation of thin sections), the samples were saturated with hot epoxy resin.
The samples were prepared and macrofossils were studied under a MBS-9 binocular microscope. The macroimages of fossils were taken during the surface treatment by glycerin. The microstructures were studied and photomicrographs were taken in transmitted and reflected light, as well as under UV fluorescence using a Zeiss REM 5 microscope and a Zeiss Axio Imager A1 microscope equipped with an Axio-CamMRc-5 digital camera (Carl Zeiss, Germany). The fluorescence method is used during the study of organic (highly carbonaceous) microfossils in thin sections prepared using Canada balsam. Excited by UV radiation, Canada balsam along with the quartz–alumosilicate matrix of silty mudstones is evenly luminescent, which reveals nonfluorescent organic remnants and their fine microstructure invisible in transmitted light. The paleontological material is stored at the Geological Institute, Russian Academy of Sciences (GIN RAS, Moscow), collection no. 14801.
FACIES CHARACTERISTICS OF THE DEPOSITS AND ORGANIC MACROFOSSILS OF THE URA FORMATION
Sedimentary Environments of the Ura Basin
Mostly silty-clay deposits of the Ura Formation accumulated in deep-sea (below storm wave base) conditions on a low-angle (~1°) homoclinal ramp with ubiquitous surficial (no more than a few tens of centimeters thick) landslides and mud flows initiated by seismic impact (Petrov, 2018b). The amount of the deformed sedimentary beds in the section often reaches 90%. It is suggested that the mass-flow sedimentation atypical of similar very low-angle ramps was related to the intense processes of methanogenesis in the bottom layers. This conclusion is supported by C isotopic data for the early diagenetic carbonates (Petrov and Pokrovsky, 2020). Most fine-grained siliciclastic particles of the Ura Formation, evidently, have an aeolian origin, which is in agreement with the grain-size distribution, as well as the structural and mineralogical features of the Ura sediments (Petrov, 2018b). A significant amount of the aeolian material, in turn, suggests increasing aridity of that period. A small amount of allothigenic carbonate in silty mudstones of the Ura Formation was related to wave erosion of the carbonate platform in the outer part of the basin, which propagated along the entire territory of the Ura Uplift during the Kalancha time. The progradation front of this platform contains the richest taphocenosis of the Ura acanthomorphs, although the poor assemblages of this microbiota are found at various levels of the section in distal basin facies (Vorob’eva and Petrov, 2020).
Organic Detritus in the Ura Sedimentary Rocks
In addition to microfossils, the Ura mudstones contain abundant organic macrofossils in the form of small-sized (1–10 mm) detritus and elongated (up to 10 cm), locally, structured fragments (films) (Figs. 2a, 2b). Similar fragments remained unbroken during transportation inside landslide and even in mass flow layers (Petrov, 2018b). Similar variously deformed but relatively flexible and plastic films are known from sedimentary rocks of various ages and they are typically interpreted as remnants of benthic microbial mats (Schieber, 1999, 2007; Simonson and Carney, 1999). In contrast to amorphous and internally unstructured fragments of the microbial mats, however, some Ura fragments are characterized by repeated forms with clear boundaries and various internal structure that is distinct from microbial. The small-sized detritus contains numerous oval or conical films, whereas the large-sized fragments contain elongated ribbons with smooth margins. The internal structures of some fragments, which are observed in thin sections, also noticeably differ from typical clotted or crinkly laminated microbial structures. These structures consist of straight clear parallel films, intercalating filaments, or micron-sized fibers (Figs. 2c–2e). Uniformly organized fibrous fabrics are observed in oblique sections of some least degraded ribbon fragments (Fig. 2f). The structures of strongly degraded fragments are composed of series of adjacent films, which are similar in transverse section to structures of microbial mats; however, in contrast, they are constant in thickness and have clearer outer boundaries. It is evident that the majority of buried detritus was subjected to significant anaerobic bacterial destruction with the formation of syngenetic framboidal pyrite, the amount of which is generally proportional to the degree of degradation of the observed biogenic structures (Figs. 2g, 2h).

Morphology and structure of large organic detritus in silty mudstones of the Ura Formation. (a, b) Sections of large film fragments on the surface of the mass flow beds; (c) transverse section of the mass flow bed (the lower part of the image) and mud flow (the upper part); (d, e) structures of detritus buried inside the mass flow bed; (f) discontinuous hatched fibrous structures of ribbon-like fragment of Liulingjitaenia alloplecta in oblique longitudinal section; (g, h) degraded fragments of detritus with syngenetic framboidal pyrite. Thin polished sections in transmitted light (c, d) and in fluorescent light from UV radiation (f–h).
Thus, a noticeable part of the organic detritus of the Ura Formation represents some organisms or colonies, which differ from mat-forming microbial communities in composition and organization. The remnants of these organisms were the detritus-forming component in the fine-grained Ura sediments.
