

Abstract
Bacterial infection or abnormal colonization in the gastrointestinal system is associated with subsets of inflammatory bowel disease and colorectal cancer. Here we demonstrated essential roles of ubiquitin-specific protease 25 (USP25) in experimental colitis, bacterial infections and colon cancer. Knockout or pharmacologic inhibition of USP25 potentiated immune responses after induction of experimental colitis or bacterial infections that promoted clearance of infected bacteria and resolution of inflammation and attenuated Wnt and SOCS3–pSTAT3 signaling, which inhibited colonic tumorigenesis. USP25 levels were positively or negatively correlated with Fusobacterium nucleatum colonization and β-catenin levels or SOCS3 levels in human colorectal tumor biopsies, respectively, and predicted poor prognosis of patients with cancers in the gastrointestinal system. Our findings suggest USP25 as a promoter and druggable target for gastrointestinal infections and cancers.







Similar content being viewed by others


Data availability
The RNA-seq and 16S rRNA-seq data that support the findings of this study have been deposited in the Gene Expression Omnibus under accession codes GSE136727 and GSE143003, which can be found in Supplementary Tables 1 and 4, respectively.
Hhuman GI cancer data were derived from GEPIA (gepia.cancer-pku.cn). The dataset derived from this resource that supports the findings of this study is available at http://gepia.cancer-pku.cn/detail.php?gene=USP25### (cutoff-high, 75%, cutoff-low, 25%; datasets: COAD, READ, STAD). Source data for Figs. 1–7 and Extended Data Fig. 1–8 have been provided as Source Data Figs. 1–7 (numerical source data), Source Data Extended Data Figs. 1–8 (numerical source data), Source Data Figs. 1–3 and 6–7 (uncropped gels) and Source Data Extended Data Figs. 2 and 5 (uncropped gels). All other data supporting the findings of this study are available from the corresponding author upon reasonable request. Source data are provided with this paper.
References
Kaser, A., Zeissig, S. & Blumberg, R. S. Inflammatory bowel disease. Annu. Rev. Immunol. 28, 573–621 (2010).
Somineni, H. K. & Kugathasan, S. The microbiome in patients with inflammatory diseases. Clin. Gastroenterol. Hepatol. 17, 243–255 (2019).
Jostins, L. et al. Host–microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 491, 119–124 (2012).
Hugot, J. P. et al. Association of Nod2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 411, 599–603 (2001).
Ogura, Y. et al. A frameshift mutation in Nod2 associated with susceptibility to Crohn’s disease. Nature 411, 603–606 (2001).
Kobayashi, K. S. et al. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 307, 731–734 (2005).
Maeda, S. et al. Nod2 mutation in Crohn’s disease potentiates NF-κB activity and IL-1β processing. Science 307, 734–738 (2005).
Sokol, H. et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl Acad. Sci. USA 105, 16731–16736 (2008).
Darfeuille-Michaud, A. et al. Presence of adherent Escherichia coli strains in ileal mucosa of patients with Crohn’s disease. Gastroenterology 115, 1405–1413 (1998).
Ohkusa, T. et al. Induction of experimental ulcerative colitis by Fusobacterium varium isolated from colonic mucosa of patients with ulcerative colitis. Gut 52, 79–83 (2003).
Bray, F. et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: Cancer J. Clin. 68, 394–424 (2018).
Lasry, A., Zinger, A. & Ben-Neriah, Y. Inflammatory networks underlying colorectal cancer. Nat. Immunol. 17, 230–240 (2016).
Dow, L. E. et al. APC restoration promotes cellular differentiation and reestablishes crypt homeostasis in colorectal cancer. Cell 161, 1539–1552 (2015).
Greten, F. R. et al. IKKβ links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell 118, 285–296 (2004).
Schwitalla, S. et al. Loss of p53 in enterocytes generates an inflammatory microenvironment enabling invasion and lymph node metastasis of carcinogen-induced colorectal tumors. Cancer Cell 23, 93–106 (2013).
Tauriello, D. V. F. et al. TGFβ drives immune evasion in genetically reconstituted colon cancer metastasis. Nature 554, 538–543 (2018).
Akira, S., Uematsu, S. & Takeuchi, O. Pathogen recognition and innate immunity. Cell 124, 783–801 (2006).
