
Abstract
Post-transcriptional autoregulation of gene expression is common in bacteria but many fewer examples are known in eukaryotes. We used the yeast collection of genes fused to GFP as a rapid screen for examples of feedback regulation in ribosomal proteins by overexpressing a non-regulatable version of a gene and observing the effects on the expression of the GFP-fused version. We tested 95 ribosomal protein genes and found a wide continuum of effects, with 30% showing at least a 3-fold reduction in expression. Two genes, RPS22B and RPL1B, showed over a 10-fold repression. In both cases the cis-regulatory segment resides in the 5’ UTR of the gene as shown by placing that segment of the mRNA upstream of GFP alone and demonstrating it is sufficient to cause repression of GFP when the protein is over-expressed. Further analyses showed that the intron in the 5’ UTR of RPS22B is required for regulation, presumably because the protein inhibits splicing that is necessary for translation. The 5’ UTR of RPL1B contains a sequence and structure motif that is conserved in the binding sites of Rpl1 orthologs from bacteria to mammals, and mutations within the motif eliminate repression.
Similar content being viewed by others

Introduction
Feedback regulation is common in many biological processes. In metabolic pathways the end product can often inhibit one of the enzymes in the pathway to set the flux through the pathway at the appropriate level1,2. Gene expression is also often regulated by feedback mechanisms. This commonly occurs with a transcription factor that regulates its own expression, referred to as autoregulation. One of the first transcription factors to be studied in detail, the lambda repressor, was found to be autoregulated both positively and negatively, allowing it to maintain its in vivo concentration in a narrow range3,4. Autoregulation has been found to be among the most common network motifs in bacterial transcription5,6. Studies on the regulatory network in yeast also identify many examples of autoregulation7,8,9. Mathematical analyses have characterized the properties and advantages of autoregulatory networks10,11,12.
Although less well-studied, autoregulation also occurs for proteins involved in post-transcriptional steps of gene expression. For example, many splicing factors regulate their own expression13,14,15,16,17. Recently developed methods for high-throughput analysis of RNA-protein interactions have identified many RNA-binding proteins, some of which are associated with their own mRNAs18,19,20,21,22,23,24,25. By itself that does not prove they are autoregulatory, but it seems likely to be a consequence of such binding. In most of those cases, both transcription factors and RNA-binding proteins, the normal function of the protein is to bind DNA or RNA and often to regulate gene expression. The fact that they can regulate their own expression is not surprising given the advantages of such feedback processes, it only requires that the gene’s own DNA or RNA be included in the target list for the protein. Binding sites are often short and, because unconstrained nucleic acids can evolve rapidly, sites that offer a selective advantage are likely to be obtained through random mutagenesis processes.
There are also examples of proteins whose primary function is not in gene regulation but that have a secondary role in regulating their own expression. Many of these are proteins that bind to RNA, but whose primary functions are not involved in controlling gene expression. For example, most of the ribosomal proteins in E. coli are subject to feedback regulation26,27. Ribosomal proteins are expressed as part of transcription units (operons) composed of other ribosomal proteins. Autoregulation by one of the proteins in the operon is typically sufficient to control the expression of all the other genes in the operon by translational coupling, where translation of an upstream gene in an operon is required for translation of the downstream genes. The ribosomal proteins are all RNA-binding proteins, having as their primary target the rRNAs of the ribosome. To become autoregulatory, the mRNA simply has to evolve a sequence that is a molecular mimic of the primary target site, but with lower affinity so that binding of the rRNA is saturated before the regulatory site becomes bound by the protein27,28,29,30. There are also examples of tRNA synthetase genes in bacteria and yeast that have evolved a similar regulatory site, where the mRNA mimics the tRNA that the synthetase gene normally binds to, but with lower affinity31,32,33,34,35. Particularly interesting are cases where the protein’s normal function does not involve binding to RNA but it is found in screens for RNA-binding proteins36,37. In some cases, proteins with alternative primary functions have been shown to be direct regulators of their own translation38,39,40,41,42. Such examples highlight the enormous functional capacity of RNA where it can become a sensor of the cellular environment and autonomously regulate its own fate, as exemplified in cases where no protein is required, such as riboswitches43,44,45,46,47. When the effector being recognized by the mRNA is its own gene product, the result is autoregulation of gene expression.
The fundamental characteristic of feedback regulation of gene expression is that if the activity of a gene product, which is usually proportional to its concentration, is higher than the set point, or “desired” level for the cell, then its expression is reduced, and conversely, if the activity is too low, the expression is increased. This relationship is true regardless of the mechanism by which the feedback regulation occurs, whether it involves a complex network of interactions or is simply the result of direct autoregulation by the gene product itself. Once examples of feedback regulation of gene expression are obtained, the mechanism can be determined by additional experiments. The collection of yeast strains with genes fused to green fluorescent protein (GFP)48 provides an excellent resource to screen for examples of feedback regulation. By introducing into those strains an inducible copy of a gene for the same protein, but lacking all potential cis-regulatory elements (cre-less), an observed reduction in the level of GFP after induction indicates some feedback mechanism controlling the expression of the wild-type gene. Further analyses are required to determine the step in the expression process that is being regulated, whether it is transcription initiation, any of the processes leading to the mature mRNA, any step in translation, or even post-translational enhancement of protein degradation. We are particularly interested in finding new examples of post-transcriptional autoregulation of protein expression so our initial focus is on ribosomal proteins, which are commonly translationally autoregulated in bacteria but27,28,49,50 for which many fewer examples are known in yeast. Ribosome synthesis in yeast is subject to feedback regulation in part by alternative functions of ribosomal proteins51,52,53,54,55. There are several examples of post-transcriptional autoregulation by yeast ribosomal proteins, most often through inhibition of splicing necessary for protein expression. For example, RPL22B, RPS14B, and RPL30 all have introns within the N-terminus of the coding sequence and splicing is inhibited by binding of the encoded protein56,57,58,59. Remarkably, the ortholog of Rpl30 in the archaeon Sulfolobus acidocarldarius can bind to the same mRNA target and inhibit splicing60. RPS9A and RPS9B both have introns within the N-terminus of the coding region and both genes are subject to feedback regulation by inhibition of splicing61. The orthologs of Rps9 are involved in autoregulation in several other eukaryotic species and even in bacteria27,61. RPS28B does not contain an intron but is autoregulated by a different mechanism where binding of the Edc3 decapping enzyme to the 3′ UTR is regulated by the Rps28 protein, leading to mRNA degradation62,63. These cases are all consistent with examples from bacteria where ribosomal protein synthesis is regulated post-transcriptionally, and it seems likely that a directed search for feedback regulation among yeast ribosomal protein genes could uncover more examples, leading us to utilize the yeast GFP-fusion collection.
