
Abstract
Due to its unique structure and properties, human breast milk lactoferrin (hLF) has many nutritional and health-promoting functions in infants, including protection against inflammation and bacterial infections. The lack of LF in breastmilk or formula can result in the weakening of the infant’s immune system. Noncompetitive polarization fluorescence immunoassay (FPIA) is a promising method for hLF quantification in milk and dairy products, which does not require the separation of the bound and free protein and allows to avoid time-consuming sample preparation. The use of fluorescently labeled single-domain camelid antibodies (nanobodies) for protein recognition in FPIA makes it possible to quantify relatively large antigens, in particular, hLF. In this work, we used previously obtained fluorescein isothiocyanate (FITC)-conjugated anti-hLF5 and anti-hLF16 nanobodies, which selectively recognized two different human lactoferrin epitopes, but did not bind to goat lactoferrin. The kinetics of hLF interaction with the FITC-labeled nanobodies was studied. The dissociation constant (KD) for the anti-LF5 and anti-LF16 nanobodies was 3.2 ± 0.3 and 4.9 ± 0.4 nM, respectively, indicating the high-affinity binding of these nanobodies to hLF. We developed the FPIA protocol and determined the concentration of FITC-labeled anti-hLF5 and anti-hLF16 nanobodies that provided the optimal fluorescence signal and stable fluorescence polarization value. We also studied the dependence of fluorescence polarization on the hLF concentration in the noncompetitive FPIA with FITC-anti-hLF5 nanobody. The detection limit for hLF was 2.1 ± 0.2 µg/ml and the linear range for determining the hLF concentration was 3-10 µg/ml. FPIA is commonly used to assay low-molecular-weight substances; however, the use of fluorescently labeled nanobodies allows quantification of high-molecular-weight proteins. Here, we demonstrated that FPIA with fluorescently labeled nanobodies can be used for hLF quantification in milk.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
INTRODUCTION
Lactoferrin (LF) is an 80-kDa protein (690 amino acids) from the transferrin family. Its amino acid sequence is 60% identical to the sequence of plasma transferrin [1, 2]. LF was for the first time isolated by Sorensen in 1939 from cow milk. In 1960, Johanson purified LF (hLF) from human breastmilk [1]. LF has be also identified in many biological fluids, such as saliva, tears, sperm, and mucous membrane secretions. It is an important component of the neutrophil granules of leukocytes [1, 3]. hLF is involved in multiple biological processes, including organism defense against pathogens and suppression of tumor growth; it possesses the antimicrobial, antibacterial, antiviral, and antiparasitic activities and regulates cell proliferation, cell differentiation, and activity of ribonuclease A [4-6]. LF efficiently binds Fe3+ ions (KD, ~10–20 M) [7] and plays an important role in the control of the free iron content in body fluids [8].
The concentration of LF in human breastmilk is higher than in cow milk [9]. Moreover, the content of LF significantly varies in different species and at different lactation periods. Human colostrum contains over 5 g/liter hLF vs. 2-3 g/liter in the breastmilk. The content of LF in cow colostrum is ~0.8 g/liter and only 0.03-0.49 g/liter in cow milk. Such high concentration of hLF in the colostrum protects infants from bacterial infections and inflammation. If the hLF content in mother’s milk is low, it is recommended to add hLF to the infant’s diet, especially in the case of prematurely born infants. It is also important for the formula-fed babies, when the breastfeeding is impossible because of the mother’s illness or treatment with drugs that can negatively affect the infant’s health [10]. Pasteurization of breastmilk used for the feeding of premature and sick infants, as well as the processing of milk and milk products, can disturb the structure and, therefore, the function of LF and its properties [5]. In view of this, the need for efficient and low-cost methods for the quantitative assay of hLF in breastmilk and milk products is obvious.
Multiple methods have been developed for the LF detection and quantification in milk (mostly, cow’s milk), including enzyme-linked immunoassay (ELISA) [11], surface plasmon resonance with immunosensors [12], capillary electrophoresis [13], ultrahigh-performance liquid chromatography combined with tandem mass spectrometry (UHPLC-MS/MS) [14], and reversed-phase HPLC (RP-HPLC) [15, 16]. hLF can also be detected by the homogeneous immunochemical method using excitation transfer and phase-resolved fluorometry [17]. The most common method for the hLF detection is ELISA; several commercial ELISA kits are available on the market now, for example, see https://www.hycultbiotech.com/hk329-02. This kit for sandwich ELISA allows to quantify hLF in human biological fluids after their dilution to the assay working concentrations with a high sensitivity (0.4-100 ng/ml) within 3.5 h. The UHPLC-MS/MS method is especially sensitive and can be used for assaying LF in the processed milk and milk products. However, this method was developed using peptides from bovine LF hydrolysate as standards, which means that UHPLC-MS/MS cannot distinguish between the native and denatured LF. The content of native LF can be determined by RP-HPLC, but the results of this assay differ from the results obtained by HPLC. The sensitivity of RP-HPLC might be insufficient for the LF detection in commercially sold milk and milk products with a low LF content. The problem of LF enrichment for the following quantification by HPLC was solved by using the HITRAPTM Heparin HP [18] and aptamer [19] columns, but these methods require time-consuming sample preparation, high-cost equipment, and highly qualified personnel.