Macrofossils of the Ura Formation
The macroscopic representatives of the Miaohe biota are found in the upper part of the formation, in beds with well-known acanthomorphic microbiota (Moczydłowska and Nagovitsin, 2012; Sergeev et al., 2011) (Plates I, II). The taphocenosis is dominated by helically twisted ribbons of Liulingjitaenia alloplecta Chen et Xiao. The ribbons straight narrow and nearly identical in width (typically, 2–3 mm) exhibit neither a beginning nor an end. In a typical locality (the Miaohe biota of China), their maximum length reaches 15–20 cm (Xiao et al., 2002). The maximum length of the ribbons in our material is ~8 cm. Numerous aggregations of unidirectional specimens are typically observed (Plate I, figs. 1–6). The morphological structure of the macrofossils analyzed is a matter of debate. According to some researchers, these forms represented a dense bundle of twisted filaments (Chen and Xiao, 1992; Xiao et al., 2020; Ye et al., 2017). A simple thick rope is an obvious example of the external morphology of these forms. In another interpretation, they had the form of an empty tube with numerous spiral folds due to twisting (Steiner, 1994; Xiao et al., 2002). This is evident from the analysis of the Ura phytoleims preserved simultaneously on the bottom and the top of the same bed (Plate I, fig. 1); however, these organisms mostly likely had the form of a helically twisted ribbon rather than a twisted bundle of filaments or twisted cylindrical tube. In tightly twisted specimens, the spiral coils could close up, thus resembling a twisted filamentous bundle, whereas the coil junctions could imitate an empty strongly twisted tube. At the same time, the same specimens often exhibit the variable direction of the ribbon curls. The various areas of the same ribbon were twisted in opposite directions (Figs. 3a, 3b). In addition, some parts of the spiral could be turned into a flat sinusoidal curved meandering ribbon (Plate I, fig. 2, the upper part of the image). Amid the specimens of the Ura Liulingjitaenia, one can observe various forms of these organisms depending on the character and the degree of twisting of the ribbons. In our material, however, the ribbons with unidirectional twisting (either to the left or to the right) are significantly dominant. The ribbons of Liulingjitaenia alloplecta have unclear fibrous fabric, which shows clear filaments of different thickness (10 to 50 µm) (Plate I, figs. 5, 7, 8). Some filaments are formed by chains of spherical aggregates of secondary framboidal pyrite (Plate I, fig. 9). All filaments are helically twisted, emphasizing a frame of the entire spiral construction.

Plate I . Liulingjitaenia alloplecta M. Chen et Xiao, 1992. (1, 2, 8) Specimen GIN no. 14801-20024-1: (2, 8) enlarged fragments; (3–5, 7, 9) specimen GIN no. 14801-20009-1: (4, 5, 7, 9) enlarged fragments; (6) specimen GIN no. 14801-20017-1. All specimens are from the Ura Formation.

Macrofossils of the Ura Formation. (1–3) Gen. et sp. indet. 1: (1, 2) specimen GIN no. 14801-20012-1; (3) specimen GIN no. 14801-20012-2; (4, 5) Gen. et sp. indet. 2, specimen GIN no. 14801-20018-1; (6) Jiuqunaoella simplicis Chen, 1991: specimen GIN no. 14801-20014-1; (7) carbonaceous conical film, specimen GIN no. 14801-17140-1; (8–10) Tawuia dalensis Hofmann, 1979: (8) specimen GIN no. 14801-17148-1; (9) specimen GIN no. 14801-17148-2; (10) specimen GIN no. 14801-20019-1. All specimens are from the Ura Formation.

Interpretation of fragment of colony of Liulingjitaenia alloplecta (а) and fragment of ribbon impression showing the change in the direction of twisting (b). Straight arrows indicate the areas which exhibit the change in the direction of the ribbon twisting (from right to left, from left to right).
Our interpretation follows the observations of these filamentous structures in paired impressions. At mechanical division of the bed, the compression of the fossil organism can be divided into the lower and upper halves. In our case, the filamentous structures in the paired impressions become symmetrically inverted mirror images; thus, the bottom and the top of the bed exhibit divided layers of filaments and the lower and the upper parts of the ribbon spiral. The transverse or oblique sections of the ribbons, which are composed of several filament layers, are also observed in thin sections (Fig. 2f). This initial form of Liulingjitaenia alloplecta is indirectly supported by very consistent linearity of phytoleims observed along their entire length. A spirally twisted ribbon, as well as a tube, is the most stable to an inflection along all directions. In contrast to the tube, however, this ribbon is flexible for both bending and extension. After cessation of a strain, the ribbon returned to the original state without deformations. It is possible that this architecture facilitated the increase in stability of the construction of a narrow thin elongated body, which could have been relevant under turbulent conditions.
Numerous one-stage buried specimens of L. alloplecta indicate a colonial mode of life of this species. In colony aggregates, the individuals grew in one direction and crossed each other at a low (<5°) angle, forming loose entanglements (Fig. 3). The margins of adjacent spirals fixed the construction of the entire colony, preventing the destruction during transport; thus, their transported and buried fragments represented in vivo organization of these colonies. No holdfasts of these organisms have been revealed in the fossil material, which can indicate their floating or seafloor covering style of habitat. The absence of the identified Liulingjitaenia remains in deep-sea distal parts of the basin could also be related both to coastal (sublittoral) habitat and to various burial conditions. The finding of micro- and macroscopic fossil organisms in the same narrow facies zones of the Ura Basin indicates their affinity for the same biofacies and suggests a leading role of taphonomy in the facies distribution of both fossil communities (Vorob’eva and Petrov, 2020).
Another representative of the Miaohe biota is Jiuqunaoella simplicis Chen (Plate II, fig. 6). These wormlike organisms were found only on one surface, although their possible small (few millimeters) fragments locally occur among abundant carbonaceous detritus in various layers of the section together with rich assemblages of the Ura acanthomorphic acritarchs. Our specimens are compressions of tubular organisms <1 cm long with closed typically curved ends and rare transverse folds of possible compression.
In addition to Liulingjitaenia and Jiuqunaoella, the Ura sedimentary rocks contain various segmented ribbon-like impressions described as Gen. et sp. indet. 1 (Plate II, figs. 1–3) and Gen. et sp. indet. 2 (Plate II, figs. 4, 5). In morphology and sizes, they cannot be ascribed to any known taxa, whereas the amount and quality of the material prevent description of new species. The representatives of Gen. et sp. indet. 1 slightly resemble Calyptrina Sokolov (Sokolov, 1965; Xiao et al., 2002; Ye et al., 2017), whereas Gen. et sp. indet. 2 are similar to Proterozoic fossils Protoarenicola Wang (Wang, 1982). Among the abundant organic detritus on the surfaces of beds, the Ura sedimentary deposits host abundant oval carbonaceous films ascribed to Tawuia dalensis Hofmann (Plate II, figs. 8–10), as well as conical compressions (Plate II, fig. 7). In contrast to the ones described above, these fossils occur along the entire section of the Ura Formation.
