

Abstract
The production and consumption of animal-source foods must be transformed to mitigate negative environmental outcomes, including greenhouse gas emissions and land-use change. However, livestock are also key for food production and for livelihoods in some settings, and they can help preserve biodiversity and certain ecosystems. Previous studies have not yet fully explored sustainability limits to the use of grazing lands for food production in the context of biodiversity. Here we explore ‘biodiversity limits’ to grassland ruminant production by estimating the meat and milk production from domestic ruminants limited to grazing areas and stocking densities where livestock can contribute to the preservation or restoration of biodiversity. With biodiversity-friendly grazing intensities at 0–20% biomass removal depending on aridity, this take on biodiversity limits corresponds to 9–13% and 26–40% of the current grassland-based milk and meat production, respectively. This equals only 2.2 kg of milk and 0.8 kg of meat per capita per year, globally, but altered management and moving from meat-specialized to meat-and-dairy systems could increase the potential production while still remaining within this approach to biodiversity limits.
Similar content being viewed by others


Main
Meat and dairy are important food resources that are valuable and desirable to individuals and cultures around the world1. Livestock rearing supports the livelihoods of 600 million smallholder farmers and pastoralists, and livestock market chains employ 1.3 billion people globally2,3.
Livestock also put considerable pressures on ecosystems, climate, waters and biodiversity. Livestock rearing on land is one of three key drivers behind land-use change and the single largest driver of habitat loss4. In addition, livestock contribute substantially to climate change5. Global livestock production has increased nearly fivefold since 19616, while the world population has only tripled. As the demand for milk and meat increases, so does the pressure of livestock production on ecosystems7,8,9.
Livestock also provide substantial value for society. A diet containing small amounts of meat from grass-fed livestock requires less arable land than a purely plant-based diet because ruminants can transform inedible biomass to highly nutritious food, acting as net protein providers10. Livestock can also act as peoples’ financial savings, represent rich cultural capital and maintain biocultural landscapes highly valued by people2. Some lands have unique flora and fauna that have co-evolved and been maintained through grazing by domestic animals11. These are places where human–livestock interactions have shaped biodiversity over a long time, such as the nomadic, pastoralist communities in Mongolia and Inner Mongolia migrating across the steppe12; Andean mountainous drylands where alpaca, llamas and sheep grazing at low intensity has given rise to outstanding endemic species richness13; and European semi-natural pastures that harbour exceptional biodiversity and rich cultural values14,15.
In efforts to better understand and describe the areas and ways to sustainably rear livestock, there is an ongoing scholarly debate on when to use what lands and under what management conditions. Parts of this discourse concern grazing livestock and what should serve as a suitable reference point for where and how grazing could or should take place16,17 in relation to the vast areas of grazing lands that have expanded since pre-industrial times18,19,20. However, no previous studies have explored limits to grassland use for domestic ruminant production based on biodiversity. Impacts from grazing on biodiversity are highly variable21, in that high-intensity grazing and management most often are harmful to biodiversity22,23,24,25,26, while low-intensity grazing can maintain and support it22,23,25,26,27, with some variability between taxa and temporal scales28.
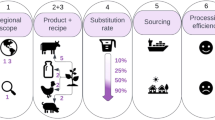
In this study, we estimate how much meat and milk could be produced from global grazing lands, using domestic livestock, while safeguarding biodiversity, which is our demarcation of ‘biodiversity limits’. To do this, we use theory and empirical findings on the positive and negative effects of domestic livestock grazing on biodiversity to select grazing areas and set grazing intensities on the basis of what could preserve a certain level of biodiversity. We use a three-step methodology (Fig. 1): (1) using the History Database of the Global Environment (HYDE) v.3.2 (ref. 29) to identify the lands where biodiversity conservation can be aligned with grazing, (2) adjusting grazing intensities to levels compatible with grassland biodiversity conservation and (3) estimating the potential production of milk and meat on the basis of current national herd structures and feed conversion ratios of grass-based production. We also do sensitivity tests of the underlying assumptions through a range of alternative scenarios.

The steps are (1) selecting areas on the basis of the historical legacy of grazing, (2) applying a biodiversity-limited grazing intensity and (3) using current herd structures, feed intakes and yields to calculate the potential production of milk and meat. CR, converted rangelands; HNR-HI, historical natural rangelands under current high-intensity use; SNP-HI, semi-natural pastures under current high-intensity use. Herd ratios, feed intake and yield data from ref. 38. AGB calculated using the methodology of ref. 59. Credit: animal icons, Pixabay.
In identifying the lands to be used for grazing, we limit the analysis to areas where grazing already exists, as land-use competition by livestock needs to be reduced globally8,30,31. Other studies have used Pleistocene conditions as baselines to study land-cover changes or historic grazing by wild (including now extinct) herbivores16,32. Here, however, we choose a more recent baseline, the pre-industrial year 1700 (in the Holocene), as we consider that baseline more relevant for this study on grazing by (managed) domestic animals.
We include all HYDE ‘natural rangelands’ grazed under low-intensity practices (Methods). For grazing lands in Olson forest biomes deforested after 1700, we assume biodiversity benefits from reforestation, land restoration and/or rewilding22,33,34,35,36, excluding them from domestic grazing. However, where historical human–livestock interactions have shaped landscapes and biodiversity11,37, we allow continuous low-intensity grazing, identified by pre-1700 use—that is, before the widespread expansion of modern agricultural practices. If grazing lands in forest biomes were grazed in 1700 and are still grazed, we assume those grazing lands hold unique biodiversity and include them in our analysis. Similarly, areas in non-forest biomes currently under high-intensity use are included if they were natural rangelands in 1700.