Based on the analysis of published data and our own experience, we suggested the fluorescence polarization immunoassay (FPIA) with fluorescently labeled single-domain antibodies (nanobodies) for the hLF detection and quantification in human breastmilk. FPIA does not involve elaborate procedures, e.g., separation of free and bound protein, time-consuming sample preprocessing, and labor-intensive procedures typical for the food quality assays and various clinical and biomedical tests [20]. FPIA can be performed in less than 5 min, and its calibration curve is stable and does not have to be repeated for each assay. We believe that FPIA can be very useful for rapid and precise quantification of hLF in breastmilk. Most FPIA protocols are competitive assays, in which the analyzed compound and fluorescently labeled analyte (tracer) compete for the binding with the antibody. Originally, FPIA was developed for the detection of low-molecular-weight compounds, such as antibiotics, mycotoxins, and pesticides [20, 21]. Theoretically, the value of fluorescence polarization (FP) signal can vary from 0 to 0.5; the calculations typically use the fluorescence millipolarization (mP) value. FP depends on the hydrodynamic radius (relative size) of the fluorescent compound. The binding of the low-molecular-weight (less than 5 kDa) tracer to the protein or antibody of a significantly larger size noticeably increases the hydrodynamic radius and results in strong changes in the degree of FP (ΔmP). If the analyzed compound is large (e.g., average-size protein of ~60 kDa), ΔmP is small, which hinders the use of FPIA because to the small difference in the hydrodynamic radii of the free analyte and analyte bound to an antibody or a protein.
The problem of quantification of large molecules can be solved by using the noncompetitive FPIA. Thus, the noncompetitive FPIA with fluorescently labeled Fab fragments was used in the development of the C-reactive protein quantitative assay [22]. The possibility of FPIA application for the detection of viral particles was demonstrated in [23-25]. However, the use of these variants of noncompetitive FPIA is limited by the high background and low sensitivity of the method. Beside the problem of small changes in the FP signal upon the binding to high-molecular-weight molecules, another drawback of FIPA is nonspecific adsorption. Therefore, further FIPA development should involve the search for the low-molecular-weight molecules for antigen binding.
One of the promising approaches in the development of protein-specific FPIA is the use of miniature camelid single-domain antibodies (nanobodies) with a molecular weight of 12-15 kDa (2 × 4 nm). Nanobodies (nanoantibodies, VHH) are recombinant proteins corresponding to the variable antigen-binding domain of camelid antibodies that normally consist of a homodimer of truncated heavy chains and lack the light chains. These antibodies have been discovered in the representatives of the Camelidae family and cartilaginous fishes [26, 27]. The nanobodies are very stable within a broad range of temperatures and pH values and can rapidly renature with a full restoration of their activity. They bind to specific antigen epitopes (e.g., enzyme active sites, small pockets on the protein surface) that are not typically recognized by the classical antibodies. The Tillib laboratory has worked on the production and use of nanobodies [28-31], in particular, highly specific nanobodies against hLF [29], for the last two decades. In recent years, the use of nanobodies in immuno-biotechnology has become increasingly common; the nanobodies are currently viewed as auxiliary tools to the traditional antibodies and their derivatives. Due to its small molecular mass, fluorescently labeled nanobody can be used to recognize protein targets of a considerably larger size in noncompetitive FPIA.
The goal of this work was to demonstrate the principal possibility of using single-domain antibodies in the noncompetitive FPIA for the antigen detection in biological fluids, namely, for hLF quantification in human breastmilk using previously developed anti-hLF nanobodies.
MATERIALS AND METHODS
Reagents. FITC (fluorescein 5-isothiocyanate isomer 1; Sigma-Aldrich, USA), clones producing camelid anti-hLF5 (20.1 kDa) and anti-hLF16 (20 kDa) nanobodies [29], hLF (80 kDa), and four samples of human breastmilk (nos. 1-4) were kindly provided by E. R. Sadchikova (Institute of Gene Biology, Russian Academy of Science, Moscow). Bovine serum albumin (BSA), human transferrin (hTF, a mixture of apo- and holoprotein), human serum albumin (HSA), human lysozyme (LYZ), and casein (Cas) were from Sigma-Aldrich.