Morphological Similarity and Possible Biological Nature of Liulingjitaenia and Jiuqunaoella
The absence of cellular microstructures and biomarker data indicate the biological relation of these macroscopic fossils only on the basis of the general comparative morphology. Chen and Xiao (1992), who first described the genus Liulingjitaenia, compared these forms with cyanobacteria. Steiner (1994) also shared this viewpoint and ascribed these fossils to large cyanobacteria colonies. Later, Xiao et al. (2002) suggested that siphonous green algae are the possible morphological analogs of Liulingjitaenia. At present, the genus Liulingjitaenia belongs to macrophytes (Xiao et al., 2020), although some authors do not exclude their cyanobacterial nature (Ye et al., 2017). Actually, the cyanobacteria colonies tend to self-organization, locally producing ordered macrostructures (Sim et al., 2012; Sumina and Sumin, 2013). The cyanobacteria trichomes (especially, when they occur in clumps with rare exceptions), however, are surrounded by a dense common sheath of exopolymeric substance. This sheath protects both the cells of the individual trichome and the entire colony and its preservation in a fossil record is much higher in comparison with cells. No sheath can be identified for Liulingjitaenia; only remains of the parallel filaments of a small amount of diffuse organic substances are observed between them. This does not allow us to consider these fossils the possible remains of anomalously large colonies of filamentous cyanobacteria, which produced no extracellular polymers and were characterized by complex level of organization atypical of cyanobacteria.
Liulingjitaenia alloplecta is similar in morphology to some Late Ediacaran and Early Paleozoic forms, in particular, Late Vendian–Early Cambrian taxon Harlaniella Sokolov. Ivantsov (2013) indicates a morphological similarity between Liulingjitaenia alloplecta and species Harlaniella ingriana Ivantsov, which was recognized by him, suggesting that L. alloplecta is only a taphonomic variety of the empty tube H. ingriana.
The Cambrian monotype genus Fuxianospira Chen et Zhou is the closest to Liulingjitaenia in morphology and size. Although this taxon is described as an entire twisted body, several specimens of Fuxianospira gyrata exhibit fragments of detached filamentous structures (e.g., Wang et al., 2021, figs. 4-C, 5-A(S1)). These structures are similar to L. alloplecta. Initially, Fuxianospira was described as a macroalga (Chen and Zhou, 1997) and was later interpreted as a possible coprolite (Steiner et al., 2005). Further detailed analysis of these fossils did not show any typical coprolite features and enabled the identification of Fuxianospira (as well as Liulingjitaenia) as siphonous algae (LoDuca et al., 2015; Wang et al., 2021). These authors compare Fuxianospira with cladophore green alga Chaetomorpha, the fibrous twisted structure of which is formed by series of helically arranged microfibrillae inside a cellulose cellular wall.
The macrofossils of another Ediacaran genus Jiuqunaoella were primarily interpreted as remnants of wormlike animals (Chen and Xiao, 1991; Ding et al., 1996) and were further ascribed to macrophytes (Xiao et al., 2002). A weak transverse fragmentation on Jiuqunaoella impressions is the compression folds of the cylindrical body. These macrofossils were interpreted as remains of a tubular organism, the morphology of which is similar to some species of coenocytic green algae (Xiao et al., 2002). Thus, the carbonaceous compressions of the Ura Formation are of macroalgae in nature. These fossils also have morphological analogs in the post-Shuram and Lower Cambrian beds.
SYSTEMATIC PALEONTOLOGY
Genus Jiuqunaoella Chen in M. Chen et Xiao, 1991 emend. Xiao, Yuan, Steiner et Knoll, 2002
Jiuqunaoella simplicis Chen in M. Chen et Xiao, 1991 emend. Xiao, Yuan, Steiner et Knoll, 2002
Plate II, fig. 6
Jiuqunaoella simplicis Chen: Chen and Xiao, 1991, p. 320, pl. 5, fig. 6; pl. 6, figs. 1, 2; Chen et al., 1994, pp. 396–397, pl. 4, figs. 7–9; Ding et al., 1996, p. 112, pl. 32, fig. 5; pl. 33, figs. 1, 4, 10; pl. 34, fig. 11; Xiao et al., 2002, p. 363, figs. 7.2–7.4, 8.1; Yuan et al., 2002, p. 61, fig. 69; Grazhdankin et al., 2007, p. 75, figs. 2g, 2h; Ye et al., 2017, p. 31, figs. 19A–19E; Bykova et al., 2020, p. 3, fig. 1A.
Vendotaenia sp.: Yuan et al., 1995, p. 98, pl. 2, fig. 9.
Jiuqunaoella convoluta Li: Ding et al., 1996, p. 112, pl. 33, >figs. 2, 3.
Jiuqunaoella minia Li: Ding et al., 1996, pp. 112–113, pl. 32, fig. 6.
Jiuqunaoella sp.: Ding et al., 1996, pl. 34, fig. 6.
Description. Carbonaceous ribbon-like structures on the bedding surface of rocks. The ribbons are long, smooth, curved, closed from one end. The terminations are elongated curved, with rare transverse compression folds. The length of the ribbons is up to 35 mm. The width of the ribbons is consistent along the entire length (1.1–1.2 mm), narrowing only to the terminations to 0.5–0.7 mm.
Occurrence. China, the upper part of the Doushantuo Formation, Miaohe Member, Ediacaran. Russia: East European Platform, Lyamitsa Formation, Upper Vendian; Siberian Platform, Khatyspyt Formation, Upper Vendian; Baikal–Patom Highland, Ura Formation, Vendian.
Material. Two incomplete specimens.
Genus Liulingjitaenia M. Chen et Xiao, 1992
Liulingjitaenia alloplecta M. Chen et Xiao, 1992
Plate I, figs. 1–9
Liulingjitaenia alloplecta Chen et Xiao: 1992, p. 515, pl. 1, figs. 1–4; Grazhdankin et al., 2007, p. 76, fig. 3f; Wang еt al., 2007, p. 833, pl. 2, figs. 2–5; Marusin et al., 2011, p. 661, fig. 2b; Wang еt al., 2014, p. 971, figs. 4g, 4h; Ye еt al., 2017, p. 35, figs. 21A–21H.
