
Abstract
The traditional application of the sclerotium of Lignosus rhinocerotis (tiger’s milk mushroom) by the indigenous folks as tonic and remedy to treat a variety of ailments has been documented in Malaysia. Indigenous communities claimed to have consumed the decoction to boost their alertness during hunting. Mental alertness is believed to be related to neuronal health and neuroactivity. In the present study, the cell viability and neuritogenic effects of L. rhinocerotis sclerotium hot aqueous and ethanolic extracts and crude polysaccharides on rat pheochromocytoma (PC-12) cells were studied. Interestingly, the hot aqueous extract exhibited neuritogenic activity comparable to NGF in PC-12 cells. However, the extracts and crude polysaccharides stimulated neuritogenesis without stimulating the production of NGF in PC-12 cells. The involvements of the TrkA receptor and MEK/ERK1/2 pathway in hot aqueous extract-stimulated neuritogenesis were examined by Trk (K252a) and MEK/ERK1/2 (U0126 and PD98059) inhibitors. There was no significant difference in protein expression in NGF- and hot aqueous extract-treated cells for both total and phosphorylated p44/42 MAPK. The neuritogenic activity in PC-12 cells stimulated by hot aqueous and ethanolic extracts and crude polysaccharides of L. rhinocerotis sclerotium mimicking NGF activity via the MEK/ERK1/2 signaling pathway is reported for the first time.
Similar content being viewed by others


Introduction
Medicinal mushrooms and their extracts have a long and rich history of use in traditional oriental medicines as mycomedicines1. Increasingly, many are being regarded as functional foods and nutraceuticals. The neuroactivities of medicinal mushrooms are under intense study and research. Phan et al. (2014)2 reviewed a number of studies of medicinal mushrooms, revealing the promises of medicinal mushrooms as useful therapeutic agents in the management and/or treatment of neurodegenerative disorders.
In Malaysia, Lignosus rhinocerotis (Cooke) Ryvarden is also known as ‘tiger’s milk mushroom’ in English or ‘cendawan susu rimau’ in Malay. It is considered as a unique “National Treasure” that can only be found in a small geographic region in Southern China, Thailand, Malaysia, Indonesia, Philippines, Papua New Guinea, New Zealand and Australia3. In Malaysia, L. rhinocerotis is the most popular medicinal mushroom used by the indigenous communities of Peninsular Malaysia4. The benefits of its underground tuber or sclerotium (where most of the nutritional and medicinal components are deposited) compared to its basidiocarp are well documented (Table 1). According to the ethnopharmacological reports, the sclerotium is sliced, boiled and drunk as an effective tonic for overall wellness and to treat several ailments including fever, cough, asthma, chronic hepatitis, gastric ulcer, cancer and food poisoning4,5,6,7 (Table 1). Indigenous communities also drank this decoction to increase their alertness during hunting (tacit knowledge). Moreover, Tan et al. (2012)3 documented that the consumption of tiger’s milk mushroom improved stamina and alertness in healthy people. Increment of the mental alertness is believed to be related to the neuroactivity and neuronal communication network in brain. Based on the traditional practice as the basic for the scientific study, we explored L. rhinocerotis, the Malaysia’s treasure mushroom, notably for the neuritogenic activity in vitro.
Neuritogenesis is a dynamic phenomenon associated with neuronal differentiation, where the neurons generate and extend their neurites to form a functional network8. Neuritogenic activity is important for the maintenance and regeneration of the neuronal communications network and has become one of the focuses of study in the search for preventive and therapeutic agents for neurodegenerative disorders. The neurotrophic factors (neurotrophins), exemplified by nerve growth factor (NGF), play distinctive roles in promoting neuronal survival9, proliferation, development, neuritogenesis and maintaining the neurons functions10. Nerve growth factor was identified as a potential therapeutic agent for the treatment of neurodegenerative disorders11. A decrease in NGF in the brain is believed to be the main cause of neuronal dysfunction and neurodegenerative disorders, remarkably Alzheimer’s disease12. Further, Capsoni et al. (2000)13 presented the evidence that a decrease in NGF in mice’s brain led to neurodegeneration and Alzheimer’s-like symptoms. Learning ability and memory were improved in aged anti-nerve growth factor transgenic mice after administration of NGF13. However, the neuroactivity of NGF is restricted due to its large molecular polypeptide structure. It is unstable and unable to cross the blood-brain barrier14. Smaller molecules that mimic and/or enhance the NGF activity have become the core focus in the search for preventive and therapeutic agents for neurodegenerative disorders15. Among the natural sources explored for NGF mimics, medicinal mushrooms have shown huge potential2.
Nerve growth factor was found to activate the mitogen-activated protein kinase (MAPK) signaling pathway that mediates the phosphorylation including the mitogen-activated protein kinase kinase/extracellular signal-regulated kinase (MEK/ERK) pathway16. The MAPK/ERK pathway is a chain of proteins in the cell that participate by Ras, Raf, MEK1/2 and ERK1/2 proteins. The MAPK/ERK cascade is a signal transduction pathway that involves a large variety of processes such as differentiation, proliferation, apoptosis, cell cycle progression, cell migration and metabolism17. The MEK/ERK1/2 signalling cascade is indeed crucial in both in vitro18 and in vivo for neuronal cell survival, axonal regeneration19 and neuritogenesis in extract-treated mouse dorsal root ganglia neurons20.
The MEK/ERK1/2 signaling pathway is believed to be the major cascade for NGF to stimulate neuritogenesis in PC-12 cells18. The PC-12 cell line is widely used as an in vitro model system to investigate the neuritogenic activity of NGF and NGF mimics and NGF responsive signaling pathways15,18,21,22. Nerve growth factor stimulates differentiation of PC-12 cells into a sympathetic neuronal-like phenotype and extend axon-like outgrowth23. PC-12 cells treated with NGF have been found to stop proliferating and differentiate into neuronal-like cells with neurite outgrowth23. In vitro neuritogenesis stimulated by hot aqueous extract of Ganoderma sp.24, Pleurotus giganteus (Berk.) Karunarathna & K. D. Hyde25 and Ganoderma neo-japonicum Imazeki26 in PC-12 cells were mediated via MEK/ERK1/2 signaling pathway.
A number of studies revealed that the sclerotium of Polyporus rhinocerus (synonym of L. rhinocerotis) demonstrated anti-tumour27,28,29, immunomodulating30,31,32 and antioxidant33 activities in in vitro (Table 1). However, very limited information is available on the neuronal activity of L. rhinocerotis. Our recent findings revealed that the hot aqueous extract of L. rhinocerotis sclerotium stimulated neuritogenesis in PC-12 cells34. To further investigate the neuritogenic activity of the L. rhinocerotis sclerotium, in the present study, we aimed to (i) compare the cell viability and neuritogenic effects of PC-12 cells of three different preparations, including hot aqueous and ethanolic extracts and crude polysaccharides of L. rhinocerotis sclerotium, (ii) investigate whether the extracts and crude polysaccharides stimulate the production of NGF and (iii) investigate the involvement of NGF responsive signaling pathway (MEK/ERK1/2) in L. rhinocerotis sclerotium-stimulated neuritogenesis in PC-12 cells.