Results
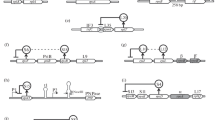
Feedback regulation of protein expression requires that when the activity of the protein, usually proportional to its concentration, is higher than the homeostasis point of the cell, its expression is reduced, and when the activity is lower than that point, its expression is increased. This allows the cell to maintain expression in a narrow range around its set point. The collection of yeast genes fused to GFP48 provides an excellent resource to screen for genes that exhibit feedback regulation. A version of the gene is synthesized that is lacking any potential cis-regulatory elements (a cre-less version of the gene), under the control of an inducible promoter. The cre-less version is synthesized with alternative 5′ and 3′ UTRs, mCherry is fused to the C-terminus in place of the GFP of the wild-type gene, any introns are removed, and the codons of the gene are shuffled64 to maintain the wild-type protein sequence while altering the mRNA sequence sufficiently that we expect any cis-regulatory elements that overlap the coding sequence would be eliminated. We use the GAL1 promoter for induction of the cre-less gene (Fig. 1a). If there is feedback regulation, overexpression of the cre-less gene (monitored by mCherry fluorescence) will lead to a decrease in the expression of the wildtype gene, which is monitored by GFP fluorescence. Identification of feedback regulated genes does not provide information about the mechanism of action, and further analyses are required to determine the step in the expression process that is regulated. The scheme for gene synthesis and fluorescent detection is summarized in Supplementary Fig. S1.

a One endogenous gene is fused to GFP and a cre-less (cis-regulatory element-less) version of the same gene is fused to mCherry. The cre-less version lacks the native 5′ and 3′ UTRs and any introns and has shuffled codons to eliminate potential regulatory sites in the cre-less mRNA. After inducing the cre-less gene, possible autoregulation is detected by a decrease in GFP levels. b An example of autoregulation with the gene PDC1. Overexpression of the cre-less PDC1 (seen as an increase in mCherry fluorescence) leads to a decrease of the endogenous PDC1 (seen as a decrease in GFP fluorescence).
The pyruvate decarboxylase gene PDC1 is transcriptionally autoregulated65. To test our strategy, we synthesized a cre-less copy of PDC1, fused to mCherry and under control of the GAL1 promoter. Figure 1b shows the change in the fluorescence signal of both mCherry and GFP after 10 h of induction, when the GFP signal is reduced about 10-fold. Note that there is a subset of cells that do not induce mCherry fluorescence, which results in the shoulder seen on the GFP fluorescence signal. To simplify measurements of the change in expression, in all further examples we use the median of the log of the GFP measurement between the two conditions, which may underestimate the true magnitude of the effect.
Screen of ribosomal proteins
We are primarily interested in identifying cases of post-transcriptional autoregulation, examples of which are common among ribosomal genes in bacteria27,29,30,50. Among the known examples of yeast ribosomal genes that are post-transcriptionally autoregulated, repressing proper splicing of the pre-mRNA is often the mechanism56,57,58,59,60,61. Many yeast ribosomal proteins have paralogs which are identical, or nearly so, to each other and for those cases it is sufficient to make a cre-less version from only one of the paralogs and to test the effects on expression of both wild-type (with GFP-fusion) paralogs. GFP expression of 60 large subunit ribosomal protein genes, including 25 paralogous pairs, both with and without induction of the cre-less gene, are shown in Fig. 2a, b shows the same for 35 small subunit ribosomal protein genes, including 16 paralogous pairs, and the fluorescence signal from control cells lacking a GFP-fusion gene. Shown are the median of the log (fluorescence) values of two or more measurements. Figure 2c shows both the induced and uninduced measurements in the same plot for all genes (data available in Supplementary Data 1). Two genes, RPL1B and RPS22B, have greater than 10-fold decrease in expression after induction (marked with ** and above the 1-log dotted line), and another 28 genes have greater than 3-fold decrease in expression (marked with *). Cases where both paralogs are decreased by similar amounts may represent examples of increased rates of protein degradation of the overexpressed proteins51,52. In ten cases (24% of paralogous pairs) one paralog is decreased in expression by greater than 2-fold compared to the other paralog (marked with #) and those are the most likely candidates for gene specific repression. Of the known examples described above, we see a large reduction in expression of RPL22B. We did not see a large reduction in expression for RPS14B, but it is expressed at very low levels, consistent with previous reports that the ratio of RSP14A to RSP14B is 10:156. We also did not see a large reduction in expression of RPS28B, likely because its regulation requires the 3′ UTR which is disrupted in the GFP-fusion genes62,63. This indicates one limitation of our approach, that the GFP fusion to the C-terminus of the protein disrupts the normal 3′ UTR, and cis-regulatory elements residing in that region will likely be missed. RPL30 and RPS9A/B are missing from our GFP collection and could not be tested. The two genes we observe with the largest effects, RPS22B and RPL1B, have not, to our knowledge, been previously shown to be autoregulated, nor have most of the regulated examples we observe.

a Sixty large ribosomal subunit proteins were screened for expression changes. Dots show the log10 expression level for each biological replicate, and the bars indicate the mean values, for both induced and uninduced conditions. Genes with more than a 10-fold reduction of their endogenous protein after induction of their cre-less protein are highlighted with two stars (**). Genes with a 3-fold to 10-fold reduction are highlighted by one star (*). Genes with one paralog effected by >2-fold more than the other paralog are highlighted with #. b Thirty-five small ribosomal subunit proteins were screened for expression changes. c Scatterplot showing the uninduced vs. induced log10 GFP levels for large ribosomal subunit proteins (triangles) and small ribosomal subunit proteins (circles). RPL1B (red circle) and RPS22B (orange triangle) showed the highest levels of autoregulation. The black line passing through the center represents equal GFP levels for induced and uninduced, while the orange dotted line represents a 10-fold reduction in the induced compared to uninduced. The black triangle is BAC177 with no GFP-tagged proteins.