We stress that our study focuses on grazing lands only. There is also potential for meat and milk production in mixed crop–livestock systems that can be compatible with biodiversity. However, estimating such potentials is outside the scope of this study; our estimate of the production potential is based solely on the biomass from grazing lands. We moreover use low-intensity grazing rates (0–20% of aboveground biomass (AGB), 10% at baseline) to ensure grassland biodiversity compatibility (Methods). Using data from Herrero et al.38, we maintain the national ruminant composition and use current milk and meat yields in relation to feed intake in grassland-based systems (Fig. 1). For instance, if a country has a 50:50 ratio of dairy cows to beef cattle in such systems, we retain this proportion but adjust the animal numbers on the basis of available forage biomass. Our estimates presume continuous use of grazing lands for domestic livestock (cattle, sheep and goats) rather than rewilding, though the presence of livestock might threaten certain biodiversity (for example, mammals28).
Results
We included 23.5 million km2 (73% of currently grazed areas) (Fig. 2) in our assessment on the basis of our take on biodiversity limits. We estimated that this land annually produces 2.5 billion tonnes of AGB (dry matter), of which we allocated 10% (low-intensity grazing) to ruminants. In our model, this results in an annual production of 4.7 million tonnes of meat from cattle, and 1.5 million tonnes of meat from small ruminants (Fig. 3). This is approximately 28% of the current production in grassland-based systems39, or 6.3% of the total global ruminant meat production6 (Fig. 4).

Credit: basemap, Esri, Garmin International, Inc., US Central Intelligence Agency (The World Factbook), National Geographic Society.

The map base layer shows the total protein production per country, and the pie charts show the share of total production of bovine meat (slaughter weight), bovine milk, small ruminant meat (slaughter weight) and small ruminant milk per region. The size of the pie charts shows how much of the global production occurs in a region. EASA, East and Southeast Asia; EEUR, Eastern Europe; EUR, Western Europe; LAM, Latin America and the Caribbean; MNA, Middle East and North Africa; NAM, North America; OCE, Oceania; RF, Russian Federation; SAS, South Asia; SSA, sub-Saharan Africa. Credit: basemap, Esri, Garmin International, Inc., US Central Intelligence Agency (The World Factbook), National Geographic Society.

a,b, Percentage of production within biodiversity limits compared to the current total production in all production systems (a) and compared to the current total production in grassland-based production systems (b). In a, the bars show the baseline, and the whiskers show the total minimum and maximum output of meat and milk across the scenarios (the dots on the whiskers) of the sensitivity analysis. The horizontal lines in a and b show the global average percentages.
Meat production was geographically concentrated in Oceania (1.6 million tonnes; 25% of all production), Latin America and the Caribbean (1.5 million tonnes; 24%), sub-Saharan Africa (0.9 million tonnes; 16%) and North America (0.6 million tonnes; 10%). The production of beef dominated in all regions except East and Southeast Asia, where 40% of the global production of meat from sheep and goats (small ruminants) occurred (another 19% occurred in Oceania and 18% in sub-Saharan Africa) (Fig. 3).
In Oceania, the potential beef production within biodiversity limits was 82% of the current grassland-based beef production and about half of the current production of mutton and goat meat. Similarly, production within biodiversity limits equalled about 45% of beef in North America and sub-Saharan Africa and about 45% of small ruminant meat in East and Southeast Asia and Eastern Europe in current grassland-based systems (Fig. 4).
The total annual global potential production of cow’s milk within biodiversity limits was estimated to be 15.9 million tonnes, and the production of milk from sheep and goats was estimated at 1.1 million tonnes. The total global production represents 9% of the milk currently produced in grassland-based systems39 and 1.5% of the current total global milk production6 (Fig. 4).
The potential production of milk was also concentrated to four regions of the world: Oceania (4.8 million tonnes; 28% of all production), Latin America and the Caribbean (3.7 million tonnes; 22%), the Middle East and North Africa (3.3 million tonnes; 19%) and sub-Saharan Africa (3 million tonnes; 18%) (Fig. 3).
In our model, no milk production occurs in North America, and no small ruminant milk production occurs in Oceania, following our use of the current national compositions of large and small ruminants in different systems for our allocation of feed38. These regions currently have no dairy in grassland-based systems (Methods), and we therefore allocate no feed to such systems.
In Australia and sub-Saharan Africa, the potential production of milk within biodiversity limits was 45% and 19%, respectively, of the current total annual milk production. Across all other regions, the corresponding number was between 0% and 6%. For meat, the potential production within biodiversity limits amounted to larger shares of the current grassland-based production than for milk, but with considerable variation; from ∼1% in South Asia to ∼20% in Oceania (including Australia) (Fig. 4).
Globally, this potential production translates to approximately 2.2 kg of milk and 0.8 kg of meat per person annually, equalling 6 and 2 g per day (with the 2021 population of 7.8 billion) or an average daily supply of 0.5 g of ruminant protein per person.
Per capita production potential varied between countries. The top four for milk were Australia (180 kg per capita per yr), Kazakhstan (135 kg per capita per yr), Uruguay (92 kg per capita per yr) and Mongolia (70 kg per capita per yr). Per capita milk production was less than 1 kg yr−1 for 102 countries and less than 10 kg yr−1 for 32 countries. For 133 countries, grazing-based production within biodiversity limits equalled less than 10% of current consumption. However, for Guyana (69%), Australia (60%), Kazakhstan (47%), Zimbabwe (43%), Uruguay (41%) and Madagascar (29%), it equalled larger shares, though sometimes compared with low current consumption. In Angola, Mozambique and Gabon, milk production from grazing lands exceeded current consumption by 63%, 18% and 3%, respectively (Supplementary Information).