Results and Discussions
The effects of hot aqueous and ethanolic extracts and crude polysaccharides on viability of PC-12 cells
The viability of cells in complete F-12 K medium was considered as 100%. The survival and proliferation of treated cells decreased in a concentration-dependent manner for hot aqueous extract (Fig. 1A), ethanolic extract (Fig. 1B) and crude polysaccharides (Fig. 1C). Hot aqueous extract and crude polysaccharides increased the cells proliferation significantly (p < 0.05) at low concentration, 9.77 μg/ml, compared to the negative control (cells in complete F-12 K medium only). The percentage of viable cells decreased gradually as the concentration of the extracts and crude polysaccharides were increased. The percentage of viable cells in hot aqueous and ethanolic extracts-treated cells were reduced significantly (p < 0.05) starting at 156.25 μg/ml compared to negative control, while the viability of crude polysaccharides-treated cells was reduced significantly (p < 0.05) starting at a low concentration, 19.53 μg/ml, compared to negative control. The required concentrations to inhibit the cell growth by 50% (IC50) for hot aqueous, ethanolic extracts and crude polysaccharides were 3223.98 μg/ml, 372.30 μg/ml and 2718.72 μg/ml, respectively. The cytoxicity of the hot aqueous extract was significantly (p < 0.05) lower in the in vitro PC-12 cell model when compared to the crude polysaccharides and ethanolic extract. In a study done by Lee et al. (2011, 2013)35,36, it was reported that in the in vivo model, no treatment-related chronic toxicity was detected in Sprague Dawley rats after a long term (180 days) oral administration of the L. rhinocerotis sclerotium freeze-dried powder (cultivar TM02) at daily dosage up to 1,000 mg/kg. Further, blood biochemical parameters related to toxicity were reported as normal. In that study, however, Lee et al. (2011, 2013) did not estimate the levels of the mushroom in the blood.

The effects of hot aqueous and ethanolic extracts and crude polysaccharides of L. rhinocerotis sclerotium on viability of PC-12 cells.
Cells were incubated with extracts or crude polysaccharides at concentrations from 0 to 2500 μg/ml for 48 h. (A) hot aqueous extract, (B) ethanolic extract and (C) crude polysaccharides. The mean absorbance obtained using complete F-12 K medium with cells only (negative control) was designated 100% of cell viability. Results are shown as means ± standard deviation (n = 3). *p < 0.05 compared to the respective negative control 100%.
The neuritogenic effects of hot aqueous and ethanolic extracts and crude polysaccharides in PC-12 cells
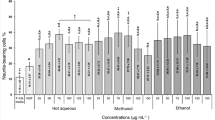
Neurite extension of PC-12 cells was regarded as an index of neuritogenesis. All the tested concentrations of hot aqueous and ethanolic extracts and crude polysaccharides stimulated neuritogenesis in PC-12 cells after 48 h of incubation (Fig. 2). The percentage of neurite-bearing cells in all the tested concentrations of extracts and crude polysaccharides treated cells were increased significantly (p < 0.05) compared to the negative control (cells in complete F-12 K medium only). The percentage of neurite-bearing cells was concentration-dependent (from 5 μg/ml to 100 μg/ml) of extracts and crude polysaccharides tested. There was an initial increase of the percentage of neurite bearing cells on increasing concentrations of extracts and crude polysaccharides, followed by a decrease at concentrations above 25 μg/ml. All the extracts and crude polysaccharides stimulated maximal neuritogenesis in PC-12 cells at 25 μg/ml. The hot aqueous extract stimulated highest percentage of neurite-bearing cells (20.99 ± 1.01%), followed by the ethanolic extract (17.35 ± 0.66%) and crude polysaccharides (16.40 ± 0.26%), at 25 μg/ml. The IC50 value of the cytotoxic activity of the hot aqueous extract, ethanolic extract and crude polysaccharides and was approximately 129-, 15- and 109- fold higher than their optimum concentration that stimulated neuritogenesis, 25 μg/ml. Our previous study34 reported that 50 ng/ml of NGF was the optimum concentration for neuritogenesis in PC-12 cells. In the present study, cells treated with 50 ng/ml of NGF served as a positive control. Interestingly, there was no significant difference (p > 0.05) in the percentage of neurite-bearing cells between 50 ng/ml of NGF- and 25 μg/ml of hot aqueous extract-stimulated neuritogenesis. This is in agreement with the previous studies, reported that 20 μg/ml of hot aqueous extract of L. rhinocerotis sclerotium34 and L. rhinocerotis mycelium37 stimulated neuritogenesis in PC-12 cells that comparable to NGF.

The neuritogenic effects of hot aqueous and ethanolic extracts and crude polysaccharides in PC-12 cells.
Cells were incubated with NGF (50 ng/ml), extracts or crude polysaccharides (5 to 100 μg/ml) for 48 h. Cells in complete F-12 K medium served as a negative control. Cells treated with 50 ng/ml of NGF served as a positive control. Data were expressed as means ± standard deviation (n = 3). *p < 0.05 compared to the negative control. #p < 0.05 compared to the positive control (50 ng/ml of NGF).
The hot aqueous extraction is the most commonly used method by the indigenous communities and traditional Chinese medicine (TCM) physicians to prepare decoctions, tonics or essences from medicinal mushrooms. In general, mushroom’s polysaccharides are targeted as the active components in the hot aqueous extract38. Mushroom’s polysaccharides include α-glucans and β-glucans, that known as effective anti-tumor and immunomodulatory agents39. Besides carbohydrates and proteins, Lau et al. (2013)29 reported that the hot aqueous extract of L. rhinocerotis sclerotium contained a higher percentage of phenolics than the cold aqueous extract. Lau et al. (2013)29 further suggested that secondary metabolites such as triterpenes and alkaloids may be present in L. rhinocerotis sclerotium extracts. Secondary metabolites isolated from different mushrooms that were reported to promote neuritogenesis in vitro and/or in vivo, include hericenones40, erinacines41, scabronions42, termitomycesphins43 and cyrneines44. These neuroactive compounds may be targeted as preventive and therapeutic agents for neurodegenerative disorders. In the present study, the hot aqueous extract of L. rhinocerotis sclerotium showed potent neuritogenic activity compared to the ethanolic extract and crude polysaccharides. These findings suggested that the hot aqueous extract containing chemical constituents that are neuroactive.