Eoscytosiphon longitubulosum Ding: Ding et al., 1996, p. 88, pl. 22, figs. 3, 6.
Liulingjitaenia alloplecta Chen et Xiao, emend. Steiner: Xiao еt al., 2002, p. 362, figs. 6.7–6.9, 6.12–6.13 (and synonymic in this work); Yuan еt al., 2002, p. 62, figs. 71, 72.
Problematicum type B: Hofmann and Mountjoy, 2010, p. 1315, fig. 7E.
Liulingjitaenia alloplecta: An еt al., 2015, p. 248, fig. 4h.
Description. Long narrow ribbons as carbonaceous compressions on the bedding surface. The ribbons are straight or slightly curved, twisted in oblique-transverse direction. The twisting of the ribbon is dense, up to 8 whorls per 1 cm of length. The width of the ribbons ranges from 1 to 3.5 mm and is consistent along the entire length. No termination of the ribbons is observed. The length of the fragments is up to 8 cm. The ribbons consist of the longitudinal thin (10–30 µm) filaments (Plate I, figs. 5, 7–9).
Remarks. In initial diagnosis, Chen and Xiao (1992) described Liulingjitaenia alloplecta as a long thallus that has a spiral form and consists of longitudinal filaments. The diagnosis was later corrected and L. alloplecta was described as narrow ribbon impressions of initially tube membrane organisms or colonies of organisms with spiral folds formed by twisting (Steiner, 1994; Xiao et al., 2002). Later, most researchers returned to the original diagnosis of Chen and Xiao (1992) (Hofmann and Mountjoy, 2010; Xiao et al., 2013, 2020; Ye et al., 2017). Our material shows that the organism was a volumetric spiral-twisted ribbon rather than a twisted cylinder. The ribbon was formed by fine fiber filaments oriented along the growth line. This structure of the body is most similar to the primary diagnosis of Liulingjitaenia (Chen and Xiao, 1992), where the filament bundles (rather than the folds of the tube) make a typical texture of phytoleims.
Occurrence. South China: Hubei Province, the upper part of the Doushantuo Formation, Miaohe Member, Ediacaran; Guizhou Province, the upper part of the Doushantuo Formation, Wenghui biota, Ediacaran. Canada: Britain Columbia, the upper part of the Miette Group, Ediacaran. Russia: Central Urals, Perevalok Formation, Upper Vendian; Siberian Platform, Khatyspyt Formation, Upper Vendian; Baikal–Patom Highland, Ura Formation, Vendian.
Material. More than 30 specimens.
Gen. et sp. indet. 1
Plate II, figs. 1–3
Description. Fragments of curved ribbons preserved as prominent impressions of narrow convex crinkles, which form in places of deep folds of the organism body and organic substances between them. The lateral margin of the ribbon is curved. The length of incomplete fragments is up to 30 mm. The width of the ribbons varies from 2.0 to 2.5 mm but slightly narrows to the ends to 1 mm. The lancet-shaped crinkles are 0.7–1.0 mm wide in a medium part. They locally cross the ribbon along the entire width and often reach only one-third or half of the width of the impression. The crinkles are oriented at an angle of 90° relatively to the phytoleims. The intervals between crinkles are 0.4–0.9 mm.
Comparison. From the genus Calyptrina Sokolov, 1967, they differ in smaller sizes and the absence of a typical “cone-in-cone” tubular structure.
Occurrence. Russia, Baikal–Patom Highland, Ura Formation, Vendian.
Material. Two incomplete specimens.
Gen. et sp. indet. 2
Plate II, figs. 4, 5
Description. Fragments of organism preserved as segmented carbonaceous ribbon-like compressions on the bedding surface of rocks. The ribbons are smooth-walled, rigid, straight, and gently curved from one termination. The fragments are 15 to 25 mm in length and the width is relatively constant (1.2–1.5 mm) along the compressions and slightly narrows only to the termination to 0.5–0.9 mm. The length of the segments is uneven (from 0.8 to 2.2 mm). In junction of the segments, the outer contour of the ribbons is slightly constricted. The intrasegmental constrictions are straight and clear.
Comparison. From the genus Protoarenicola Wang, 1982, emend. Dong et al., 2008, our specimens differ in rarer segmentation and the absence of terminal disks.
Occurrence. Russia, Baikal–Patom Highland, Ura Formation, Vendian.
Material. Two incomplete specimens.
CONCLUSIONS
The carbonaceous macrofossils typical of the younger post-Shuram (<570 Ma) Mioahe biota are found in the Lower Vendian (pre-Shuram, 580–570 Ma) fine-grained siliciclastic rocks of the Ura Formation together with diverse assemblages of acanthomorphic acritarchs. The Ura macrofossils include the genera Liulingjitaenia alloplecta and Jiuqunaoella simplicis, as well as unrecognized segmented ribbon-like impressions and carbonaceous compressions corresponding to the diagnosis of Tawuia dalensis.
The extended ribbon-like remnants of L. alloplecta, which were characterized by the in vivo form of helically twisted ribbons, are most abundant in the Ura Formation. These organisms formed large floating or bottom covering colonies composed of unidirectional individuals. The majority of the Ura macrofossils included macrophytes, which were a significant detritus-dominant component of the paleobasin. Together with eukaryote microphytoplankton, this detritus composed a main part of buried organic matter.
The macrophytes of the Ura Formation coexisted with the Early Ediacaran microbiota and, evidently, composed a common ecosystem, which was formed just before the Shuram event and was significantly modified after it.
REFERENCES
An, Z., Jiang, G., Tong, J., Tian, L., Ye, Q., Song, H., and Song, H., Stratigraphic position of the Ediacaran Miaohe biota and its constraints on the age of the upper Doushantuo δ13C anomaly in the Yangtze Gorges area, South China, Precambrian Res., 2015, vol. 271, pp. 243–253. https://doi.org/10.1016/j.precamres.2015.10.007
Bobrov, A.K., Geologiya Predbaikal’skogo kraevogo progiba (Geology of the Cis-Baikalian Foredeep), Moscow: Nauka, 1964 [in Russian].