It is possible that changes in expression could be influenced by the change in media required for induction, with 0.2% galactose added to the existing 2% raffinose as the carbon source. To test that we measured expression of each GFP strain, but without the cre-less gene plasmid, in both conditions (data in Supplementary Data 2). The results show very little change in GFP in inducing conditions, and in fact it is in the wrong direction to contribute to gene repression. After induction the log-GFP levels rose by a mean 0.06 (with a standard deviation of 0.06).
Autoregulation of RPS22B
Upon induction of the cre-less version of RPS22 (the two paralogs code for identical proteins), the expression of wild-type RPS22B (with GFP fusion) is reduced over 10-fold at 10 h while RPS22A has only a modest reduction (Fig. 2b). The RPS22A gene has no introns whereas the RPS22B gene has two introns, including one in the 5′ UTR that contains a conserved, predicted secondary structure66. That intron contains seven AUG codons and is a substrate for RNase III-mediated cleavage if not spliced67, both of which suggest that splicing is required for translation of the mRNA. In fact, deletion of the 5′ UTR intron increases expression of the RPS22B gene several-fold68. The simplest hypothesis is that the 5′ UTR intron of the RPS22B gene is the cis-regulatory site required for autoregulation, probably via inhibition of splicing. To test that we integrated the gene for GFP, expressed from the constitutive TEF2 promoter whose activity is unaffected by galactose induction69, into the yeast chromosome in place of a putative gene of unknown function (chromosome II: YBR032W). Two different 5′ UTRs were placed upstream of GFP, the complete RPS22B UTR and a “post-spliced” RSP22B UTR with the intron removed. Both strains were transformed with the plasmid containing the cre-less version of RPS22B (Fig. 3a). GFP fluorescence was measured for each strain, with and without the plasmid, and with and without induction (Fig. 3b, c, Supplementary Data 3). The strain with the spliced 5′ UTR showed nearly identical expression with or without the plasmid and with or without induction (Fig. 3b, c, blue traces), indicating that translation of the spliced mRNA is not repressed. In the strain with the complete RPS22B 5′ UTR, GFP expression is nearly 3-fold lower in cells without the plasmid and with the plasmid but without induction. We expect this is due to repression by the endogenous Rps22 protein in the cells. When the cre-less plasmid is induced to overexpress Rps22, the expression of GFP is further reduced about 6-fold (Fig. 3b, c, green traces), similar to the repression of the wild-type RPS22B-GFP strain (Fig. 2b). This indicates that, similar to several other autoregulated ribosomal protein genes, repression occurs by inhibiting splicing. Interestingly, the bacterial ortholog of Rps22 is S870 and it is also involved in autoregulation49. One anomaly is worth noting. The original cre-less version of Rps22 had unintentionally left the stop codon at the end of the gene, so that it was not fused with mCherry, but it showed the autoregulation. When that was corrected to make a cre-less version fused to mCherry, autoregulation was no longer observed, suggesting interference with RNA binding by the mCherry fusion. It is possible the same thing happens with other of our mCherry fusion constructs, which is another reason we could be underestimating the true number of autoregulated ribosomal protein genes.

a Two reporter constructs were made to test the sequence requirements for autoregulation by overexpressed cre-less Rps22. One construct had the wild-type RPS22B 5’ UTR (containing a 557 bp intron) placed upstream of GFP and driven by the TEF2 promoter. A second construct was identical except that the intron was removed. Both constructs were separately integrated into the YBR032W locus and transformed with the plasmid for expressing cre-less Rps22. b Histograms of log10 GFP intensity from flow cytometry of cells containing either the UTR-with-intron reporter (green) or the UTR-without-intron reporter (blue). Cells were measured as either uninduced (lighter-shaded colors) or after induction of cre-less Rps22 (darker-shaded colors). Each histogram curve represents cells picked from a single colony after transformation of the cre-less plasmid. c Bar plot comparing the log GFP levels of the two reporter constructs in cells with or without the plasmid for expression of cre-less Rps22 and with or without galactose induction. Circles represent the measurements of individual transformants and the bar is the mean, also listed in the table below.
Autoregulation of RPL1B
The expression of the RPL1B gene is reduced over 10-fold after 10 h of induction of the cre-less version (Fig. 2a). The paralog RPL1A showed less than a 2-fold reduction in expression (Fig. 2a). RPL1B has no introns and we surmised that the 5′ UTR may be the regulatory region. The gene has a short 5′ UTR of 64 bases that is highly conserved within the senso stricto yeast species71 (Fig. 4a). We place the full 5′ UTR before the chromosomal GFP gene (as we did for the RPS22B UTR described above) and measured about a 10-fold reduction in expression after induction of the cre-less gene (Supplementary Fig. S2, green graphs). This indicates that the 5′ UTR is sufficient to confer feedback regulation by Rpl1. We also tested different versions of the gene on the plasmid fused to mCherry. Besides the cre-less version (RPL1B-cl), we tested three alternative constructs (Supplementary Fig. S3): wild-type for both the 5′ UTR and coding region (RPL1B-wt); wild-type for the 5′ UTR with a shuffled coding region (RPL1B-cd); and mutant 5′ UTR but wild-type coding region (RPL1B-mt). After induction the two constructs containing the wild-type 5′ UTR (RPL1B-wt and RPL1B-cd) produced about 2-fold less mCherry fusion protein than those with the mutant 5′ UTR (RPL1B-mt and RPL1B-cl), consistent with the regulatory element being within the 5′ UTR (Supplementary Fig. S4). Although the effect is smaller, the constructs with the wild-type 5′ UTR also reduce the repression of GFP-tagged RPL1B, presumably as a result of elevated level of Rpl1-mCherry protein. The wild-type and cre-less versions of RPL1B-mCherry both reduce RPL1A expression equivalently, about 3-fold after 10 and 20 h of induction (Supplementary Fig. S5). To further test the evidence for whether Rpl1 directly interacts with the 5’ UTR sequence of its own message we designed an Electrophoretic mobility shift assay (EMSA) experiment with purified Rpl1 (see “Methods” section) and a synthetic RNA, containing the UTR sequence. The presence of a distinct slower migrating band in lanes 2–3 (Supplementary Fig. S6) indicates direct binding between Rpl1 and the 5′-UTR of its own message.