Labeling of anti-hLF nanobodies with FITC. Using earlier obtained bacterial producers [29], anti-hLF5 and anti-hLF16 nanobodies specifically recognizing hLF (but not goat LF) were purified from the bacterial periplasm by the metal-chelating chromatography and characterized by chromatography, electrophoresis, and enzyme immunoassay (ELISA) as described in [29]. The nanobodies were conjugated with FITC, purified as described in [32], and characterized spectrophotometrically to determine the protein content and the extent of modification of protein amino groups. The concentration of labeled FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies was determined as 0.68 and 0.71 mg/ml, respectively, using the corresponding molar extinction coefficients ε280 = 29780 and 28260 M–1 cm–1 (molar extinction coefficient of FITC is 73,000 M–1 cm–1) [33].
Enzyme-linked immunoassay (ELISA) of hLF was performed with an hLF ELISA kit (XEMA, Russia) as recommended by the manufacturer. The optical density in the plate wells was measured at 450 nm with a Bio-Rad 680 microplate reader (Bio-Rad, USA).
Noncompetitive FPIA was performed using a portable Sentry-200 fluorimeter (Ellie, USA) at λex = 495 and λem = 530 nm. The working solutions of the FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies were prepared so that the florescence of the solutions was approx. 200,000 AU. The concentration of prepared hLF solution was checked spectrophotometrically in several dilutions using ε280 = 8.85·104 M−1 cm−1 [34]. The working nanobody solution (950 µl) was mixed with 50 µl of hLF solution (0, 0.01, 0.1, 1, 3, 10, 20, 40, 100, and 1000 µg/ml) and incubated for 5 min, followed by measuring fluorescence polarization (mP) and fluorescence intensity. Each experiment was conducted in two biological and three technical repeats. The cross-reactivity of the used nanobodies was evaluated by measuring FP after adding 50 µl of Cas, BSA, HAS, or hTF (0, 1, 10, 100, and 1000 µg/ml) to 950 µl of the FITC-anti-hLF nanobody working solution.
hLF quantification in milk. hLF was added to goat milk to the final concentration of 0.5 and 1 mg/ml. The milk was centrifuged for 15 min at 4000g to remove fat, diluted 20, 40, and 80 times with 10 mM phosphate buffer (pH 7.4) containing 150 mM NaCl (PBS) and used in noncompetitive FPIA. To validate the results of FPIA, all milk samples were analyzed by ELISA using the hLF ELISA kit (at present, this kit undergoes the procedure of state registration and is produced as a pilot batch).
RESULTS AND DISCUSSION
Here we used earlier obtained single-domain camelid anti-hLF5 and anti-hLF16 nanobodies developed against two different hLF epitopes [29]. Each nanobody bound almost all hLF in milk samples [29], thus suggesting that the recognized epitopes remained accessible the interaction with the nanobodies in hLF monomers and oligomers. According to the preliminary data, the anti-hLF5 nanobodies lost the ability to bind the peptides obtained by the partial hLF hydrolysis with pepsin, while the anti-hLF16 nanobody bound a small 35-kDa peptide in the hydrolysate. Therefore, it is possible that at least one nanobody (anti-hLF16) can bind not only the full-size protein, but its peptide as well (or denatured protein containing this peptide). We found no other differences between the anti-hLF5 and anti-hLF16 nanobodies when they were individually used as the tracers. We assumed that the influence of possible partial denaturation of hLF on the hLF detection with these nanobodies was negligent. Both nanobodies were highly selective toward hLF and did not bind goat LF [29].
The nanobodies used in this work were adapted, i.e., contained at the C-terminus a long linker sequence (28 amino acids of the long variant of the noncanonical camelid antibody hinge region) followed by the HA-tag (YPYDVPDYA) and His6-tag. The linear part of the linker contained four easily accessible lysine residues that can be used for the conjugation of other molecules.
The anti-hLF5 and anti-hLF16 nanobodies were labeled with FITC The nanobody/FITC ratio was determined from the spectra of the labeled nanobodies in PBS (pH 7.4) using equation (1):
where A492 is the conjugate absorbance at the FITC maximum absorption wavelength (492 nm); A280 is the sample absorbance at the nanobody maximum absorption wavelength (280 nm); εanti-hLF is the molar extinction coefficient at 280 nm; εFITC is the FITC molar extinction coefficient at 492 nm (73,000 M−1 cm−1); and C is the correction factor for FITC at 280 nm (C = 0.35). The FITC/nanobody ratio was approximately 1 : 1 and was found to be the optimal, as further increase in the label/protein ratio did not lead to the increase in the fluorescence signal.
The development of efficient diagnostics system requires full characterization of the specificity and affinity of the used antibodies [35]. For the analytical method based on FPIA, it is important to determine the constant for the antigen-antibody binding in solution, which might significantly differ from the binding constant for the surface-immobilized antibody. Hence, we evaluated the affinities of the FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies toward hLF to estimate the possibility of using them for the LF quantification in milk by FPIA.
