
Abstract
In order to study the distribution of phylogenetic mtDNA lineages of Arctic charr Salvelinus alpinus in their contact zones in the north of East Siberia we analyzed nucleotide sequences of mtDNA control region of charr from 10 Arctic populations in the area from the Yenisei to the Lena. At the Putorana plateau, haplotypes of Atlantic and Siberian subgroups of Eurasian group were recorded, in the Khatanga River basin, haplotypes of Atlantic subgroup, in the Lena River delta, haplotypes of Siberian subgroup and of Bering group were observed. Some Siberian haplotypes found at the Putorana and in the Lena delta, have been earlier registered in other regions of East Siberia. New findings, along with published materials allow to specify the ranges of these three phylogenetic lineages and the margins of their contact zones; they also evidence wide sympatry of Atlantic and Siberian haplotypes in Taimyr water bodies and support close relationship of charr of Siberian subgroup from all main areas of their distribution.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
In Arctic charr Salvelinus alpinus and closely related species/forms, 5 phylogenetic groups of mtDNA control region haplotypes (Atlantic, Siberian, Acadian, Arctic and Beringian) were revealed (Brunner et al., 2001). We unite the first two groups in one Eurasian group and consider them as its subgroups (Gordeeva et al., 2018). Charr bearing haplotypes of these groups/subgroups have vicariant partly overlapping ranges (Brunner et al., 2001; Alekseyev et al., 2009; Moore et al., 2015; Osinov et al., 2017; Gordeeva et al., 2018; Salisbury et al., 2019). The areas of overlap are of special interest as the zones of postglacial secondary contacts and hybridization between these Pleistocene lineages (Osinov et al., 2015, 2017). Contact zones of Beringian and Arctic groups were studied in the north-west of North America (Moore et al., 2015) and in the north-east of Eurasia (Esin et al., 2017; Oleinik et al., 2017; Osinov et al., 2018); those of Arctic, Atlantic and Acadian groups, in Labrador and Newfoundland (Moore et al., 2015; Salisbury et al., 2019). In the north of East Siberia, the zones of secondary contact between Atlantic and Siberian, Siberian and Beringian groups/subgroups are known (Radchenko, 2003, 2004; Alekseyev et al., 2009; Osinov et al., 2015, 2017; Gordeeva et al., 2018). Genetic studies of charr from these remote areas are based on small number of individuals and populations and need to be significantly extended.
According to the initial description (Brunner et al., 2001), the Atlantic group is distributed in Eurasia to the east up to Kola Peninsula and the Siberian group, to the west up to Finland and Spitsbergen. Such a western position of the border between the ranges of the two groups was due to the fact that the authors included in the latter group several haplotypes, which were later transferred to the first one (Gordeeva et al., 2018). Taking into account this transfer and the new data, the updated range of the Siberian subgroup is restricted to continental water bodies of East Siberia; its present-day western border is located at the Putorana Plateau (Brunner et al., 2001; Osinov et al., 2017), while the eastern border of the Atlantic subgroup is in the Khatanga Bay basin (Brunner et al., 2001; Alekseyev et al., 2009). At the territory between the Khatanga and Kola Peninsula charr of the Atlantic subgroup were found in the Pyasina basin, including lakes of Putorana (Alekseyev et al., 2009; Moore et al., 2015; Osinov et al., 2017; Gordeeva et al., 2018), at Polar Urals and at Novaya Zemlya (Gordeeva et al., 2018).
In three water bodies of Taimyr Dolgano-Nenets region charr with haplotypes of both phylogenetic lineages were observed: in Lake Lama (based on restriction analysis of a section of ATPase6/ND4L: Radchenko, 2003; based on the analysis of nucleotide sequences of cytochrome b gene: Radchenko, 2004; Osinov et al., 2015; of mtDNA control region: Osinov et al., 2017), in Lake Kungasalakh and in the Novaya River (Alekseyev et al., 2009). All this shows that in Taimyr region a zone of secondary contact between Atlantic and Siberian subgroups is located (Osinov et al., 2015, 2017), but its precise boundaries are not yet known.