Walsh, D., McCarthy, J., O’Driscoll, C. & Melgar, S. Pattern-recognition receptors–molecular orchestrators of inflammation in inflammatory bowel disease. Cytokine Growth Factor Rev. 24, 91–104 (2013).
Rakoff-Nahoum, S., Paglino, J., Eslami-Varzaneh, F., Edberg, S. & Medzhitov, R. Recognition of commensal microflora by Toll-like receptors is required for intestinal homeostasis. Cell 118, 229–241 (2004).
Fukata, M. et al. Toll-like receptor-4 is required for intestinal response to epithelial injury and limiting bacterial translocation in a murine model of acute colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 288, G1055–G1065 (2005).
Dheer, R. et al. Intestinal epithelial Toll-like receptor 4 signaling affects epithelial function and colonic microbiota and promotes a risk for transmissible colitis. Infect. Immun. 84, 798–810 (2016).
Zhong, B. et al. Ubiquitin-specific protease 25 regulates TLR4-dependent innate immune responses through deubiquitination of the adaptor protein TRAF3. Sci. Signal. 6, ra35 (2013).
Lin, D. et al. Induction of USP25 by viral infection promotes innate antiviral responses by mediating the stabilization of TRAF3 and TRAF6. Proc. Natl Acad. Sci. USA 112, 11324–11329 (2015).
Xu, D. et al. USP25 regulates Wnt signaling by controlling the stability of tankyrases. Genes Dev. 31, 1024–1035 (2017).
Kinchen, J. et al. Structural remodeling of the human colonic mesenchyme in inflammatory bowel disease. Cell 175, 372–386 (2018).
Tabula Muris, C. et al. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 562, 367–372 (2018).
Kitajima, S., Takuma, S. & Morimoto, M. Changes in colonic mucosal permeability in mouse colitis induced with dextran sulfate sodium. Exp. Anim. 48, 137–143 (1999).
Yang, Y. et al. Fusobacterium nucleatum increases proliferation of colorectal cancer cells and tumor development in mice by activating Toll-like receptor 4 signaling to nuclear factor-κB, and up-regulating expression of microRNA-21. Gastroenterology 152, 851–866 (2017).
Abed, J. et al. Fap2 mediates Fusobacterium nucleatum colorectal adenocarcinoma enrichment by binding to tumor-expressed Gal-GalNAc. Cell Host Microbe 20, 215–225 (2016).
Rubinstein, M. R. et al. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 14, 195–206 (2013).
Kostic, A. D. et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 14, 207–215 (2013).
Yu, T. et al. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell 170, 548–563 (2017).
Tomkovich, S. et al. Locoregional effects of microbiota in a preclinical model of colon carcinogenesis. Cancer Res. 77, 2620–2632 (2017).
Suzuki, A. et al. CIS3/SOCS3/SSI3 plays a negative regulatory role in STAT3 activation and intestinal inflammation. J. Exp. Med. 193, 471–481. (2001).
Li, Y. et al. Disease-related expression of the IL6/STAT3/SOCS3 signalling pathway in ulcerative colitis and ulcerative colitis-related carcinogenesis. Gut 59, 227–235 (2010).
Wrigley, J. D. et al. Identification and characterization of dual inhibitors of the USP25/28 deubiquitinating enzyme subfamily. ACS Chem. Biol. 12, 3113–3125 (2017).
Parikh, K. et al. Colonic epithelial cell diversity in health and inflammatory bowel disease. Nature 567, 49–55 (2019).
Schmitt, M. et al. Paneth cells respond to inflammation and contribute to tissue regeneration by acquiring stem-like features through SCF/c-Kit signaling. Cell Rep. 24, 2312–2328 (2018).
Adolph, T. E., Mayr, L., Grabherr, F. & Tilg, H. Paneth cells and their antimicrobials in intestinal immunity. Curr. Pharm. Des. 24, 1121–1129 (2018).
Price, A. E. et al. A map of Toll-like receptor expression in the intestinal epithelium reveals distinct spatial, cell type-specific, and temporal patterns. Immunity 49, 560–575 (2018).
Bersudsky, M. et al. Non-redundant properties of IL-1α and IL-1β during acute colon inflammation in mice. Gut 63, 598–609 (2014).
Hasegawa, M. et al. Protective role of commensals against Clostridium difficile infection via an IL-1β-mediated positive-feedback loop. J. Immunol. 189, 3085–3091 (2012).