Bobrov, A.K. Stratigrafiya i paleogeografiya otlozhenii verkhnego dokembriya Yuzhnoi Yakutii (Stratigraphy and Paleogeography of Upper Precambrian Rocks in Southern Yakutia), Yakutsk: Yakutsk. Knizhn. Izd., 1979 [in Russian].
Bykova, N., LoDuca, S.T., Ye, Q., Marusin, V., Grazhdankin, D., and Xiao, S., Seaweeds through time: Morphological and ecological analysis of Proterozoic and early Paleozoic benthic macroalgae, Precambrian Res., 2020, vol. 350, p. 105875. https://doi.org/10.1016/j.precamres.2020.105875
Canfield, D.E., Poulton, S.W., and Narbonne, G.M., Late Neoproterozoic deep-ocean oxygenation and the rise of animal life, Science, 2007, vol. 315, pp. 92–95. https://doi.org/10.1126/science.1135013
Canfield, D.E., Knoll, A.H., Poulton, S.W., Narbonne, G.M., and Dunning, G.R., Carbon isotopes in clastic rocks and the Neoproterozoic carbon cycle, Am. J. Sci., 2020, vol. 320, pp. 97–124. https://doi.org/10.2475/02.2020.01
Chen, M. and Xiao, Z., Discovery of the macrofossils in the Upper Sinian Doushantuo Formation at Miaohe, eastern Yangtze Gorges, Scientia Geol. Sinica, 1991, no. 4, pp. 317–324.
Chen, M. and Xiao, Z., Macrofossil biota from upper Doushantuo Formation in eastern Yangtze Gorges, China, Acta Palaeontol. Sinica, 1992, vol. 31, pp. 513–529.
Chen, J.Y. and Zhou, G.Q., Biology of the Chengjiang fauna, Bull. Nat. Mus. Sci., 1997, vol. 10, pp. 11–106.
Chen, M., Xiao, Z., and Yuan, X., A new assemblage of megafossils – Miaohe biota from Upper Sinian Doushantuo Formation, Yangtze Gorges, Acta Palaeontol. Sinica, 1994, vol. 33, pp. 391–403.
Chumakov, N.M., Glaciations of the Earth. History, stratigraphic significance, and role in the biosphere, in Tr. GIN RAN. Vyp. 611 (Trans. Geol. Inst. Russ. Acad. Sci. Vol. 611), Moscow: GEOS, 2015 [in Russian].
Chumakov, N.M., Pokrovskii, B.G., and Melezhik, V.A., Geological history of the Patom Complex, Late Precambrian, Central Siberia, Dokl. Earth Sci., 2007, vol. 413, no. 3, pp. 355–358.
Chumakov, N.M., Semikhatov, M.A., and Sergeev, V.N., Vendian reference section of southern Middle Siberia, Stratigr. Geol. Correl., 2013, vol. 21, no. 4, pp. 359–382. https://doi.org/10.7868/S0869592X13040029
Cloetingh, S., McQueen, H., and Lambeck, K., On a tectonic mechanism for regional sea level variations, Earth Planet. Sci. Lett., 1985, vol. 75, pp. 157–166. https://doi.org/10.1016/0012-821X(85)90098-6
Ding, L., Li, Y., Hu, X., Xiao, Y., Su, C., and Huang, J., Sinian Miaohe Biota, Beijing: Geol. Publ. House, 1996.
Dong, L., Xiao, S., Shen, B., Yuan, X., Yan, X., and Peng, Y., Restudy of the worm-like carbonaceous compression fossils Protoarenicola, Pararenicola, and Sinosabellidites from early Neoproterozoic successions in North China, Palaeogeogr., Palaeoclimatol., Palaeoecol., 2008, vol. 258, pp. 138–161. https://doi.org/10.1016/j.palaeo.2007.05.019
Dornbos, S.Q., Oji, T., Kanayama, A., and Gonchigdorj, S., A new Burgess shale-type deposit from the Ediacaran of western Mongolia, Sci. Rep., 2016, vol. 6, pp. 1–5. https://doi.org/10.1038/srep23438
Fan, H., Nielsen, S.G., Owens, J.D., Auro, M., Shu, Y., Hardisty, D.S., Horner, T.J., Bowman, C.N., Young, S.A., and Wen, H., Constraining oceanic oxygenation during the Shuram excursion in South China using thallium isotopes, Geobiology, 2020, vol. 18, pp. 348–365. https://doi.org/10.1111/gbi.12379
Gong, Z., Kodama, K.P., and Li, Y.-X., Rock magnetic cyclostratigraphy of the Doushantuo Formation, South China, and its implications for the duration of the Shuram carbon isotope excursion, Precambrian Res., 2017, vol. 289, pp. 62–74. https://doi.org/10.1016/j.precamres.2016.12.002
Grazhdankin, D.V., Nagovitsin, K.E., and Maslov, A.V., Late Vendian Miaohe-type ecological assemblage of the East European Platform, Dokl. Earth Sci., 2007, vol. 417, no. 1, pp. 1183–1187.