To develop the FPIA protocol, we used the working solutions of FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies with the concentrations that provided the optimal fluorescence signal and, therefore, ensured stable FP value. The optimal signal/noise ratio was observed within the concentration interval of 2.5-5 nM. The FP value for the FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies was 125 ± 1 mP.
The kinetics of the FITC-anti-hLF5 and FITC-anti-hLF16 binding with hLF was studied in a broad protein concentration range. The standard solutions of hLF (0.125 pM-12 µM) were prepared in PBS. To measure the FP signal, 50 µl of the standard hLF solution was mixed with 950 µl of the FITC-anti-hLF5 or FITC-anti-hLF16 working solution. The tube was placed in a portable Sentry-200 fluorimeter, and the kinetics of hLF (final concentration, 6 pM-625 nM) binding by the nanobodies was recorded for 1 h. Figure 1 shows the time dependence of the FP signal at the hLF concentration of 10 nM. The observed rapid increase in the signal intensity and establishment of equilibrium within 1-2 min at room temperature indicated a high-affinity binding of hLF by the nanobodies. In further experiments, the FP signal was measured after 5 min of the reaction mixture incubation. It is known that LF can form associates; the extent of its oligomerization depends on the concentration of hLF and salts (KCl, NaCl), as well as on the time of protein storage in solution [36]. To exclude the effect of oligomerization, we studied the stability of the FP signal in a wide range of hLF concentration (6 pM-625 nM). At any concentration tested, the FP signal reached its maximum within 2 min and then remained unchanged for 1 h. This suggests that no hLF aggregation took place in the studied solution (otherwise, the FP signal would have increased). We also studied the kinetics of hLF interaction with FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies after preincubation of hLF standard solutions (125 pM-12 µM) for different periods of time before the addition of the nanobodies. However, even after hLF preincubation at room temperature for 8 h, the FP signal remained the same (data not shown), thus indicating the absence of the influence of possible hLF oligomerization on the assay results.

Kinetics of hLF (10 nM) binding with FITC-anti-hLF5 (1) and FITC-anti-hLF16 (2) nanobodies (concentration of both nanobodies, 5 nM) in PBS (pH 7.4) at 25°C. The measurements were performed in triplicate; the data are shown as mean ±SD.
FPIA is a common method for characterization of the protein–ligand interactions [34, 36]. The equation (2) describes the binding between the tracer (FITC-anti-LF) and hLF:
If the concentration of FITC-anti-LF is much less that the hLF concentration, FP can be expressed by the equation (3) [37]:
where mPmin is FP of the free FITC-anti-LF5, mPmax is FP of the hLF*FITC-anti-LF5 complex; [hLF] is the hLF concentration; and KD is the dissociation constant of the hLF*FITC-anti-LF complex.
Figure 2 shows the binding curves for the interaction of FITC-anti-LF5 and FITC-anti-LF16 tracers (2.5 nM) with different hLF concentrations. As seen from the figure, the FP value increased with the increase in the hLF concentration and reached its maximum upon full tracer binding to hLF. The change in FP (ΔmP) was 30-35 AU due to the use of single-domain low-molecular-weight (~20 kDa) nanobodies. When we used three independently produced batches of labeled nanobodies, the variations in the FP value did not exceed 10%.

The dependence of FP signal on the hLF concentration in the presence of 2.5 nM FITC-anti-LF5 (1) and FITC-anti-LF16 (2) nanobodies at pH 7.4 at 25°C. The measurements were performed in triplicate; the data are shown as mean ±SD.
The dissociation constants (KD) for the hLF complexes with the FITC-anti-LF5 and FITC-anti-LF16 nanobodies were determined at the constant tracer concentration (2.5 nM) that ensured the optimal signal/noise ratio and stable FP values. This concentration was below the expected KD and below increasing hLF concentrations that exceeded the expected KD value. The hLF concentration was varied from 0 to 50 nM. The FP signals (mP) were approximated by the equation (2). The calculated equilibrium KD for FITC-anti-LF5 and FITC-anti-LF16 was 3.2 ± 0.3 and 4.9 ± 0.4 nM, respectively, i.e., both nanobodies had sufficiently low dissociation constants indicating their high-affinity binding to hLF.
Proteins present in body fluids, as well as proteins often used in buffers, which are structurally similar to LF, might affect the FP signal and analysis accuracy. Hence, we investigated the influence of several nonspecific proteins (BSA, HSA, LYZ, Cas, hTF) potentially capable of cross-reacting with the nanobodies in the noncompetitive FPIA with the FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies (Fig. 3). The tested proteins (1-1000 µg/ml) were added to the nanobody working solutions, and the FP signal was measured after 5 min of incubation. At all the concentrations used, the nonspecific proteins did not affect the FP signal, indicating the high selectivity of the FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies toward hLF (Fig. 3).