Gao, S. J. et al. Interleukin-18 genetic polymorphisms contribute differentially to the susceptibility to Crohn’s disease. World J. Gastroentero. 21, 8711–8722 (2015).
Harrison, O. J. et al. Epithelial-derived IL-18 regulates Th17 cell differentiation and Foxp3(+) Treg cell function in the intestine. Mucosal Immunol. 8, 1226–1236 (2015).
Tak, T., Tesselaar, K., Pillay, J., Borghans, J. A. & Koenderman, L. What’s your age again? Determination of human neutrophil half-lives revisited. J. Leukoc. Biol. 94, 595–601 (2013).
Sylvia, C. J. The role of neutrophil apoptosis in influencing tissue repair. J. Wound Care 12, 13–16 (2003).
Hart, J. Inflammation. 1: its role in the healing of acute wounds. J. Wound Care 11, 205–209 (2002).
Zhao, J., Kim, K. A. & Abo, A. Tipping the balance: modulating the Wnt pathway for tissue repair. Trends Biotechnol. 27, 131–136 (2009).
Bradford, E. M. et al. Epithelial TNF receptor signaling promotes mucosal repair in inflammatory bowel disease. J. Immunol. 199, 1886–1897 (2017).
Isomoto, H. et al. Sustained IL-6/STAT-3 signaling in cholangiocarcinoma cells due to SOCS-3 epigenetic silencing. Gastroenterology 132, 384–396 (2007).
Carow, B. & Rottenberg, M. E. SOCS3, a major regulator of infection and inflammation. Front. Immunol. 5, 58 (2014).
Brennan, C. A. & Garrett, W. S. Gut microbiota, inflammation, and colorectal cancer. Annu. Rev. Microbiol. 70, 395–411 (2016).
Zhong, B. et al. Negative regulation of IL-17-mediated signaling and inflammation by the ubiquitin-specific protease USP25. Nat. Immunol. 13, 1110–1117 (2012).
Zhao, Y. et al. USP2a supports metastasis by tuning TGF-β signaling. Cell Rep. 22, 2442–2454 (2018).
Yang, X. O. et al. Regulation of inflammatory responses by IL-17F. J. Exp. Med. 205, 1063–1075 (2008).
Li, S. et al. Pathogen blocks host death receptor signalling by arginine GlcNAcylation of death domains. Nature 501, 242–246 (2013).
Zhang, M. et al. USP18 recruits USP20 to promote innate antiviral response through deubiquitinating STING/MITA. Cell Res. 26, 1302–1319 (2016).
Liuyu, T. et al. Induction of OTUD4 by viral infection promotes antiviral responses through deubiquitinating and stabilizing MAVS. Cell Res. 29, 67–79 (2019).
Acknowledgements
We thank Y. Pan (Fourth Military Medical University) for valuable suggestions, S. Li (Huazhong Agricultural University) for reagents and technical help and members of the Zhong laboratory and the core facilities of the Medical Research Institute for technical help. This study was supported by grants from the National Key Research and Development Program of China (2018TFE0204500 and 2018YFC1004601), Natural Science Foundation of China (31671454 and 31930040), Fundamental Research Funds for Central Universities (2042020kf0207 and 2042020kf0042), Natural Science Foundation of Hubei Province (2018CFA016) and Medical Science Advancement Program (Basic Medical Sciences) of Wuhan University (TFJC2018004).
Author information
Authors and Affiliations
Contributions
B.Z. designed and supervised the study; X.-M.W. performed the core experiments; C.Y. and Y.Z. helped with mouse breeding and colon cancer modeling; Z.X. and P.W. performed bioinformatics analysis; Z.-G.X., W.Y., D.L. and B.X. collected CRC samples, produced tissue arrays and performed IHC analysis; J.-Y.F. and C.D. provided reagents; B.Z., X.-M.W. and D.L. wrote the paper; all the authors analyzed data.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 USP25 deficiency inhibits DSS-induced colitis independently of type I IFN signaling.