For meat, the highest per capita production in our model occurred in Australia (60 kg per capita per yr), Uruguay (38 kg per capita per yr) and Mongolia (33 kg per capita per yr). In 129 countries, annual per capita meat production was less than 1 kg, and in 26 countries, it was less than 10 kg. In 129 countries, production within biodiversity limits represented less than 10% of current consumption, and in 144 countries, it represented less than 50%. In Namibia (94%), Botswana (70%) and the Congo (65%), the potential production equalled larger shares than current consumption, though compared with low current consumption. However, in Mozambique, Australia, Angola, Guyana and Uruguay, meat production within biodiversity limits exceeded current per capita consumption by 121%, 64%, 36%, 34% and 30%, respectively (Supplementary Information).
Sensitivity of findings to critical assumptions
We tested the sensitivities of the results to our four key assumptions: (1) grazing land selection, (2) grazing intensity, (3) the historical legacy of land use and (4) herd structures and livestock productivities (Table 1 and Supplementary Information).
Grazing areas
We tested reforesting grazing lands in forest biomes (SNP-HI and CR in Fig. 1) (A), assuming no restoration potential from reintroduced low-intensity grazing in areas with current high-intensity management (HNR-HI and SNP-HI in Fig. 1) (B) and low-intensity grazing on all HYDE grazing lands (C) (Table 1). In scenarios A and B, milk and meat production was reduced by ∼5% compared with the baseline, while in scenario C, production increased by around 70%. This larger effect (70%) compared with the change in area (−27%, 23.5 million km2 instead of 32) results from removing production from high-yielding pastures (Fig. 1).
Grazing intensity
Drawing on theory and empirical findings on the relationship between grazing intensity, hydroclimate and biodiversity impacts (Supplementary Information), we tested removing 20% instead of 10% of biomass, which obviously resulted in doubled production (D), and increasing the biomass removal to 20% in wetter areas but removing all grazing from hyper-arid and arid areas, as even 10% removal may negatively impact biodiversity in drier contexts (F) (Table 1 and Supplementary Information). This increased milk and meat production by ∼42% compared with the baseline (Supplementary Data 1).
Historical legacy
We distributed ruminants on lands where the historical legacy of grazing has shaped the ecosystem and grassland biodiversity, using the year 1700 as the baseline reference point. The expansion and intensification of land use have varied geographically, however29—for example, following colonization. We therefore also tested the years 1800 (E) and 1500 (G) (Table 1 and Supplementary Table 1). The selected area changed by 8.8% (E) and −0.7% (G), leading to an increase of milk and meat production by 14.8% and 0.7% compared with the baseline.
Herd structures and productivity
Our baseline used current herd structures and productivities. In the data used38, most high-income countries have no grassland-based dairy; these regions currently operate mixed systems with more grains and concentrates. New Zealand, Australia, Norway and Switzerland are exceptions. We estimated the effects of Europe and North America adopting similar grassland-based production systems as those of these countries (H) (Table 1). In this scenario, milk production increased by 74.6%, but meat production decreased by 3.4% compared with the baseline (Supplementary Tables 2 and 3).
Discussion
Production corresponds to 5–30% of healthy meat protein recommendations
Grazing lands managed under our take on biodiversity limits (with current herd structures and productivity levels) would yield modest meat and milk outputs: 2 g of meat and 6 g of milk per person daily on a global scale, totalling 0.5 g of protein per capita (with a population of 7.8 billion). Previous research restricting livestock to grass from grazing lands but without biodiversity considerations found that approximately 9 g of protein per person per day could be produced40. These studies assume continued use of all global grazing lands (∼32 million km2 compared with 23.5 million km2 used here) at current or higher utilization rates, partially explaining the difference.
However, even when we include all grazing lands from the HYDE database (32.3 million km2), total production increases by only 70% for milk and 64% for meat, which gives a daily per capita protein production of ∼0.8 g (Supplementary Information and Supplementary Table 4). Land area inclusion is thus not the main explaining factor for the difference between our results and previous results. Nor is it the grassland utilization rate: all else being equal, even if 100% of the AGB was grazed (that is, a tenfold increase of feed input), the daily protein supply would still equal only ∼5 g per capita. Hence, the difference between our results and those of previous studies is mainly explained by differences in livestock productivities and herd structures, such as the assumptions about the number of dairy-producing animals in relation to specialized meat production (see discussion below).
Applying our biodiversity limits to the previous estimated potential production of 9 g of protein40 by reducing grazing lands by 27% (23.5 million km2 instead of 32), and reducing grazing intensity by half, yields an approximate output of ∼3 g of protein per person daily. This suggests that grazing lands under this take on biodiversity limits could provide around half or a few grams of protein per capita per day. To put this in perspective, the EAT-Lancet Commission31 recommended 9 g (range 0–18 g) of protein from ruminant meat and milk daily. Our results indicate that 5–30% (0.5 to ∼3 g) of this could be supplied by minimal-condition grazing systems aligned with our approach to biodiversity limits.
Future research could explore biodiversity limits to diverse mixed systems, incorporating various feeds from croplands to potentially increase meat and milk production. Although using cropland for feed is usually considered inefficient land use10, well-designed mixed crop–livestock systems offer several advantages. For instance, integrating perennial forage crops into annual cropping systems can promote nitrogen fixing and soil carbon sequestration and reduce the need for pesticides, also benefiting biodiversity41,42. Some argue that forage from such systems is not in direct competition with food43. Additionally, crop residues such as cereal stover is another feed source that can be utilized in ruminant production without competing with food (though other competing uses exist). These resources are already vital in ruminant production in many low-income settings38. To quantify this potential of livestock production from such resources could be an important topic for further research.