The same refers to the zone of contact between Arctic charr with haplotypes of Siberian subgroup and with haplotypes of Beringian group introgressed from northern Dolly Varden S. malma malma, which is situated to the East, in coastal regions from the Anabar to the Lena (Alekseyev et al., 2009; Gordeeva et al., 2018). Here haplotypes of both lineages co-occur in one lake in the Olenek Bay basin, while in the Anabar basin and in the Lena delta region they were recorded in different lakes. Analogous areas, in which Arctic charr or closely related Taranets charr captured mtDNA of northern Dolly Varden, were found in the Kolyma-Okhotsk region (Radchenko, 2003, 2004; Alekseyev et al., 2009; Osinov et al., 2017, 2018), and in Kamchatka (Esin et al., 2017).
The aim of the present study is to provide new data about the distribution of Arctic charr with haplotypes of three phylogenetic lineages in the zones of their secondary contacts in the north of East Siberia for specifying the boundaries of these zones and extending knowledge about phylogeography of this species.
MATERIALS AND METHODS
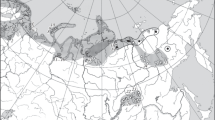
The material (39 individuals of Arctic charr) was collected in 10 Arctic water bodies of East Siberia (Table; Fig. 1a). It includes samples taken by the authors in the Khatanga basin, at Putorana Plateau (Fig. 1b) and in the Lena delta (Fig. 1c) in 2018–2020, as well as samples from two latter regions collected by the expeditions of the Department of Ichthyology of Moscow State University (MSU) in 1975–1976 and, with the participation of the second and the last authors, in 1979.

Distribution of three phylogenetic lineages of mtDNA control region haplotypes of Arctic charr Salvelinus alpinus in the north of Siberia: (а) schematic map of the region with ranges of groups and subgroups of mtDNA haplotypes; (b, c) satellite photographs of the western part of Putorana Plateau (b) and of a section of Olenek branch of the Lena River delta (c). The location of populations, in which mtDNA was studied: 1–9—our data; B—Brunner et al., 2001; A—Alekseyev et al., 2009; M—Moore et al., 2015; O—Оsinov et al., 2017, G—Gordeeva et al., 2018); ( ) and (
) and ( )—Atlantic subgroup of Eurasian group (including haplotypes SIB1-3,5,9), (⚪) and (
)—Atlantic subgroup of Eurasian group (including haplotypes SIB1-3,5,9), (⚪) and ( )—Siberian subgroup of Eurasian group, (⚫) and (
)—Siberian subgroup of Eurasian group, (⚫) and ( )—Beringian group (Arctic charr with haplotypes introgressed from northern Dolly Varden). Water bodies: 1—Lake Glubokoe, 2—Lake Sobach’e, 3—Lake Keta, 4—Lake Khantaiskoe, 5—Lake Ayan, 6—Lake Bayadzhaga, 7—Lake Dal’nee, 8—Lake Perekhodnoe, 9—Lake Bulkurka-2, 10—Lake Ladannakh-Kyuele (Forelevoe, Aranastakh). Scale: 20 km.
)—Beringian group (Arctic charr with haplotypes introgressed from northern Dolly Varden). Water bodies: 1—Lake Glubokoe, 2—Lake Sobach’e, 3—Lake Keta, 4—Lake Khantaiskoe, 5—Lake Ayan, 6—Lake Bayadzhaga, 7—Lake Dal’nee, 8—Lake Perekhodnoe, 9—Lake Bulkurka-2, 10—Lake Ladannakh-Kyuele (Forelevoe, Aranastakh). Scale: 20 km.