The morphology of PC-12 cells stained with anti-neurofilament-200 antibody
Neurofilaments are neuron specific intermediate filament proteins (8–10 nm) that are located in axons, found specifically in most mature neurons45. Neurofilaments are composed predominantly of distinct subunits, namely neurofilament light (NF-L), medium (NF-M) and heavy (NF-H)46. The anti-NF-200 antibody recognizes both phosphorylated and non-phosphorylated forms of heavy neurofilament subunit NF-H at 180–220 kDa. Immunostaining of neurofilaments confirmed the neuritogenesis was stimulated by NGF, hot aqueous, ethanolic and crude polysaccharides extracts (Fig. 3). PC-12 cells nuclei were stained blue by DAPI and neurofilaments were stained green by anti-NF-200 antibody labelled with FITC. Cells were elongated and exhibited significant neurite extensions in NGF-, hot aqueous extract-, ethanolic extract- and crude polysaccharides-stimulated neuritogenesis.

Morphology of PC-12 cells stained with anti-NF-200 antibody.
Cells were incubated with or without NGF (50 ng/ml), hot aqueous extract (25 μg/ml), ethanolic extract (25 μg/ml) or crude polysaccharides (25 μg/ml) for 48 h. Cells in complete F-12 K medium served as a negative control. Cells treated with 50 ng/ml of NGF served as a positive control. Nuclei stained blue and neurofilaments stained green. Scale bars represent 50 μM. Arrows indicate neurite outgrowth.
The concentration of extracellular NGF of hot aqueous and ethanolic extracts- and crude polysaccharides-treated cells
The increase in extracellular NGF in cell supernatant showed the ability of tested compounds to induce NGF production by PC-12 cells47. The concentration of extracellular NGF of PC-12 cells without treatment (negative control) was detected at 65.64 pg/ml (Fig. 4). The concentration of extracellular NGF in 50 ng/ml of NGF-treated cells (positive control) was 353.42 pg/ml, which was approximately five fold increase compared to the negative control. However, there was no significant difference (p > 0.05) in the concentration of extracellular NGF between all the tested concentrations of the extracts and crude polysaccharides and the negative control.

Extracellular NGF concentration in supernatants of NGF-, extracts- or crude polysaccharides-treated PC-12 cells.
Cells were incubated with or without NGF (50 ng/ml), extracts or crude polysaccharides (25 to 100 μg/ml) for 48 h. Cells in complete F-12 K medium served as a negative control. Cells treated with 50 ng/ml of NGF served as a positive control. Data were expressed as means ± standard deviation (n = 3). Means with different alphabets show significant difference (p < 0.05).
The present findings showed that the concentration of extracellular NGF in 50 ng/ml of NGF-treated cells increased by approximately 81% compared to the non-treated cells. Based on the amount of extracellular NGF measured and percentage of neurite bearing cells observed in positive control (NGF-treated cells), neuritogenesis in PC-12 cells may be NGF-dependent. Number of studies documented the potential of extracts and compounds of edible and medicinal mushrooms to stimulate the biosynthesis and secretion of NGF in vitro40,41,42,44,47,48. According to Lai et al. (2013)48, hot aqueous extract of Hericium erinaceus (Bull.: Fr.) Pers. (lion’s mane mushroom) stimulated the production of NGF in NG108–15 cells, a hybrid neuronal cell line derived from mouse neuroblastoma and rat glioma. The concentration of the extracellular NGF in NG108–15 cells treated with 50 μg/ml of hot aqueous extract of H. erinaceus was 21.4% higher, compared to the cells treated with 20 ng/ml of NGF48. Compounds from H. erinaceus, including hericenone C-E40, erinacine A-C41 stimulated biosynthesis of NGF and exhibited neuritogenesis in astroglial cells. In a recent study by Phan et al. (2014)47 reported that hericenones E potentiated NGF-induced neuritogenesis in PC-12 cells by stimulating the production of NGF that was almost two times higher than that of positive control (50 ng/ml of NGF). However, in the present study, L. rhinocerotis sclerotium extracts and crude polysaccharides stimulated the neuritogenic activity without stimulating the production of NGF in PC-12 cells. These findings showed that the extracts might contain NGF-like compound(s) (NGF mimics or substitute for NGF) that mimic the neuritogenic activity of the NGF.
The involvement of NGF responsive signaling pathway in hot aqueous extract-stimulated neuritogenesis
The NGF responsive pathway, TrkA-MEK1/2-ERK1/2 was selected as the targeted cascade for neuritogenic activity in PC-12 cells. The Trk and MEK/ERK1/2 inhibitors, namely K252a, U0126 and PD98059 significantly (p < 0.05) blocked the NGF- and hot aqueous extract-stimulated neuritogenesis (Fig. 5). The K252a, U0126 and PD98059 decreased the percentage of neurite-bearing cells by approximately 82.13%, 86.15% and 91.56% in NGF-treated cells and 80.97%, 86.68% and 84.59% in hot aqueous extract-treated cells, respectively. The significant (p < 0.05) reduction of neurite stimulation activity was also observed in the negative control with the addition of the inhibitors.

The effects of the specific inhibitors of TrkA and MEK/ERK1/2 on NGF-, or hot aqueous extract-stimulated neuritogenesis.
Cells were pre-treated with K252a, U0126, PD98059 for one hour before the treatment with NGF (50 ng/ml) or hot aqueous extract (25 μg/ml). Cells in complete F-12 K medium served as a negative control. Cells treated with 50 ng/ml of NGF served as a positive control. A control (without inhibitor) was used in each treatment group. Data were expressed as means ± standard deviation (n = 3). *p < 0.05 compared to the respective controls.
TrkA is a cell surface transmembrane receptor tyrosine kinase for NGF and activated TrkA is critical for activation of the Ras/MAPK signaling pathway49,50. Nerve growth factor binds to its high affinity receptor, TrkA to initiates the NGF responsive pathways, such as the MEK/ERK1/2 signaling pathway, to kick start neuritogenesis. Once the TrkA was phosphorylated, it became a scaffolding structure and recruits proteins that ultimately propagate the MEK/ERK signaling pathway51. Specific inhibitors of protein kinase served as powerful tool to study the kinase activities in selected signaling pathway2. K252a acts as a specific and potent inhibitor of Trk receptor, inhibits the phosphorylation of NGF-induced TrkA and selectively blocks the activities of NGF in PC-12 cells52. In the present study, the neuritogenic activity of hot aqueous extract was blocked 80.97% by K252a, parallel to the inhibition effect of K252a (82.13%) towards NGF-treated cells. These findings showed that the hot aqueous extract-stimulated neuritogenesis was TrkA-dependant in PC-12 cells.