‘Grassland-based’ reflects a diverse and complex reality
We used the best available data on global livestock productivities and herd structures38, reflecting the real (although somewhat outdated) production situation. These data are, however, highly uncertain with substantial data gaps due to the vast number of livestock globally, variety in production systems, the difficulty of ascertaining exact feed intakes and limitations in nations’ data collection capacity and reporting. Grassland-based systems also sometimes use stover and ‘occasional’ feed, impacting productivity. Since we are estimating the production potential from grazing, we have assumed feed intake solely from grazing. This may have resulted in slight overestimations of production potentials in some instances. An update of these data would greatly aid further research.
Also, we used the Food and Agriculture Organization (FAO) standard39 categorizations of livestock systems as ‘grassland-based’ where more than 90% of livestock feed come from forage and where more than 10% of feed is produced on-farm (Fig. 1 and Methods). This implies uncertainties as, in reality, these are rather loose classifications; forage can be nutritious silage or poorer feeds, and ‘more than 90%’ can imply 100% or indeed ‘only’ 90%. The general pattern of these categories in the data38 reflects the fact that feed and yields vary considerably depending on the level of management, technology, access to resources and so on, and that high-income, high-input regions are generally more productive. We did not make assumptions of intensification in low-productivity regions, as we aimed to make the scenarios reflect real input:output ratios as best as possible, but the approach and results show that there is room to increase production within biodiversity limits by altering herd structures and improving productivity. The dairy scenario for Europe and North America illustrated the regional potential for increased production with herd redistribution to optimize feed utilization. A shift to grassland-based dual milk–meat systems would increase production from the same amount of feed and land. Breeding and improved animal health could also increase productivity without increased feed requirements44. However, high-yielding, intensive production systems cannot be fitted within the low-intensity principles used in our approach.
The data at the national level mask sub-national diversity, especially in large and climatically diverse countries, and our approach does not spatially distribute livestock types accordingly. Because our method for calculating the available biomass is spatially explicit, though, the non-spatially-explicit livestock distribution does not impact the amount of potential production.
Lastly, terms such as ‘grassland’, ‘grazing land’, ‘pasture’ and ‘rangeland’ have plural and contested meanings (see, for example, ref. 45). Estimates of their global distribution are uncertain46. We have used HYDE v.3.2 as it is well established and widely used29 and is based on reported FAO statistics and models continuously incorporating various historical data47. Estimates of grazing lands are generally consistent compared to a satellite-based approach and various assumptions of per capita land use (see the Supplementary Information of refs. 29,48), but localized studies have found both overestimates49 and underestimates50 of historical grazing lands, and there are parallel schools of thought disagreeing with HYDE’s reliance on the Olson biome way of conceptualizing landscapes (for example, ref. 17). In light of this, we welcome further refinement of the approach and urge readers to keep in mind the exploratory nature of this study.
Biodiversity impacts
This approach to limiting ruminants could aid biodiversity restoration both within and outside grazing areas. First, our approach implies an overall extensification of the use of grazing lands (Supplementary Information). Considering the degraded state of 23–50% of global grasslands21,51 and the increasing shares of intensive pastures29 with monoculture grass cultivation and high nutrient deposition that is harmful to most organisms11, extensification would probably benefit biodiversity overall. In some areas, restoration efforts could be needed.
In some places, there could be room for increased grazing intensity within grassland biodiversity limits—that is, beyond the 20% biomass removal maximally used here. Gaining a more detailed understanding of the quantified relationship between stocking densities, landscape configuration and biodiversity impacts across a wide range of geographic locations is key for further research. It would also be important to analyse biodiversity limits to ruminant livestock while considering biodiversity impacts from the herd structures—that is, the mix of large and small ruminants52, mobility and grazing management strategies (mob, rotational, continuous and so on). Some scholars, for example, argue that mimicking guilds of wild herbivores allows for higher herbivore densities without negative ecosystem impacts, but they caution against assuming that livestock could replace more complex wild grazing16.
Moreover, our approach leaves nearly 10 million km2 of current grazing lands without livestock. Biodiversity benefits on these areas depend on the state of the land and the surrounding landscape, as well as any restoration efforts; land sparing does not automatically lead to nature restoration53. In overgrazed or degraded areas, rewilding could restore ecosystems, and assemblages of wild herbivores often provide more functional diversity than domestic livestock because of different body sizes, diets and so on54. There are diverse potential uses of this land, and nature restoration, especially rewilding, needs a social licence to operate, including mitigation measures for potential land-use conflicts. Reforestation in response to reduced land-use needs for food production can also offer potential for climate change mitigation55.
Lastly, we set the biomass removal in our analysis on the basis of no or minimal impact on biodiversity from grazing. In reality, however, global international agreements or local or regional goals or regulations may desire to allow for more negative impacts on biodiversity—for example, depending on trade-offs with other desirable outcomes, or in relation to the state of the area in question. With greater allowance for negative impacts on biodiversity comes greater potential production of milk and meat, and it would be up to the political processes to determine what levels and types of biodiversity impacts would be acceptable.
Challenges to managing low-intensity grazing systems
The type of systems that are in line with the low-intensity, landscape-determined practices in our approach may be incompatible with current animal husbandry conditions and market demands. Increased marketization in pastoral regions can change grazing management and cause overgrazing56, and the human–livestock systems that have shaped biodiversity-rich landscapes throughout history have been driven not by market interests but by locally available resources and striving to meet local needs. Also, fences, roads, degraded land, urban areas and privatized land fragment the landscapes shaped by human–livestock interactions. We could not account for production-reducing effects from land fragmentation in our calculations, but it is possible that this would reduce production even further since the feasibility of low-intensity grazing, especially in arid areas, can depend on the mobility of animals57.