The surveyed water bodies significantly differ in their characteristics (Table 1). Three lakes in the Lena delta (Bulkurka-2, Dal’nee and Perekhodnoe), as well as the earlier studied lakes Gusinka ans Bulkurka (Alekseyev et al., 2009) are small low-lying nameless (the names were given by us and by the members of the expedition of MSU) thermokarst lakes located along a ~50-km section of the Olenek branch of the delta. These lakes are inhabited by monomorphic populations of Arctic charr represented by large form. A lake in the spurs of the Kharaulakh mountain range (Neelov Bay basin), which is designated at topographic maps as Ladannakh-Kyuele appears in the paper by Savvaitova and Maksimov (1980) under local name Forelevoe. The co-ordinates and the description given by Borisov (1932) leave no doubt that it is Lake Aranastakh; half-a-century earlier he described charr from this lake as a separate species “Yakut charr” S. jacuticus. In this lake, small and large forms of charr were found (Savvaitova and Maksimov, 1980), but the form of the studied individual is unknown.
The lakes of Putorana Plateau are large deep-water mountain lakes of glacial-tectonic origin hosting sympatric Arctic charr forms. Our samples included large long-rakered charr and small “typtushka” from Lake Khantaiskoe, “giant” charr, large lacustrine-riverine charr and small “putoranchik” from Lake Ayan, Drjagin’s charr and juveniles of “Boganida charr” from Lake Sobach’e. The latter were assigned to this form because they were caught in the lower section of River Khoronen, a tributary of Lake Sobach’e, where only “Boganida charr” spawns. Drjagin’s charr and the third form from Lake Sobach’e, the goggle-eye charr, spawn in the lake proper and were never observed in the river. The studied individuals from lakes Glubokoe and Keta also belong to “Boganida charr”. Bayadzhaga (the Khatanga basin) is a small lake inhabited by large form of Arctic charr.
Charr were sampled with 10, 20, 25, 30 and 40 mm knot-to-knot mesh size gillnets. DNA was isolated from adipose fins, pieces of muscles or of male gonads stored in 96% ethanol using commercial kit Diatom DNA Prep 100 (OOO Isogen Laboratory, Russia). A 534–543 bp fragment including the entire left domain of mitochondrial DNA control region was sequenced using the primers and procedures described in Alekseyev et al. (2009). For editing chromatograms and aligning sequences, the Geneious program (Kearse et al., 2012) was used. mtDNA fragments sequenced in this study, as well as in our previous study (Gordeev et al., 2018) overlap with the fragments sequenced by Brunner et al. (2001), Alekseyev et al. (2009), Moore et al. (2015) and Osinov et al. (2017) by 499–507 base pairs. New sequences revealed in this study were deposited in GenBank under accession numbers MT586474 (BER24), MT586475 (ATL32) and MT586476 (SIB35). Since sometimes similar (within the overlapping fragment) haplotypes are assigned different designations and different haplotypes are assigned the same designations, we use names of haplotypes, which were deposited in GenBank/NCBI first and in the cases of duplicate names give the first letter of the first author’s name in brackets. In order to verify the assignment of charr from new lakes Bayadzhaga and Bulkurka-2 to S. alpinus, we assessed their diagnostically important meristic characters, the numbers of gill rakers (sp.br.) and pyloric caeca (pc). Data on morphology and ecology of charr from other lakes under study can be found in papers by Borisov (1932); Savvaitova and Maksimov (1980); Romanov (1983, 1996, 2003); Savvaitova (1989); Pavlov et al. (1994); Pavlov (1997) and Pichugin (2009).
RESULTS AND DISCUSSION
Sequencing revealed 16 variable positions, including one indel, defining 8 haplotypes: three new ones (ATL32, SIB35 and BER24) and 5 earlier published ones: ATL21(M) (Moore et al., 2015), SIB4 (Brunner at al., 2001), SIB10, SIB20 and BER10 (Alekseyev et al., 2009). ATL21(M) is equivalent within the overlapping fragment to haplotype ATL20(O) (Osinov et al., 2017); SIB4, to haplotypes SIB31 (Moore et al, 2015) and SIB34 (Alekseyev et al., 2019).