(a, b) Body weight change (a) and gross morphological change and lengths of colons (b) of Usp25-/-Ifnar1-/- mice (n = 3) and Ifnar1-/- mice (n = 3) that were given 2% DSS in drinking water for 5 d, followed by normal drinking water for another 4 d. (c) Representative histological images (left) and pathological scores (right) of H&E stained colon sections of Usp25-/-Ifnar1-/- mice (n = 3) and Ifnar1-/- mice (n = 3) in (a). (d–f) Total cell number (d), representative flow cytometry images and analysis of the percentages and cell number of neutrophils (CD11b+Gr-1+) and macrophages (CD11b+F4/80+) (E), CD8+, CD4+, CD3+CD4+IFNγ+, CD3+CD4+IL−17γ+, and CD3+CD4+Foxp3+ cells (f) from lamina propria in Usp25+/+ (n = 3) and Usp25-/- (n = 3) mice that were given 2.5% DSS in drinking water for 5 d, followed by normal drinking water for another 3 d. (g–i) Total cell number (g), representative flow cytometry images and analysis of the percentages and cell number of neutrophil (CD11b+Gr-1+) and macrophage (CD11b+F4/80+) (h), CD8+, CD4+, CD3+CD4+IFNγ+, CD3+CD4+IL-17γ+, and CD3+CD4+Foxp3+ cells (i) of lamina propria from Usp25-/-Ifnar1-/- mice (n = 3) and Ifnar1-/- mice (n = 3) that were given 2% DSS in drinking water for 5 d, followed by normal drinking water for another 3 d. *P < 0.05; **P < 0.01 (two-tailed student’s t-test). Scale bars represent 0.4 mm. Data are representative of two independent experiments (Graphs show mean ± S.D.). Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.1.
Extended Data Fig. 2 USP25 deficiency in hematopoietic cells does not affect immune cell homeostasis or differentiation in the colon lamina propria after DSS treatment.
(a) Flow cytometry analysis of the blood from irradiated CD45.1+ mice that were adoptively transferred Usp25+/+ (n = 5) and Usp25-/- (n = 5) bone marrow cells (1×106) (CD45.2+) for 6 weeks. (b–d) Total cell number (b), representative flow cytometry images and analysis of the percentages and cell number of neutrophil (CD11b+Gr-1+) and macrophage (CD11b+F4/80+) (c), CD8+, CD4+, CD3+CD4+IFNγ+, CD3+CD4+IL-17γ+, and CD3+CD4+Foxp3+ cells (d) of lamina propria from mice in (a) that were given 2.5% DSS in drinking water for 5 d, followed by normal drinking water for another 3 d. (e) qRT-PCR analysis of Usp25 mRNA levels (left graph) (n = 4 for each time point) and immunoblot analysis (n = 2 for each time point) of USP25 protein in colon epithelial cells isolated from wild-type mice treated with 2.5% DSS for 0–4 d. Data are representative of two independent experiments (Graphs show mean ± S.D., two-tailed student’s t-test.). Uncropped immunoblot images are shown in Uncropped_gels_Extended_Data_Fig.2. Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.2.
Extended Data Fig. 3 USP25 deficiency inhibits DSS-induced colitis dependently on gut bacteria.
(a) Gene set enrichment analysis of IRF-dependent genes of Usp25+/+ (n = 2) and Usp25-/- (n = 2) mice that were given 2.5% DSS in drinking water for 5 d, followed by normal drinking water for another 2 d. (b) qRT-PCR analysis of IRF-dependent genes in the colons of Usp25+/+ (n = 2 at day 0, n = 4 at day 4 and 6) and Usp25-/- (n = 2 at day 0, n = 4 at day 4 and 6) mice that were given 2.5% DSS in drinking water for 4 or 6 d. (c) qRT-PCR analysis of the indicated genes in the colons from Usp25-/-Ifnar1-/- (n = 3) and Ifnar1-/- (n = 3) mice that were given 2% DSS in drinking water for 5 d, followed by normal drinking water for another 3 d. (d) A scheme of antibiotics treatment and colitis induction. Mice were fed with sterile water containing ampicillin (1 g/L), metronidazole (1 g/L), vancomycin (0.5 g/L), and neomycin (1 g/L). Four weeks later, the mice were used for 2% DSS-colitis analysis. (e) Survival of Usp25+/+ (n = 6) and Usp25-/- (n = 8) mice treated as in (d). (f, g) Body weight change (f), gross morphological change and lengths of colons, and representative H&E stained colon sections (g) of Usp25+/+ (n = 7) and Usp25-/- (n = 6) mice that were treated as in (d). (h) qRT-PCR analysis of the indicated genes in the colons of Usp25+/+ (n = 3) and Usp25-/- (n = 3) mice that were treated as in (d). *P < 0.05; **P < 0.01; ***P < 0.001 (two-tailed student’s t-test). Scale bars represent 0.4 mm. Data are representative of two (b, c, h) and combined two (e–g) independent experiments (Graphs show mean ± S.D.). Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.3.