Both U0126 and PD98059 are selective and potent inhibitors of MEK 1 and MEK 253. Priming PC-12 cells with U0126 and PD98059 will inhibit the phosphorylation and activation of MEK/ERK1/2 and eventually diminish cell differentiation and neuritogenesis54. Nishina et al. (2006)55 demonstrated that the activation of MAPK by lysophosphatidylethanolamine, a neuroactive compound extracted from Grifola frondosa (maitake mushroom) was suppressed by U0126, but not by K252a. The study suggested that the MEK/ERK1/2 signaling pathway was involved in lysophosphatidylethanolamine-induced neuritogenesis in PC12 cells, but was not through the activation of TrkA receptor55. Phan et al. (2014)47 reported that the neuritogenic activity potentiated by hericenone E was found to be partially blocked (46%) by K252a and almost completely blocked by U0126 and PD98059, suggested that hericenone E potentiated NGF-stimulated neuritogenesis in PC12 cells was partially mediated by TrkA and MEK/ERK1/2 dependent. In the present study, all three inhibitors (K252a, U0126 and PD98059) successfully attenuated the NGF- and hot aqueous extract-stimulated neuritogenesis in PC-12 cells, showed that the hot aqueous extract of L. rhinocerotis sclerotium mimicked the NGF neuritogenic activity by binding to the TrkA receptor and activated the MEK/ERK1/2 signaling pathway in PC-12 cells.
The protein expression of total p44/42 MAPK (ERK1/2) and phosphorylated p44/42 MAPK (Thr202/Tyr204) in hot aqueous extract-treated cells
Endogenous level of total p44/42 MAPK (ERK1/2) and phosphorylated p44/42 MAPK (Thr202/Tyr204) were quantified by ELISA. The magnitude of absorbance for the developed colour is proportional to the quantity of MAPK protein expressed by PC-12 cells. The expression of both total and phosphorylated p44/42 MAPK protein in PC-12 cells treated with NGF and hot aqueous extract were significantly (p < 0.05) higher compared to the negative control (Fig. 6). Both total and phosphorylated p44/42 protein concentrations in NGF-treated cells were higher compared to the hot aqueous extract-treated cells and negative control (p < 0.05). However, there was no significant difference (p > 0.05) in protein expression between NGF- and hot aqueous extract-treated PC-12 cells for both total and phosphorylated p44/42, pERK1 and pERK2.

Protein expression of total p44/42 MAPK (Erk1/2) and phosphorylated p44/42 MAPK (Thr202/Tyr204) activation in PC-12 after 48 h of incubation with NGF or hot aqueous extract.
Cells were incubated with or without NGF (50 ng/ml) or hot aqueous extract (25 μg/ml) for 48 h. Cells in complete F-12 K medium served as a negative control. Cells treated with 50 ng/ml of NGF served as a positive control. Total p44/42 and phosphorylated p44/42 protein level were measured by determination of the optical intensity (OD). Data were expressed as means ± standard deviation (n = 3). Different alphabets indicate significant difference (p < 0.05).
Activation of NGF responsive pathway by mushroom extract is crucial in the preliminary search of neuroactive compound(s) that may mimic the neuritogenic activity of NGF. Activation of MEK/ERK1/2 provided the biochemical evidence for neuritogenesis and the presence of neurite-stimulating agent(s) in the extract. Cheung et al. (2000)24 reported that G. lucidum aqueous extract promoted neuritogenic and neuroprotective activity via Ras/ERK pathway in PC-12 cells, by demonstrating the phosphorylation of ERK1 and ERK2. In the present study, the endogenous level of total and phosphorylated p44 (ERK1) and p42 (ERK2) proteins were elevated, suggesting that the phosphorylation and activation of ERK1/2 were involved in the stimulation of neuritogenesis in PC-12 cells by the hot aqueous extract of L. rhinocerotis sclerotium.
Immunofluorescence study demonstrated the protein expression of phosphorylated p44/42 MAPK (Thr202/Tyr204) in hot aqueous extract-treated cells
Immunofluorescence staining was served as a visualize support to the protein expression of phosphorylated p44/42 MAPK (Thr202/Tyr204) (Fig. 7). The intensity of the immunofluorescence staining demonstrated the protein expression level of phosphorylated p44/42 protein in PC-12 cells. The intensity of immunofluorescence staining of phosphorylated p44/42 protein was higher in NGF- (Fig. 7B) and hot aqueous extract- (Fig. 7C) treated cells than the negative control (Fig. 7A). An MEK/ERK1/2 inhibitor, U0126 was used as a control to ensure the involvement of MEK/ERK1/2 signaling pathway in neuritogenesis. Pre-treatment with U0126 in PC-12 cells showed lower signal intensity (Fig. 7D–F). Results showed that neuritogenesis in PC-12 cells was dependent on the activation of MEK/ERK1/2 signaling pathway.

Immunocytochemistry study of phosphorylated p44/42 MAPK (Thr202/Tyr204) in NGF- or hot aqueous extract-treated PC-12 cells after 48 h of incubation.
(A) Negative control, cells in complete F-12 K medium only. (B) Positive control, cells treated with 50 ng/ml of NGF. (C) Cells treated with 25 μg/ml of hot aqueous extract. (D–F) Cells were pre-treated with 10 μM U0126 prior to the treatment with NGF or hot aqueous extract. Phosphorylated p44/42 protein stained cell body in green. DAPI stained nuclei in blue. Scale bar = 50 μm.
Conclusions
The hot aqueous and ethanolic extracts and crude polysaccharides of L. rhinocerotis sclerotium stimulated neuritogenesis in PC-12 cells. All the concentrations of the extracts and crude polysaccharides tested for neuritogenic activity were not cytotoxic to PC-12 cells. The hot aqueous extract (25 μg/ml) stimulated neuritogenic activity that was comparable to NGF (50 ng/ml). The extracts and crude polysaccharides stimulated neuritogenic activity but did not stimulate the production of NGF in PC12 cells. The neuritogenic activity of NGF- and hot aqueous extract may be mediated through the phosphorylation of TrkA receptor and ERK1/2 signaling pathway in PC-12 cells. Lignosus rhinocerotis sclerotium may contain neuroactive compound(s) that mimic the neuritogenic activity of NGF and induce neuritogenesis in PC-12 cells via the NGF responsive pathway, TrkA-MEK1/2-ERK1/2 signaling pathway.