In the contact zone of Siberian subgroup and Beringian group in the Lena delta region, Siberian haplotypes SIB10 and SIB35 were revealed in Lake Perekhodnoe, while Beringian haplotypes were revealed in lakes Dal’nee, Bulkurka-2 (BER10) and Ladannakh-Kyuele (BER24). It should be noted that earlier only one Siberian (SIB25) and one Beringian (BER10) haplotypes were reported from northern coastal regions of East Siberia (Alekseyev et al., 2009; Gordeeva et al., 2018). Lake Perekhodnoe is located close to the lakes, in which haplotype BER10 was found—in particular, only 6 km from Lake Dal’nee. Phenotypically charr from lakes Dal’nee, Bulkurka-2 and Ladannakh-Kyuele are Arctic charr, not Dolly Varden, since they have on average not less than 23.5 gill rakers and 34 pyloric caeca thus fitting into the variation limits of these diagnostic characters in the former species (Fig. 2).

Mean number of gill rakers (sp.br.) against mean number of pyloric caeca in populations of Arctic charr Salvelinus alpinus of Europe and Siberia ((– –)—Atlantic, (−⚪−)—Siberian subgroup of Eurasian group of haplotypes, (⚫)—Beringian group of haplotypes, charr from lakes 7, 9 and 10 in the Lena delta region) and northern Dolly Varden S. malma malma from Chukotka and Kamchatka ((- -▲- -)—Beringian group of haplotypes), based on original and literature data (Borisov, 1932; Esipov, 1952; Kolyushev, 1967; Nilsson, Filipsson, 1971; Balon, Penczak, 1980; Savvaitova, Maksimov, 1980; Chereshnev, 1982; Klemetsen, 1984; Chereshnev et al., 1989; Pervozvanskii and Shustov, 1999; Esin and Markevich, 2017). Lake numbers correspond to those in Fig. 1.
–)—Atlantic, (−⚪−)—Siberian subgroup of Eurasian group of haplotypes, (⚫)—Beringian group of haplotypes, charr from lakes 7, 9 and 10 in the Lena delta region) and northern Dolly Varden S. malma malma from Chukotka and Kamchatka ((- -▲- -)—Beringian group of haplotypes), based on original and literature data (Borisov, 1932; Esipov, 1952; Kolyushev, 1967; Nilsson, Filipsson, 1971; Balon, Penczak, 1980; Savvaitova, Maksimov, 1980; Chereshnev, 1982; Klemetsen, 1984; Chereshnev et al., 1989; Pervozvanskii and Shustov, 1999; Esin and Markevich, 2017). Lake numbers correspond to those in Fig. 1.
Within the contact zone of Atlantic and Siberian subgroups, in Lake Bayadzhaga (the Khatanga basin) Atlantic haplotype ATL32 was revealed, in Lake Khantaiskoe, in two charr forms (long-rakered charr and “typtushka”)—haplotype ATL21(M), which was earlier found in Lake Lama charr (Moore et al., 2015; Osinov et al., 2017). These are the easternmost and the southernmost extremes of the distribution of the Atlantic subgroup in East Siberia. Lake Bayadzhaga is situated near the border of the range of charr with Beringian group haplotypes, which were recorded in the neighbouring Anabar basin (Gordeeva et al., 2018). Long-rakered charr from Lake Khantaiskoe have the highest number of gill rakers among all Arctic charr of Atlantic subgroup (sp.br. 30–38 (33.1), Romanov, 2003); in this character they are similar with charr from some Transbaikalian populations, which belong to Siberian subgroup, for instance, with small-form charr from Lake Tokko (sp.br. 31–37 (33.6), Alekseyev et al., 2021). In the light of the results obtained this similarity can be qualified as a case of parallelism, though we cannot rule out the possibility of the substitution of mtDNA of charr of Siberian subgroup with that of Atlantic subgroup resulting from introgressive hybridization.