Fairer distribution of livestock production and consumption
With our approach, the potential production varied considerably across regions, and our theoretical model is a ‘world free of trade’ (all production is ‘consumed’ domestically or regionally). For example, East and Southeast Asia, home to nearly 30% of the global population, would produce only 5% of the milk and 11% of the meat, corresponding to low per capita availability (0.3 kg of meat and 0.4 kg of milk per person yearly). Conversely, Oceania, with only 0.5% of the global population, would produce 28% of the milk and 25% of the meat, leading to high regional per capita availability (37 kg of meat and 114 kg of milk per capita yearly). Currently, livestock product consumption exacerbates inequality, with large economies exploiting grazing lands in other regions; the European Union, the USA, China and Japan are responsible for >50% of global grazing land exploitation56. Given historical and present resource infringement with colonial roots, the redistribution of milk and meat is crucial for justice in sustainable food system development.
In this study, we explored the concept of biodiversity limits to ruminant livestock production based on both the selection of grazing lands and the intensity of grazing. On the selected areas, with low-intensity practices and using current herd structures and productivity levels, global annual production of milk in our model was ∼17 million tonnes, and that of meat was ∼6.2 million tonnes, with variability across regions. To put this in perspective, current global milk production from cattle, sheep and goats is about 1.1 billion tonnes, and meat production from these species is 99 million tonnes6. Our conclusions remained consistent across sensitivity analyses of the core assumptions. Our results contribute to the growing body of literature on changes to ruminant food production and consumption to curb negative biodiversity impacts, a critical sustainability goal. We encourage further research to develop and customize this approach for regional and local contexts. Future studies should also explore the production potential from croplands while considering farmland biodiversity.
Methods
Step 1: selecting grazing areas
We used HYDE, which combines human population data, estimates of historic human populations and agricultural land-use statistics with satellite information and allocation algorithms. HYDE shows human-induced land-use change for the past 12,000 years29,48,58. We used version 3.2, in which different types of grazing lands can be distinguished on the basis of the intensity of the use. In HYDE v.3.2, data for grazing lands are gathered primarily from country-reported statistics from the FAO since 1960 and to the current date, while historical data are modelled on the basis of population and per capita land-use estimates29. Grazing lands are allocated spatially using the European Space Agency land-cover classes and sequential allocation rules based on the probability of a land-cover class containing either cropland or grazing land. Lastly, grazing lands are divided into pastures and rangelands on the basis of population density and aridity, and rangelands are further divided into natural rangelands and converted rangelands on the basis of whether they exist within forest biomes or not, using the Olson/World Wildlife Fund (WWF) ecoregions29. HYDE features raster data at a 5′ spatial resolution, equal to grid cells that are approximately 85 km2.
Most of the spatial analysis was conducted using ArcGIS Pro v.3.0.3. Some supplementary tasks were performed in R 4.3.3 software.
In HYDE v.3.2, natural rangelands are “natural grasslands, shrublands, woodlands, wetlands, and deserts” where there is low-intensity grazing and where no conversion of the natural vegetation has occurred29. They constitute around 66% of HYDE’s total grazing lands. In our analysis, we considered grazing by domestic livestock as an integral, natural part of natural rangeland ecosystems and therefore included all areas for the year 2017 (the most recent data), for a total of 21.4 million km2 (Figs. 1 and 2).
Converted rangelands in HYDE v.3.2 are low-intensity grazing areas, but unlike natural rangelands, they exist in forest biomes and hence have involved conversion of the natural vegetation to make room for grassland establishment29. From the perspective of biodiversity impacts of grazing, the historical legacy of foraging management is key. To account for this, we used the number of km2 of land present in each grid cell in 1700, but only for areas still present in 2017—that is, if 50 km2 in a grid cell was converted rangeland in 1700, and ≥50 km2 existed in the same grid cell in 2017, we included 50 km2 in our analysis. That way, we captured lands where long-standing and continuous use and grazing have shaped unique ecosystems with high biodiversity. In all other areas—that is, converted rangelands that were established later than 1700 and thus do not have a long historical legacy of grazing—we assumed that the removal of livestock and reforestation, restoration and/or rewilding of these areas would be more beneficial for biodiversity (see, for example, ref. 33). This approach led us to including 0.8 million km2 of HYDE converted rangelands in our analysis (Figs. 1 and 2).
Historical natural rangelands under current high-intensity management (HNR-HI, Fig. 1) cannot immediately be gathered from HYDE v.3.2 because all high-intensity grazing areas are classified as pastures. To single out these areas, we first separated data for pastures for 2017 on the basis of whether they existed within or outside a Olson/WWF forest ecoregion. We did this to differentiate between grazing areas that exist in a grassland/rangeland biome (that is, non-forest biomes) and semi-natural pastures that exist in forest biomes. To capture historical natural rangelands that are currently under high-intensity management, we compared maps of pastures outside forest biomes (current high-intensity management) with historical (year 1700) maps of natural rangelands and only included areas that are present in both datasets (Supplementary Table 4). That way, we allowed grazing in our analysis only on lands where grazing is naturally a part of the landscape dynamics (that is, natural rangelands), and we excluded livestock from all other intensively managed pastures. This approach resulted in the inclusion of 0.3 million km2 (Figs. 1 and 2).
Intensively managed pastures are also found in forest biomes. Applying the same reasoning as for converted rangelands, we based the inclusion/exclusion of semi-natural pastures under current high-intensity use (SNP-HI, Fig. 1) on the historical legacy of grazing. We singled out pastures in forests by overlaying the HYDE data with the Olson/WWF ecoregions. Again, we compared data for the year 1700 and required that the area was a pasture in 1700 and was still present in 2017 to be included in the analysis (Fig. 1). Unlike converted rangeland areas, however, these grazing areas are under high-intensity management as of today (2017), but we assumed that there is scope for restoring biodiversity by reintroducing low-intensity management (see, for example, the review in ref. 22 as well as refs. 34,35,36 for experimental studies). This led us to include 1 million km2 of current (2017) pastures in forest biomes (Figs. 1 and 2).