a Alignment of the 5′ UTR sequence of RPL1B across different species of saccharomyces shows a high level of conservation. b The predicted structure of the RPL1B 5′ UTR with hairpin stem 1 (HP1) in orange and hairpin stem 2 (HP2) in blue. The red arrow indicates A at position 43 which differs from the G nucleotide seen in the consensus motif bound by Rpl1 in other species. c The common sequence and structure motif bound by Rpl1 in other species. The red arrow indicates the G nucleotide that is changed to A in RPL1B’s 5′ UTR sequence. d Variant sequences of the RPL1B 5′ UTR that were used to probe the sequence and structure requirements for autoregulation by Rpl1. Variants included deletions from the 5′ and 3′ ends, mutations to disrupt the hairpin stem structures, as well as compensating mutations to restore them, and mutations affecting the core GGAAG of the motif. The log GFP expression for uninduced (light bar) and induced (dark bar) cells are shown to the right for each sequence. * indicates UTR sequences with great than 3-fold repression.
Having verified that there is a post-transcriptional cis-regulatory element in the 5′ UTR of RPL1B that responds to the Rpl1 concentration, we sought to identify the sequence features required for binding by Rpl1. The RNAStructure web server72 (see “Methods” section) predicts the same minimum free energy secondary structure and maximum expectation secondary structure for the 64 base long 5’ UTR (Fig. 4b). The structure has two primary stems, with hairpin stem 1 (HP1) shown in orange and hairpin stem 2 (HP2) shown in blue. Remarkably, every known binding site for Rpl1 and its orthologs in other species contains a common sequence and structure motif, shown in Fig. 4c. This includes the binding sites for E. coli ribosomal protein L1 on both the L11 mRNA and the 23SrRNA73, and ribosomal proteins from several other bacteria and archaea on mRNA and rRNA binding sites49,74. It also includes the binding sites on stems H77 and H78 of 28SrRNA of both human and yeast75,76. The RPL1B UTR structure contains the same sequence and structure motif except for a single G to A change shown by the arrows in Fig. 4b, c.
To test if those sequence and structure features are required for regulation by Rpl1, we made several variants of the RPL1B 5′ UTR and placed them upstream of GFP driven by the TEF2 promoter. Figure 4d shows the wild-type 5’UTR with the Hairpin1 and Hairpin2 stems marked. Below that are 12 different variants and the logarithm of the GFP expressions from cells both uninduced (−Gal) and induced (+Gal) for the expression of the cre-less Rpl1 protein. The log expression values and the differences between induced and uninduced are also shown in the Table 1 (all data are available in Supplementary Data 4). The wild-type UTR is repressed by 0.70 (log reduction in GFP), verifying that the 5′ UTR is the regulatory region. Removing the bases at either the 5′ or 3′ sides of the structured region reduces but does not eliminate repression, but removing both together does eliminate repression, perhaps by altering the structure of the mRNA. To test the importance of the hairpin structures we modified the 5′ half of each stem to eliminate the structure, and then compensated by modifying the 3′ half to make a complementary sequence and recover the secondary structure. HP1-mut modifies the 5′ half of the HP1 sequence to eliminate the structure and it completely abolishes repression. HP1-cmp restores the predicted secondary structure and regains regulatory activity, in fact to a slightly higher level than the wild-type sequence.
We mutated HP2 in two different ways, first altering only the upper three bases in the 5′ half (HP2-mut1) and then by altering the upper four bases in the 5′ half (HP2-mut2). Both eliminate repression, demonstrating the importance of HP2 for the regulatory site. When the upper three bases are compensated (HP2-cmp1), repression is restored. However, when the upper four bases of HP2 are compensated (HP2-cmp2), there is still no regulation. This highlights the importance of the first G in the conserved sequence GGGAG shown in Fig. 4c. To further test the importance of the bases in the conserved motif we altered the GGAAG to GGTTG (3muts, which also contains an additional mutation to maintain the wild-type structure), and the repression is eliminated. In another variant we changed the GGAAG to GCAAC, along with two additional changes to maintain the secondary structure (4muts), and again the repression is eliminated. Finally, we tested the importance of the base that differs between the Rpl1B 5′ UTR (the A with the arrow in Fig. 4b) and the conserved G in the binding site motif (the arrow in Fig. 4c). The A to G variant (A43G) is still repressed, indicating that either base is acceptable at that position for regulatory activity. However, the expression of GFP in the non-induced state is lower for the mutant A43G than for the wild-type sequence, perhaps because it provides a higher affinity binding site for the intrinsic Rpl1 protein in the cell.
Discussion
Many recent studies use high-throughput methods to identify protein-mRNA interactions, some of which may be regulatory interactions. However, most of those approaches will only identify common events, where a protein regulates many genes. Discoordination of protein and mRNA levels adds to the complexity of these studies and any inferences that can be drawn. In addition, cases of autoregulation, such as in bacteria, would be missed because the proteins have only a single target and are not, primarily, regulatory proteins. Non-coding segments of RNA can evolve rapidly and acquire various roles including regulation of gene expression. They can be highly sensitive and autonomous sensors of the cellular environment and determine their own fate, such as to be translated or not. Riboswitches are an especially compelling example of such autonomous regulation because no protein is involved in the feedback response. Sensing protein concentrations can also be accomplished by RNAs and is likely to be much more common than is currently known, but because of the single target limitation to detection, directed searches are needed to identify such cases. We have used the GFP-fusion collection in yeast as a means of rapidly screening for examples of feedback regulation. We observed a continuum of repression levels (and a few unaffected genes) with 30% of ribosomal genes repressed by over 3-fold by expression of the cre-less gene, and that is likely an underestimate of the true number due to some limitations of our approach. In many cases both paralogs are affected similarly and those are likely due to increased protein degradation when excess protein is expressed. Seven examples have a greater than 3-fold reduction in expression and greater than 2-fold difference between paralogs. Those are the most likely candidates for autoregulation. We find that RPS22B regulates its own splicing, which is required for translation and is similar to a few other known autoregulatory examples in yeast. RPL1B, on the other hand, appears to regulate its own translation via binding to a sequence and structure motif in the 5′ UTR that is remarkably conserved in bacteria, archea and eukaryotes from yeast to mammals. We know that motif is necessary but it does not appear to be sufficient as changes outside of the conserved region can also affect regulation. More work is required to identify the complete mechanism, and to uncover the regulatory domains of the remaining examples we identified. The GFP-fusion collection in yeast provides an outstanding resource for identifying feedback regulation of all types, but the development of more flexible approaches will be necessary to do similar searches in more complex genomes.