Methods
Preparation of hot aqueous and ethanolic extracts and crude polysaccharides of L. rhinocerotis sclerotium
Lignosus rhinocerotis sclerotium light brown and dry fluffy freeze-dried powder (LiGNO™ cultivar TM02) was purchased from Ligno Biotech Sdn. Bhd., Malaysia56. Every batch of cultivar TM02 freeze-dried powder is identified and validated by the internal transcribed spacer regions of ribosomal RNA57. The hot aqueous extraction was carried out according to Wong et al. (2007)58 with modification. Briefly, the freeze-dried powder was soaked in distilled water at a ratio of 1:20 (w/v) and was agitated on a shaker at 150 rpm at room temperature, overnight. Then, the mixture was double boiled in a water bath for 30 min and cooled to room temperature. The mixture was then centrifuged at 7,800 × g for 15 min and the supernatant was collected and filtered through Whatman no. 4 filter paper. The resulting hot aqueous extract was freeze-dried and kept at −20 °C prior to use. For the ethanolic extract, the freeze-dried powder was soaked in 80% ethanol (v/v in distilled water) at room temperature for three days and the process was repeated three times. The ethanol was evaporated using a rotary evaporator (Eyela N-1000). The resulting ethanolic extract was kept in −20 °C prior to use. The crude polysaccharides were extracted according to the alkaline extraction method of Ojha et al. (2010)59. The freeze-dried powder was soaked in 4% (w/v) sodium hydroxide (NaOH) and heated in 80 °C water bath for 45 min, then the mixture was centrifuged at 7,800 × g for 45 min. Supernatant was collected and precipitated with absolute ethanol at a ratio of 1:5 (v/v). The mixture was kept for 12 h at 4 °C. The precipitated polysaccharides were centrifuged at 7,800 × g for 45 min. The residue was dialysed using diethylaminoethyl (DEAE) cellulose bag for 4 h. The crude polysaccharides were freeze-dried and kept at −20 °C prior to use.
In vitro cell culture
The PC-12 cells (American Type Culture Collection, ATCC) were maintained in complete Kaighn’s Modification of Ham’s F-12 (F-12 K) Medium (Sigma) supplemented with 15% of heat-inactivated horse serum and 2.5% of heat-inactivated Fetal bovine serum (PAA Laboratories) at 37 ± 2 °C in a 5% CO2-humidified incubator. The cells were passaged every 2 to 3 days upon 80% confluent.
Assessment of the effects of hot aqueous and ethanolic extracts and crude polysaccharides on viability of PC-12 cells
Cells were plated at a density of 1 × 104 cells per well in 96-well plates and incubated overnight at 37 °C in a 5% CO2-humidified incubator. Then, the supernatant was carefully replaced with freshly prepared extracts or crude polysaccharides (0–2500 μg/ml) in complete F-12 K medium. After 48 h of incubation, 3-(4,5-dimethythiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was performed as described previously26. The extent of the reduction of MTT was determined by measurement of the absorbance at 540 nm with 690 nm as background absorbance with an ELISA microplate reader (Sunrise, Tecan). The complete F-12 K medium was the blank and cells incubated in the medium only were denoted as the negative control. The 50% inhibitory concentration (IC50) was interpolated from the response curve.
Assessment of neuritogenic activity of hot aqueous and ethanolic extracts and crude polysaccharides in PC-12 cells
Stimulation of neuritogenesis in PC-12 cells
Cells were plated at a density of 5 × 103 cells per well in 12-well plates and then treated with freshly prepared extracts or crude polysaccharides (25 to 100 μg/ml) in complete F-12 K medium. Cells treated with 50 ng/ml of NGF (Sigma) served as a positive control, while cells in complete F-12 K medium without treatment served as a negative control. Assay plates were incubated for 48 h at 37 ± 2 °C in a 5% CO2-humidified incubator prior to scoring the neurite-bearing cells.
Quantification of neurite bearing-cells
Differentiated cells were counted by visual examination of the field. A neurite-bearing cell was defined as a cell with one or more axon-like extension that was double or more the length of the cell body diameter58. Ten selected microscopic fields with an average of 200–300 cells per well were assessed under an inverted microscope (Nikon Eclipse TS100). The images were captured with a QImaging Go-3 color CMOS Camera (QImaging) and by the image processor system, Image-Pro Insight (MediaCybernetics). The percentage of neurite-bearing cells was evaluated by scoring the proportion of neurite-bearing cells to the total number of cells in a well.
Immunofluorescence staining of neurofilaments
Immunofluorescence assay was carried out according to Schimmelpfeng et al. (2004)60. Briefly, cells were seeded at a density of 5 × 103 cells per well in 12-well micro-chamber (ibidi). The cells were treated with the extracts or crude polysaccharides for 48 h at 37 ± 2 °C in a 5% CO2-humidified incubator. After fixing with 4% paraformaldehyde, the cells were incubated with anti-neurofilament 200 antibody produced in rabbit (1:80; Sigma) and followed by fluorophore-conjugated secondary antibody, anti-rabbit IgG-fluorescein isothiocyanate (FITC) antibody produced in sheep (1:80; Sigma). Finally, the cells were mounted with ProLong® gold antifade reagent with 4-6-Diamidino-2-phenylindole (DAPI) (Life Technologies Corporation). The slides were observed under fluorescence illumination using FITC and DAPI filters and images were captured with Nikon’s Imaging Software, NIS-Elements.
Quantification of the concentration of extracellular NGF in cell culture supernatant
Cells were plated at a density of 1 × 104 cells per well in 96-well plates. The cells were treated with freshly prepared extracts or crude polysaccharides (25 to 100 μg/ml) in complete F-12 K medium for 48 h. The cell culture supernatant was collected, centrifuged at 1500 × g for 15 min and maintained at 0–4 °C prior to assay. The samples were diluted with sample diluent at a ratio of 1:2 (v/v). The amount of NGF in culture supernatant was measured by using ChemiKineTM nerve growth factor sandwich enzyme-linked immunosorbent assay (ELISA) kit (Chemicon® International Inc.) according to the manufacturer’s protocol.
Elucidation of the involvement of NGF responsive pathway in the hot aqueous extract-stimulated neuritogenesis in PC-12 cells
Treatment with specific inhibitors of signaling pathway
The inhibitors for Trk receptor (K252a) and MEK/ERK1/2 signaling pathway (U0126, PD98059) (all Sigma, USA) were used. Stock solutions (10 mM) of the inhibitors were prepared in dimethyl sulfoxide (DMSO) and stored at −20 °C in the dark. Final concentrations of 100 nM of K252a, 10 μM of U0126 and 40 μM of PD98059 were prepared by diluting in complete F-12 K medium before use. Cells were pre-incubated either with or without the inhibitor for one hour at 37 ± 2 °C in a 5% CO2-humidified incubator, respectively before the treatment with 50 ng/ml of NGF or 25 μg/ml of hot aqueous extract (the concentration that stimulated maximal percentage of neurite-bearing cells). Cells were then incubated for 48 h prior to scoring the neurite-bearing cells.