The binding of nonspecific proteins with FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies at pH 7.4 at 25°C. The concentration of hLF was 40 µg/ml; the concentration of other proteins was 1000 µg/ml. The measurements were performed in triplicate; the data are shown as mean ±SD.
The concentration of hLF in milk was determined using the recovery test. Neither anti-hLF5, nor anti-hLF16 nanobody reacted with goat LF, hence, we used natural goat milk to which known amounts of hLF were added.
Analysis of substances in milk is complicated by the presence of multiple constituents that can affect the analytical signal (matrix effect). The most common approach to reducing the matrix effect is dilution aimed to decrease the concentration of potentially interfering compounds. The milk was centrifuged to remove fats and then used for preparing a series of two-fold dilutions in PBS. The optimal dilution for the use in analysis was determined by adding 50 µl of diluted milk sample to the working solutions of FITC-anti-hLF5 and FITC-anti-hLF16 nanobodies and measuring the FP signal after 5 min of incubation. Milk samples diluted 8 to 10 times (final dilution in the analyzed solution, 160 to 200 times) produced the FP value similar to that of the FITC-anti-hLF5 and FITC-anti-hLF16 solutions mixed with 50 µl of buffer (data not shown). Therefore, already at the 160-fold dilution, the matrix effect was eliminated, whereas the high content of LF in milk (0.5-1 mg/ml) allowed its quantification by FPIA in the milk diluted more than 200 times.
We chose the FITC-anti-hLF5 nanobody for the use in FPIA, because its affinity to hLF was higher than the affinity of FITC-anti-hLF16. Figure 4a shows the dependence of the FP on the hLF concentration in the noncompetitive FPIA with the FITC-anti-hLF5 nanobody and the linear fragment of this curve (Fig. 4b). The detection limit was determined as 2.1 ± 0.2 µg/ml (mP0 + 3 SD); the linear range for the hLF quantification was 3-50 µg/ml.

Normalized dependence of the FP signal on the hLF concentration in the noncompetitive FPIA with FITC-anti-hLF5 nanobodies (a) and its linear fragment (b) at pH 7.4 and 25°C. The measurements were performed in triplicate; the data are shown as mean ±SD.
To verify the reproducibility of the method used for the nanobody labeling, the FITC-anti-hLF5 conjugate was synthesized and purified as described in [32] in three independent experiments. The difference in the calibration curves for the three produced batches of nanobodies and analytical characteristics of these nanobodies differed by less than 10%.
Since the anti-hLF5 nanobody was highly selective toward hLF and did not bind goat LF [29], we used goat milk to confirm the possibility of hLF detection in milk. For this, hLF was added to the goat milk (final concentration, 0.5 and 1 mg/ml), the samples were diluted 20, 40, 80 times (as mentioned above, the matrix effect disappeared already after 10-fold milk dilution); and each dilution was tested for the FP value and hLF concentration (Table 1).
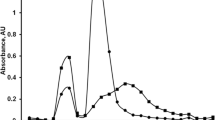
The same samples were assayed for the hLF content by ELISA. Figure 5 shows the dependence of the absorbance (A) on the hLF concentration for the standard hLF solutions. Milk samples for ELISA were diluted 200, 400, and 800 times. The results of measurements are shown in Table 1. No increase in the analytical signal was found by FPIA or ELISA in the goat milk containing no hLF, which indicates a highly specific binding between hLF and used nanobody.

Calibration curve for hLF ELISA. The measurements were performed in duplicate; the data are shown as mean ±SD.
As seen from Figs. 4b and 5, the detection limit for ELISA was lower that for FPIA (0.3 and 2.1 µg/ml, respectively). However, the content of LF in milk is 1 mg/ml or more and dilution of milk samples allows to eliminate the matrix effect that might influence the accuracy of the assay. FPIA is much less laborious and time-consuming method that reduced the time of assay from 3.5 h to 5 min. Moreover, the calibration curve for the used nanobody was stable, which means it does not have to be constructed each time the samples are analyzed.
To verify the possibility of using the developed FPIA for the hLF analysis in human breastmilk, we tested four breastmilk samples (nos. 1-4). In parallel, the content of hLF in the same samples was determined by traditional ELISA (hLF ELISA kit from XEMA).
Table 2 shows that the hLF concentrations determined by FPIA and ELISA in the breastmilk samples diluted 100 and 150 times were close to the expected ones.
In order to obtain reliable results, breastmilk had to be diluted more than 1000 times when tested by ELISA, since this assay is more sensitive than FPIA; however, it is also more laborious and time-consuming.