Grazhdankin, D.V., Balthasar, U., Nagovitsin, K.E., and Kochnev, B.B., Carbonate-hosted Avalon-type fossils in Arctic Siberia, Geology, 2008, vol. 36, pp. 803–806. https://doi.org/10.1130/G24946A.1
Grotzinger, J.P., Fike, D.A., and Fischer, W.W., Enigmatic origin of the largest-known carbon isotope excursion in Earth’s history, Nat. Geosci., 2011, vol. 4, pp. 285–292. https://doi.org/10.1038/ngeo1138
Hofmann, H.J. and Mountjoy, E.W., Ediacaran body and trace fossils in Miette Group (Windermere Supergroup) near Salient Mountain, British Columbia, Canada, Can. J. Earth Sci., 2010, vol. 47, pp. 1305–1325. https://doi.org/10.1139/e10-070
Ivantsov, A. Yu., New data on Late Vendian problematic fossils from the genus Harlaniella, Stratigr. Geol. Correl., 2013, vol. 21, no. 6, pp. 592–600. https://doi.org/10.7868/S0869592X13060069
Ivantsov, A.Yu., Gritsenko, V.P., Konstantinenko, L.I., and Zakrevskaya, M.A., Revision of the problematic Vendian microfossil Beltanelliformis (=Beltanelloides, Nemiana), Paleontol. J., 2014, vol. 48, no. 13, pp. 1423–1448. https://doi.org/10.1134/S0031030114130036
Khomentovsky, V.V., Postnikov, A.A., Karlova, G.A., Kochnev, B.B., Yakshin, M.S., and Ponomarchuk, V.A., The Vendian of the Baikal–Patom Upland, Siberia, Russ. Geol. Geophys., 2004, vol. 45, no. 4, pp. 465–484.
Kochnev, B.B. and Karlova, G.A., New data on biostratigraphy of the Vendian Nemakit-Daldynian Stage in the Southern Siberian Platform, Stratigr. Geol. Correl., 2010, vol. 18, no. 5, pp. 492–504.
Kochnev, B.B., Pokrovskii, B.G., Kuznetsov, A.B., and Marusin, V.V., C and Sr isotope chemostratigraphy of Vendian–Lower Cambrian carbonate sequences in the central Siberian Platform, Russ. Geol. Geophys., 2018, vol. 59, no. 6, pp. 585–605.
Kolosov, P.N., Stratigrafiya verkhnego dokembriya yuga Yakutii (Upper Precambrian Stratigraphy of Southern Yakutia), Novosibirsk: Nauka, 1975 [in Russian].
Lambeck, K., Rouby, H., Purcell, A., Sun, Y., and Sa-mbridge, M., Sea level and global ice volumes from the Last Glacial Maximum to the Holocene, PNAS, 2014, vol. 111, pp. 15296–15303. www.pnas.org/cgi/doi/10.1073/ pnas.1411762111
Leonov, M.V. and Rud’ko, S.V., Finding of the Ediacaran–Vendian fossils in the Far Taiga Deposits, Patom highlands, Stratigr. Geol. Correl., 2012, vol. 20, no. 5, pp. 497–500. https://doi.org/10.31857/S0024497X20030052
LoDuca, S.T., Wu, M.Y., Zhao, Y.L., Xiao, S.H., Schiffbauer, J.D., Caron, J.-B., and Babcock, L.E., Reexamination of Yuknessia from the Cambrian of China and first report of Fuxianospira from North America, J. Paleontol., 2015, vol. 89, pp. 899–911. https://doi.org/10.1017/jpa.2016.3
Marusin, V.V., Grazhdankin, D.V., and Maslov, A.V., Redkino Stage in evolution of Vendian macrophytes, Dokl. Earth Sci., 2011, vol. 436, no. 2, pp. 197–202.
Melezhik, V.A., Fallick, A.E., and Pokrovsky, B.G., Enigmatic nature of thick sedimentary carbonates depleted in 13C beyond the canonical mantle value: the challenges to our understanding of the terrestrial carbon cycle, Precambrian Res., 2005, vol. 137, pp. 131–165. https://doi.org/10.1016/j.precamres.2005.03.010
Melezhik, V.A., Pokrovsky, B.G., Fallick, A.E., Kuznetsov, A.B., and Bujakaite, M.I., Constraints on 87Sr/86Sr of Late Ediacaran seawater: insight from Siberian high-Sr limestones, J. Geol. Soc., 2009, vol. 166, pp. 183–191. https://doi.org/10.1144/0016-76492007-171
Metelkin, D.V., Vernikovsky, V.A., and Kazansky, A.Yu., Tectonic evolution of the Siberian paleocontinent from the Neoproterozoic to the Late Mesozoic: Paleomagnetic record and reconstructions, Russ. Geol. Geophys., 2012, vol. 53, no. 7, pp. 675–688. https://doi.org/10.1016/j.rgg.2012.05.006
Minguez, D., Kodama, K.P., and Hillhouse, J.W., Paleomagnetic and cyclostratigraphic constraints on the synchroneity and duration of the Shuram carbon isotope excursion, Johnnie Formation, Death Valley Region, CA, Precambrian Res., 2015, vol. 266, pp. 395–408. https://doi.org/10.1016/j.precamres.2015.05.033
Moczydłowska, M. and Nagovitsin, K.E., Ediacaran radiation of organic-walled microbiota recorded in the Ura Formation, Patom Uplift, East Siberia, Precambrian Res., 2012, vol. 198-199, pp. 1–24. https://doi.org/10.1016/j.precamres.2011.12.010
Narbonne, G.M., Neoproterozoic oceans and early animals, Science, 2010, vol. 328, pp. 53–54. https://doi.org/10.1126/science.1188688
Pelechaty, S.M., Integrated chronostratigraphy of the Vendian System of Siberia: implications for a global stratigraphy, J. Geol. Soc., 1998, vol. 155, pp. 957–973. https://doi.org/10.1144/gsjgs.155.6.0957
Petrov, P.Yu., Postglacial deposits of the Dal’nyaya Taiga Group: Early Vendian in the Ura Uplift, Siberia. Communication 1. Barakun Formation, Lithol. Miner. Resour., 2018a, vol. 53, no. 5, pp. 417–429. https://doi.org/10.31857/S0869592X20040109
Petrov, P.Yu., Postglacial deposits of the Dal’nyaya Taiga Group: Early Vendian in the Ura Uplift, Siberia. Communication 2. Ura and Kalancha formations and history of the basin, Lithol. Miner. Resour., 2018b, vol. 53, no. 6, pp. 473–488. https://doi.org/10.1134/S0024497X18060083
Petrov, P.Yu. and Pokrovsky, B.G., Carbon Isotope Evidence of methanogenesis in sediments of the Dal’nyaya Taiga Group (Lower Vendian of the Patom Basin, Siberia), Lithol. Miner. Resour., 2020, vol. 55, no. 2, pp. 83–94. https://doi.org/10.31857/s0024497x20020068
Pokrovskii, B.G., Melezhik, V.A., and Buyakaite, M.I., Carbon, oxygen, strontium, and sulfur isotopic compositions in late Precambrian rocks of the Patom Complex, central Siberia: Communication 1. Results, isotope stratigraphy, and dating problems, Lithol. Miner. Resour., 2006a, vol. 41, no. 5, pp. 450–474.