Step 2: limiting grazing intensity
When combining all four land categories, our analysis included 23.5 million km2 globally. On the total area, we used data on AGB from Piipponen et al.59. Their method uses temperature as a predictor to allocate part of the net primary productivity aboveground, takes the interannual variability of biomass growth (in 2001–2015) into account and reduces grazing on slopes and under tree canopies; this method is therefore argued to be a state-of-the-art approach for estimating grassland carrying capacity59.
While the geographic coverage in Piipponen et al.59 largely overlapped with ours, there were some discrepancies in data availability for different regions compared with HYDE v.3.2, especially in dry regions with low AGB productivity, such as large areas in Saudi Arabia and North Africa (Supplementary Fig. 1). To fill in the data gaps, we calculated AGB for these missing areas by using the methodology of Piipponen et al.59 and running the simulation over 2001–2015 with 500 runs; we used the median of these runs for the final GIS layers (Supplementary Information). We were thus able to obtain AGB data for over 99% of our selected area (Supplementary Fig. 2).
Many studies that analyse the impact on biodiversity from grazing use qualitative estimates (such as low, moderate and heavy) without articulating actual stocking densities24,26,28. Building on reviews and models of the relationship between biodiversity and livestock density, Alkemade et al.22 and Petz et al.23 quantified the relationship between stocking density and biodiversity at the global scale, using mean species abundance (MSA) as a measurement of biodiversity. Their studies suggest that at 10% AGB (in net primary productivity) removal, biodiversity is intact (MSA = 100%)23. Drawing on this, we set 10% biomass removal (MSA is intact) as our baseline grazing intensity.
Step 3: calculating potential production of milk and meat
To quantify the global, regional and national production potentials of ruminant milk and meat, we determined the composition of large (cattle) and small (sheep and goats) ruminants for each country on the basis of mirroring the current ruminant herd structure in grassland-based systems (Supplementary Table 5). These are systems where >90% of livestock feed comes from forage and >10% of the feed is produced on-farm (originally developed by Seré and Steinfeld60 and now used, for example, in FAO’s Gridded Livestock of the World61). By only using data for grassland-based systems, we established herd compositions that are to the largest extent possible determined by the local availability of feed for grazing, and not by external factors such as imported feed. We used data on herd structures of grassland-based production systems from Herrero et al.38 and kept the ratio of dairy cows to bovine beef animals to dairy ewes/does and small ruminants for meat constant for each country (Supplementary Information). One consequence of this is that in our analysis, there is no dairy production in some regions (for example, North America and large parts of Europe), as dairy production systems in these regions use higher shares of non-grass feed sources than what is allowed within the current standard categorization of production systems as grassland-based (for example, in FAO’s Global Livestock Environmental Assessment Model (GLEAM)). This definition of grassland-based is also what we refer to in the comparisons to current production.
On the basis of the selected areas and the corresponding total available biomass for each country, we allocated feed in accordance with the constant ratios between the different animal categories. For example, if country X has a 50:50 split between dairy cows and beef cattle in their current grassland-based production systems, feed was allocated in our calculation so that this remained unchanged, but the total number of animals was adjusted (Supplementary Fig. 3). To do this, we used feed intake coefficients from Herrero et al.38 and could thus calculate how many animals could be kept in each country under our restrictions.
Lastly, using data from Herrero et al.38 on yields of ruminant milk and meat in these grassland-based systems (Supplementary Table 6), we calculated the total potential production of meat and milk. Using country-level human population data from FAOSTAT, we calculated the per capita consumption that would result from this production.
All AGB and feed intake was calculated as dry matter. The production potential was calculated in Excel v.16.82 For protein conversions, see Supplementary Table 7.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The HYDE v.3.2 data and Olson/WWF Ecoregions GIS maps are open access, and links for retrieval are listed in the Supplementary Information. The AGB data and calculations are based on Piipponen et al.59 and are open access (see ‘Code availability’). Open-access background data from FAOSTAT and GLEAM v.3 can be found in Supplementary Data 1, and links for retrieval are in the Supplementary Information. All data for the results are shared in Supplementary Data 1. The data on herd structures, feed intake and yields from Herrero et al.38 are not open access and thus not included in Supplementary Data 1; access must be requested from owners of the dataset.
Code availability
The code for the analysis of data gaps between selected areas and AGB data builds on Piipponen et al.59. The input data and source code can be accessed via Zenodo at https://doi.org/10.5281/zenodo.6366896 (ref. 62) and via GitHub at https://github.com/jpiippon/cc_rsd_repo.
References
Chiles, R. M. & Fitzgerald, A. J. Why is meat so important in Western history and culture? A genealogical critique of biophysical and political–economic explanations. Agric. Hum. Values 35, 1–17 (2018).
Herrero, M., Thornton, P. K., Gerber, P. & Reid, R. S. Livestock, livelihoods and the environment: understanding the trade-offs. Curr. Opin. Environ. Sustain. 1, 111–120 (2009).
Pastoralist Knowledge Hub (FAO, 2020); https://www.fao.org/pastoralist-knowledge-hub/news/detail/en/c/1330121/
Díaz, S. et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 366, eaax3100 (2019).
Clark, M. et al. Global food system emissions could preclude achieving the 1.5 ° and 2 °C climate change targets. Science 370, 705–708 (2020).
FAOSTAT—Data (FAO, 2022); http://www.fao.org/faostat/en/#data
The Future of Food and Agriculture—Alternative Pathways to 2050 (FAO, 2018).