Quantification of protein expression of total p44/42 MAPK (ERK1/2) and phosphorylated p44/42 MAPK (Thr202/Tyr204)
Cells were pre-incubated either with or without the MEK/ERK1/2 inhibitor, U0126 for one hour at 37 ± 2 °C in a 5% CO2-humidified incubator, before the treatment with 50 ng/ml of NGF or 25 μg/ml of hot aqueous extract for 48 h. Cells were washed and harvested in PBS and centrifuged at 10,000 × g for 10 min at 4 °C. Cell pellets were resuspended in cell lysis and protein extraction buffer (Thermo Fisher Scientific Inc.) with protease inhibitor cocktail and 1 mM of phenylmethylsufonyl fluoride (Sigma) on ice and vortex every 10 min for three times. Then, the extracted proteins were centrifuged and pelleted at 10,000 × g for 10 min at 4 °C. Supernatants were aliquoted into clean micro centrifuge tubes and kept cool prior to assay. The protein expression was quantified by using the total p44/42 MAPK (ERK1/2) and phosphorylated p44/42 MAPK (Thr202/Tyr204) sandwich ELISA kits (Cell Signaling Technology) according to the manufacturer’s protocol. The absorbance was recorded at 450 nm with an ELISA microplate reader (Sunrise, Tecan). Concentration of the protein antibodies of every treatment were quantified according to the standard graph and multiplied by the appropriate dilution factor.
Immunofluorescence study of protein expression of phosphorylated p44/42 MAPK (Thr202/Tyr204)
Cells were pre-incubated either with or without the U0126 inhibitor for one hour at 37 ± 2 °C in a 5% CO2-humidified incubator before the treatment with 50 ng/ml of NGF or 25 μg/ml of hot aqueous extract. Cells were then incubated for 48 h prior to immunofluorescence staining. Protein expression of phosphorylated p44/42 MAPK (Thr202/Tyr204) (1:100; Cell Signaling Technology) detection antibody was used as the primary antibody. The slides were observed under fluorescence illumination using FITC and DAPI filters and images were captured with Nikon’s Imaging Software, NIS-Elements.
Statistical analysis
All the experimental data were expressed as the mean ± standard deviation (SD) of triplicate values. Statistical differences between groups were assessed using one-way analysis of variance (ANOVA) of a minimum of three independent experiments and Duncan’s multiple range test (DMRT), p < 0.05 was considered to be significant.
Additional Information
How to cite this article: Seow, S. L.-S. et al. Lignosus rhinocerotis (Cooke) Ryvarden mimics the neuritogenic activity of nerve growth factor via MEK/ERK1/2 signaling pathway in PC-12 cells. Sci. Rep. 5, 16349; doi: 10.1038/srep16349 (2015).
References
Wasser, S. P. & Weis, A. L. Therapeutic effects of substance occurring in higher basidiomycetes mushrooms: A modern perspective. Critical Reviews in Immunology. 19(1), 65–96 (1999).
Phan, C. W., Wong, W. L., David, P., Naidu, M. & Sabaratnam, V. Therapeutic potential of culinary-medicinal mushrooms for the management of neurodegenerative diseases: diversity, metabolite and mechanism. Critical Reviews in Biotechnology. 1–14 (2014), doi: 10.3109/07388551.2014.887649.
Tan, C. S., Ng, S. T., Yap, Y. H. Y., Lee, S. S., Lee, M. L. et al. Breathing new life to a Malaysia lost national treasure – the tiger’s milk mushroom (Lignosus rhinocerotis) in Mushroom Science XVIII: Proceedings of the18th Congress of the International Society for Mushroom Science (eds Zhang, J. et al.) 66–71 (Beijing, China, 2012).
Lee, S. S., Chang, Y. S. & Noraswati, M. N. R. Utilization of macrofungi by some indigenous communities for food and medicine in Peninsular Malaysia. Forest Ecology and Management. 257, 2062–2065 (2009).
Burkill, I. H., Birtwistle, W., Foxworthy, F. W., Scrivenor, J. B. & Watson, J. B. A Dictionary of the Economic Products of the Malay Peninsula, volume I & II. Kuala Lumpur: Governments of Malaysia and Singapore by the Ministry of Agriculture and Co-operatives. (Malaysia, 1966).
Chang, Y. S. & Lee, S. S. Utilisation of macrofungi species in Malaysia. Fungal Diversity. 15, 15–22 (2004).
Huang, N. L. Identification of the scientific name of Hurulingzhi. Acta Edulis Fungi. 6, 30–32 (1999).
Da Silva, J. S. & Dotti, C. G. Breaking the neuronal sphere: regulation of the actin cytoskeleton in neuritogenesis. Nature Reviews Neuroscience. 3, 694–704 (2002).
Kromer, L. F. Nerve growth factor treatment after brain injury prevents neuronal death. Science. 235, 214–216 (1987).
Thoenen, H. Neurotrophins and neuronal plasticity. Science. 270, 593–598 (1995).
Koliatsos, V. E., Clatterbuck, R. E., Nauta, W. H. J., Knusel, B., Burton, L. E. et al. Recombinant human nerve growth factor prevents degeneration of basal forebrain cholinergic neurons in primates. Annals of Neurology. 30, 831–840 (1991).
Hefti, F. & Weiner, W. J. Nerve growth factor and Alzheimer’s disease. Annals of Neurology. 20, 275–281 (1986).
Capsoni, S., Ugolini, G., Comparini, A., Ruberti, F., Berardi, N. et al. Alzheimer-like neurodegeneration in aged anti-nerve growth factor transgenic mice. Proceedings of the National Academy of Sciences of the United States of America. 97, 6826–6831 (2000).
Granholm, A. C., Sanders, L. A., Reyland, M. E., Hoernig, G. R., Shen, L. et al. Transplantation of fetal tissues from trophic factor knockout mice to wildtype and heterozygous siblings. In: Abstract American Society for Neural Transplantation. (Clearwater, FL, 1998).
Williams, B. & Dwyer, D. S. Structure-based discovery of low molecular weight compounds that stimulate neurite outgrowth and substitute for nerve growth factor. Journal of Neurochemistry. 110(6), 1876–1884 (2009).
Kao, S. C., Jaiswal, R. K., Kolch, W. & Landreth, G. E. Identification of the mechanisms regulating the differential activation of the MAPK cascade by epidermal growth factor and nerve growth factor in PC12 cells. The Journal of Biological Chemistry. 276(21), 18169–18177 (2001).
Roskoski, R. ERK1/2 MAP kinases: structure, function and regulation. Pharmacological Research. 66, 105–143 (2012).