Pokrovskii, B.G., Melezhik, V.A., and Buyakaite, M.I., Carbon, oxygen, strontium, and sulfur isotopic compositions in late Precambrian rocks of the Patom Complex, central Siberia: Communication 2. Nature of carbonates with ultralow and ultrahigh δ13C values, Lithol. Miner. Resour., 2006b, vol. 41, no. 6, pp. 576–587.
Pokrovskii, B.G. and Bujakaite, M.I., Geochemistry of C, O, and Sr isotopes in the Neoproterozoic carbonates from the southwestern Patom paleobasin, southern Middle Siberia, Lithol. Miner. Resour., 2015, vol. 50, no. 2, pp. 144–169. https://doi.org/10.7868/S0024497X15010048
Powerman, V., Shatsillo, A., Chumakov, N., Kapitonov, I., and Hourigan, J., Interaction between the Central Asian Orogenic Belt (CAOB) and the Siberian Craton as recorded by detrital zircon suites from Transbaikalia, Precambrian Res., 2015, vol. 267, pp. 39–71. https://doi.org/10.1016/j.precamres.2015.05.015
Rooney, A.D., Cantine, M.D., Bergmann, K.D., Gómez-Pérez, I., Al Baloushi, B., Boag, T.H., Busch, J.F., Sperling, E.A., and Strauss, J.V., Calibrating the coevolution of Ediacaran life and environment, Proc. Natl. Acad. Sci. U.S.A., 2020, vol. 117, no. 29, pp. 16824–16830. https://doi.org/10.1073/pnas.2002918117
Rowland, S.M. and Rodriguez, M.G., A multicellular alga with exceptional preservation from the Ediacaran of Nevada, J. Paleontol., 2014, vol. 88, pp. 263–268. https://doi.org/10.1666/13-075
Rud’ko, S.V., Petrov, P.Yu., Kuznetsov, A.B., Shatsillo, A.V., and Petrov, O.L., Refined δ13C trend of the Dal’nyaya Taiga Group of the Ura Uplift (Vendian, southern part of Middle Siberia), Dokl. Earth Sci., 2017, vol. 477, no. 2, pp. 1449–1453. https://doi.org/10.7868/S0869565217350183
Rud’ko. S.V., Kuznetsov. A.B., and Petrov. P.Yu., Strontium isotope composition in limestones of the Dal’nyaya Taiga Group in the Patom Basin: Vendian reference section of Siberia, Lithol. Miner. Resour., 2020, vol. 55, no. 3, pp. 206–217. https://doi.org/10.31857/S0024497X20030052
Rud’ko, S.V., Kuznetsov, A.V., Petrov, P.Yu., Sitkina, D.R., and Kaurova, O.K., Pb–Pb dating of the Dal’nyaya Taiga Group in the Ura uplift of southern Siberia: Implication of C-isotopic and biotic events in the Ediacaran, Precambrian Res., 2021, vol. 362. https://doi.org/10.1016/j.precamres.2021.106285
Sahoo, S.K., Planavsky, N.J., Jiang, G., Kendall, B., Owens, J.D., Wang, X., Shi, X., Anbar, A.D., and Lyons, T.W., Oceanic oxygenation events in the anoxic Ediacaran ocean, Geobiology, 2016, vol. 14, pp. 457–468. https://doi.org/10.1111/gbi.12182
Schieber, J., Microbial mats in terrigenous clastics: The challenge of identification in the rock record, Palaios, 1999, vol. 14, pp. 3–12. https://doi.org/10.2307/3515357
Schieber, J., Microbial mats on muddy substrates — examples of possible sedimentary features and underlying processes, in Atlas of Microbial Mat Features Preserved within the Siliciclastic Rock Record, Atlases in Geoscience 2, Schieber, J., Bose, P.K., Eriksson, P.G., Banerjee, S., Sarkar, S., Altermann, W., and Catuneanu, O., Eds., Elsevier, 2007, pp. 117–134.
Sergeev, V.N., Knoll, A.H., and Vorob’eva, N.G., Ediacaran microfossils from the Ura Formation, Baikal-Patom Uplift, Siberia: Taxonomy and biostratigraphic significance, J. Paleontol., 2011, vol. 85, pp. 987–1011. https://doi.org/10.1666/11-022.1
Sim, M.S., Liang, B., Petroff, A., Evans, A., Klepac-Ceraj, V., Flannery, D.T., Walter, M.R., and Bosak, T., Oxygen-dependent morphogenesis of modern clumped photosynthetic mats and implications for the Archean stromatolite record, Geosciences, 2012, vol. 2, pp. 235–259. https://doi.org/10.3390/geosciences2040235
Simonson, B.M. and Carney, K.E., Roll-up structures: evidence of in situ microbial mats in Late Archaean deep shelf environments, Palaios, 1999, vol. 14, pp. 13–24. https://doi.org/10.2307/3515358
Sokolov, B.S., Sokolov, B.S., Ancient Early Cambrian deposits and sabelliditids, in Vsesoyuz, simp. po paleontologii dokembriya i rannego kembriya (All-Union Symp. on Precambrian and Lower Cambrian Paleontology), Novosibirsk, 1965, pp. 78–91.
Sovetov, J.K., Vendian foreland basin of the Siberian cratonic margin: Paleopangean accretionary phases, Rus. J. Earth Sci., 2002, vol. 4, no. 5, pp. 363–387. https://doi.org/10.2205/2002ES000092
Steiner, M., Die Neoproterozoischen megaalgen sudchinas, Berl. Geowiss. Abh., R. (E), 1994, vol. 15, pp. 1–146.