Table 2 shows that the concentration of hLF in breastmilk determined by FPIA correlated well with the data obtained by the more traditional ELISA.
To verify the use of FPIA for the hLF analysis breastmilk, we tested only 4 breastmilk samples that have been used in the studies by our colleagues before and have been already diluted and stored frozen. Here, we demonstrated that FPIA can be used for reliable hLF quantification in real biological specimens without significant modifications of the developed method.
In conclusion, we demonstrated a principal possibility of using single-domain antibodies as recognizing agents for assaying target antigens (LF) in biological fluids (milk) by noncompetitive FPIA. Compared to the traditional ELISA, FPIA is less time consuming (5 min); the antigen-antibody binding takes place in a homogeneous medium and requires no complicated manipulations. FPIA demonstrated a high sensitivity and selectivity toward hLF in human breastmilk and samples of goat milk with added hLF; the obtained results correlated with the data obtained by ELISA. Fluorescently labeled single-domain anti-LF5 and anti-LF16 nanobodies are promising agents for developing a test system for hLF quantification in human breastmilk and biological fluids by FPIA.
Abbreviations
- FITC:
-
fluorescein 5-isothiocyanate isomer 1
- FP:
-
fluorescence polarization
- FPIA:
-
fluorescence polarization immunoassay
- hLF:
-
human lactoferrin
- LF:
-
lactoferrin
References
González-Chávez, S. A., Arévalo-Gallegos, S., and Rascón-Cruz, Q. (2009) Lactoferrin: Structure, function and applications, Int J. Antimicrob. Agents, 33, 301.e1-301.e8, https://doi.org/10.1016/j.ijantimicag.2008.07.020.
Karav, S., German, J. B., Rouquié, C., Le Parc, A., and Barile, D. (2017) Studying lactoferrin N-glycosylation, Int. J. Mol. Sci., 18, 870, https://doi.org/10.3390/ijms18040870.
Rosa, L., Cutone, A., Lepanto, M. S., Paesano, R., and Valenti, P. (2017) Lactoferrin: a natural glycoprotein involved in iron and inflammatory homeostasis, J. Mol. Sci., 18, 1985, https://doi.org/10.3390/ijms18091985.
Adlerova, L., Bartoskova, A., and Faldyna, M. (2008) Lactoferrin: a review, Vet. Med., 53, 457-468, https://doi.org/10.17221/1978-VETMED.
Murphy, M. E., Kariwa, H., Mizutani, T., Tanabe, H., Yoshimatsu, K., Arikawa, J., and Takashima, I. (2001) Characterization of in vitro and in vivo antiviral activity of lactoferrin and ribavirin upon hantavirus, J. Vet. Med. Sci., 63, 637-645, https://doi.org/10.1292/jvms.63.637.
Legrand, D., and Mazurier, J. (2010) A critical review of the roles of host lactoferrin in immunity, Biometals, 23, 365-376, https://doi.org/10.1007/s10534-010-9297-1.
Baker, E. N. (2004) Lactoferrin and iron: structural and dynamic aspects of binding and release, BioMetals, 17, 209-216, https://doi.org/10.1023/B:BIOM.0000027694.
Nagasako, Y., Saito, H., Tamura, Y., Shimamura, S., and Tomita, M. (1993) Iron-binding properties of bovine lactoferrin in iron-rich solution, J. Dairy Sci., 76, 1876-1881, https://doi.org/10.3168/jds.S0022-0302(93)77520-7.
Fox, P. F., Uniacke-Lowe, T., McSweeney, P., and O’Mahony, J. (2015) Biologically Active Compounds in Milk, in: Dairy Chemistry and Biochemistry, Springer. Cham, https://doi.org/10.1007/978-3-319-14892-2_11.
Sadchikov, P. E., Goldman, I. L., Namazova-Baranova, L. S., Yatsyk, G. V., Borovik, T. E., Chernousov, A. D., Romanchenko, A. I., Sadchikova, E. R., Lukoyanova, O. L., Zvonkova, N. G., and Belyaeva, I. A. (2016) Lactoferrin in the problem of ant-infection protection of infants of the first year of life [in Russian], Pediatr. Farmakol., 13, 607-613, https://doi.org/10.15690/pf.v13i6.1677.
Chen, P.-W., and Mao, F.C. (2004) Detection of lactoferrin in bovine and goat milk by enzyme-linked immunosorbent assay, J. Food Drug Anal., 12, 9, https://doi.org/10.38212/2224-6614.2653.
Tomassetti, M., Martini, E., Campanella, L., Favero, G., Sanzò, G., and Mazzei, F. (2013) Lactoferrin determination using flow or batch immunosensor surface plasmon resonance: Comparison with amperometric and screen-printed immunosensor methods, Sens. Actuat. B Chem., 179, 215-225, https://doi.org/10.1016/j.snb.2012.09.096.