Methods
GAL1 vector and yeast strains
The synthetic genes were cloned into a custom plasmid, MBJ1-mod5 (Supplementary Fig. S7). The vector backbone was pMW102-empty-MORF. The main features of this vector are a β-lactamase gene, replication origin for selection in E. coli, the URA3 gene for selection in ura- yeast strains and GAL1 promoter for induction with galactose and the mCherry coding region that can be fused to the cre-less gene. The His5 terminator site with 5′-NheI and 3′-XhoI restriction sites was synthesized by G-blocks from IDT. The mCherry sequence was cloned out from pMVS124-pACT1 (a generous gift from Max Staller) with a 3′-NheI and 5′-AvrII restriction sites by PCR using. pMW102-empty-MORF was linearized using primers to incorporate PacI and AvrII restriction sites downstream of the GAL1 promoter. One microgram of purified mCherry, His5 terminator and linearized vector backbone were digested with NheI, AvrII, XhoI, and PacI for 15 min and gel purified. The digested DNAs were ligated with T4 DNA ligase for 15 min at room temperature and transformed into DH5α cells. The sequences of the selected clones were verified by Sanger sequencing.
The cre-less genes were synthesized and integrated directly into the MBJ1-mod5 vector by Twist Biosciences (San Francisco, CA). The genes were designed to be lacking any cis-regulatory elements (cre-less) by eliminating any introns, replacing the 5′ UTR with an alternative sequence, and shuffling the synonymous codons of the gene using the program CodonShuffle64. The sequences of the shuffled coding regions are provided in the Supplementary Data 5.
The parental GFP-tagged yeast strains (S288C) were taken from the Yeast GFP library48 (a gift from Heather True-Krob). GFP-tagged strains were transformed with the plasmid containing the corresponding cre-less gene driven by the GAL1 promoter. Two or more independent clones were assayed for each sample.
Yeast transformation with cre-less plasmid
Yeast strains from the GFP collection (MATα SUC2 gal2 mal2 mel flo1 flo8-1 hap1 ho bio1 bio6) were grown in YPD medium (1% yeast extract, 1% bacto-peptone and 2% glucose) at 30 °C, overnight in 96 well plate format or in individual culture tubes. The cells were inoculated in 1 ml fresh YPD (10% v/v) and grown to OD600 = 1. The cells were collected by centrifugation at 2000 × g for 2 min and then mixed with 0.6–1 μg of the cre-less plasmid in buffer containing 100 mM LiOAc, 50% PEG (MW 3350). The resulting mixture was incubated for 5 min and subjected to heat shock at 40 °C for 20 min. The cells were mixed with 200 μl of fresh YPD, incubated at 30 °C with shaking and plated on selective medium (SD-URA). Several colonies from each plate were collected after 2–3 days for galactose induction.
Galactose induction and flow cytometry assay
Liquid cultures were inoculated and grown overnight in 400 µl SD-URA with 2% raffinose in 96 deep well plates at 30 °C. Overnight cultures were diluted into both SD-URA with 2% raffinose (uninduced) and SD-URA with 2% raffinose and 0.2% galactose (induced). Cells were grown at 30 °C for 10 h.
Two-hundred microliter cultures were transferred to 96-well plates and assayed on a CytoFLEX (Beckman Coulter). The live cells were gated and 10,000 events were acquired.
RPL1B Variants
Three additional RPL1B variants were synthesized (Supplementary Fig. S3) beside the cre-less version of the gene (RPL1B-cl). The RPL1B-wt retained the wild type sequence of native RPL1B with the 5′ UTR. The RPL1B-cd retained the 5′ UTR of the wild type sequence but the internal mRNA sequence was shuffled and was identical to the cre-less sequence. The RPL1B-mt was identical to the wild type coding sequence but lacked the 5′ UTR. All of these variations of RPL1B sequences were fused with mCherry sequence and cloned into MJB1 plasmid as described above.
Reporter gene assay
To construct yeast strains expressing GFP with either the RPS22B UTR or RPL1B UTR variants a new plasmid (BAC690-TEF) was designed. BAC690-TEF (Supplementary Fig. S8) was designed based on the BAC690_Euroscarf vector, which has an eGFP ORF 5′ to an ADH1 terminator77. The TEF2 promoter sequence was amplified using PCR from purified yeast saccharomyces cerevisiae (S288C) genomic DNA. The TEF2 promoter was cloned into the BAC690_Euroscarf vector 5′ of the eGFP sequence. UTR variants were cloned upstream of eGFP using NEB’s HiFi DNA Assembly. A PCR amplicon was made containing the TEF2 promoter, UTR variant, eGFP, and ADH1 terminator, as well as a kanamycin phosphotransferase cassette for integration in the yeast genome. The integration location was a dubious ORF (YBR032W).
Transformed cells were grown on YPD plates containing 200 µg/ml G418 to select for the integrated kanamycin cassette. Correct integrations were verified by colony PCR of the 5′ and 3′ junctions. Integrated strains were then transformed with their corresponding cre-less plasmid. Galactose induction and flow cytometry measurements were done as described above.
RNA structure prediction
We used the Matthews lab RNAstructure prediction web server72 (https://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html) version 6.0.1 with default parameters. It produces both a minimum free energy predicted secondary structure and a maximum expectation prediction secondary structure, which may be different. The structures shown in Fig. 4b, c were created using the StructureEdit program from the same package.