Steiner, M., Zhu, M., Zhao, Y., and Erdtmann, B.D., Lower Cambrian Burgess shale-type fossil associations of South China, Palaeogeogr., Palaeoclimatol., Palaeoecol., 2005, vol. 220, pp. 129–152. https://doi.org/10.1016/j.palaeo.2003.06.001
Sumina, E.L. and Sumin, D.L., Morphogenesis in a community of filamentous cyanobacteria, Russ. J. Development. Biol., 2013, vol. 44, no. 3, pp. 158–172. https://doi.org/10.7868/S0475145013030087
Vorob’eva, N.G. and Petrov, P.Yu., Microbiota of the Barakun Formation and biostratigraphic characteristics of the Dal’nyaya Taiga Group: Early Vendian of the Ura Uplift (Eastern Siberia), Stratigr. Geol. Correl., 2020, vol. 28, no. 4, pp. 365–380. https://doi.org/10.31857/S0869592X20040109
Vorob’eva, N.G. and Sergeev, V.N., Stellarossica gen. nov. and the Infragroup Keltmiides infragroup. nov.: Extremely large acanthomorph acritarchs from the Vendian of Siberia and the East European Platform, Paleontol. J., 2018, vol. 52, no. 5, pp. 563–573. https://doi.org/10.1134/S0031031X18040141
Wang, G., Late Precambrian Annelida and Pogonophora from the Huainan of Anhui Province, Bull. Tianjin Inst. Geol. Miner. Resour., 1982, vol. 6, pp. 9–22.
Wang, Y., Wang, X., and Huang, Y., Macroscopic algae from the Ediacaran Doushantuo Formation in northeast Guizhou, South China, Earth Sci. J. China Univ. Geosci., 2007, vol. 32, pp. 828–844.
Wang, Y., Wang, Y., Du, D., and Wang, X., The correlation between macroscopic algae and metazoans in the Ediacaran: a case study on the Wenghui biota in northeastern Guizhou, South China, J. Aust. Earth Sci., 2014, vol. 61, pp. 967–977. https://doi.org/10.1080/08120099.2014.956231
Wang, P.-L., LoDuca, S.T., Wu, M.-Y., Tang, Y.-G., and Sun, Z.-X., Benthic primary producers in exceptionally preserved Cambrian biotas of North China, Palaeoworld, 2021. https://doi.org/10.1016/j.palwor.2021.01.009
Witkosky, R. and Wernicke, B.P., Subsidence history of the Ediacaran Johnnie Formation and related strata of southwest Laurentia: implications for the age and duration of the Shuram isotopic excursion and animal evolution, Geosphere, 2018, vol. 14, pp. 2245–2276. https://doi.org/10.1130/GES01678.1
Xiao, S. and Narbonne, G.M., The Ediacaran Period, in Geologic Time Scale. Vol. 1, Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M., Eds., Oxford: Elsevier, 2020, pp. 521–561. https://www.doi.org/10.1016/B978-0-12-824360-2.00018-8
Xiao, S., Yuan, X., Steiner, M., and Knoll, A.H., Macroscopic carbonaceous compressions in a terminal Proterozoic shale: a systematic reassessment of the Miaohe biota, South China, J. Paleontol., 2002, vol. 76, pp. 347–376. https://doi.org/10.1017/S0022336000041743
Xiao, Sh., Droser, M., Gehling, J.G., Hughes, I.V., Wan, B., Chen, Z., and Yuan, X., Affirming life aquatic for the Ediacaran biota in China and Australia, Geology, 2013, vol. 41, pp. 1095–1098. https://doi.org/10.1130/G34691.1
Xiao, S., Gehling, J.G., Evans, S.D., Hughes, I.V., and Droser, M.L., Probable benthic macroalgae from the Ediacaran Member, South Australia, Precambrian Res., 2020, vol. 350. https://doi.org/10.1016/j.precamres.2020.105903
Ye, Q., Tong, J., An, Z., Hu, J., Tian, L., Guan, K., and Xiao, S., A systematic description of new macrofossil material from the upper Ediacaran Miaohe Member in South China, J. Syst. Palaeontol., 2017, vol. 17, pp. 183–238. https://doi.org/10.1080/14772019.2017.1404499
Yuan, X., Li, J., and Chen, M., Development and their fossil records of metaphytes from late Precambrian, Acta Palaeontol. Sinica, 1995, vol. 34, pp. 90–102.
Yuan, X., Xiao, S., Yin, L., Knoll, A.H., Zhou, C., and Mu, X., Doushantuo Fossils: Life on the Eve of Animal Radiation, Hefei: China Univ. Sci. Technol. Press, 2002.
Zhang, F., Xiao, S., Romaniello, S.J., Hardisty, D., Li, C., Melezhik, V., Pokrovsky, B., Cheng, M., Shi, W., Lenton, T.M., and Anbar, A.D., Global marine redox changes drove the rise and fall of early animals, Geobiology, 2019, vol. 17, pp. 594–610. https://doi.org/10.1111/gbi.12359
ACKNOWLEDGMENTS
We are grateful to D.V. Grazhdankin, E.Yu. Golubkova, and M.A. Fedonkin for valuable criticism and suggestions, which improved the paper, as well as G.S. Feofanov for extensive help in preparation and conducting of field works.
Funding
This work was supported by the Russian Foundation for Basic Research, project no. 9-05-00155, and the Russian Science Foundation, project no. 20-77-10066.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare no conflicts of interest.
Additional information
Translated by I. Melekestseva
Reviewers E.Yu. Golubkova, D.V. Grazhdankin, and M.A. Fedonkin
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Petrov, P.Y., Vorob’eva, N.G. Representatives of the Miaohe Biota from the Ediacaran (Vendian) Pre-Shuram Strata of the Patom Highland, Siberia. Stratigr. Geol. Correl. 30, 52–64 (2022). https://doi.org/10.1134/S0869593822010051
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0869593822010051