Chen, H., Wang, Z., Fan, F., Shi, P., Xu, X., Du, M., and Wang, C. (2021) Analysis method of lactoferrin based on uncoated capillary electrophoresis, eFood, 2, 147-153, https://doi.org/10.2991/efood.k.210720.001.
Zhang, J., Lai, S., Cai, Z., Chen, Q., Huang, B., and Ren, Y. (2014) Determination of bovine lactoferrin in dairy products by ultra-high performance liquid chromatography-tandem mass spectrometry based on tryptic signature peptides employing an isotope-labeled winged peptide as internal standard, Anal. Chim. Acta, 829, 33-39, https://doi.org/10.1016/j.aca.2014.04.025.
Palmano, K. P., and Elgar, D. F. (2002) Detection and quantitation of lactoferrin in bovine whey samples by reversed-phase high-performance liquid chromatography on polystyrene-divinylbenzene, J. Chromatogr. A, 947, 307-311, https://doi.org/10.1016/s0021-9673(01)01563-1.
Tsakali, E., Chatzilazarou, A., Houhoula, D., Koulouris, S., Tsaknis, J., and Van Impe, J. (2019) A rapid HPLC method for the determination of lactoferrin in milk of various species, J. Dairy Res., 86, 238-241, https://doi.org/10.1017/s0022029919000189.
Nithipatikom, K., and McGown, L. B. (1987) Homogeneous immunochemical technique for determination of human lactoferrin using excitation transfer and phase-resolved fluorometry, Anal. Chem., 59, 423-427, https://doi.org/10.1021/ac00130a010.
Chen, M. X., Wen, F., Zhang, Y. D., Li, P., Zheng, N., and Wang, J. Q. (2019) Determination of native lactoferrin in milk by HPLC on HiTrap Heparin HP column, Food Anal. Method., 12, 2518-2526, https://doi.org/10.1007/s12161-019-01572-x.
Wang, N., Jiang, X., Xu, X., Liu, Y., Liu, L., Lu, A., Lu, J., and Luan, Y. (2021) An aptamer affinity column for purification and enrichment of lactoferrin in milk, J. Chromatography B, 1178, 122724, https://doi.org/10.1016/j.jchromb.2021.122724.
Hendrickson, O. D., Taranova, N. A., Zherdev, A.V., Dzantiev, B. B., and Eremin, S. A. (2020) Fluorescence polarization-based bioassays: new horizons, Sensors, 20, 7132, https://doi.org/10.3390/s20247132.
Zhang, H., Yang, S., De Ruyck, K., Beloglazova, N., Eremin, S. A., De Saeger, S., Zhang, S., Shen, Ji., and Wang, Z. (2019) Fluorescence polarization assays for chemical contaminants in food and environmental analyses, Trends Anal. Chem., 114, 293-313, https://doi.org/10.1016/j.trac.2019.03.013.
Nishiyama, K., Fukuyama, M., Maeki, M., Ishida, A., Tani, H., Hibara, A., Tokeshi, M. (2021) One-step non-competitive fluorescence polarization immunoassay based on a Fab fragment for C-reactive protein quantification, Sensors Actuators B Chem., 326, 128160, https://doi.org/10.1016/j.snb.2020.128982.
Nishiyama, K., Takeda, Y., Takahash, K., Fukuyama, M., Maeki, M., Ishida, A., Tani, H., Shigemura, K., Hibara, A., Ogawa, H., and Tokeshi, M. (2021) Non-competitive fluorescence polarization immunoassay for detection of H5 avian influenza virus using a portable analyzer, Anal. Bioanal. Chem., 413, 4619-4623, https://doi.org/10.1007/s00216-021-03193-y.
Nishiyama, K., Takahashi, K., Fukuyama, M., Kasuya, M., Imai, A., Usukura, T., Maishi, N., Maeki, M., Ishida, A., Tani, H., Hida, K., Shigemura, K., Hibara, A., and Tokeshi, M. (2021) Facile and rapid detection of SARS-CoV-2 antibody based on a noncompetitive fluorescence polarization immunoassay in human serum samples, Biosens. Bioelectron., 190, 113414, https://doi.org/10.1016/j.bios.2021.113414.
Takeda, Y., Yonezawa, Y., Asake, S., Ogawa, H., and Imai, K. (2020) A fluorescence polarization immunoassay for the rapid detection of antibody against influenza A virus in chicken and goat sera, J. Vet. Diagn. Invest., 32, 887-891, https://doi.org/10.1177/1040638720960046.