IPBES Global Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (eds Brondízio, E. S. et al.) (IPBES, 2019); https://ipbes.net/global-assessment-report-biodiversity-ecosystem-services
IPCC: Summary for Policymakers. In Climate Change 2021: The Physical Science Basis (eds Masson-Delmotte, V. et al.) (Cambridge Univ. Press, 2021); https://www.ipcc.ch/report/ar6/wg1/
van Zanten, H. et al. Defining a land boundary for sustainable livestock consumption. Glob. Change Biol. 24, 4185–4194 (2018).
Bengtsson, J. et al. Grasslands—more important for ecosystem services than you might think. Ecosphere 10, e02582 (2019).
Briske, D. et al. Strategies to alleviate poverty and grassland degradation in Inner Mongolia: intensification vs production efficiency of livestock systems. J. Environ. Manage. 152, 177–182 (2015).
Arzamendia, Y. et al. The Puna pastoralist system: a coproduced landscape in the central Andes. Mt. Res. Dev. 41, R38–R49 (2021).
Wilson, J. B., Peet, R. K., Dengler, J. & Pärtel, M. Plant species richness: the world records. J. Veg. Sci. 23, 796–802 (2012).
Plieninger, T. et al. Wood-pastures of Europe: geographic coverage, social–ecological values, conservation management, and policy implications. Biol. Conserv. 190, 70–79 (2015).
Fløjgaard, C., Pedersen, P. B. M., Sandom, C. J., Svenning, J. C. & Ejrnæs, R. Exploring a natural baseline for large-herbivore biomass in ecological restoration. J. Appl. Ecol. 59, 18–24 (2022).
Zimov, S. A., Zimov, N. S., Tikhonov, A. N. & Chapin, F. S. Mammoth steppe: a high-productivity phenomenon. Quat. Sci. Rev. 57, 26–45 (2012).
Houghton, R. A. et al. Changes in the carbon content of terrestrial biota and soils between 1860 and 1980: a net release of CO2 to the atmosphere. Ecol. Monogr. 53, 236–262 (1983).
Ramankutty, N., Evan, A. T., Monfreda, C. & Foley, J. A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 22, GB1003 (2008).
Pongratz, J., Reick, C., Raddatz, T. & Claussen, M. A reconstruction of global agricultural areas and land cover for the last millennium. Glob. Biogeochem. Cycles 22, GB3018 (2008).
Dengler, J., Biurrun, I., Boch, S., Dembicz, I. & Torok, P. Grasslands of the Palaearctic Biogeographic Realm: Introduction and Synthesis. Encyclopedia of the World’s Biomes Vols 3–5 (Elsevier, 2020).
Alkemade, R., Reid, R. S., van den Berg, M., de Leeuw, J. & Jeuken, M. Assessing the impacts of livestock production on biodiversity in rangeland ecosystems. Proc. Natl Acad. Sci. USA 110, 20900–20905 (2013).
Petz, K. et al. Mapping and modelling trade-offs and synergies between grazing intensity and ecosystem services in rangelands using global-scale datasets and models. Glob. Environ. Change 29, 223–234 (2014).
Zhou, G. et al. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis. Glob. Change Biol. 23, 1167–1179 (2017).
Herrero-Juregui, C. & Oesterheld, M. Effects of grazing intensity on plant richness and diversity: a meta-analysis. Oikos 127, 757–766 (2018).
Wang, C. & Tang, Y. A global meta-analyses of the response of multi-taxa diversity to grazing intensity in grasslands. Environ. Res. Lett. 14, 114003 (2019).
Gao, J. & Carmel, Y. A global meta-analysis of grazing effects on plant richness. Agric. Ecosyst. Environ. 302, 107072 (2020).
Filazzola, A. et al. The effects of livestock grazing on biodiversity are multi-trophic: a meta-analysis. Ecol. Lett. 23, 1298–1309 (2020).
Klein Goldewijk, K., Beusen, A., Doelman, J. & Stehfest, E. Anthropogenic land use estimates for the Holocene—HYDE 3.2. Earth Syst. Sci. Data 9, 927–953 (2017).
IPCC Special Report on Climate Change and Land (eds Shukla, P. R. et al.) (in the press); https://www.ipcc.ch/srccl/
Willett, W. et al. Food in the Anthropocene: the EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 393, 447–492 (2019).
Pedersen, R. Ø., Faurby, S. & Svenning, J. Late‐Quaternary megafauna extinctions have strongly reduced mammalian vegetation consumption. Glob. Ecol. Biogeogr. 32, 1814–1826 (2023).
Torres, A. et al. Measuring rewilding progress. Phil. Trans. R. Soc. B 373, 20170433 (2018).
Calladine, J., Baines, D. & Warren, P. Effects of reduced grazing on population density and breeding success of black grouse in northern England. J. Appl. Ecol. 39, 772–780 (2002).
Louault, F., Pillar, V. D., Aufrère, J., Garnier, E. & Soussana, J. F. Plant traits and functional types in response to reduced disturbance in a semi-natural grassland. J. Veg. Sci. 16, 151–160 (2005).
Walker, K. J. et al. Recreation of lowland heathland on ex-arable land: assessing the limiting processes on two sites with contrasting soil fertility and pH. J. Appl. Ecol. 44, 573–582 (2007).
Milchunas, D., Sala, O. & Lauenroth, W. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 132, 87–106 (1988).
Herrero, M. et al. Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc. Natl Acad. Sci. USA 110, 20888–20893 (2013).
GLEAM v3.0 Dashboard (FAO, 2023); https://foodandagricultureorganization.shinyapps.io/GLEAMV3_Public/
Schader, C. et al. Impacts of feeding less food-competing feedstuffs to livestock on global food system sustainability. J. R. Soc. Interface 12, 20150891 (2015).