Extended Data Fig. 4 USP25 deficiency potentiates bacterial infection-induced expression of cytokines.
(a) Image (left) and cell number (right) of spleens from Usp25+/+ (n = 4) and Usp25-/- (n = 4) mice that were orally infected with C rodentium (1 ×109 CFU) for 14 days. (b, c) Flow cytometry analysis of spleenocytes strained with fluorescence-conjugated antibodies against the indicated surface from mice in (a) (n = 4). (d, e) qRT-PCR analysis of the expression of the indicated genes in the colons from Usp25+/+ (n = 3) and Usp25-/- (n = 3) mice that were orally infected with C rodentium (1 ×109 CFU) for 14 days (d) (n = 3 for Usp25+/+; n = 4 for Usp25-/-) or S typhimurium (1 ×107 CFU) for 6 days (e) (n = 5). (f, g) qRT-PCR analysis of the expression of the indicated genes in Usp25+/+ and Usp25-/- colon organoids (n = 3, technical replicates) that were infected with C rodentium and S typhimurium (CFU of 5×106 or 1×107) for 1 h (f) or stimulated with LPS (10 mg/ml) for 0–8 h (g). (h) Immunohistochemistry analysis (IHC) of USP25 and fluorescent in situ hybridization (FISH) of F. nucleatum in human colorectal cancer tissues (n = 18 for the + or - groups). (i) Correlation analysis (left) and representative images (right) of USP25 protein levels (IHC) and F. nucleatum colonization (FISH) in CRC tissues (n = 18). Scale bars represent 0.2 mm. (j) qRT-PCR analysis of the expression of USP25 in the colons from Usp25+/+mice day (n = 4 for each time point) that were orally infected with C rodentium (1 ×109 CFU) at 0, 5, 10 day. (k) qRT-PCR analysis of the expression of USP25 in the colons from Usp25+/+mice day (n = 4 for each time point) that were orally infected S typhimurium (1 ×107 CFU) at 0, 2, 4. (l) Representative images and tumor counts of colons from ApcMin/+ (n = 10) and ApcMin/+ Usp25-/- (n = 7) mice that were daily fed with F. nucletum (1 ×108 CFU) for 8 weeks beginning at 6 weeks of age. (m, n) FISH (m) and statistical (n) (n = 10 mice for ApcMin/+; n = 3 mice for ApcMin/+ Usp25-/-) analysis of F. nucleatum of colon tumors from mice in (l). Black scale bars represent 2 mm. Red scale bars represent 0.1 mm. *P < 0.05; **P < 0.01; ***P < 0.001 (two-way ANOVA for j and k; two-tailed student’s t-test for a–e, h, l and n). Data are representative of two independent experiments (Graphs show mean in f and g or mean ± S.D. in a–e, h, j–n). Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.4.
Extended Data Fig. 5 Negative correlation of USP25 expression level and prognosis of gastrointestinal cancers.