Hamers-Casterman, C., Atarhouch, T., Muyldermans, S., Robinson, G., Hamers, C., Songa, E.B, Bendahman, N., and Hamers, R. (1993) Naturally occurring antibodies devoid of light chains, Nature, 363, 446-448, https://doi.org/10.1038/363446a0.
Flajnik, M. F., and Kasahara, M. (2010) Origin and evolution of the adaptive immune system: genetic events and selective pressures, Nat. Rev. Genet., 11, 47-59, https://doi.org/10.1038/nrg2703.
Tillib, S. V., Ivanova, T. I., and Vasilev, L. A. (2010) Fingerprint-like analysis of “nanoantibody”. Selection by phage display using two helper phage variants, Acta Naturae, 2, 85-93, https://doi.org/10.32607/20758251-2010-2-3-85-93.
Tillib, S. V., Privezentseva, M. E., Ivanova, T. I., Vasilev, L. F., Efimov, G. A., Gurskiy, Ya. G., Georgiev, G. P., Goldman, I. L., and Sadchikova, E. R. (2014) Single-domain antibody-based ligands for immunoaffinity separation of recombinant human lactoferrin from the goat lactoferrin of transgenic goat milk, J. Chromatogr. B, 949-950, 48-57, https://doi.org/10.1016/j.jchromb.2013.12.034.
Goryainova, O. S., Ivanova, T. I., Rutovskaya, M. V., and Tillib, S. V. (2017) A Method for the parallel and sequential generation of single-domain antibodies for the proteomic analysis of human blood plasma [in Russian], Mol. Biol. (Mosk.), 51, 985-996, https://doi.org/10.7868/S0026898417060106.
Tillib, S. V. (2020) Prospective applications of single-domain antibodies in biomedicine [in Russian], Mol. Biol. (Mosk.), 54, 362-373, https://doi.org/10.31857/S0026898420030167.
Schreiber, A. B., and Yfimovich, J. (1983) Quantitative fluorometric assay for detection and characterization of Fc receptors, Meth. Enzymol., 93, 147-155, https://doi.org/10.1016/s0076-6879(83)93039-2.
Pace, C. N.Vajdos, F., Fee, L., Grimsley, G., and Gray, T. (1995) How to measure and predict the molar absorption coefficient of a protein, Protein Sci., 11, 2411-2423, https://doi.org/10.1002/pro.5560041120.
Chung, T., and Raymond, K. (1993) Lactoferrin: the role of conformational changes in its iron binding and release, J. Am. Chem. Soc., 115, 6765-6768, https://doi.org/10.1021/ja00068a037.
Mukhametova, L. I., Krylov, V. B., Soloviev, A. S., Yashchunsky, D. V., Matveev, A. L., Tikunova, N. V., Eremin, S. A., and Nifantyev, N. E. (2021) Characterization of affinity of the anti-β-(1→3)-d-glucan monoclonal antibody 3G11 by the fluorescence polarization immunoassay, Izv. Akad. Nauk, Ser. Khim., 5, 975-980.
Nevinskii, A. G., Soboleva, S. E., Tuzikov, F. V., Buneva, V. N., and Nevinsky, G. A. (2009) DNA, oligosaccharides, and mononucleotides stimulate oligomerization of human lactoferrin, J. Mol. Recognit., 22, 330-342, https://doi.org/10.1002/jmr.952.
Zhang, S., Chen, L., Kumar, S., Wu, L., Lawrence, D. S., and Zhang, Z.-Y. (2007) An affinity-based fluorescence polarization assay for protein tyrosine phosphatases, Methods, 42, 261-267, https://doi.org/10.1016/j.ymeth.2007.02.008.
Acknowledgments
The authors thank E. R. Sadchikova (Institute of Gene Biology) for kindly providing purified hLF and human breastmilk samples.
Funding
This work was supported by the Russian Science Foundation (project no. 20-14-00305).
Author information
Authors and Affiliations
Contributions
S. A. Eremin, L. I. Mukhametova, and S. V. Tillib developed the concept, supervised the study, and discussed the results; L. I. Mukhametova and D. A. Arutyunyan performed FPIA experiments; O. S. Goryainova, T. I. Ivanova, and S. V. Tillib produced and labeled nanobodies; L. I. Mukhametova wrote the manuscript; S. A. Eremin and S. V. Tillib edited the text.
Corresponding authors
Ethics declarations
The authors declare no conflict of interest. This article contains no description of studies involving humans or animals performed by any of the authors.
Rights and permissions
Open access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution, and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mukhametova, L.I., Eremin, S.A., Arutyunyan, D.A. et al. Fluorescence Polarization Immunoassay of Human Lactoferrin in Milk Using Small Single-Domain Antibodies. Biochemistry Moscow 87, 1679–1688 (2022). https://doi.org/10.1134/S0006297922120227
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0006297922120227