In Lake Sobach’e, in one individual of Drjagin’s charr Atlantic haplotype ATL32 was revealed and in two individuals of “Boganida charr” Siberian haplotype SIB20 was found, which was earlier observed only in Transbaikalia (Alekseyev et al., 2009; Gordeeva et al., 2018). In Lake Glubokoe, haplotype SIB10 was found in “Boganida charr”. Taking into account that earlier (Gordeeva et al., 2018) we reported Atlantic haplotype ATL22(M) from this lake, the list of water bodies with sympatric haplotypes of two subgroups is supplemented with two more lakes. It is important to note that in these three lakes of the Noril’skaya-Pyasina system (Lama, Sobach’e and Glubokoe) and in their tributaries Siberian haplotypes were found in “Boganida charr”, while Atlantic haplotypes (including SIB9) were found in other forms (Dryagin’s charr, goggle-eye charr, mountain charr (Brunner et al., 2001; Osinov et al., 2017; Gordeeva et al., 2018; present study)). The correspondence of these forms to two phylogenetic lineages so far revealed only in few individuals is highly interesting, but needs verification using more representative material. In Lake Keta such correspondence is violated since the studied individual of “Boganida charr” had Atlantic haplotype ATL21(M). However, misidentification of this individual cannot be ruled out since it was not preserved. In all forms from Lake Ayan we found Siberian haplotype SIB4, which was earlier revealed in two individuals of “putoranchik” from this lake (Brunner et al., 2001; Moore et al., 2015). In the latter paper it is designated as SIB31, although it is completely identical to 499-bp fragment in the sequence reported by Brunner et al (550 bp). SIB31 and SIB4 correspond to haplotype SIB34 reported for the large form of Arctic charr from Lake Maloe Leprindo in Transbaikalia (Alekseyev et al., 2019). Thus, presently 3 haplotypes are observed in common for Taimyr and Transbaikalia (SIB4, SIB10, SIB20).
Haplotype SIB10 is now found in all main areas of distribution of Siberian group: in Transbaikalia, in the basins of upper-middle Yana and Indigirka, in the Lena data and in Taimyr. Obviously, this most widespread haplotype is ancestral for Siberian subgroup (Gordeeva et al., 2018). We believe that Siberian group originated in the first half of Neopleistocene in giant Lena-Vilui paleolake (Lake Lena, Yakut “Sea”), which emerged at the territory of East Siberia in the periods of Pleistocene glaciations as the result of blocking of the northern flow of the Lena by the glacier gliding down from Verkhoyansk Range (Enikeev, 2009). This lake discharged into the Yenisei basin via the Vilui and the Nizhnyaya Tunguska and reached close to the main continental mountainous regions of East Siberia, which are nowadays inhabited by Arctic charr (Stanovoi and Oymyakon highlands, Verkhoyansk, Sette-Daban, Suntar-Khayata ranges), and also to northern coastal regions. It connected with Putorana via the Nizhnyaya Tunguska basin and lower reaches of the Yenisei. Dispersing throughout this lake, charr could quickly and easily reach these regions and settle in them. The dispersal of Siberian subgroup was probably provided exclusively by freshwater charr, unlike that of Atlantic subgroup, which was mainly provided by anadromous charr. Haplotype SIB10 obviously emerged in Lena-Vilyui lake, spread throughout this lake and in different regions gave rise to unique haplotypes restricted to these areas.
Thus, in the northern part of Arctic charr range from the Yenisei to the Lena new mitochondrial haplotypes of three phylogenetic lineages—of the Beringian group and of the Atlantic and Siberian subgroups of Eurasian group, as well as new populations of charr bearing these haplotypes were revealed; wider limits of the distribution of Atlantic group and its wider overlap with the range of Siberian group in the Khatanga basin and in the region of Putorana Plateau were demonstrated. New evidence was received that in lakes of Noril’skaya-Pyasina system (with the possible exception of Lake Keta) Siberian subgroup is represented by “Boganida charr” and Atlantic subgroup, by other charr forms dwelling in these lakes. The finding of three southern haplotypes of Siberian subgroup in the northern part of Arctic charr range supports the earlier conclusion about common origin and close phylogenetic relationships of Siberian group charr from different regions of East Siberia (Alekseyev et al., 2009; Gordeeva et al., 2018).