(a) The correlation of USP25 transcript expression with overall survival in gastrointestinal cancer patients (n = 187 for USP25 high and low patients, GEPIA database). (b) Immunoblot analysis of USP25 in the colon tumors from Usp25+/+ mice after AOM/DSS induction for 12 weeks (left), Vil-Cre;Trp53fl/fl mice at the 20th week after the first AOM injection (middle), or ApcMin/+ mice at 20-week-old (right). (c) qRT-PCR analysis of the expression of the indicated genes in the colon from Usp25+/+ (n = 5) and Usp25-/- (n = 5) mice after AOM/DSS induction for 9 weeks. (d-f) qRT-PCR analysis of the expression of the indicated genes in the colon tumors from Usp25+/+ (n = 5) and Usp25-/- (n = 5) mice after AOM/DSS induction (d), Vil-Cre;Trp53fl/fl (n = 6) and Vil-Cre;Trp53fl/flUsp25-/- (n = 6) mice at the 18th week after the first AOM injection (e), or ApcMin/+ (n = 5) and ApcMin/+Usp25-/- (n = 5) mice at 5-month-old (f). (g) qRT-PCR of the Wnt signaling genes in the Usp25+/+ (n = 3, technical replicates) and Usp25-/- (n = 3, technical replicates) colon organoids stimulated with Wnt3a (50 ng/ml) for 4 h. (h) Immunoblot analysis of the indicated proteins in Usp25+/+ (n = 3, technical replicates) and Usp25-/- (n = 3, technical replicates) colon organoids that were left untreated or stimulated with Wnt3a (50 ng/ml) for 4 h. (i) IHC images and the intensity quantification of Axin2, SOCS3, pStat3 and TNKS proteins in small intestine tumors from ApcMin/+ (n = 6) and ApcMin/+Usp25-/- (n = 6) mice at 5-month-old. (j, k) Immunoblot analysis and the intensity quantification of the indicated proteins in tumors from Usp25+/+ (n = 6) and Usp25-/- (n = 6) mice after AOM/DSS induction (j), or Vil-Cre;Trp53fl/fl (n = 5) and Vil-Cre;Trp53fl/flUsp25-/- (n = 5) mice at the 18th week after the first AOM injection (k). *P < 0.05; **P < 0.01; ***P < 0.001 (two-way ANOVA for g; two-tailed student’s t-test for c–f, i, j, k). Red and black scale bars represent 50 mm and 1 mm, respectively. Data are representative of at least three independent experiments (Graphs show mean in g or mean ± S.D. in c–f and i–k). Uncropped immunoblot images are shown in Uncropped_gels_Extended_Data_Fig.5. Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.5.
Extended Data Fig. 6 Synthesis and toxicity analysis of AZ1.
(a, b) IHC images of USP25, b-Catenin, SOCS3 and pSTAT3 proteins (a) in patient CRC tissues (n = 36) and correlation analysis of USP25 with b-Catenin, SOCS3 and pSTAT3 proteins expression level (b). (c, d) The synthetic method (c) and HPLC chromatography (d) of the inhibitor AZ1. (e, f) Weight change (e) and images of spleens and lymph nodes (f) of 7-week-old male Usp25+/+ (n = 5) and Usp25-/- (n = 5) mice that were daily injected of PBS or AZ1 (40 mg/kg body weight) by gavage. Scale bars represent 0.4 mm. Data are representative of two experiments (a, e, f) (Graphs show mean ± S.D., two-tailed student’s t-test). Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.6.
Extended Data Fig. 7 Selectivity analysis of AZ1 on USP25.
(a, b) qRT-PCR analysis of the expression of the indicated genes in Usp25+/+ (n = 3, technical replicates) and Usp25-/- (n = 3, technical replicates) colon organoids infected with C. rodentium (a) or treated with Wnt3a (50 ng/ml) (b) in the presence or absence of AZ1 (10 mM) stimulation for 4 h. (c) Growth rate (as indicated by the OD600 values) of different bacteria that were inoculated to fresh LB culture medium (1:200) for the indicated time points after overnight culture (n = 3, technical replicates). (d) Shannon diversity (left), bacterial relative abundance (middle) and PCoA analysis from 16 S rRNA seq in feces of WT mice fed with PBS (n = 5 mice) or AZ1 (n = 5 mice) for 15 days. (e) Shannon diversity (left), bacterial relative abundance (middle) and PCoA analysis from 16 S rRNA seq in feces of WT mice treated by 2.5% DSS for 5 d followed by normal water for 2d in the presence of PBS (n = 4 mice) or AZ1 (n = 5 mice) by gavage. (f, g) Body weight change (f) and gross morphological change and lengths of colons (g) of Usp25-/- mice that were given 2.5% DSS in drinking water for 5 d, followed by normal drinking water for another 2 d in the presence or absence of seven successive daily gavage of PBS (n = 8 mice) or AZ1 (40 mg/kg) (n = 8 mice). (h, i) Weight change (h) and image and colon lengths (i) of Il10-/- mice (8-week old) housed in conventional open cage conditions and injected with PBS (n = 9 mice) or AZ1 (40 mg/kg) (n = 11 mice) by gavage every three days starting from 12-week old for 8 successive weeks. (j, k) qRT-PCR of the distal colon tissues (j) (n = 5 mice for PBS and AZ1 groups) and HE staining of colon (g) (PBS, n = 9 mice; AZ1, n = 11 mice) of Il10-/- mice treated as in (h). *P < 0.05; **P < 0.01; ***P < 0.001. (two-tailed Student’s t-test). Scale bars represent 300 μm. Data are representative of three (a–e) or two (j) independent experiments or combined two independent experiments (f–i) (Graphs show mean in a-c or mean ± S.D. in d-j). Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.7.