Vaudry, D., Stork, P. J. S., Lazarovici, P. & Eiden, L. E. Signaling pathways for PC-12 cell differentiation: making the right connections. Science. 296(5573), 1648–1649 (2002).
Naidu, M., David, P., Asher, R. & Fawcett, J. Expression of Akt and MAPK in the normal and regenerating peripheral nerve and their dorsal root ganglia. Malaysian Journal of Biochemistry and Molecular Biology. 17, 16–19 (2009).
Wong, K. H., Naidu, M., David, P., Bakar, R. & Sabaratnam, V. Neuroregenerative potential of lion’s mane mushroom, Hericium erinaceus (Bull.: Fr.) Pers. (Higher Basidiomycetes), in the treatment of peripheral nerve injury (Review). International Journal of Medicinal Mushrooms. 14, 427–446 (2012).
Parmar, P. K., Coates, L. C., Pearson, J. F., Hill, R. M. & Birch, N. P. Neuroserpin regulates neurite outgrowth in nerve growth factor-treated PC12 cells. Journal of Neurochemistry. 82, 1406–1415 (2002).
Teng, K. K., Angelastro, J. M., Cunningham, M. E. & Greene, L. A. Cultured PC12 cells: A model for neuronal function, differentiation and survival in Cell Biology: A Laboratory Handbook. 3rd ed. (ed. Colin, J. E. ) 171–176 (USA: Elsevier Science, 2006).
Greene, L. A. & Tischler, A. S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proceedings of the National Academy of Sciences. 73, 2424–2428 (1976).
Cheung, W. M. W., Hui, W. S., Chu, P. W. K., Chiu, S. W. & Ip, N. Y. I. Ganoderma extract activates MAP kinases and induces the neuronal differentiation of rat pheochromocytoma PC12 cells. FEBS Letters. 486, 291–296 (2000).
Phan, C. W., Wong, W. L., David, P., Naidu, M. & Sabaratnam, V. Pleurotus giganteus (Berk.) Karunarathna & K. D. Hyde: nutritional value and in vitro neurite outgrowth activity in rat pheochromocytoma cells. BMC Complementary and Alternative Medicine. 12, 102 (2012), doi: 10.1186/1472-6882-12-102.
Seow, S. L. S., Naidu, M., David, P., Wong, K. H. & Sabaratnam, V. Potentiation of neuritogenic activity of medicinal mushrooms in rat pheochromocytoma cells. BMC Complementary and Alternative Medicine. 13, 157 (2013), doi: 10.1186/1472-6882-13-157.
Lai, C. K. M., Wong, K. H. & Cheung, P. C. K. Antiproliferative effects of sclerotial polysaccharides from Polyporus rhinocerus Cooke (Aphyllophoromycetideae) on different kinds of leukemic cells. International Journal of Medicinal Mushrooms. 10, 255–264 (2008).
Lee, M. L., Tan, N. H., Fung, S. Y., Tan, C. S. & Ng, S. T. The antiproliferative activity of sclerotia of Lignosus rhinocerus (Tiger Milk Mushroom). Evidence-based complementary and alternative medicine. eCAM, 697603 (2012), doi: 10.1155/2012/697603.
Lau, B. F., Abdullah, N., Aminudin, N. & Lee, H. B. Chemical composition and cellular toxicity of ethnobotanical-based hot and cold aqueous preparations of the tiger’s milk mushroom (Lignosus rhinocerotis). Journal of Ethnopharmacology. 150, 252–262 (2013).
Wong, K. H., Lai, C. K. M. & Cheung, P. C. K. Stimulation of human innate immune cells by medicinal mushroom sclerotial polysaccharides. International Journal of Medicinal Mushrooms. 11, 215–223 (2009).
Wong, K. H., Lai, C. K. M. & Cheung, P. C. K. Immunomodulatory activities of mushroom sclerotial polysaccharides. Food Hydrocolloids 25(2), 150–158 (2011).
Guo, C., Wong, K. H. & Cheung, P. C. Hot water extract of the sclerotium of Polyporus rhinocerus Cooke enhances the immune functions of murine macrophages. International Journal of Medicinal Mushrooms. 13(3), 237–44 (2011).
Yap, Y. H. Y., Tan, N., Fung, S., Aziz, A. A., Tan, C. et al. Nutrient composition, antioxidant properties and anti-proliferative activity of Lignosus rhinocerus Cooke sclerotium. Journal of the Science of Food and Agriculture. 93, 2945–2952 (2013).
Eik, L. F., Naidu, M., David, P., Wong, K. H., Tan, Y. S. et al. Lignosus rhinocerus (Cooke) Ryvarden: a medicinal mushroom that stimulates neurite outgrowth in PC-12 cells. Evidence-based Complementary and Alternative Medicine. eCAM, 320308 (2012), doi: 10.1155/2012/320308.
Lee, S. S., Tan, N. H., Fung, S. Y., Pailoor, J. & Sim, S. Evaluation of the sub-acute toxicity of the sclerotium of Lignosus rhinocerus (Cooke), the Tiger Milk mushroom. Journal of Ethnopharmacology. 138, 192–200 (2011).
Lee, S. S., Enchang, F. K., Tan, N. H., Fung, S. Y. & Pailoor, J. Preclinical toxicological evaluations of the sclerotium of Lignosus rhinocerus (Cooke), the tiger milk mushroom. Journal of Ethnopharmacology. 147, 157–163 (2013).
John, P. A., Wong, K. H., Naidu, M., Sabaratnam, V. & David, P. Combination effects of curcumin and aqueous extract of Lignosus rhinocerotis mycelium on neurite outgrowth stimulation activity in PC-12 cells. Natural Product Communications. 8, 711–714 (2013).
Mizuno, T. The extraction and development of antitumour-active polysaccharides from medicinal mushrooms in Japan. International Journal of Medicinal Mushrooms. 1, 9–29 (1999).
Vannucci, L., Krizan, J., Sima, P., Stakheev, D., Caja, F. et al. Immunostimulatory properties and antitumor activities of glucans (Review). International Journal of Oncology. 43, 357–364 (2013).
Kawagishi, H., Ando, M., Sakamoto, H., Yoshida, S., Ojima, F. et al. Hericenones C, D and E, stimulators of nerve growth factor (NGF)-synthesis, from the mushroom Hericium erinaceum. Tetrahedron Letters. 32, 4561–4564 (1991).
Kawagishi, H., Shimada, A., Shirai, R., Okamoto, K., Ojima, F. et al. Erinacines A, B and C, strong stimulators of nerve growth factor (NGF)-synthesis, from the mycelia of Hericium erinaceum. Tetrahedron Letters. 35, 1569–1572 (1994).