REFERENCES
Alekseyev, S.S., Bajno, R., Gordeeva, N.V., et al., Phylogeography and sympatric differentiation of the Arctic charr Salvelinus alpinus (L.) complex in Siberia as revealed by mtDNA sequence analysis, J. Fish Biol., 2009, vol. 75, no. 2, pp. 368–392. https://doi.org/10.1111/j.1095-8649.2009.02331.x
Alekseyev, S.S., Pichugin, M.Yu., Gordeeva, N.V., et al., Reproductive strategies and the origin of parapatric and sympatric forms of Arctic charr Salvelinus alpinus (Salmonidae) in the system of lakes Bol’shoe Leprindo and Maloe Leprindo (Northern Transbaikalia), J. Ichthyol. 2019, vol. 59, no. 4, pp. 527–544. https://doi.org/10.1134/S0032945219040015
Alekseyev, S.S., Samusenok, V.P., Yur’ev, A.L., et al., Morphological and ecological differentiation of sympatric forms of Arctic charr Salvelinus alpinus (Salmonidae) in lake Tokko (northern Transbaikalia), J. Ichthyol., 2021, vol. 61, no. 1, pp. 109–127. https://doi.org/10.1134/S0032945221010021
Balon, E.K. and Penczak, T., The dwarfed charr of Dösener See, an alpine lake in Austria, in Charrs: Salmonid Fishes of the Genus Salvelinus, Persp. Vertebr. Sci. vol. 1, Balon, E.K., Ed., Hague: Dr. W. Junk, 1980, pp. 773–794.
Borisov, P.G., New species of charrs—Salvelinus jacuticus n. sp., in Materialy k ikhtiofaune basseina r. Leny (Materials on Ichthyofauna of the Lena River Basin), Moscow: Akad. Nauk SSSR, 1932, pp. 1–42.
Brunner, P.C., Douglas, M.R., Osinov, A., et al., Holarctic phylogeography of Arctic charr (Salvelinus alpinus L.) inferred from mitochondrial DNA sequences, Evolution, 2001, vol. 55, no. 3, pp. 573–586. https://doi.org/10.1554/0014-3820(2001)055[0573:hpoacs]2.0.co;2
Chereshnev, I.A., Taxonomic status of anadromous charr of genus Salvelinus (Salmonidae) of eastern Chukotka, Vopr. Ikhtiol., 1982, vol. 22, no. 6, pp. 922–936.
Chereshnev, I.A., Gudkov, P.K. and Neiman, M.Yu., First data on the biology of anadromous Dolly Varden from the basin of River Chegitun’ (Arctic coast of eastern Chukotka), Vopr. Ikhtiol., 1989, vol. 29, no. 1, pp. 68–83.
Enikeev, F.I., Pleistocene glaciations in the Eastern Transbaikalia and the southeast of Middle Siberia, Geomorfologiya, 2009, vol. 2, pp. 33–49. https://doi.org/10.15356/0435-4281-2009-2-33-49
Esin, E.V. and Markevich, G.N., Gol’tsy roda Salvelinus aziatskoi chasti severnoi Patsifiki (Charrs of Genus Salvelinus from the Asian Part of Northern Pacific), Petropavlovsk-Kamchatskiy: Kamchatpress, 2017.
Esin, E.V., Bocharova, E., Mugue, N.S., and Markevich, G.N., Occurrence of sympatric charr groups, Salvelinus, Salmonidae, in the lakes of Kamchatka: a legacy of the last glaciations, J. Fish Biol., 2017, vol. 91, no. 2, pp. 628–644. https://doi.org/10.1111/jfb.13378
Esipov, V.K., Ryby Karskogo morya (Fishes of the Kara Sea), Leningrad: Akad. Nauk SSSR, 1952.