Protein purification
The cDNA sequence of Rpl1B was codon optimized for expression in E. coli and cloned into SBP-PRDM vector. The ORF was cloned downstream of the T7 promoter and was flanked by EcoRI and NotI restriction sites. An 18-nucleotide (CACCATCACCATCACCAT) sequence was attached to the 3′-end of the ORF for encoding an additional six amino-acid (HHHHHH) long peptide (His-tag) for purification. Expression of Rpl1B was achieved in E. coli BL21 (DE3) cells. Two-fifty microliter E. coli culture was grown in LB media until OD600 = 0.7 and induced by IPTG to 1 mM final concentration for 18 h. After lysing the cells by sonication, proteins were separated from the cellular debris by centrifugation at 15,000 rpm. The supernatant was filtered and loaded directly on a column prepacked with HisPur Ni-NTA resin (Thermo Scientific). After washing the column with buffer containing 50 mM Tris.HCl, pH 8.5, and 20 mM imidazole the protein was eluted in buffer containing 50 mM Tris.HCl, pH 8.5, 350 mM KCl, 20 mM MgCl2 and 200 mM imidazole. The protein was stored in a final binding buffer with 50 mM Tris.HCl, pH 8.5, 350 mM KCl, 20 mM MgCl2, 0.1% NP-40 and 2% Glycerol. Buffer exchange was performed by spinning the sample at 400 rpm using Amicon ultra-4 centrifugal filter unit with a 10 kDa molecular weight cutoff. The protein was visualized by running a 4–15% Tris-glycine PAGE gel with 0.1% SDS at 200 V for 1 h at room temperature. The gels were stained by InstantBlue (Sigma) and visualized by a Bio-Rad ChemiDoc imaging system.
RNA substrate production
Wild type RPL1B-UTR was cloned in a pUC19 vector under a T7 promoter. DNAs were amplified by PCR following the Phusion High-Fidelity PCR (NEB, catalog # E0553L) protocol. One microgram of the PCR product (linearized duplex DNA with 5′ T7 promoter and UTR or the control DNA) was used in a 50 μl in vitro transcription reaction following manufacturer’s protocol (NEB). The mixture of DNA and enzyme was incubated at 37 °C for 16 h. The RNA mixture was purified by Monarch RNA cleanup kit. The RNA concentration was measured by NanoDrop and adjusted to a 400 nM final concentration for EMSA.
Electrophoretic mobility shift assay
The protein−DNA binding reactions used buffer containing 50 mM Tris.HCl at pH 8.5, 350 mM KCl, 20 mM MgCl2, 0.1% NP-40, and supplemented with 10% glycerol. Either control RNA or RPL1B- UTR RNAs at a final concentration of 400 nM were incubated with 0.1 to 1 μg of purified Rpl1B for 30 min in 10 μl reaction volumes at 4 °C. The reaction mixtures were run on a 7.5% Tris-glycine PAGE gel at 80 V for 2 h in the cold room. The gels were stained with SYBR Green I stain for 20 min. The RNA in the bound (slow migrating) and unbound (fast migrating) bands were visualized by a BioRad imager with a 520 nM bandpass filter.
Statistics and reproducibility
Biological replicates, from two or three independent transformants for each cre-less gene, were measured for GFP expression with and without induction. The means for each condition (shown in Fig. 2 and available in Supplementary Data 1) were compared to rank genes by their level of repression. The same procedure was used for the other GFP measurements and results are provided in Supplementary Data files 2–4.
Data availability
The cytometry data are available from the Flow Repository (https://flowrepository.org) using accession ID FR-FCM-Z2UK. Source data underlying plots shown in figures are provided in Supplementary Data 1–4. All other relevant data are also available from the authors upon request.
A subset of the plasmids created for the project are available from Addgene (https://www.addgene.org). These are: pMJB1, the backbone into which the cre-less genes were inserted (ID: 160429); pMJB1-RPL1BCl, which includes the cre-less RPL1B gene (ID: 160430); pMJB1-RPS22B-Cl, which include the cre-less RPS22B gene (ID: 160431); pBAC690-TEF, which expresses eGFP under control of the TEF2 promoter for the study of variant 5′ UTS (ID: 160432). All plasmids and yeast strains used in the study are also available by request from the authors.
References
Atkinson, D. E. Regulation of enzyme function. Annu. Rev. Microbiol. 23, 47–68 (1969).
Pardee, A. B. Molecular-basis of biological regulation-origins from feedback inhibition and allostery. Bioessays 2, 37–40 (1985).
Ptashne, M. et al. Autoregulation and function of a repressor in bacteriophage lambda. Science 194, 156–161 (1976).
Ptashne, M., Johnson, A. D. & Pabo, C. O. A genetic switch in a bacterial virus. Sci. Am. 247, 128–130 (1982).
Alon, U. Network motifs: theory and experimental approaches. Nat. Rev. Genet. 8, 450–461 (2007).
Shen-Orr, S. S., Milo, R., Mangan, S. & Alon, U. Network motifs in the transcriptional regulation network of Escherichia coli. Nat. Genet. 31, 64–68 (2002).
Harbison, C. T. et al. Transcriptional regulatory code of a eukaryotic genome. Nature 431, 99–104 (2004).
Hughes, T. R. & de Boer, C. G. Mapping yeast transcriptional networks. Genetics 195, 9–36 (2013).
MacIsaac, K. D. et al. An improved map of conserved regulatory sites for Saccharomyces cerevisiae. BMC Bioinform. 7, 113 (2006).
Rosenfeld, N., Elowitz, M. B. & Alon, U. Negative autoregulation speeds the response times of transcription networks. J. Mol. Biol. 323, 785–793 (2002).
Savageau, M. A. Comparison of classical and autogenous systems of regulation in inducible operons. Nature 252, 546–549 (1974).
Wall, M. E., Hlavacek, W. S. & Savageau, M. A. Design of gene circuits: lessons from bacteria. Nat. Rev. Genet. 5, 34–42 (2004).
Lareau, L. F. & Brenner, S. E. Regulation of splicing factors by alternative splicing and NMD is conserved between kingdoms yet evolutionarily flexible. Mol. Biol. Evol. 32, 1072–1079 (2015).
Pervouchine, D. et al. Integrative transcriptomic analysis suggests new autoregulatory splicing events coupled with nonsense-mediated mRNA decay. Nucleic Acids Res. 47, 5293–5306 (2019).
Sun, Y. et al. Autoregulation of RBM10 and cross-regulation of RBM10/RBM5 via alternative splicing-coupled nonsense-mediated decay. Nucleic Acids Res. 45, 8524–8540 (2017).