Extended Data Fig. 8 AZ1 has minimal effect on colon tumorigenesis in the USP25 deficient background.
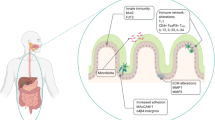
(a) A scheme of colon tumor induction in Vil-Cre;Trp53fl/flUsp25-/- mice (upper). Representative images and tumor counts of colons from Vil-Cre;Trp53fl/flUsp25-/- mice that were weekly injected with AOM (10 mg/kg) for six successive weeks, followed by injection of PBS (n = 6 mice) or AZ1 (20 mg/kg) (n = 6 mice) by gavage every three days from 12th to 18th weeks after initial AOM injection (lower). (b) qRT-PCR of the indicated genes in colon tumors from Vil-Cre;Trp53fl/flUsp25-/- mice treated in (a) (n = 3 mice for PBS and AZ1 groups). (c) A model for USP25 to modulate colonic infections, inflammations and colon cancer. USP25 inhibits the expression of anti-bacteria genes, promotes bacterial infections and inflammation in the colon, downregulates SOCS3 to facilitate activation of STAT3, and promotes Wnt signaling to increase the stemness of colonic epithelial cells, which are inducers of colon cancer. Pharmacologically inhibition of USP25 impairs inflammation, restricts bacterial infections and inhibits tumorigenesis in the colon. Two-tailed student’s t-test. Data are combination (a) or representative (b) of two independent experiments (Graph show mean ± S.D.). Numerical source data for the experiments in this figure can be found in Numerical_Source_Data_Extended_Data_Fig.8.
Supplementary information
Source data
Source Data Fig. 1
Numerical source data.
Source Data Fig. 1
Uncropped gels.
Source Data Fig. 2
Numerical source data.
Source Data Fig. 2
Uncropped gels.
Source Data Fig. 3
Numerical source data.
Source Data Fig. 3
Uncropped gels.
Source Data Fig. 4
Numerical source data.
Source Data Fig. 5
Numerical source data.
Source Data Fig. 6
Numerical source data.
Source Data Fig. 6
Uncropped gels.
Source Data Fig. 7
Numerical source data.
Source Data Fig. 7
Uncropped gels.
Source Data Extended Data Fig. 1
Numerical source data.
Source Data Extended Data Fig. 2
Numerical source data.
Source Data Extended Data Fig. 2
Uncropped gels.
Source Data Extended Data Fig. 3
Numerical source data.
Source Data Extended Data Fig. 4
Numerical source data.
Source Data Extended Data Fig. 5
Numerical source data.
Source Data Extended Data Fig. 5
Uncropped gels.
Source Data Extended Data Fig. 6
Numerical source data.
Source Data Extended Data Fig. 7
Numerical source data.
Source Data Extended Data Fig. 8
Numerical source data.
Rights and permissions
About this article

Cite this article
Wang, XM., Yang, C., Zhao, Y. et al. The deubiquitinase USP25 supports colonic inflammation and bacterial infection and promotes colorectal cancer. Nat Cancer 1, 811–825 (2020). https://doi.org/10.1038/s43018-020-0089-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s43018-020-0089-4
- Springer Nature America, Inc.
This article is cited by
-
USP54 is a potential therapeutic target in castration-resistant prostate cancer
BMC Urology (2024)
-
Targeting ubiquitin specific proteases (USPs) in cancer immunotherapy: from basic research to preclinical application
Journal of Experimental & Clinical Cancer Research (2023)
-
Ubiquitin-specific peptidase 25 exacerbated osteoarthritis progression through facilitating TXNIP ubiquitination and NLRP3 inflammasome activation
Journal of Orthopaedic Surgery and Research (2023)
-
Statistical modeling of gut microbiota for personalized health status monitoring
Microbiome (2023)
-
p53 promotes peroxisomal fatty acid β-oxidation to repress purine biosynthesis and mediate tumor suppression
Cell Death & Disease (2023)