Gordeeva, N.V., Alekseyev, S.S., Kirillov, A.F., et al., Distribution, composition and relationships of phylogenetic groups of Arctic charr Salvelinus alpinus (L.) (Salmoniformes, Salmonidae) in the European part of Russia and in Siberia as revealed by the analysis of nucleotide sequences of mitochondrial DNA, J. Ichthyol., 2018, vol. 58, no. 6, pp. 808–818. https://doi.org/10.1134/S0032945218050089
Kearse, M., Moir, R., Wilson, A., et al., Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data, Bioinformatics, 2012, vol. 28, no. 12, pp. 1647–1649. https://doi.org/10.1093/bioinformatics/bts199
Klemetsen, A., The Arctic charr speciation problem as seen from northern Norway, Proc. Int. Symp. “Biology of the Arctic Charr,” Winnipeg: Univ. of Manitoba Press, 1984, pp. 65–77.
Kolyushev, A.I., Morphology and systematic position of charr from lakes of Kola Peninsula, Tr. Karel. Otd., Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 1967, vol. 5, no. 1, pp. 269–273.
Moore, J.-S., Bajno, R., Reist, J.D., and Taylor, E.B., Post-glacial recolonization of the North American Arctic by Arctic char (Salvelinus alpinus): genetic evidence of multiple northern refugia and hybridization between glacial lineages, J. Biogeogr., 2015, vol. 42, no. 11, pp. 2089–2100. https://doi.org/10.1111/jbi.12600
Nilsson, N.-A. and Filipsson, O., Characteristics of two discrete populations of Arctic charr (Salvelinus alpinus L.) in a north Swedish lake, Rep.–Inst. Freshwater Res., Drottningholm, 1971, vol. 51, pp. 90–108.
Oleinik, A.G., Skurikhina, L.A. and Kukhlevsky, A.D., Secondary contact between two divergent lineages of charrs of the genus Salvelinus in the Northwest Pacific, Russ. J. Genet., 2017, vol. 53, no. 11, pp. 1221–1233. https://doi.org/10.1134/S1022795417110084
Osinov, A.G., Senchukova, A.L., Mugue, N.S., et al., Speciation and genetic divergence of three species of charr from ancient Lake El’gygytgyn (Chukotka) and their phylogenetic relationships with other representatives of the genus Salvelinus, Biol. J. Linn. Soc., 2015, vol. 116, pp. 63–85. https://doi.org/10.1111/bij.12559
Osinov, A.G., Volkov, A.A., Alekseyev, S.S., et al., On the origin and phylogenetic position of Arctic charr (Salvelinus alpinus complex, Salmonidae) from Lake Cherechen` (middle Kolyma River basin): controversial genetic data, Polar Biol., 2017, vol. 40, pp. 777–786. https://doi.org/10.1007/s00300-016-2000-4
Osinov, A.G., Pavlov, D.A., and Volkov, A.A., On the origin of the lacustrine charr Salvelinus alpinus complex from the Kolyma and Sea of Okhotsk basins, J. Ichthyol., 2018, vol. 58, no. 3, pp 353–370. https://doi.org/10.1134/S0032945218030104
Pavlov, S.D., Sympatric forms of charrs (gen. Salvelinus) from Ayan Lake (Taimyr Peninsula), J. Ichthyol., 1997, vol. 37, no. 7, pp. 485–494.
Pavlov, S.D., Savvaitova, K.A. and Maksimov, V.A., The relationships of sympatric groups of Arctic charr in Lake Sobach’e (Noril’skaya-Pyasina aquatic system), in Sistematika, biologiya i biotekhnika razvedeniya lososevykh ryb (Systematics, biology and farming of salmonids), St. Petersburg: Gos. Nauchno-Issled. Inst. Ozern. Rechn. Rybn. Khoz., 1994, pp. 148–151.