Mäder, P. et al. Soil fertility and biodiversity in organic farming. Science 296, 1694–1697 (2002).
Tuck, S. L. et al. Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J. Appl. Ecol. 51, 746–755 (2014).
Karlsson, J. O. Livestock as Resource Users and Landscape Managers—a Food Systems Perspective (Swedish University of Agricultural Sciences, 2022).
Gerber, P. J. et al. Tackling Climate Change through Livestock—a Global Assessment of Emissions and Mitigation Opportunities. (Food and Agriculture Organisation of the United Nations, 2013).
Sayre, N. F., Davis, D. K., Bestelmeyer, B. & Williamson, J. C. Rangelands: where anthromes meet their limits. Land 6, 31 (2017).
Fetzel, T. et al. Quantification of uncertainties in global grazing systems assessment. Glob. Biogeochem. Cycles 31, 1089–1102 (2017).
Klein Goldewijk, K., Dekker, S. C. & van Zanden, J. L. Per-capita estimations of long-term historical land use and the consequences for global change research. J. Land Use Sci. 12, 313–337 (2017).
Klein Goldewijk, K., Beusen, A., Van Drecht, G. & De Vos, M. The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12,000 years. Glob. Ecol. Biogeogr. 20, 73–86 (2011).
Li, S., He, F., Zhang, X. & Zhou, T. Evaluation of global historical land use scenarios based on regional datasets on the Qinghai–Tibet area. Sci. Total Environ. 657, 1615–1628 (2019).
Li, F. et al. Towards quantification of Holocene anthropogenic land-cover change in temperate China: a review in the light of pollen-based REVEALS reconstructions of regional plant cover. Earth Sci. Rev. 203, 103119 (2020).
Newbold, T. et al. Has land use pushed terrestrial biodiversity beyond the planetary boundary? A global assessment. Science 353, 291–288 (2016).
Gao, J. & Carmel, Y. Can the intermediate disturbance hypothesis explain grazing–diversity relations at a global scale? Oikos 129, 493–502 (2020).
Figueiredo, F., Jahi, G. M. & Frédéric, C. Sparing or expanding? The effects of agricultural yields on farm expansion and deforestation in the tropics. Biodivers. Conserv. https://doi.org/10.1007/s10531-022-02540-4 (2023).
Trouwborst, A. & Svenning, J. C. Megafauna restoration as a legal obligation: international biodiversity law and the rehabilitation of large mammals in Europe. Rev. Eur. Comp. Int. Environ. Law 31, 182–198 (2022).
Sun, Z. et al. Dietary change in high-income nations alone can lead to substantial double climate dividend. Nat. Food 3, 29–37 (2022).
Li, C., Wu, X., Chen, K. & Chen, G. Global pastureland use as reflected in inter-regional supply chain. J. Environ. Manage. 322, 116016 (2022).
Thompson Hobbs, N. et al. Fragmentation of rangelands: implications for humans, animals, and landscapes. Glob. Environ. Change 18, 776–785 (2008).
Klein Goldewijk, K. Estimating global land use change over the past 300 years: the HYDE database. Glob. Biogeochem. Cycles 15, 417–433 (2001).
Piipponen, J. et al. Global trends in grassland carrying capacity and relative stocking density of livestock. Glob. Change Biol. https://doi.org/10.1111/GCB.16174 (2022).
Seré, C. & Steinfeld, H. World Livestock Production Systems: Current Status, Issues and Trends. (Food and Agriculture Organisation of the United Nations, 1996).
Gilbert, M. et al. Global distribution data for cattle, buffaloes, horses, sheep, goats, pigs, chickens and ducks in 2010. Sci. Data 5, 180227 (2018).
Piipponen, J. et al. Data for ‘Global trends in grassland carrying capacity and relative stocking density of livestock’. Zenodo https://doi.org/10.5281/zenodo.6366896 (2022).
Acknowledgements
We thank H. van Zanten, F. van der Plas and F. Cozim-Melges for contributing to early discussions on the conceptualization and methodology of this study. We also thank M. Herrero for sharing unpublished data and volunteering time to answer questions about the use of the data. We acknowledge the following funding: the Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning, grant no. 2019-00403 (K.R.S.); the European Research Council Horizon 2020 research and innovation programme, grant no. 819202 (J.P.); and GRAZELIFE, grant no. LIFE18PRE/NL002 (J.R.-L.).
Funding
Open access funding provided by Stockholm University.
Author information
Authors and Affiliations
Contributions
K.R.S. contributed to the conceptualization, methodology development, data collection, and analysis, and led the article writing and revisions, author group coordination and acquisition of funding. L.J.G. contributed to the conceptualization, methodology development, article writing and revisions, and acquisition of funding. R.L. contributed to the conceptualization, methodology development and article revisions. J.P. contributed to the method for grassland carrying capacity, writing on methods and article revisions. P.V.R. contributed to the methodology development, data analysis and article revisions. J.R.-L. contributed to the conceptualization, methodology development and article revisions. E.R. contributed to the conceptualization, methodology development, data collection, analysis, article writing and revisions, and acquisition of funding.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Sustainability thanks Imke J. M. de Boer, An Notenbaert and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Figs. 1–3, Tables 1–7, Methods description and Lists of data.
Supplementary Data 1
Data for the results, sensitivity checks and open access background data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Resare Sahlin, K., Gordon, L.J., Lindborg, R. et al. An exploration of biodiversity limits to grazing ruminant milk and meat production. Nat Sustain (2024). https://doi.org/10.1038/s41893-024-01398-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41893-024-01398-4
- Springer Nature Limited