Nagengast, A. A., Stitzinger, S. M., Tseng, C. H., Mount, S. M. & Salz, H. K. Sex-lethal splicing autoregulation in vivo: interactions between SEX-LETHAL, the U1 snRNP and U2AF underlie male exon skipping. Development 130, 463–471 (2003).
Penalva, L. O., Lallena, M. J. & Valcarcel, J. Switch in 3’ splice site recognition between exon definition and splicing catalysis is important for sex-lethal autoregulation. Mol. Cell Biol. 21, 1986–1996 (2001).
Ascano, M., Gerstberger, S. & Tuschl, T. Multi-disciplinary methods to define RNA-protein interactions and regulatory networks. Curr. Opin. Genet. Dev. 23, 20–28 (2013).
Hannigan, M. M., Zagore, L. L. & Licatalosi, D. D. Mapping transcriptome-wide protein-RNA interactions to elucidate RNA regulatory programs. Quant. Biol. 6, 228–238 (2018).
Matia-Gonzalez, A. M., Laing, E. E. & Gerber, A. P. Conserved mRNA-binding proteomes in eukaryotic organisms. Nat. Struct. Mol. Biol. 22, 1027–1033 (2015).
Mitchell, S. F., Jain, S., She, M. & Parker, R. Global analysis of yeast mRNPs. Nat. Struct. Mol. Biol. 20, 127–133 (2013).
Ramanathan, M., Porter, D. F. & Khavari, P. A. Methods to study RNA-protein interactions. Nat. Methods 16, 225–234 (2019).
Castello, A. et al. Insights into RNA biology from an atlas of mammalian mRNA-binding proteins. Cell 149, 1393–1406 (2012).
Gerstberger, S., Hafner, M. & Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet. 15, 829–845 (2014).
Mitchell, S. F. & Parker, R. Principles and properties of eukaryotic mRNPs. Mol. Cell 54, 547–558 (2014).
Nomura, M. Journey of a molecular biologist. Annu Rev. Biochem. 80, 16–40 (2011).
Nomura, M., Gourse, R. & Baughman, G. Regulation of the synthesis of ribosomes and ribosomal components. Annu. Rev. Biochem. 53, 75–117 (1984).
Nomura, M., Yates, J. L., Dean, D. & Post, L. E. Feedback regulation of ribosomal protein gene expression in Escherichia coli: structural homology of ribosomal RNA and ribosomal protein MRNA. Proc. Natl Acad. Sci. USA 77, 7084–7088 (1980).
Deiorio-Haggar, K., Anthony, J. & Meyer, M. M. RNA structures regulating ribosomal protein biosynthesis in bacilli. RNA Biol. 10, 1180–1184 (2013).
Meyer, M. M. rRNA Mimicry in RNA Regulation of Gene Expression. Microbiol. Spectr. 6, https://doi.org/10.1128/microbiolspec.RWR-0006-2017 (2018).
Romby, P. & Springer, M. Bacterial translational control at atomic resolution. Trends Genet. 19, 155–161 (2003).
Springer, M. et al. Autogenous control of Escherichia coli threonyl-tRNA synthetase expression in vivo. J. Mol. Biol. 185, 93–104 (1985).
Torres-Larios, A. et al. Structural basis of translational control by Escherichia coli threonyl tRNA synthetase. Nat. Struct. Biol. 9, 343–347 (2002).
Frugier, M. & Giege, R. Yeast aspartyl-tRNA synthetase binds specifically its own mRNA. J. Mol. Biol. 331, 375–383 (2003).
Levi, O. & Arava, Y. mRNA association by aminoacyl tRNA synthetase occurs at a putative anticodon mimic and autoregulates translation in response to tRNA levels. PLoS Biol. 17, e3000274 (2019).
Castello, A., Hentze, M. W. & Preiss, T. Metabolic enzymes enjoying new partnerships as RNA-binding proteins. Trends Endocrinol. Metab. 26, 746–757 (2015).
Hentze, M. W., Castello, A., Schwarzl, T. & Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 19, 327–341 (2018).
Andrake, M. et al. DNA polymerase of bacteriophage T4 is an autogenous translational repressor. Proc. Natl Acad. Sci. USA 85, 7942–7946 (1988).
Gold, L. Posttranscriptional regulatory mechanisms in Escherichia coli. Annu. Rev. Biochem. 57, 199–233 (1988).
McPheeters, D. S., Stormo, G. D. & Gold, L. Autogenous regulatory site on the bacteriophage T4 gene 32 messenger RNA. J. Mol. Biol. 201, 517–535 (1988).
Russel, M., Gold, L., Morrissett, H. & O’Farrell, P. Z. Translational, autogenous regulation of gene 32 expression during bacteriophage T4 infection. J. Biol. Chem. 251, 7263–7270 (1976).
Tuerk, C., Eddy, S., Parma, D. & Gold, L. Autogenous translational operator recognized by bacteriophage T4 DNA polymerase. J. Mol. Biol. 213, 749–761 (1990).
Breaker, R. R. Prospects for riboswitch discovery and analysis. Mol. Cell 43, 867–879 (2011).
Mandal, M. & Breaker, R. R. Gene regulation by riboswitches. Nat. Rev. Mol. Cell Biol. 5, 451–463 (2004).
McCown, P. J., Corbino, K. A., Stav, S., Sherlock, M. E. & Breaker, R. R. Riboswitch diversity and distribution. RNA 23, 995–1011 (2017).
Stormo, G. D. & Ji, Y. Do mRNAs act as direct sensors of small molecules to control their expression? Proc. Natl Acad. Sci. USA 98, 9465–9467 (2001).
Vitreschak, A. G., Rodionov, D. A., Mironov, A. A. & Gelfand, M. S. Riboswitches: the oldest mechanism for the regulation of gene expression? Trends Genet. 20, 44–50 (2004).
Huh, W. K. et al. Global analysis of protein localization in budding yeast. Nature 425, 686–691 (2003).
Fu, Y., Deiorio-Haggar, K., Anthony, J. & Meyer, M. M. Most RNAs regulating ribosomal protein biosynthesis in Escherichia coli are narrowly distributed to Gammaproteobacteria. Nucleic Acids Res. 41, 3491–3503 (2013).