Pervozvanskii, V. Ya. and Shustov, Yu. A., Dwarf form of char Salvelinus alpinus (Salmonidae) in Lake Verkhni Neris (Paanayarvi National Park, Karelia), J. Ichthyol., 1999, vol. 39, no. 1, pp. 125–126.
Pichugin, M.Yu., The development of an artificial hybrid and revealing elements of reproductive isolation between sympatric forms of Dryagin’s char and Salvelinus alpinus complex (Salmonidae) from Sobachye mountain lake (Taimyr), J. Ichthyol., 2009, vol. 49, no. 3, pp. 236–248.
Radchenko, O.A., Variability of mitochondrial DNA in populations of lake charrs from genus Salvelinus in the Far East and Siberia, J. Ichthyol., 2003, vol. 43, no. 4, pp. 539–547.
Radchenko, O.A., Variability of nucleotide sequences of mitochondrial DNA cytochrome b gene in charrs of the genus Salvelinus, Russ. J. Genet., 2004, vol. 40, no. 3. pp. 244–254.
Romanov, V.I., Ecological structure of charr (genus Salvelinus) from the Khantaiskoe Lake, Vopr. Geogr. Sib., 1983, no. 14, pp. 73–88.
Romanov, V.I., Population structure of charrs (genus Salvelinus) from the Keta Lake, Materialy konferentsii “Zadachi i problemy razvitiya rybnogo khozyaistva na vnutrennikh vodoemakh Sibiri” (Proc. Conf. “Challenges and Problems of Development of Fishery in Inland Water Bodies of Siberia”), Tomsk, 1996, pp. 97–98.
Romanov, V.I., Morphological peculiarities of mass forms of charr (genus Salvelinus) from the Khantaiskoe Lake, Vestn. Tomsk. Gos. Pedagog. Univ., Ser. Estestv. Tochnye Nauki, 2003, no. 4 (36), pp. 66–72.
Salisbury, S.J., McCracken, G.R., Keefe, D., et al., Extensive secondary contact among three glacial lineages of Arctic char (Salvelinus alpinus) in Labrador and Newfoundland, Ecol. Evol., 2019, vol. 9, no. 4, pp. 2031–2045. https://doi.org/10.1002/ece3.4893
Savvaitova, K.A., Arkticheskie gol’tsy (struktura populyatsionnykh sistem, perspektivy khozyaistvennogo ispol’zovaniya) (Arctic Charrs: Structure of Population Systems and Prospective Economic Use), Moscow: Agropromizdat, 1989.
Savvaitova, K.A. and Maksimov, V.A, Speciation in charr of genus Salvelinus (Salmoniformes, Salmonidae) from the Lena delta lakes, Zool. Zh., 1980, vol. 59, no. 12, pp. 1820–1830.
ACKNOWLEDGMENTS
The authors thank K.A. Savvaitova, V.A. Maksimov, V.R. Filin and S.Sh. Bagiryan-Mikhailova (Moscow State University) for the organization and implementation of the sampling expedition of the Moscow State University in 1975–1979; A.A. Lomov (Moscow State University) for preserving and providing the samples; S.V. Maksimov, S.D. Pavlov and Yu.V. Chebotareva (Moscow State University) for field assistance.
Funding
The study was funded by Russian Foundation for Basic Research (project no. 18-04-00092) and conducted under Government basic research programs of Institute of General Genetics (NVG) no. 0112-2019-0001 and Institute of Developmental Biology (SSA), no. 0088-2021-0019.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflicts of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by S.S. Alekseyev
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gordeeva, N.V., Alekseyev, S.S., Kirillov, A.F. et al. New Data about the Distribution of Three Phylogenetic Lineages of Arctic Charr Salvelinus alpinus (Salmonidae) in their Contact Zones in the North of East Siberia. J. Ichthyol. 61, 701–708 (2021). https://doi.org/10.1134/S0032945221050064
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0032945221050064