Obara, Y., Nakahata, N., Kita, T., Takaya, Y., Kobayashi, H. et al. Stimulation of neurotrophic factor secretion from 1321N1 human astrocytoma cells by novel diterpenoids, scabronines A and G. European Journal of Pharmacology. 370, 79–84 (1999).
Qi, J., Ojika, M. & Sakagami, Y. Termitomycesphins A–D, novel neuritogenic cerebrosides from the edible Chinese mushroom Termitomyces albuminosus. Tetrahedron. 56, 5835–5841 (2000).
Marcotullio, M. C., Pagiotti, R., Maltese, F., Mwankie, G. N. O., Hoshino, T. et al. Cyathane diterpenes from Sarcodon cyrneus and evaluation of their activities of neuritegenesis and nerve growth factor production. Bioorganic and Medicinal Chemistry. 15, 2878–2882 (2007).
Lee, M. K., Xu, Z., Wong, P. C. & Cleveland, D. W. Neurofilaments are obligate heteropolymers in vivo. J. Cell Biol. 122, 1337–1350 (1993).
Liem, R. K. H., Yen, S. H., Salomon G. B. & Shelanski M. L. Intermediate filaments in nervous tissue. J. Cell Biol. 79, 637–645 (1978).
Phan, C. W., Lee, G. S., Hong, S. L., Wong, Y. T., Brkljača, R. et al. Hericium erinaceus (Bull.:Fr) Pers. cultivated in tropical conditions: Isolation of hericenones and demonstration of NGF-mediated neurite outgrowth in PC12 cells via MEK/ERK and PI3K-Akt signaling pathways. Food & Function. 00, 1–3 (2014).
Lai, P. L., Naidu, M., Sabaratnam, V., Wong, K. H., David, R. S. et al. Neurotrophic Properties of the Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Higher Basidiomycetes) from Malaysia. International Journal of Medicinal Mushrooms. 15, 539–554 (2013).
Frade, J. M. & Barde, Y. A. Nerve growth factor: two receptors, multiple functions. Bioessays. 20, 137–145 (1998).
Limpert, A. S., Karlo, J. C. & Landreth, G. E. Nerve growth factor stimulates the concentration of TrkA within lipid rafts and extracellular signal-regulated kinase activation through c-Cbl-associated protein. Molecular and Cellular Biology. 27, 5686–5698 (2007).
Egea, J., Espinet, C., Soler, R. M., Peiro, S., Rocamora, N. et al. Growth factor activation of the extracellular signal-regulated kinase pathway is modulated by Ca2+ and calmodulin. Molecular and Cellular Biology. 20, 1931–1946 (2000).
Koizumi, S., Contreras, M. L., Matsuda, Y., Hama, T., Lazarovici, P. et al. K-252a: A specific inhibitor of the action of nerve growth factor on PC 12 cells. The journal of Neuroscience. 8(2), 715–721 (1988).
Favata, M. F., Horiuchi, K. Y., Manos, E. J., Daulerio, A. J., Stradley, D. A. et al. Identification of a novel inhibitor of mitogen-activated protein kinase kinase. The Journal of Biological Chemistry. 273(29), 18623–18632 (1998).
Kawamata, T., Yamaguchi, T., Shin-ya, K. & Hori, T. Divergence in signaling pathways involved in promotion of cell viability mediated by bFGF, NGF and EGF in PC12 cells. Neurochemical Research. 28(8), 1221–1225 (2003).
Nishina, A., Kimura, H., Sekiguchi, A., Fukumoto, R., Nakajima, S. et al. Lysophosphatidylethanolamine in Grifola frondosa as a neurotrophic activator via activation of MAPK. Journal of Lipid Research. 47, 1434–1443 (2006).
Vikineswary, S. & Chang, S. T. Edible and medicinal mushrooms for sub-health intervention and prevention of lifestyle diseases. Technology Trends, Tech Monitor. Jul-Sept, 33–43 (2013).
Tan, C. S., Ng, S. T., Vikineswary, S., Lo, E. P. & Tee, C. S. Genetic markers for identification of a Malaysian medicinal mushroom Lignosus rhinocerus (Cendawan susu rimau). Acta Hort (ISHS). 859, 161–167 (2010).
Smalheiser, N. R. & Schwartz, N. B. Kinetic analysis of ‘rapid onset’ neurite formation in NG108-15 cells reveals a dual role for substratum-bound laminin. Brain Res. 431(1), 111–121 (1987).
Ojha, A. K., Chandra, K., Ghosh, K. & Islam, S. S. Glucans from the alkaline extract of an edible mushroom, Pleurotus florida, cv Assam Florida: isolation, purification and characterization. Carbohydrate Research. 345, 2157–2163 (2010).
Schimmelpfeng, J., Weibezahn, K. F. & Dertinger, H. Quantification of NGF-dependent neuronal differentiation of PC-12 cells by means of neurofilament-L mRNA expression and neuronal outgrowth. Journal of Neuroscience Methods. 139(2), 299–306 (2004).
Acknowledgements
This research is supported by University of Malaya High Impact Research Grant UM-MoE UM.C/625/1/HIR/MoE/ASH/01 (H-23001-G000008) and UM.C/625/1/HIR/MoE/SC/02 from the Ministry of Education, Malaysia and grants PG110-2012B, J-21001-76536 and UMRG RP005B-13 AFR.
Author information
Authors and Affiliations
Contributions
S.L.-S.S. and L.-F.E. performed the experiments and analysed the data; S.L.-S.S. drafted the manuscript; M.N., P.D. and K.-H.W. conceived and designed the experiments and edited the manuscript. V.S. provided the grant, coordinated and monitored the research and edited the manuscript. All authors reviewed and approved the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Seow, SS., Eik, LF., Naidu, M. et al. Lignosus rhinocerotis (Cooke) Ryvarden mimics the neuritogenic activity of nerve growth factor via MEK/ERK1/2 signaling pathway in PC-12 cells. Sci Rep 5, 16349 (2015). https://doi.org/10.1038/srep16349
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep16349
- Springer Nature Limited
This article is cited by
-
Neuritogenic and in vitro antioxidant activities of Malaysian Gracilaria manilaensis Yamamoto & Trono
Journal of Applied Phycology (2018)
-
Effect of lower chlorinated hydroxylated-polychlorobiphenyls on development of PC12 cells
Environmental Science and Pollution Research (2018)
-
6-shogaol, a neuroactive compound of ginger (jahe gajah) induced neuritogenic activity via NGF responsive pathways in PC-12 cells
BMC Complementary and Alternative Medicine (2017)
-
The potential neuritogenic activity of aqueous extracts from Morchella importuna in rat pheochromocytoma cells
Food Science and Biotechnology (2017)