Dean, D. & Nomura, M. Feedback regulation of ribosomal protein gene expression in Escherichia coli. Proc. Natl Acad. Sci. USA 77, 3590–3594 (1980).
de la Cruz, J. et al. Feedback regulation of ribosome assembly. Curr. Genet. 64, 393–404 (2018).
Warner, J. R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 24, 437–440 (1999).
Warner, J. R. & McIntosh, K. B. How common are extraribosomal functions of ribosomal proteins? Mol. Cell 34, 3–11 (2009).
Chaker-Margot, M. Assembly of the small ribosomal subunit in yeast: mechanism and regulation. RNA 24, 881–891 (2018).
Chaker-Margot, M. & Klinge, S. Assembly and early maturation of large subunit precursors. RNA 25, 465–471 (2019).
Fewell, S. W. & Woolford, J. L. Jr. Ribosomal protein S14 of Saccharomyces cerevisiae regulates its expression by binding to RPS14B pre-mRNA and to 18S rRNA. Mol. Cell Biol. 19, 826–834 (1999).
Gabunilas, J. & Chanfreau, G. Splicing-mediated autoregulation modulates Rpl22p expression in Saccharomyces cerevisiae. PLoS Genet. 12, e1005999 (2016).
Macias, S., Bragulat, M., Tardiff, D. F. & Vilardell, J. L30 binds the nascent RPL30 transcript to repress U2 snRNP recruitment. Mol. Cell 30, 732–742 (2008).
Vilardell, J., Chartrand, P., Singer, R. H. & Warner, J. R. The odyssey of a regulated transcript. RNA 6, 1773–1780 (2000).
Vilardell, J., Yu, S. J. & Warner, J. R. Multiple functions of an evolutionarily conserved RNA binding domain. Mol. Cell 5, 761–766 (2000).
Plocik, A. M. & Guthrie, C. Diverse forms of RPS9 splicing are part of an evolving autoregulatory circuit. PLoS Genet. 8, e1002620 (2012).
Badis, G., Saveanu, C., Fromont-Racine, M. & Jacquier, A. Targeted mRNA degradation by deadenylation-independent decapping. Mol. Cell 15, 5–15 (2004).
He, F., Li, C., Roy, B. & Jacobson, A. Yeast Edc3 targets RPS28B mRNA for decapping by binding to a 3’ untranslated region decay-inducing regulatory element. Mol. Cell Biol. 34, 1438–1451 (2014).
Katz, L. & Burge, C. B. Widespread selection for local RNA secondary structure in coding regions of bacterial genes. Genome Res. 13, 2042–2051 (2003).
Eberhardt, I. et al. Autoregulation of yeast pyruvate decarboxylase gene expression requires the enzyme but not its catalytic activity. Eur. J. Biochem. 262, 191–201 (1999).
Hooks, K. B., Naseeb, S., Parker, S., Griffiths-Jones, S. & Delneri, D. Novel intronic RNA structures contribute to maintenance of phenotype in Saccharomyces cerevisiae. Genetics 203, 1469–1481 (2016).
Danin-Kreiselman, M., Lee, C. Y. & Chanfreau, G. RNAse III-mediated degradation of unspliced pre-mRNAs and lariat introns. Mol. Cell 11, 1279–1289 (2003).
Parenteau, J. et al. Introns within ribosomal protein genes regulate the production and function of yeast ribosomes. Cell 147, 320–331 (2011).
Peng, B., Williams, T. C., Henry, M., Nielsen, L. K. & Vickers, C. E. Controlling heterologous gene expression in yeast cell factories on different carbon substrates and across the diauxic shift: a comparison of yeast promoter activities. Micro. Cell Fact. 14, 91 (2015).
Ban, N. et al. A new system for naming ribosomal proteins. Curr. Opin. Struct. Biol. 24, 165–169 (2014).
Cliften, P. et al. Finding functional features in Saccharomyces genomes by phylogenetic footprinting. Science 301, 71–76 (2003).
Reuter, J. S. & Mathews, D. H. RNAstructure: software for RNA secondary structure prediction and analysis. BMC Bioinform. 11, 129 (2010).
Said, B., Cole, J. R. & Nomura, M. Mutational analysis of the L1 binding site of 23S rRNA in Escherichia coli. Nucleic Acids Res. 16, 10529–10545 (1988).
Kohrer, C., Mayer, C., Neumair, O., Grobner, P. & Piendl, W. Interaction of ribosomal L1 proteins from mesophilic and thermophilic Archaea and Bacteria with specific L1-binding sites on 23S rRNA and mRNA. Eur. J. Biochem. 256, 97–105 (1998).
Anger, A. M. et al. Structures of the human and Drosophila 80S ribosome. Nature 497, 80–85 (2013).
Bernier, C. R. et al. RiboVision suite for visualization and analysis of ribosomes. Faraday Discuss. 169, 195–207 (2014).
Sheff, M. A. & Thorn, K. S. Optimized cassettes for fluorescent protein tagging in Saccharomyces cerevisiae. Yeast 21, 661–670 (2004).
Acknowledgements
Funding for this work came from NIH grants R01HG000249 and R21GM126307. F.B. was supported by R25HG006687. We thank Dr. Max Staller for the gift of a plasmid used in this work and Dr. Heather True-Krob for the gift of the yeast GFP-fusion collection.
Author information
Authors and Affiliations
Contributions
B.R., D.G., F.B., J.A.Y.C., and M.A.W. performed the experiments. B.R., D.G., M.A.W., and G.D.S. designed the experiments, analyzed the data and participated in writing and revising the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Roy, B., Granas, D., Bragg, F. et al. Autoregulation of yeast ribosomal proteins discovered by efficient search for feedback regulation. Commun Biol 3, 761 (2020). https://doi.org/10.1038/s42003-020-01494-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-020-01494-z
- Springer Nature Limited
This article is cited by
-
A single-cell transcriptome atlas of human euploid and aneuploid blastocysts
Nature Genetics (2024)
-
Contrasting outcomes of genome reduction in mikrocytids and microsporidians
BMC Biology (2023)
-
Effects of aneuploidy on cell behaviour and function
Nature Reviews Molecular Cell Biology (2022)
-
Intron-mediated induction of phenotypic heterogeneity
Nature (2022)